Keywords
hyperoside, natural product, effect, therapy, pharmacology, review.
hyperoside, natural product, effect, therapy, pharmacology, review.
Plants have been used as folkloric sources of medicinal agents since the beginning of humankind. Following the emergence of modern medicine and single pure drugs, plant-derived active principles and their semi-synthetic and synthetic analogs have served as a major route to new pharmaceuticals (Lee, 1999). Hyperoside, a flavonoid compound (Magnus et al., 2020), is isolated from Hypericaceae (Figure 1), Rhododendron ponticum L. Rosaceae, Campanulaceae, Lamiaceae, and Berberis (Guo et al., 2020). Hyperoside, which is the main active ingredient of many Hypericum and Crataegus plants, has anti-inflammatory, anti-oxidant, anti-tumor, anti-bacterial and anti-viral, anti-coagulant, anti-platelet and lipid-lowering, hypoglycemic, and other pharmacological activities. It can protect the liver, kidney, and nervous, cardio-cerebrovascular, and cardio-cerebrovascular systems. Over recent years, the antidepressant, anti-neurodegenerative, and bone protective effects of hyperoside have also attracted people’s attention. Clinically, several drugs containing hyperoside or made from hyperoside have been widely used (Chen et al., 2020; He et al., 2016; Qian et al., 2007; Wang et al., 2011; Wei et al., 2009; Wu et al., 2019; Zou et al., 2004).

With the continuous development of scientific research, numerous comprehensive studies have been conducted for hyperoside, including those addressing many pharmacological effects. Hyperoside can effectively prevent age-related kidney damage, protect human primary melanocytes from oxidative damage induced by H2O2, and liver fibrosis induced by heart failure in rats (Guo et al., 2019b; Liu et al., 2018a; Yang et al., 2016). Hyperoside can also effectively induce the apoptosis of breast cancer cells, autophagy, and apoptosis of non-small cell lung cancer cells, and inhibit the survival and proliferation of lung cancer cells induced by hypoxia (Chen et al., 2020; Fu et al., 2016; Qiu et al., 2019). Considering anti-inflammatory effects, hyperoside glycosides can effectively reduce allergic airway inflammation, inhibit lipopolysaccharide-induced inflammation of microglia, and depress pro-inflammatory cytokines in human lung epithelial cells infected with Mycoplasma pneumonia (Fan et al., 2017; Liu et al., 2019a; Ye et al., 2017), all of which show the great potential of hyperoside for the development of a new drug. Therefore, this article reviews the existing pharmacological effects of hyperoside to provide references for further research on hyperoside.
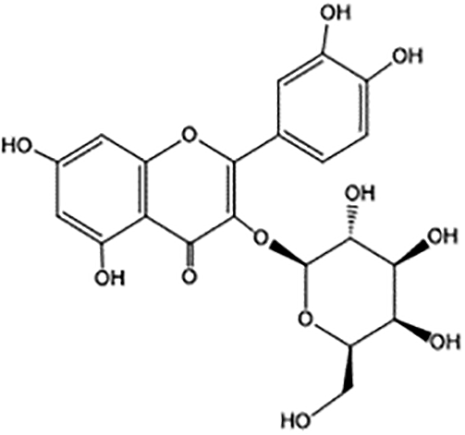
The chemical structure of hyperoside (Figure 2) is composed of two phenyl rings (A ring and B ring), a hexaoxy heterocyclic ring (C ring), and a galactoside (D ring). It is a pale-yellow needle crystal with a melting point of 227–229°C. The rotation rate is −83°C (C=0.2, pyridine), which is easily soluble in ethanol, methanol, acetone, and pyridine. It reacts with magnesium hydrochloric acid powder by producing fuchsia color, ferric chloride by producing green color, and naphthol by positive reaction (Lavie et al., 1995; Liu et al., 2019b). Compared with synthetic drugs, natural plant drugs have fewer side effects and higher economic value. Hyperoside is found in a variety of plants. For example, the content range of hyperoside is 2.790–6.502 mg/g in Cuscuta chinensis (Lin et al., 2007), 3.64 μg/100 mg in the dried Geranii robertiani herba (Fodorea et al., 2005), 0.85–2.7 mg/g in the hawthorn buds (Jakstas et al., 2004), and 4.6–4.9 mg/g in St. John’s Wort (Bertoli et al., 2008). However, the content is relatively low in plants, where it is mainly synthesized from rutin.
As early as 1996, Wang et al. (1996) found that hyperoside has a protective effect on myocardial ischemia/reperfusion injury. After years of follow-up studies by other scholars, the effect has been reaffirmed. Hyperoside has a protective effect on cardiomyocyte injury induced by hypoxia/reoxygenation through anti-oxidant properties. After hyperoside preconditioning, the activities of Bnip3, Bax, and caspase3 decrease, and the expression of Bcl-2 increases (Xiao et al., 2017). Hyperoside can alleviate heart failure by inducing autophagy, inhibiting apoptosis, changing the level of apoptosis-related proteins, and promoting the autophagy of H9C2 cells induced by angiotensin II. Most importantly, hyperoside can reduce the heart/body weight ratio and the cross-sectional area of cardiomyocytes (Guo et al., 2020). Recent studies proved that (Wang et al., 2018b) hyperoside can also prevent cardiac hypertrophy by blocking the activation of the AKT signal pathway and protecting heart remodeling caused by pressure overload. In the model of oxidative injury induced by high glucose, hyperoside protects cardiomyocytes from oxidative stress induced by high glucose by activating PI3K/AKT/Nrf2 signal pathway (Wang et al., 2018a). After the hyperoside intervention, cardiac function parameters of cardiomyocytes injured by ischemia-reperfusion injury obtained significant improvements. The possible mechanism is that hyperoside-activated anti-oxidant Nrf2 signal pathway and decreased the level of endoplasmic reticulum stress and oxidative stress (Hou et al., 2016). However, some studies suggested that the protective effect of hyperoside on ischemia–reperfusion injury of isolated rat cardiomyocytes may be related to the activation of the ERK signal pathway, thus promoting the phosphorylation of extracellular signal-regulated protein kinase, improving myocardial contractile function, and reducing the myocardial infarct size (Li et al., 2013).
The effect of hyperoside on cerebral ischemia–reperfusion injury has also been previously reported. Hyperoside was administered to rats at doses of 6.25, 12.5, and 25 mg·kg−1, which can significantly improve abnormal neurological symptoms. Among them, 12.5 and 25 mg·kg−1 significantly reduced infarct weight, inhibited the increase of MDA and NO in rats’ cerebral cortex, increased the CBF of the cerebral cortex, and protected cerebral infarction by reducing lipid peroxidation and NO (Chen et al., 1998). Liu et al. (2012) studied primary cultured rat neurons and created ischemia–reperfusion cell model (OGDR), reporting that hyperoside could significantly reduce the damage caused by OGDR to neurons and that internal mechanisms were related to the NO signal pathway. The regulation of hyperoside may be related to the BKC a channel by activating TRPV4, reducing the concentration of Ca2+ in cells, relaxing blood vessels, forming a new therapeutic target for protecting ischemic brain injury, and participating in brain protection (Han et al., 2018).
