Keywords
floristic diversity, vegetation structure, vegetation distribution, forest patches, environmental factors
floristic diversity, vegetation structure, vegetation distribution, forest patches, environmental factors
Forest vegetation is an important natural resource with global ecological, economic, and social implications. Forest vegetation is an important component of the biogeochemical cycle and aids in the protection of the earth from natural and anthropogenic risks, particularly in moderating climate change. Many economic lifestyles rely on trees and shrubs for food, timber, medicinal supplies, and textiles. Religious and cultural activities depend heavily on forest resources. As a result, forest vegetation is critical to sustaining the global, national, and local balance of nature (Coomes et al., 2016). However, due to deforestation and overuse, these vital resources are dwindling. Urgent resource valuation according to site features is required for conservation and long-term use (Nzabarinda et al., 2021).
The interaction of living and non-living elements influences the dynamics of forest ecosystem vegetation (Cormack et al., 1979; Kent, 2012; Franklin, 2013). The principal sources of temporal and spatial changes in vegetation are natural plant development processes and disturbances (Hull et al., 2018). Anthropogenic and natural disturbances have a significant impact on forest ecosystem vegetation, with time and place variations. Time is considered to be a key factor in vegetation dynamics (Cormack et al., 1979). Rainfall, temperature, wind, light, humidity, and other climatic variables change throughout time and influence ecological variables that affect plant ecology and evolution. Furthermore, changes in population development contribute to a rise in anthropogenic disturbances throughout time. The interactions of natural and manmade disturbances cause ongoing vegetation dynamics within the forest ecosystem (Tuxen, 1974; Cormack et al., 1979; Franklin, 2013; Hull et al., 2018; Sales et al., 2019).
Forest vegetation is extremely variable in terms of structure, composition, growth morphology, and geographical distribution (Kent, 2012; Hull et al., 2018). Vegetation dynamics are influenced by environmental elements such as slope, elevation, aspect, and site conditions (Dwyer et al., 2022). The degree of differences in vegetation studies, however, differs depending on whether they are conducted at the biome, ecosystem, community, or population level (Musau et al., 2016). There are variations in structure and growing conditions even within the same species of vegetation. Primary features that fluctuate in place and time are species makeup, structure, and growth performance (Belay et al., 2013; Jyothi and Sureshkumar, 2018; Aide et al., 2019; Angessa et al., 2020; Dang and Trung, 2021). These parameters can be used to measure vegetation dynamics (Pease et al., 2022). Examining vegetation dynamics across different environmental patterns is critical for sustainable forest management and utilization. It also supports the design and administration of ecological restorations and management by identifying environmental factors that influence the spatial distribution and quantity of vegetation (Kent, 2012; Gemedo et al. 2014).
Ethiopia has a wide range of agroecological zones, from the top Ras-Dejen Mount to the lower Afar Depression, which results in a wide range of plant species typical of tropical climates (Bekele, 2007). However, human activities such as deforestation, agricultural land development, overgrazing, high demand for firewood and charcoal, and a lack of adequate legislative frameworks are causing alarming degradation of flora resources (Soromessa et al., 2004). Conservation actions, particularly in the study areas, are urgently needed to protect these Afromontane forests (Senbeta et al., 2014). Effective conservation strategies that assure long-term viability are required. Unfortunately, there have been very few scientific studies on forest vegetation distribution, diversity, and structure, as well as growth performances in relation to natural and anthropogenic causes (Caroline et al., 2020), particularly in the Awi zone, northwest Ethiopia.
As a result, the purpose of this investigation is to evaluate the diversity, composition, distribution, and growth performance of forest vegetation in relation to environmental circumstances. The study also investigates the impacts of slope, aspect, altitude, and agroclimatic zone on vegetation diversity, structure, and composition, while considering the interaction of human and livestock disturbances. The study gives important scientific knowledge for the protection and long-term management of natural forest vegetation. It also provides an initial for policymakers to use when developing and implementing forest restoration, environmental management, and climate change mitigation strategies.
The study was carried out in the Amhara region’s Awi administrative zone, which is located between 11° to 10°85’N latitude and 36°39’60” to 36°57’E longitude. The study area has an altitudinal range of 600 to 3500 m. a. sl., 1750 mean annual rainfall, and temperatures ranging from 170°C to 27°C. Awi (Agew-mider) has a rather level topography (Figure 1).
Agroecologically, most of the Awi administrative Zone is highland (17%), mid-altitude (72%), and lowland (11%). The rainfall distribution in the area is ideal. Geographically, the Awi zone has a total area of 8,935,520 ha of land, of which 297,133 ha (33.25%) is agricultural land and 354,396 ha (34.02%) are forest land (76,554 ha plantation and 277,842 ha natural forest). The remaining land is rangeland and grazing land, covering 217,138 ha (24.3%), and other land uses, including as infrastructure and settlement, covering 74,853 ha (8.38%) (Awi Zone Agricultural office, 2021).