The inhibitory effect of hyperoside on tumor has been confirmed in various human malignant tumor cells such as lung, breast, liver, prostate, colon, gastric, and similar. The important reason for the unrestricted growth of tumor cells is the loss of their active and physiological apoptotic ability. Therefore, inducing tumor cell apoptosis and inhibiting tumor cell proliferation are the main purposes of most anti-tumor drugs (Shi et al., 2014). Hyperoside can affect tumor cells through different mechanisms of action, including depressing proliferation, promoting apoptosis, blocking cell cycle.
Hyperoside could inhibit the survival and proliferation of hypoxia-induced non-small cell lung cancer cell line A549 in a dose-dependent manner, possibly through iron accumulation on the AMPK/HO-1 axis to combat hypoxia-induced survival and proliferation of A549 (Chen et al., 2020). In vitro experiments revealed that hyperoside could induce autophagy and apoptosis in human non-small cell lung cancer by inhibiting the activation of the Akt/mTOR/p70S6K signaling pathway (Fu et al., 2016). By inducing the inactivation of the NF-kB signaling pathway, activating caspase3 regulated by Bcl-2/Bax, and increasing lung cancer malignant tumor cell apoptosis, hyperoside can also inhibit lung cancer growth. Regulating caspase-3 and p53 inhibits the proliferation, migration, and invasion of lung cancer cells, promotes the expression of pro-apoptotic factors, and suppresses the expression of anti-apoptotic factors (Liu et al., 2016; Lü, 2016). Apoptosis is closely related to the activation of p38MAPK and JNK-induced mitochondrial death pathway (Yang et al., 2017c). Further studies proved that hyperoside mediates p38MAPK and AKT/PI3K signaling pathways, regulates the expression of genes related to migration and invasion and inhibits the invasion and migration of A549 cells (Yang et al., 2017b). Some studies have shown that hyperoside can also work synergistically with some drugs to exert its anti-tumor effect. Hyperoside can inhibit the proliferation of A549 cells by inducing cell apoptosis and G1/S phase arrest. Let-7a-5p can inhibit the proliferation of A549 cells by blocking the cell cycle in G1/S phase. At the same time, microRNA-let-7a-5p directly regulates the expression of CCND1 in A549 cells. Hyperoside combined with let-7a-5p can effectively inhibit the proliferation of A549 cells (Li et al., 2018a).
According to breast cancer research, hyperoside can reduce the production of ROS, inhibit the transcription activity of NF-kB and the expression of Bcl-2 and XIAP, promote the expression of Bax and cleaved caspase-3, and induce breast cancer cells MCF-7 and the apoptosis of 4T1 that can scavenge free radicals and has a cytotoxic effect on breast cancer MCF-7 cells (Qiu et al., 2019). Hyperoside can inhibit the viability of breast cancer cells without being cytotoxic to normal breast mammary epithelial cell lines (Agar et al., 2015). It also increases cell apoptosis and caspase-3 activity, inhibits the activation of TLR4-NF-κB signal transduction caused by paclitaxel, reduces the expression of anti-apoptotic protein Bcl-2, enhances the expression of pro-apoptotic protein Bax and pro-inflammatory cytokines IL-6, and IL-6 protein levels increase the sensitivity of paclitaxel to breast cancer cells MDA-MB-231 (Sun et al., 2020). In other types of cancer, hyperoside can also exert its anti-cancer effect. Guo et al found that hyperoside can reduce the expression of the C-Myc gene in cervical cancer C-33A and HeLa cells and promote the expression of TFRC. To inhibit the proliferation of cervical cancer cells (Guo et al., 2019a), hyperoside may induce the apoptosis of human endometrial RL952 cells through the Ca2+-related mitochondrial apoptosis pathway, and it can induce death receptor-mediated and mitochondrial-mediated apoptosis. The apoptotic pathway induces apoptosis of HT-29 human colon cancer cells, thereby inhibiting tumor growth (Guon and Chung, 2016; Li et al., 2012a). Hyperoside can inhibit GSH-Px and CAT mRNA expression by inducing caspase-dependent apoptosis and p53 signaling pathway and participating in the pro-apoptotic signal transduction of SW620 human colorectal cancer cells (Zhang et al., 2017a). It can also induce apoptosis of SW579 human thyroid squamous cell carcinoma cells, partly by up-regulating Fas during apoptosis And FasL mRNA expression and down-regulating survivin protein expression to induce apoptosis (Liu et al., 2017). Hyperoside inhibits the cycle of INS-1 and MIA PaCa-2 pancreatic cancer cells in the G2/M phase and activates caspase-3 protein expression, induces apoptosis of tumor cells, and inhibits the proliferation of osteosarcoma cells by inducing G0/G1 block in the cell cycle (Boukes and van de Venter, 2016; Zhang et al., 2014). It may also reactivate caspase-9 and caspase-9 by inhibiting BAD phosphorylation, increase the level of p27 by up-regulating LC-II in the HL-60 AML cell line; it can induce autophagy, and enhance the apoptosis-inducing effect of As2O3 on acute myeloid leukemia cells (Zhang et al., 2015). Hyperoside can also down-regulate β1-adrenergic receptors in rat C6 glioblastoma cells, reduce the β1AR density in the plasma membrane, and subsequently reduce downstream signal transduction, thus inhibiting the growth of tumor cells (Jakobs et al., 2013). High glycosides use PGRMC1-dependent autophagy to induce apoptosis and cell death of ovarian cancer cells and increase tumor cells’ sensitivity to cisplatin drugs. In combination with quercetin, it can inhibit prostate cancer cells and kidney cancer by regulating microRNA-21 Cell growth and metastasis (Li et al., 2014; Yang et al., 2015; Zhu et al., 2017).
Growing evidence has suggested that hyperoside has anti-inflammatory, anti-oxidant stress, anti-swelling, anti-bacterial and anti-viral effects in vivo and in vitro. Due to the anti-inflammatory effect, hyperoside is commonly applied in the treatment of a variety of inflammatory-related diseases.
Hyperoside can inhibit the release of lipopolysaccharide (LPS)-mediated HMGB1 and HMGB1-mediated cytoskeletal rearrangement and inhibit the HMGB1 signaling pathway to treat vasculitis (Ku et al., 2015). Zhou et al. (2018) explored the effects of hyperoside on inflammation and apoptosis of human umbilical vein endothelial cells induced by endotoxin. They found that 20 μg/L and 50 μg/L hyperoside could significantly increase the survival rate of human umbilical vein endothelial cells induced by LPS. In addition, hyperoside could decrease the mRNA expression of IL-1β, IL-6, TNF-α, and iNOS in umbilical vein endothelial cells in a dose-and time-dependent manner. Furthermore, some studies showed that hyperoside could inhibit vascular inflammation mediated by TNF-α, which is characterized by the drop of VCAM-1 expression in vascular smooth muscle cells and the adhesion ability of monocytes to vascular smooth muscle cells, where hyperoside (10, 50, 100 μmol/L) dose-dependently inhibited the proliferation and migration of human RAFLSs induced by LPS, reduced the production of TNF-α, IL-6, IL-1 and MMP-9 in LPS-stimulated cells and inhibited lipopolysaccharide. Moreover, it induced p65 and IκBα phosphorylation, lipopolysaccharide-induced p65 nuclear translocation, and NF-κB DNA adhesion at 3 weeks after administration, thus significantly reducing the clinical score of collagen-induced arthritis (CIA) in mice, reduced synovial hyperplasia, inflammatory cell infiltration, and cartilage damage (Fan et al., 2017; Jang et al., 2018; Jin et al., 2016; Yang et al., 2017a). In another study, hyperoside significantly inhibited the loss of cell viability and the increase in endothelial Ca2+ content and apoptosis in HUVEC induced by H2O2, and reduced B-cell lymphoma (Bcl)-2 related X Protein (Bax). It also cleaved caspase-3 and phosphorylated p38 mRNA expression levels while increasing the mRNA expression of Bcl-2 in H2O2-induced HUVEC, thus indicating that it has a certain anti-H2O2-induced HUVEC apoptosis effect, as well as a key role in preventing cardiovascular diseases (Hao et al., 2016). Hyperoside could decrease skin inflammation by inhibiting inflammatory pathways and repairing DNA damage (Kurt-Celep et al., 2020). Animal experiments demonstrated that hyperoside could lower TNF-α and IL-1β in rat cerebral ischemia-reperfusion injury models and play an anti-inflammatory role (He et al., 2019). Hyperoside also has a protective effect on ovalbumin-induced allergic airway inflammation in mice by decreasing the levels of IL-4, IL-5, IL-1β, and IgE and reducing inflammatory cell infiltration (Ye et al., 2017).
Hyperoside can selectively block the activation of AIM2 and NLRC4 inflammatory bodies and inhibit inflammatory response. In vitro experiments illustrated that hyperoside inhibited the production of TNF-α and IL-1β and inhibited the activation of AKT, NF-κB, and extracellular regulated kinase (ERK1/2) that are mediated by HMGB1 in lipopolysaccharide-induced vascular endothelial cells (Jung et al., 2012). In rat peritoneal macrophages, hyperoside inhibited the expression of pro-inflammatory cytokines and iNOS, and significantly decreased the levels of inflammatory cytokines such as TNF-α and IL-6 (Kim et al., 2011). Hyperoside can also be applied to treat allergic inflammation aggravated by TSLP, thus reducing the expression of IL-1β, IL-6, and their mRNA, down-regulating Ca2+/RIP2/Caspase-1/NF-κB signal pathway and inhibiting the level of TSLP in human mast cell lines (Han et al., 2014).
The role of oxidative stress in cardiovascular diseases, malignant tumors, liver and kidney injury, and autoimmune diseases has been extensively verified. Hyperoside has a protective effect on oxidative stress and apoptosis of granulosa cells induced by H2O2, which is potentially exerted by reducing the expression of Bax and up-regulating the expression of Bcl-2 in granulosa cells (Wang et al., 2019). After intraperitoneal injection of hyperoside (50 mg/kg/d) into the rat model of ischemia–reperfusion injury, the activity of malondialdehyde decreased, the activity of superoxide dismutase and glutathione peroxidase increased, the expression of heme oxygenase-1 and NADPH quinone oxidoreductase-1 increased, and the apoptosis index decreased. Hyperoside could remarkably reduce the levels of ALT and AST after reperfusion and reduce the histological injury score (Shi et al., 2019). Similarly, polyphenols in the lotus chamber, including hyperoside and other compounds, were also reported to display strong anti-oxidant and anti-proliferation activity and to have the ability to effectively scavenge many kinds of free radicals such as superoxide anion (Shen et al., 2019).
In a rat model of cerebral ischemia–reperfusion injury, He et al. (Kurt-Celep et al., 2020) found that hyperoside could increase the levels of SOD, MDA, and GSH-Px, improve the total anti-oxidant capacity of the rat brain, and inhibit oxidative stress and anti-apoptosis. Gao et al. (2019) applied Saccharomyces cerevisiae as a model to study the anti-oxidant activity of hyperoside, revealing that hyperoside can reduce the level of intracellular reactive oxygen species and lipid peroxidation, and improve cell survival rate. Chen et al. (2019) proposed that hyperoside improves the activity of free radical scavenging (or power reduction) in a dose-dependent manner and has an anti-oxidant role through the REDOX reaction and covalent pathway of lyophilized water extract and phenolic components of Hyalocin. Hyperoside promotes the expansion of cord blood hematopoietic cells in vitro by reducing the level of intracellular ROS. The expansion ability of cord blood hematopoietic cells that are pretreated with hyperoside (1 μM) was 54.9±9.6 times higher than that of the control group (42.0±8.1 times), which was a noticeable difference (Zhang et al., 2018a). Hyperoside exerts a protective effect on apoptosis of retinal pigment epithelial cells by inhibiting blue light-induced poly ADP-ribose polymerase cleavage and complement C3 activation of PARP (Kim et al., 2018). Similarly, hyperoside has a protective effect on oxidative damage and cytotoxicity of renal cells simulated by oxalate, and the ability of hyperoside to enhance endogenous antioxidation and detoxification may be closely related to Nrf2/HO-1/NQO1 pathway (Chen et al., 2018b). In vitro experiments revealed that hyperoside could enhance the activity of anti-oxidant enzyme SOD/CAT/GSH-Px (Zou et al., 2017).
Yang et al. (2016) demonstrated that hyperoside protects melanocytes from oxidative damage by inhibiting p38 phosphorylation and mitochondrial apoptosis signals and activating AKT, which provides vital value for the treatment of vitiligo. By inducing an endogenous oxidation system, hyperoside up-regulates the level of Nrf2 and the binding activity of anti-oxidant response elements and increases the expression of HO-1 mRNA and protein in a time- and dose-dependent manner (Park et al., 2016). Additionally, hyperoside in hawthorn extract has been shown to have an immunomodulatory effect through its antioxidant activity, including spleen cells, NK cells, CTL cells, and macrophages; however, the specific mechanism remained unclear (Mustapha et al., 2016). Li et al. (2012b) argued that the underlying mechanism might be correlated with the activation of the ERK signal pathway. According to a recent study, hyperoside can attenuate H2O2-induced oxidative stress damage in L02 cells by inhibiting the KAP1-activated NRF2-ARE signal pathway and increasing serine kinase-3β to inhibit phosphorylation (Xing et al., 2015). The anti-oxidant activity of hyperoside is also reflected in its inhibitory effect on cell injury induced by H2O2 that up-regulates the expression of HO-1 and increases the activity of heme oxygenase-1 through the interaction of Keap1-Nrf2-ARE signal pathway (Xing et al., 2011). Hyperoside depends on the regulation of NMDA receptors of NR2A and NR2B, and significantly attenuates NMDA-induced neuronal apoptosis and prevents neuronal injury (Zhang et al., 2010). Previous studies revealed that H2O2 could induce apoptosis of hamster lung fibroblasts (V79-4). Through the intervention of hyperoside, the activity of antioxidant enzymes in V79-4 cells significantly increases, while the content of reactive oxygen species obviously decreases (Piao et al., 2008).