Agew-mider (Awi) has a total population of 1,159,386. The urban population accounts for 13% of the total population; the rest live in rural areas and rely on mixed farming lifestyles (Genet, 2022). Currently, the people in the highlands parts practices Acacia mearnsii forest farming in the Taungya farming system. The small-scale Acacia mearnsii woodlot is responsible for reclaiming acidic soil and enhancing soil fertility, but its major function is to generate money through charcoal manufacture (Wondie and Mekuria, 2018). One of the studied districts, Jawi, has a significant potential for natural forest cover (48.9%), followed by Guangua (20.5%) and Ankesha (8.27%).
Highland (2200-3600 m.ab.sl.), Mid-altitude (1300-2200 m.ab.sl.), and Lowland (500-1300 m.ab.sl.) were the primary Awi Zone classifications (Bekele, 2007; Gorfu and Ahmed, 2011). One or two districts in each agroclimatic zone were purposely chosen based on the potential for natural forest cover. To obtain sufficient biophysical data, three forest patches with high area coverage were chosen in each target district and agroclimatic zone (Alemayehu, 2007). Systematic sampling was used to acquire biophysical data in the forest under each designated forest patch. Then, using GPS and GIS tools, transects and plots were put out based on area coverage. The transect and plots were 200 meters apart. Transects were laid across the elevation gradient, followed by transects and then plots. The main plot was 20m by 20m for trees, with two 10m by 10m sapling subplots, five 5m by 5m seedling and regeneration subplots, and five 1m by 1m herbaceous species subplots within the main plots. The number of plots investigated in a particular forest varies according to the species-area curve (Kent, 2012).
To achieve this, vegetation and environmental data from nine forest patches in 72 plots were collected. Following standard inventory procedures, tree growth parameters (height and DBH) were measured with a hypsometer and diameter tape, respectively, and the life form of a species was clustered as trees (DBH > 2.5 cm and height of >2.5 m), saplings (DBH 2.5 cm and height of 1m h 2 m), and seedlings and herbs (DBH 2.5 cm and height of 1 m) at each plot (FAO, 2004; Kent, 2012). Site parameters such as slope class (gentle slope = 1-10%, mid-slope = 10.1-20%, and upper slope = 20.1-30%), aspect angle data were collected at each plot and classified as Northeast (NE), Northwest (NW), Southeast (SE), and Southwest (SW), and elevation were classified for individual forest patches were collected using GPS. The species-area-curve model was used to determine the optimal number of plots (FAO, 2006).
The vegetation characteristics were computed and compared with biodiversity indices such as the Shannon Weiner diversity index (H’), Sorenson’s similarity index (Ss), and Shannon Weiner evenness index (SEI) (Gotelli & Anne, 2013; Naidu and Kumar, 2016).
The species diversity index with the Shannon-Wiener index:
Where: H’ = Diversity of species S = Number of species Pi = the proportion of individuals abundance of the ith species
Sorenson’s similarity Coefficient (Ss)
Where: a = number of species common to both habitats; b = number of species present in the first habitat but absent in the second; c = number of species present in the second habitat but absent in the first.
Shannon Weiner evenness index (SEI)
Where: H′: Shannon Weiner diversity index, S: number of species
Where: IVI = RDe = Relative density, (RDe) + RDo = relative dominance, (RDo) + RF = relative
Forest species were identified using NDA (Natural Database Africa) software Ver. 2.0 and the Flora of Ethiopia and Eritrea book (Bekele, 2007), vegetation descriptions, and environmental patterns were analyzed using Kent (2012), Zhang et al. (2013), Lemessa et al. (2017) methods. Brawn Banquets cover analysis was used to determine vegetation cover: 1% extinct or uncommon, 1-5% bare or sporadic, 6-25% rare, 26-50% patches, 50-75% interrupted, >75% continuous. The connection of vegetation traits with environmental gradients and plot distribution was determined using principal component analysis (PCA) (Kent, 2012). All statistical studies (multivariate analysis, ANOVA, and cluster analysis) were carried out using different packages of R. Ver. 4.1.1 and PAST.Vr.3.
The possibility of obtaining new species increases up to 2.4 hectares, beyond which the species richness becomes saturated. This means that species recruitment was possible up to a 2.4 ha plot area, and then the types of species recorded afterward were repetitive in highland and mid-altitude forest patches, whereas in lowland forest patches, the possibility of getting new species saturates at a 0.6 ha plot area (Figure 2).
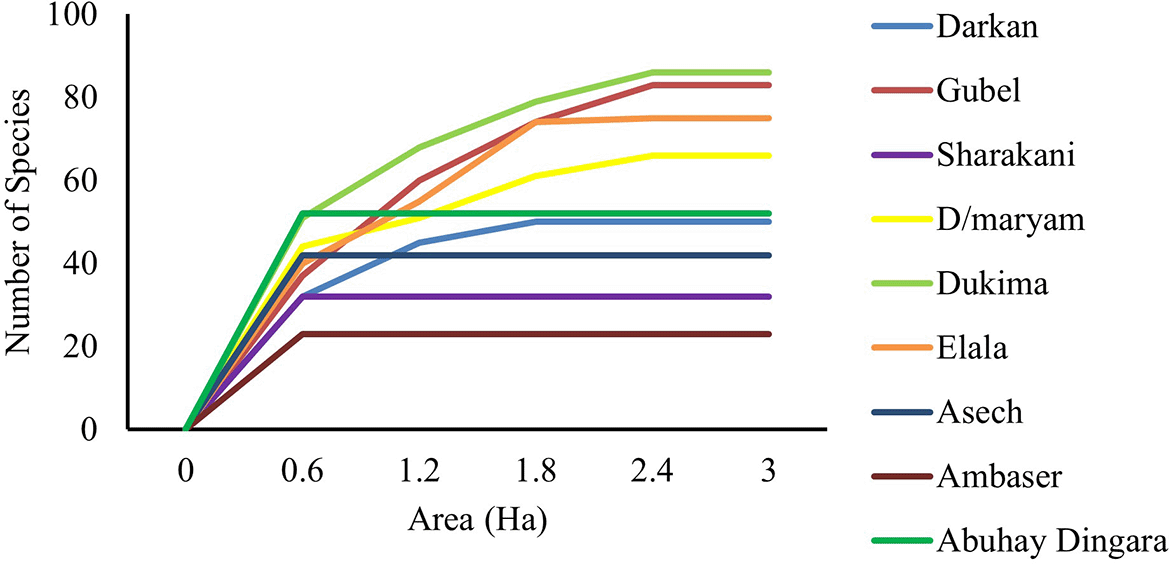
Note: Highland Forest patches: Darkan, Gubel, and Saharakani; Mid-altitude forest patches: Den Maryam, Dukima, and Elala; Lowland Forest patches: Asech, Ambaser, and Abuhay Dengara.
The species-area curve is a species distribution model that illustrates the relationship between area and species richness with an indicator of the optimal number of plots for the inventory of floristic diversity in a specific forest patch (Gray et al., 2004). At 15 plots (0.6 ha), almost all forest patches had reached saturation in terms of new species.
There is no species variability after a certain elevation range in forest patches with substantial anthropogenic disturbances. Forest patches with small-scale spatial variability and significant disturbances have comparable species, whereas forest patches with vast spatial variability contain diverse species with even distribution, and the probability of discovering new species grows with the area. In some forest ecosystems, the possibility of acquiring new species extends until the last plots before becoming saturated. The environmental gradient in tropical Africa is highly variable, resulting in a wide range of species diversity in forest patches. This theory is comparable to that of Scheiner (2003) and Rosindell and Cornell (2009), who claim that floristic species are evenly distributed across the forest and that there are new species in every aspect of the forest patches.
Floristic diversity
Floristic diversity and species richness varied significantly (p 0.008) over slope gradients in different agroclimatic zone and forest patches. Mid-altitude forest patches had the highest floristic species richness (78 species in 32 Families) (Table 2). The Fabaceae and Rosaceae families account for 39% and 29% of the total species in mid-altitude and highland forest groups, respectively. Burseraceae, Combretaceae, and Fabaceae account for 75% of all species in lowland forests. Highland and mid-altitude forest patches exhibit higher floristic diversity (2.48) than lowland forest patches (1.88). The highest floristic diversity was found in the Dukima forest (2.83), followed by the Gubel forest (2.69) and Abuhay Dengara forest (2.49) (Table 2). The species diversity on the gentle slope is the highest (2.15), followed by the medium slope (1.96) and the steep slope (1.87) (Table 1).
Mid-altitude forests have more floristic diversity and richness. This could be the transition zone between lowland and highland forest patches. Because of the optimum climatic conditions, some lowland and highland species may favor mid-altitude areas. That is, vegetation characteristics, such as floristic diversity and species availability, have site-specific patterns, with some species preferring the extreme lower altitude of the highland and others preferring the extreme higher elevation of the lowland. This idea is expressed by Hawk et al. (1977), Franklin (2013), Gemedo et al. (2014), and Senbeta et al. (2014) who show that the species richness of plant patches changes reliably along environmental gradients. Another study by White and Hood (2004) stated that floristic diversity and distribution in forest patches are influenced by topographic and underlying features. Furthermore, according to Chetcuti et al. (2022) plant community distribution in landscapes is a manifestation of environmental gradients and biotic responses to these gradients.
The gentle slope is ideal for maximizing species diversity and richness. This could be because the lower areas of a forest are more suitable and productive for most floristic species. The finding supports Gemedo et al. (2014) argument that the higher richness of species diversity in gentle and flat slope areas of the forest is attributable to optimal environmental conditions for most species of life. Similar to the study of Franklin (2013), various environmental gradients and specific slopes have a direct influence on vegetation features, most notably diversity of species and richness. It is a natural law that environmental gradients and species compatibility matching regulate vegetation dynamics, specifically floristic diversity, and richness. Environmental gradients associated with natural and anthropogenic disturbance have a substantial impact on the geographical and temporal distribution of vegetation and life forms in an open natural forest. Each disturbance event, specifically in this study area—grazing and human impact—and the resulting site features distinguish a given forest patch.