Numerous studies indicated that hyperoside has a neuroprotective effect. Hyperoside has a protective effect on human dopaminergic neurons, and it can inhibit 6-hydroxydopamine induced oxidative stress of dopaminergic neurons by activating Nrf2/HO-1 signal. It can also improve the loss of neuronal vitality, lactate dehydrogenase release, excessive accumulation of ROS, and abnormal mitochondrial membrane potential induced by 6-OHDA (Ramesh et al., 2018). Most importantly, hyperoside treatment activates nuclear erythroid 2-related factor 2 that is the upstream molecule of heme oxygenase-1. Simultaneously, Nrf2-dependent HO-1 signal activation is a potential target for preventing and treating Parkinson’s disease (Kwon et al., 2019).
Amyloid protein, which is thought to have an important role in Alzheimer’s disease’s pathogenesis, produces neurotoxicity by destroying the blood-brain barrier (Kumaran et al., 2018). Liu et al. (2018b) stated that hyperoside could alleviate the damage of a blood-brain barrier induced by Aβ1–42 and may be a potential drug for AD treatment. Furthermore, hyperoside can reverse mitochondrial dysfunction induced by Aβ25-35, inhibit its toxicity and apoptosis, and protect primary cultured cortical neurons by regulating PI3K/AKT/Bad/BclXL pathway (Zeng et al., 2011). Based on the in vitro ischemia model of hypoxia–glucose deprivation–reperfusion injury, it was obvious that hyperoside has a protective effect on primary cultured cortical neurons against OGD-R injury and that NO signal pathway is involved in this regulatory process (Liu et al., 2012).
As the most important detoxification organ of the human body, the metabolism of most drugs depends on the liver, and liver injury caused by drugs is inevitable. Paracetamol is widely used in clinics due to its good antipyretic and analgesic effect; however, liver injury caused by acetaminophen is unavoidable. In vitro experiments proved that hyperoside could effectively reduce the indexes of acute liver injury induced by acetaminophen. The possible mechanisms of ALT, AST, and ALP are related to hyperoside that can increase the level of glutathione and reduce the content of ROS (Jiang et al., 2019). Xie et al. (2016) proposed that Hyperoside accelerates the harmless metabolism of APAP by inhibiting the activity of CYP2E1 and increasing the expression and activity of detoxifying enzymes Sults and UGTS.
Hyperoside has a protective effect on liver fibrosis and liver injury induced by heart failure in rats, such as the decrease of hydroxyproline content and liver fibrosis area, the decrease of ALT, AST, and ALP levels, and the relief of hepatocyte edema and vacuolar degeneration (Guo et al., 2019b). By inhibiting inflammation and strengthening the anti-oxidant defense system, the protein and mRNA expression of iNOS, COX-2, and TNF-α are inhibited (Choi et al., 2011). Recently, Wang et al. (2016) suggested that hyperoside can effectively inhibit the DNA binding activity of transcription factor NF-κB, change the expression of apoptosis-related genes regulated by it, and induce HSC apoptosis in hepatic stellate cells to reduce hepatic fibrosis.
Diabetic nephropathy is one of the diabetic microvascular complications. Early selective loss of glomerular permeability has an essential role in the pathogenesis of microalbuminuria in diabetic nephropathy. Oral administration of hyperoside to 30 mg/kg/day DN mice for 4 weeks can significantly reduce microalbumin excretion and glomerular filtration but has no significant effect on glucose metabolism and lipid metabolism (Zhang et al., 2016a). Moreover, hyperoside may delay the progression of diabetic nephropathy by down regulating the phosphorylation levels of p38MAPK, IL-β, and pERK1/2 in glomerular Mesangial cells of diabetic rats and reducing the expression of TGF-κ1 and AGE/RAGE binding (Kim et al., 2016). Hyperoside can inhibit the activity of heparanase gene promoter and heparanase expression induced by ROS or high glucose in podocytes (An et al., 2017).
Wang et al. (Wu et al., 2018) proposed that Huangkui capsule and its active ingredient hyperoside could improve the proteinuria and renal dysfunction of the mouse model of early diabetic nephropathy, reduce glomerular basement membrane thickening, glomerular hypertrophy and mesangial dilatation, and alleviate the pathological changes of early DN by inhibiting Akt/mTOR/p70S6K signal activity, which was consistent with results reported in another study (Cai et al., 2017). Huangkui capsule significantly improves the renal function of rats with chronic renal failure induced by adenine and significantly inhibits Scr, BUN, UP, p-ERK1/2, α-SMA, and similar. It was recently reported that hyperoside could alleviate D-galactose-induced renal injury and delay renal aging by inhibiting AMPK-ULK1 signal-mediated autophagy (Liu et al., 2018a). Hyperoside can block ADR-induced mitochondrial division and improve renal injury in both ADR-induced nephrotic mice and cultured human podocytes (Chen et al., 2017).
Further studies revealed that hyperoside could alleviate glomerulosclerosis and improve renal function in diabetic nephropathy mice, which may be related to its promotion of the expression of MMP-2 and MMP-9 and the inhibition of the expression of TIMP-1 and Col IV, FN (Zhang et al., 2016b). Yan et al. (2014) proposed the renal fibrosis model of Wistar rats based on unilateral ureteral obstruction, reporting that the combination of hyperoside and quercetin (H:Q=1:1) could significantly inhibit the expression of smooth muscle actin and fibronectin in Mesangial cells induced by interleukin-1β (IL-1β), thus effectively interfering with the progression of renal fibrosis. As the main active component of total flavonoids of Abelmoschus, hyperoside could inhibit caspase-3 and caspase-8 expression induced by advanced glycation end product, reduce podocyte apoptosis induced by sAGEs and prevent renal injury (Lei et al., 2012). Furthermore, the combination of hyperoside and quercetin (20 mg/kg/day) can inhibit the formation of calcium oxalate stones induced by ethylene glycol in rats and significantly increase catalase levels superoxide dismutase (Zhu et al., 2014).
Hyperoside, which decreases the levels of MC3T3-E1 phosphorylated Jun N-terminal kinases and p38 induced by H2O2 in osteoblasts, has a protective effect on MC3T3-E1 of osteoblasts induced by hydrogen peroxide (Qi et al., 2020). Hyperoside can induce MC3T3-E1 differentiation of mouse preosteoblasts, and it participates in the process of promoting and inhibiting osteoclast formation (Hou et al., 2020).