Floristic species similarity
There was significant positive species similarity (p 0.024) between and among mid-altitude and highland forest ecosystems. However, there were significant floristic species dissimilarities (p 0.001) between forest patches of highland and lowland areas. There was also significant species similarity within the lowland forest ecosystem (Figure 3).
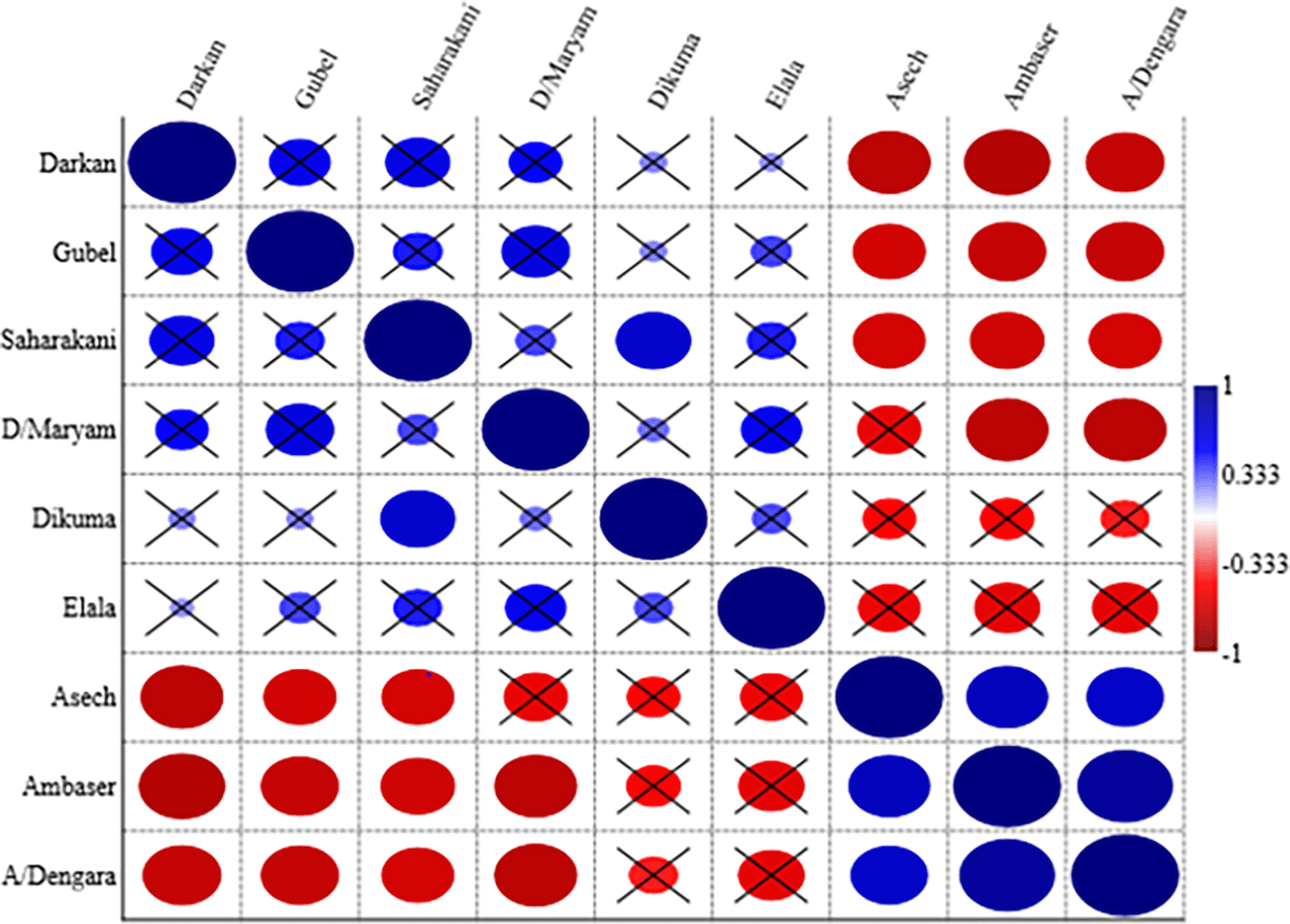
((A/Dengara = Abuhay Dengara, D/Maryam = Den Maryam); the red circle indicates a negative correlation; the blue circle indicates a positive correlation circle with X indicates a non-significant correlation (p > 0.05)).
There was a high degree of similarity (82% within lowland forest patches) and a medium degree of similarity (69% within highland forest patches). Highland and mid-altitude forests share 69% of their species presence; yet there are significant dissimilarities (68% between lowland and highland forest patches). There was also a 61% dissimilarity between mid-altitude and low-altitude forest patches.
Mid-altitude forest patches are more like highland and lowland forest patches, while the two extremes of highland and lowland forest patches are very different. Even while the degree of similarity or variability changes depending on a variety of anthropogenic and natural disturbances, the probability of species being similar within a single agroecological cluster is much higher (Della Rocca et al., 2021). The results of this study demonstrated that there was more than 50% similarity within the forest patches under the same agroclimatic zone, according to the Martín-Devasa et al. (2022) similarity index concept. Lowland forest patches, on the other hand, have unique species from highland and mid-altitude forest patches, suggesting a dissimilarity of more than 50%. According to Senbeta et al. (2014) and Lemessa et al. (2017), despite geographical distance, high floristic similarities were observed between patches located very close to each other, and dissimilar forest patches were expected if they had wide variations in altitudinal gradients and topographic aspects.
Floristic species distribution
Satellite species were identified in all elevation patterns at all forest patches, indicating that scattered species distribution with low abundance is prevalent. This revealed that there is significant deforestation and forest degradation, resulting in poor vegetation cover. Rural species distribution was similar in all elevation patterns, but the extent was less than that of satellite species. The distribution of urban species is widespread at lower and higher elevations when there is a patch of forests on the area’s hillsides. The distribution of core species was quite limited, with just a few species occurring at medium elevations where there is a transition zone and some management measures (Figure 4).
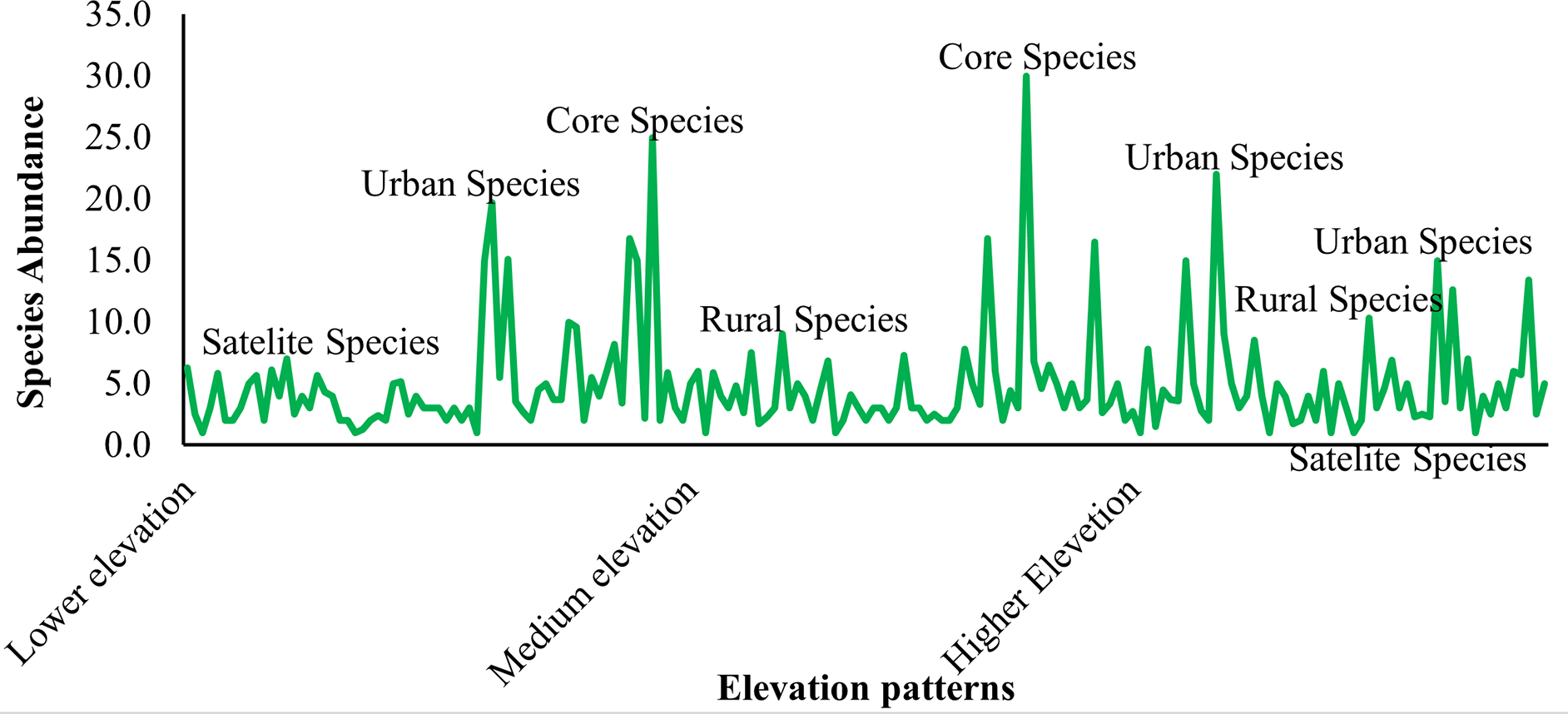
According to the hierarchical continuum paradigm Kent (2012), species with high abundance and even distribution (core species) were only found in medium elevation gradients with optimal climatic and edaphic conditions. Satellite species were detected in all agroclimatic zone and elevation gradients with low abundance and inconsistent distribution. The species were uniformly distributed and scarce in higher elevation gradients, indicating that they are rural and satellite. A variety of species distribution types were discovered in the lower elevation gradient in the order satellite > rural > urban.