After an artificial joint replacement, joint aseptic loosening often occurs, which maybe related to titanium particles’ effect on osteoblast apoptosis and autophagy. Hyperoside effectively intervenes by improving the vitality and proliferation of MC3T3-E1 cells to protect MC3T3-E1 cells from titanium particles. At the same time, the activation of the TWEEP-p38 pathway is involved in this repair process (Zhang and Zhang, 2019). Further studies revealed that hyperoside has an anti-osteoporotic effect on ovariectomized mice, which may be correlated with its inhibition of TRAF6-mediated RANKL/RANK/NF-κB signal pathway and the increase of OPG/RANKL value (Chen et al., 2018a). Artemisia annua extracts’ active components include chlorogenic acid, hyperoside, and artemisia lactone, which together inhibit osteoclast differentiation and bone resorption-related acidification that is partly achieved by down-regulating the interaction between V-ATPase and TRAF6 to reduce acidification (Lee et al., 2017). Hyperoside reduces the expression of osteopontin, sclerosin, TNF, and IL6, thereby reducing osteoblastic activity (Nash et al., 2016).
A previous experimental study in mice showed that hyperoside could exert an antidepressant effect through the monoaminergic system and up-regulation of brain-derived neurotrophic factor (Wang et al., 2016b). Hyperoside also has an antidepressant effect on CNS after intraperitoneal administration (Liu et al., 2019b). The main active components of Apocynum venetum leaves include hyperoside and isoquercitrin. Recent studies proved that the antidepressant effect of Apocynum venetum leaf extract (AVLE) on rats exposed to CUMS is similar to that of fluoxetine (10 mg/kg), which may be related to its up-regulation of hippocampal BDNF level and the inhibition of hippocampal neuronal apoptosis and oxidative stress (Li et al., 2018b; Orzelska-Górka et al., 2019). Gong et al. (2017) proposed that hyperoside can increase the expression of brain-derived neurotrophic factor BDNF in the hippocampus of rats induced by chronic mild stress, thus reversing the cognitive impairment caused by CMS and improving cognitive function. In the in vitro model of depression induced by corticosterone, hyperoside reduces the expression of BDNF and CREB genes up-regulated by Ca2+ through cAMP-CREB signaling pathway, which protects PC12 cells from corticosterone-induced neurotoxicity. The cellular mechanism of hyperoside protecting PC12 cells from corticosterone-induced neurotoxicity is closely correlated with the cAMP signaling pathway (Zheng et al., 2012). In the FST model, hyperoside displayed strong antidepressant activity by mediating and activating D2-like receptors through the dopaminergic system (Haas et al., 2011). It has recently been proposed that the extract of Hawthorn fruit can improve memory impairment in mice with Alzheimer’s disease induced by β-amyloid protein. Different from the general drug action, CPE can block the accumulation of Aβ in a concentration-dependent manner (Lee et al., 2019).
Hu et al. (Liu et al., 2012) agreed that Hawthorn ethanol extract containing hyperoside could affect lipid metabolism and significantly reduce the levels of triglyceride, total cholesterol, and low-density lipoprotein cholesterol in hyperlipidemic rats. Totally, 15 lipid metabolites, including threonine, aspartic acid, and glutamine, were identified as potential biomarkers of hyperlipidemia. Berkoz (Hu et al., 2019) discovered that hyperoside could inhibit the transformation of early fat to mature fat as 10 μM hyperoside has an inhibitory effect in the late stage of adipogenesis, 5 μM hyperoside has an anti-lipid effect in the early stage of adipogenesis, and high dose hyperoside could reduce lipid accumulation in mature adipocytes, while low dose hyperoside could inhibit adipogenesis. Zhang et al. (Berkoz, 2019) proved that hyperoside extracted from Zanthoxylum bungeanum leaves also have a certain lipid-lowering effect and can reduce LDL-C and increase HDL-C.
Platelet aggregation and thrombosis are the common pathogenesis of many vascular diseases, so anticoagulation and inhibition of platelet aggregation are the focus of many new drug researches and development. Although there are few reports on the anti-platelet effect of hyperoside, this does not negate its role in the vascular system. A simultaneous experiment in vivo and in vitro revealed that (Lee et al., 2019) hyperoside could inhibit platelet aggregation induced by collagen or thrombin in vitro, while it had enhanced antithrombotic effect in the model of arterial thrombosis and pulmonary embolism in vivo. Both isorhamnetin-3 murine O-galactose and hyperoside in water celery extract could inhibit the activity of thrombin and activator FXa and partially prolong activated thromboplastin time and plasma prothrombin time (PT). At the same time, both of them could inhibit the production of plasminogen activator inhibitor-1 induced by TNF-α; however, the anticoagulant effect and fibrinolytic activity of hyperoside were lower than those of isorhamnetin-3 murine O-galactose (Ku et al., 2014). In the model of vascular smooth muscle cells cultured with oxidized low-density lipoprotein in vitro, hyperoside could inhibit the proliferation of vascular smooth muscle cells through the oxLDL-LOX-1-ERK pathway and the activation of VSMCs and ERK (Zhang et al., 2017b).
The above-reported studies on the pharmacological effects of various aspects of hyperoside revealed that hyperoside has many pharmacological properties, different mechanisms of action, and different dosages in the treatment of various diseases. The pharmacological mechanism of hyperoside, its effect on the different cells or animals, and the dosage are summarized in Table 1 and Figure 3.