Floristic species lifeform
There were a significant variation (p < 0.03) proportions among different lifeforms in the three agroclimatic zone forest patches. Tee components account for the lion’s share of all agroecological classes in each forest patch, with average contributions of 55%, 42%, and 67% in highland, mid-altitude, and lowland forest patches, respectively (Table 2).
| Lifeforms | Highland forests | Mid-altitude forests | Lowland forests |
|---|---|---|---|
| Trees | 55A | 42A | 67A |
| Shrubs | 32B | 35AB | 28B |
| Climbers | 12C | 16C | - |
| Herbs | - | 7D | 6C |
| Mean | 33* | 25* | 33.67* |
| p-value | 0.03 | 0.04 | 0.002 |
The forest in the study area is an open natural forest that has been disturbed by anthropogenic factors. This is why lower plants become extensively degraded, and forest patches are made up of living forms with tree > shrub > climber > herb composition shares. This idea is comparable to that of Meragiaw et al. (2018) who found that tree and shrub life forms predominated in open natural forest patches. It is common for herbaceous species to experience grazing issues in the open natural forest. As a result, shrubs and trees that are resistant to grazing have the highest share.
Climbers were present in some forest patches, particularly in highland and mid-altitude forest patches, which adds to the beauty of the forest in mountainous areas. This is strongly connected to human and grazing issues, which are especially severe in lowland places with few climbing plants. Life forms, notably herbs, and climbers, are highly susceptible to disturbances. According to Mahdavi et al. (2013) and Shary et al. (2020), habitat instability is the fundamental factor that affects the proportion of life forms.
Woody species structure
Lowland and mid-altitude forest patches exhibit inverted-J structures, whereas highland forests have J-shaped structures (Figure 5A & B).
However, there are extremely few seedlings in all forests and agroclimatic zone, implying that no trees have less than 5 cm DBH. The mid-latitude forest has an inverted J-shape structure based on DBH class distributions, indicating that higher forest tree populations were found in lower diameter classes, however, this does not imply that the forest is a healthy community. According to Senbeta et al. (2014), forest patches with an inverted-J shape indicate good natural regeneration and a healthy community. There were few trees having seedlings less than 5 cm in diameter in all forest patches, indicating a lack of spontaneous regeneration (Figure 5C). In highland agroclimatic zone with intermediate anthropogenic disturbances (Table 4), the vegetation pattern has a J-shape, indicating a healthy forest ecosystem.
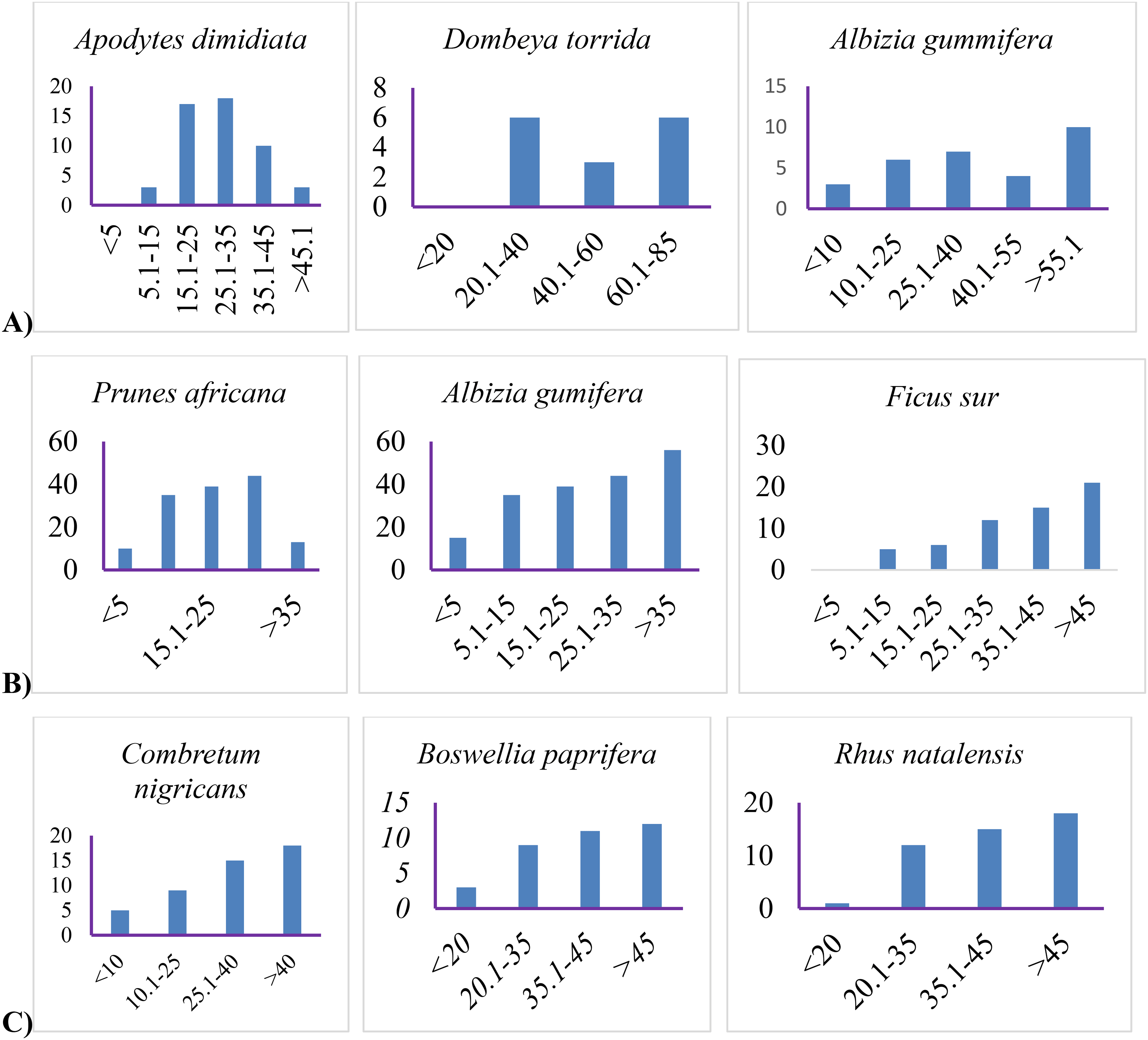
However, there is still a natural regeneration issue, and seedlings are unable to reach the sapling and tree phases. This is due to human intervention and grazing encroachment. Meragiaw et al. (2018) endorse this theory, stating that the lowest percentage of species found in the lowest and highest classes could be due to the overgrazing of young plants and selective extraction of young trees. Several scholars (Atomsa and Dibbisa, 2019; Mewded, Negash, and Awas, 2019; Dibaba et al., 2020; Tadese et al., 2021) argued that in some parts of Ethiopia woody species population structure was follow either or a combination of inverted J-shape, a J-shape, a bell-shape, and an irregular shaped pattern which is an indicator of high anthropogenic disturbances combined with environmental factors that affect natural forest community balance. As a result, such pattern variability is an indicator for immediate intervention steps to ensure the survival and sustainability of forests that are experiencing poor regeneration.