| Mechanism | Animal/cell (organ) | Model/stimulation | Dosage | Time | Effects | Ref |
|---|---|---|---|---|---|---|
| Anti-tumor effects | Pancreatic cancer cells | N/A | 10 mg/kg | 21 day | Bax/Bcl-2, Bcl-xL↑, NF-κB,NF-κBdownstream gene products↓ | (Liu et al., 2005) |
| Murine pre-osteoblast MC3T3-E1 cells | N/A | 5 μM | 4 week | Differentiation of murine pre-osteoblast MC3T3-E1 cells↑ | (Zhang and Zhang, 2019) | |
| Non-small cell lung cancer cell lines | 1% O2 | 100 μM | 24 h | AMPK phosphorylation, HO-1 expression↑ | (Chen et al., 2020) | |
| Balb/c mice | N/A | 50 mg/kg | 18 day | Viabilityand migration capability of MCF-7 and 4T1 cells, ROS production, activation of NF-κB signaling pathway↓ | (Qiu et al., 2019) | |
| HeLa and C-33A cells | N/A | 0.25, 0.5, 1, 2, 4, 8 mM | 24, 48, 72 h | C-MYC and TFRC expressions↓ | (Li et al., 2012a) | |
| A549 cells | N/A | 100 μg/ml | 24 h | Induce A549 cell apoptosis and G1/S phase arrest; protein phosphorylation of p38 and JNK↑ | (Li et al., 2018a; Yang et al., 2017c) | |
| A549 cells | N/A | 1, 2, 5 μM | 12, 24, 48 h | Nm23-H1, MTA1, TIMP-2 and MMP-2/9 expression↑, p-ATK and p-P38↓ | (Yang et al., 2017b) | |
| A549 cells | N/A | 2.5–25 mM | 30 day | Activate caspase3, inactivate NF-kB | (Lü, 2016) | |
| A549 cells, H1975 cells | N/A | 20, 40, 60, 80,100 mg/ml | 12, 24, 36, 48, 72 h | Anti-apoptotic factors expression↓, pro-apoptotic factors expression↑ | (Liu et al., 2016) | |
| Ovarian cancer cells | N/A | 50 and 100 μM | 72 h | PGrMC1-dependent autophagy to induce apoptotic cell | (Zhu et al., 2017) | |
| Colorectal cancer cell lines | N/A | 12.5, 25, 50 μM | 48 h | Inducing cell cycle G2/M phase arrest; expression of p53 and p21↑ | (Zhang et al., 2017a) | |
| SW579 cells | N/A | 5, 10, 20 mg/ml | 24 h | Expression of Fas and FasL mRNAs↑, expression of surviving protein↓ | (Liu et al., 2017) | |
| HT 29 cells lines | N/A | 0-200 μm | 24 h | Modulate Bcl 2 associated X protein and B cell lymphoma 2 expression | (Guon and Chung, 2016) | |
| HL-60 myeloid leukemia cell lines | N/A | 10, 20, 50 μmol/L | 48 h | BAD phosphorylation↓, p27 levels↑, reactivate caspase-9 | (Zhang et al., 2015) | |
| Osteosarcoma cell lines | N/A | 150 mg/ml | 24 h | Inducing G0/G1arrest | (Li et al., 2014) | |
| C6 gliobastoma cells | N/A | 1 μm | 6 day | b1AR density in plasma membrane, downstream signalling↓ | (Jakobs et al., 2013) | |
| RL952 cells | N/A | 0 to 500 μM | 24, 48, 72 h | Induce RL952 cells apoptosis via a Ca2+-related mitochondrion apoptotic pathway | (Li et al., 2012a) | |
| Anti-inflammatory effects | Human FLSs | LPS | 1 μg/ml | 3 week | TNF-α, IL-6, IL-1, MMP-9↓ | (Li et al., 2005) |
| SD rats | 2,4,6-TNBS | 25–100 mg/kg | 10 day | PGE2, TNF-α, PGE2, TNF-α, IL-1β, CRP↓, NF-κB signaling pathway↓ | (Liu et al., 2020) | |
| HDFs cells | UVB | 0.5, 1, 1.5, 2 mg/ml | 24, 48, 72h | MMP-9, inflammatory pathway↓ | (Kurt-Celep et al., 2020) | |
| SD rats | Suture-occlusion | 50 mg/kg/d | 15 day | TNF-α, IL-1β↓ | (He et al., 2019) | |
| HUVECs | LPS | 10, 20, 50 μmol/L | 12 h | mRNA expression of IL-1β, IL-6, TNFα and iNOS↓ | (Zhou et al., 2018) | |
| Murine VSMC | TNF-α | 0–10 μg/mL | 24 h | TNF-α mediated increase in VCAM-1 expression, monocyte adhesion to VSMCs↓ | (Jin et al., 2016) | |
| BALB/c mice | OVA | 5, 10, 20 mg/kg | 3 day | inflammatory cells infiltration, levels of IL-4, IL-5, IL-13, IgE↓ | (Ye et al., 2017) | |
| Male C57BL/6 mice | 3.0% DSS | 80, 120 mg/kg | 15 day | TNF-α, IL-6, COX-2, NF-κB p65↓; IL-10↑ | (Yang et al., 2017a) | |
| Microglial cells | LPS | 2.5, 5, 10, 20 μM | 24 h | IL-1β, TNF-α, expression of inducible nitric oxide synthase | (Li et al., 2016) | |
| Male C57BL/6 mice | LPS | 20, 50 μM | 6 h | HMGB1 signaling pathway↓ | (Liu et al., 2005) | |
| Male C57BL/6 mice | LPS | 0.5-5 μM | 1 h | NF-κB activation and IκB-α degradation, TNF-α, IL-6, NO↓ | (Kim et al., 2011) | |
| Male C57BL/6N mice | Ligate Carotid arteries | 50 mg/kg | 2 week | Activate Nur77 receptor | (Chen et al., 2020) | |
| VSMCs | oxLDL | 50 μg/mL | 24 h | LOX-1 expression, ERK activation, cell proliferation↓ | (Huo et al., 2014) | |
| ICR mice | N/A | 2.3 mg/kg | 24 h | Extrinsic and intrinsic blood coagulation pathways↓ | (Ku et al., 2014) | |
| HMC-1 cells | TSLP | 100, 160 μg/mL | 2 h | IL-1β, IL-6, IL-1βmRNA, IL-6RNA, calcium/RIP2/caspase-1/NF-κB↓ | (Han et al., 2014) | |
| ECV304 cells | AGEs | 50 μg/mL | 1 h | RAGE expression | (Sun et al., 2017) | |
| Antioxidant effects | Granulosa cells | H2O2 | 10, 20, 40 μM | 24 h | Bax expression↓; Bcl-2 expression,mRNA and protein levels of SHH,Gli1, and SMO↑ | (Wang et al., 2019) |
| SD rats | microaneurysm clamp | 50 mg/kg/d | 5 day | Activity of malondialdehyde,expression of caspasee3↓;activities of SOD and glutathione peroxidase, expression of heme oxygenase 1 and NAD(P)H↑ | (Shi et al., 2019) | |