Woody species dominance
According to the important value index, Schefflera abyssinica dominated highland forest patches, followed by Dombeya torrida and Albiza gumifera, whereas Syzygium guineense was the least dominant species, despite being an ecologically and economically important indigenous woody species. Because of its large basal area, Schefflera abyssinica was the dominating species, but it had a low stem density (Figure 6A). Whereas Mimusops kummel was the dominant species in mid-altitude forest patches due to its largest basal area, Albiza gumifera, and Prunus africana were the prevalent species in highland and mid-altitude forest patches. Despite being a keystone species, Teclea nobilis had the lowest IVI in mid-altitude forest patches (Figure 6B). In lowland forest patches, Oxytenanthera abyssinica and Ficus sycomorus were the most dominant species, followed by Combretum nigricans. Ziziphus mumucronta was the least prevalent species, despite being a wild edible fruit species that plays an important ecological role in lowland forest patches (Figure 6C).
Because of their economic relevance in timber and non-timber products, the species with the least dominance have suffered degradation due to over-utilization. Several studies (Kidane, Stahlmann, and Beierkuhnlein, 2012; Sales de Melo et al., 2019; Mucheye et al., 2020) contended that species with high economic value had lower important value indexes, indicating degradation. As a result, they require unique conservation priorities that promote natural regeneration as in-situ conservation and collection in arboretums as ex-situ conservation.
Height, diameter, and stem density differed significantly (p 0.03) within and within agroclimatic zones. Mid-altitude forest patches had the highest stem density (2578 trees per ha), followed by highland (1351 trees per ha) and lowland (1268 trees per ha). Highland forest patches have the highest height (25.14m) and diameter (40.41 cm), followed by mid-altitude (19.5m height and 26.8cm diameter) and lowland forest patches (16.03m height and 25.9 cm diameter) (Table 3).
Height, diameter, and stem density differed significantly (p 0.03) across slope gradients; however, changes in aspect and elevation gradients did not contribute to a significant variance (p 0.05) within forest patches. The moderate slope had the highest stem density (2260 trees per hectare), whereas the medium slope (2124 trees per ha) and steep slope (1468 trees per ha) had the lowest (Table 1).
Highland forest patches exceed mid-altitude and lowland forest patches in diameter and height growth. Better climatic conditions and anthropogenic perturbations may be substantially connected with growth performance and growth rate. This notion is comparable to that of Toledo et al. (2011), who show that climatic and edaphic conditions fluctuate with place and time, influencing growth performance appropriately. Mid-altitude forest patches, on the other hand, outperform highland and lowland forest patches in terms of stem density. Lowland forests had a poor growth performance in any case. This could be due to human and grazing effects because there was extensive prohibited cutting and overgrazing in lowland forest patches, resulting in poor growth performance (Table 4).