| IR-induced AKI mice | microaneurysm clamps | 20 mg/kg | 2 h | IR-induced mitochondrial fission↓ | (Wu et al., 2019) | |
| SD rats | Suture-occlusion | 50 mg/kg/d | 15 day | SOD,GSH-Px, MDA,Expression of Bcl-2 mRNA, p-PI3K and p-AKT proteins↑;Expression of Bax and caspase-3 mRNA proteins↓ | (Kurt-Celep et al., 2020) | |
| Saccharomyces cerevisiae | H2O2, CCl4, Cd2+ | 10, 20, 40 mg/L | 72 h | Depend on the intercellular ROS detoxification system | (Shen et al., 2019) | |
| CB-HSPC | SCF, TPO, FL | 1 μM | 7, 14 day | ROS level↓, protect cells from apoptosis | (Chen et al., 2019) | |
| Human kidney-2 (HK2) cells | Oxalic acid (OA) | 50, 100, 200 μM | 4 h | Involves the Nrf2/HO-1/NQO1 pathway | (Kim et al., 2018) | |
| Male Kunming mice | CCl4 | 100, 200, 400 mg/kg | 4 week | Activities cellular antioxidant defense enzymes ↑ | (Chen et al., 2018b) | |
| BALB/c mice | OVA | 5, 10, 20 mg/kg | 3 day | MDA↓, GSH and SOD↑ | (Jang et al., 2018) | |
| Male C57BL/6 mice | 3.0% DSS | 80, 120 mg/kg | 15 day | MDA↓, Nrf2, HO-1 and SOD↑ | (Ye et al., 2017) | |
| Primary melanocytes | H2O2 | 0.5-200 μg/ml | 2 h | Mitochondrial apoptosis signaling, p38 phosphorylation↓Activating AKT | (Zou et al., 2017) | |
| HLE-B3 cells, HUVECs | H2O2 | 100 μM, 400 μM | 12 h, 18 h | Nrf2 and its antioxidant response element-binding activity↑;Activate ERK signaling pathway | (Yang et al., 2016) | |
| HUVECs | H2O2 | 10, 15, 20 μM | 24 h | mRNA expression of Bcl-2 associated Bax↓, cleave caspase-3 and phosphorylated-p38 | (Mustapha et al., 2016) | |
| Human hepatocytes | H2O2 | 100 μM | 6 h | Activate the Nrf2-ARE signaling pathway | (Li et al., 2012b) | |
| L02 cells | H2O2 | 10–800 μM | 24 h | MAPK-dependent Keap1–Nrf2–ARE signaling pathway | (Xing et al., 2015) | |
| SD rats | NMDA | 1, 10 μM | 24 h | Modulate NR2A- and NR2B-containing NMDA receptors | (Xing et al., 2011) | |
| Chinese hamster | H2O2 | 1, 2.5, 5 μM | 30 min | Antioxidant enzyme activity↑, scavenge intracellular ROS | (Zhang et al., 2010) | |
| PC12 cells | H2O2/TBHP | 10 ~ 120 μg/ml | 8 h | Prevent PC12 cells from shrinking and apoptosis | (Wu et al., 2019) | |
| Balb/c mice | UVA and blue light | 25, 50, 100 mg/kg | 7 day | AP-1 and NF-kB activities, C3 activation and PARP cleavage↓ | (Zhang et al., 2018a) | |
| Cardiovascular protection | SD rats | Thoracic aortic constriction | 100, 200 mg/kg | 6 week | Improve cardiac function, HW/BW↓, H9C2 cells apoptosis↓,H9C2 cells autophagy↑ | (Guo et al., 2020) |
| Male C57/BL6 mice | Aortic banding | 20 mg/kg/d | 7 week | Block activation of the AKT signaling pathway | (Wang et al., 2018b) | |
| Cardiomyocyte of rats | High-glucose | 4, 8, 20 nmol/L | 48 h | Activate PI3K, AKT, Nrf2 signaling pathway | (Wang et al., 2018a) | |
| SD rats | Hypoxia/reoxygenation | 25 μM | 12 h | Bnip3, Bax, cleaved caspase3↓; expression of Bcl-2↑ | (Xiao et al., 2017) | |
| Wistar rats | I/R injury | 50 μM | 1 h | ER stress↓, activate downstream Nrf2 signaling | (Hou et al., 2016) | |
| SD rats | I/R injury | 50 mM | 10 min | Activate ERK signaling pathway | (Li et al., 2013) | |
| Neuroprotection | Male Albino Swiss mice | PCPA | 1.875, 3.75, 7·5 mg/kg | 60 min | Brain-derived neurotrophic factor (BDNF) expression↑ | (Orzelska-Gorka et al., 2019) |
| Male Wistar rats | chronic unpredictable mild stress (CUMS) | 30, 60, 125 mg/kg | 4 week | Oxidative stress, hippocampal neuronal apoptosis↓hippocampal BDNF level↑ | (Li et al., 2018b) | |
| PC12 cells | Corticosterone | 2.5, 5, 10 μg/ml | 48 h | Expression of BDNF and CREB through the signal pathway AC–cAMP–CREB↑ | ||
| Male CF1 mice | Forced Swimming Test | 10, 20, 40 mg/kg | 2 h | Activate D2-Like Receptors | (Haas et al., 2011) | |
| Hepatoprotection | C57BL/6 male mice | APAP | 0, 25, 50, 100 mg/kg | 7 day | Glutathione (GSH)↑, ROS production↓, activate Nrf2 and its downstream genes | (Jiang et al., 2019) |
| Male Wistar rats | Aortocaval fistula (ACF) | 100, 200 mg/kg | 4 week | ALT, AST, ALP↓, fibrosis area and hydroxyproline content, edema, liver cell vacuoles↓ | (Guo et al., 2019b) | |
| Male Kunming mice | CCl4 | 100, 200, 400 mg/kg | 4 week | Serum aminotransferases, MAO, lipid peroxidation↑ | (Chen et al., 2018b) | |
| Male Kunming mice | APAP | 10, 50, 100 mg/kg | 3 day | Expression and activities of detoxification enzymes including UGTs and SULTs↑, the activity of CYP2E1↓ | (Xie et al., 2016) | |
| LX-2 cells | N/A | 0.5, 1.0, 2.0 μM | 24, 48 h | The DNA-binding activity of NF-κB↓alter expression levels of NF-κB-regulated genes related to apoptosis | (Wang et al., 2016) | |
| Male ICR mice | CCl4 | 50, 100, 200 mg/kg | Antioxidative defense system↑, inflammatory response↓ | (Choi et al., 2011) | ||