Flat slopes have the highest stem density in an open natural forest, followed by medium and steep slopes. Flat slopes have higher soil fertility and ideal climatic conditions, which subsidize tree micro-sites for better plant growth and accumulation (Gemedo et al., 2014; Field, 2014; Senbeta et al., 2014). Furthermore, Afromontane forests have a high density of woody plants that vary greatly between places (Senbeta et al., 2014).
At a significant level of p 0.05, forest patches and human impacts in all agroecological clusters incur a similar degree of effect. However, there was a substantial difference in grazing impact and vegetation cover-abundance within and between agroclimatic zones (p 0.005) (Table 4). This suggests that all forest sections around the globe are being subjected to extensive tree cutting by illegal cutters and grazing encroachment. As a result, lowland forests have the biggest grazing issue, followed by mid-altitude and highland forests, and it has a substantial impact on the cover-abundance of lowland forest patches. Mid-altitude forest patches had the most abundant but interrupted vegetation cover (5 = 50-75% cover), followed by highland forests with abundant cover (4 = 26-50%) and lowland forests with less than 3 = 6-25 rare vegetation cover (Table 4). Intensive grazing and human encroachment threaten lowland forest patches. This is due to the lowland areas’ animal-based farming practices. That is why vegetation cover-abundance is greater in mid-altitude and highland forest patches than in lowland forest patches. This could be attributed to environmental compatibility as well as human and grazing intensity. Human effect was substantial in all forests and agroclimatic zone, whereas grazing impact is lowland > mid-altitude > highland.
Thus, anthropogenic disturbances with the interaction of climate variances are the primary determinants of vegetation cover-abundance. This argument is similar to Alemayehu (2007) and Cardelús et al. (2019) claim that anthropogenic disturbances are common in all forest habitats and have a detrimental impact on variety, growth performance, and vegetation cover-abundance. Furthermore, most evaluated forest patches encounter similar levels of disturbance, and the forest population is significantly threatened by habitat fragmentation, lack of regeneration, and high disturbances (Chetcuti, Kunin, and Bullock, 2022; Pinho et al., 2022).
Externally, almost all study forest patches appear intact; yet, they are open on the interior, and only large trees shield the canopy, as observed in vegetation structure (Figure 5). There is little regeneration and very little conservation. All of this is the result of extensive human encroachment and overgrazing. This result is consistent with the findings from Teketay (1992), regarding the subjectivity of most Ethiopian natural forests to strong pressure and, as a result, the risk of local extinction. In addition, White and Hood (2004) contend that grazing has an impact on natural regeneration, life in transition, and optimal productivity. As a result, grazing and human encroachment have altered growing habitats, influencing vegetation dynamics (Zhang et al., 2013; Field, 2014; Girma, Chuyong, and Mamo, 2018; Cardelús et al., 2019).
Slope, agroclimatic zone, and forest patches have a substantial influence on the vegetation features of the evaluated forest patches, with altitude and aspect having a minor influence. Forest patches in close spatial proximity have similar species diversity and composition, whereas those with large-scale altitude differences show significant disparities. Transitional forest patches that share some species between the extremes, on the other hand, show small parallels in species composition and diversity. The forests in the study area are open natural forests that are subject to anthropogenic habitat disturbances, resulting in a higher proportion of trees and shrubs. This emphasizes the importance of disturbances and habitat instability in the spread of plant life forms.
Satellite species were detected throughout all agroclimatic zone and elevation gradients, with low abundance and inconsistent distribution. This implies that native floristic species have low abundance and distribution, most likely because of deforestation and forest degradation, both of which act as indications of local species extinction. The woody species population structure in the study forest patches follows various patterns, including a combination of inverted J-shaped, J-shaped, bell-shaped, and irregular-shaped patterns, indicating high anthropogenic disturbances combined with environmental factors that affect the balance of natural forest patches, like other parts of Ethiopia.
Human activities and grazing are common in all forest patches and have a negative impact on floristic diversity, composition, growth performance, vegetation distribution, structure, and cover abundances, as well as long-term effects on habitat fragmentation. Most forest patches in various agroclimatic zones have been impacted by humans and livestock, resulting in reduced natural regeneration and high levels of deforestation and forest degradation. While the forests appear green and lush from the outside, most species have not naturally regenerated, and the canopy is dominated by massive, elderly trees that are becoming increasingly vulnerable to illegal cutting. This makes the forest patches less valuable and more vulnerable to deterioration. Woody species that are less prominent in the study forest patches have suffered deterioration because of overutilization for their economic worth in timber and non-timber products.
As a result of the observed pattern of vegetation structure, composition, and distribution variability, as well as the risk of local extinction, urgent intervention is required to ensure the survival and sustainability of sensitive woody species suffering from poor regeneration status. As a result, extraordinary conservation measures are required, with the active participation of forest researchers, development agents, and political leaders, to safeguard the few remaining natural forests. If the encroachment is not addressed, critical biological and socioeconomic forest patches will be lost soon. Conservation actions aiming at conserving vegetation and wildlife habitats should be prioritized.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)