| Renal protection | DN mice | STZ | 30 mg/kg/d | 4 week | Improve glomerular mesangial matrix expansion and podocyteprocess effacement, restore the decreased slit diaphragm protein nephrin and podocin mRNA expression and protein levels | (Zhang et al., 2016a) |
| Male SD rats | STZ | 5, 10, 15, 20 μg/ml | 72 h | Akt/mTOR/p70S6K signaling activity↓ | (Wu et al., 2018) | |
| SD male rats | D-galactose | 20 mg/kg/d | 8 week | AMPK-ULK1-mediated autophagy↓ | (Liu et al., 2018a) | |
| Male Balb/c mice | ADR | 20 mg/kg/d | 14 day | Mitochondrial fission↓ | (Chen et al., 2017) | |
| Female C57BL6 mice | STZ | 10, 30 mg/kg/d | 4 week | Increased heparanase gene (HPR1) promoter activity and heparanase expression↓ | (An et al., 2017) | |
| DN mice | STZ | 50, 200 μg/ml | 24 h | Caspase-3 and caspase-8 expressions↓ | (Zhou et al., 2012) | |
| Bone protection | Osteoblastic MC3T3-E1 cells | H2O2 | 0.3 mM | 24 h | MAPK signaling pathway, oxidative damage↓ | (Jakobs et al., 2013) |
| MC3T3-E1 cells | N/A | 200, 400 μg/ml | 6 h | Activation of the TWEAK-p38 pathway, cell viability, proliferation↑; Apoptosis, autophagy↓ | (Zhang and Zhang, 2019) | |
| RAW 264.7 cells | RANKL | 1~20 μg/mL | 5 day | Interaction of V-ATPase with TRAF6↓ | (Lee et al., 2017) | |
| Other effects | Male Kunming mice | 0.04% alloxan | 50, 100, 200 mg/kg | 6 weeks | Improve glucose tolerance, serum insulin level↓ | (Zhang et al., 2018b) |
| A549 cell | Mycoplasma pneumoniae | 0, 100, 200, 400 μg/ml | 24 h | Proliferation of MP-infected A549 cells↑, MP-induced IL-8 and TNF-α production↓ | (Kim et al., 2011) | |
| Male C57BL/6N mice | Ligate Carotid arteries | 50 mg/kg | 2 week | Activate Nur77 receptor | (Liu et al., 2020) | |
| VSMCs | oxLDL | 50 μg/mL | 24 h | LOX-1 expression, ERK activation, cell proliferation↓ | (Zhang et al., 2017b) | |
| ICR mice | N/A | 2.3 mg/kg | 24 h | Extrinsic and intrinsic blood coagulation pathways↓ | (Ku et al., 2013) | |
| HMC-1 cells | TSLP | 100, 160 μg/mL | 2 h | IL-1β, IL-6, IL-1βmRNA, IL-6RNA, calcium/RIP2/caspase-1/NF-κB↓ | (Liu et al., 2019a) | |
| ECV304 cells | AGEs | 50 μg/mL | 1 h | RAGE expression | (Zhang et al., 2013) | |
| Pseudomonas aeruginosa | N/A | 8, 16, 32, 64, 128, 256 μg/ml | 24 h | Twitching motility, adhesion, expression of lasR, lasI, rhlR and rhlI genes↓ | (Sun et al., 2017) |
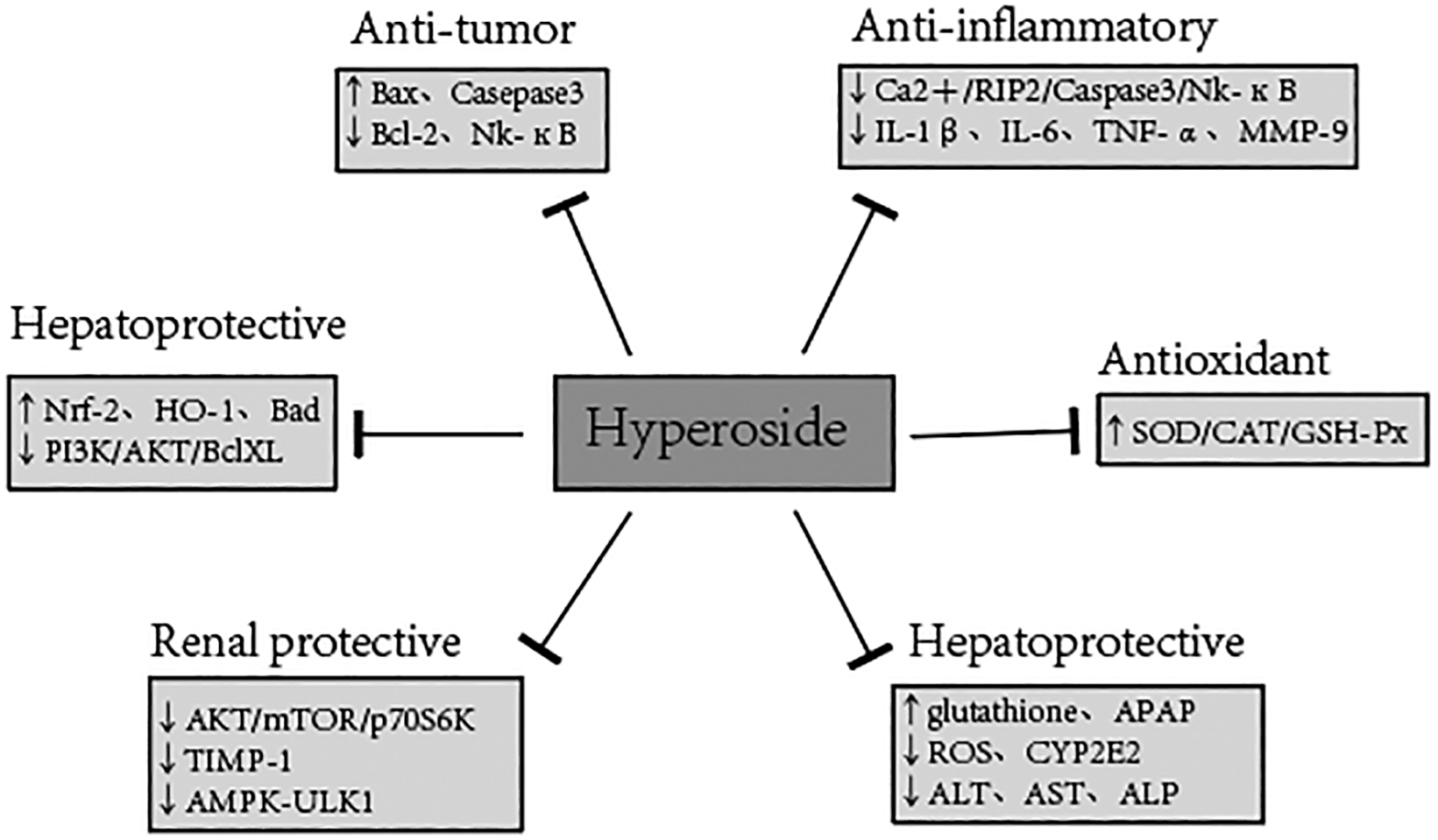
In recent years, the research on hyperoside has been greatly increased, mainly focusing on its pharmacological activity. For example, hyperoside can significantly increase the activity of antioxidant enzymes, while protecting the heart and cerebral ischemia/reperfusion injury, but also enhance immunity, liver protection, anti-depression, prevention of cardiac hypertrophy by blocking the activation of AKT signaling pathway and protecting heart remodeling caused by pressure overload (Wang et al., 2018a), inhibiting tumor cell proliferation, promoting tumor cell apoptosis, and preventing tumor cell cycle, etc. On the contrary, there are few studies on the toxicology and pharmacokinetics of hyperoside. Consequently, more clinical trials are needed for the accumulation of reliable research data under the guidance of clinical applications and medical practices. Chinese herbal medicine has been proven to be effective in the field of traditional medicine. However, its effectiveness and safety have not been globally accepted, as its mechanism remains unclear. Therefore, future studies should pay attention to the pharmacological activity of a single active ingredient and address the interaction among multiple components in the same drug.
Although the current pharmacological research of hyperoside has made great progress, due to the lack of systematic pharmacokinetic studies, the dosage of hyperoside in different research protocols is quite different, and some experimental evidence is contradictory (Guo et al., 2012). At the same time, there are few studies on the toxicology and safety of the substance, and different administration methods have also led to differences in the bioavailability and pharmacological activity of the drug. Therefore, this article makes a comparison of the pharmacological effects of hyperoside, provides a complete summary and a new perspective on the limitations of current hyperoside metabolism and toxicology studies, as well as a reference for further research on hyperoside.
In conclusion, hyperoside has multiple bio-activities, including anti-inflammatory, anti-viral, anti-cancer, and hepatoprotective effects. Further pharmacokinetic studies on hyperoside and the systematic evaluation of its metabolites’ biological activity, the dose-time-pharmacology/toxicity relationship, and the determination of biological targets and interaction modes are urgently needed.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)