Keywords
Cecal ligation & puncture, inflammatory mediators, Montelukast, NF-κB signaling cascade, Sepsis associated acute kidney injury
This article is included in the Advances in Fibroblast Research collection.
Cecal ligation & puncture, inflammatory mediators, Montelukast, NF-κB signaling cascade, Sepsis associated acute kidney injury
In this updated version of the article, several important revisions have been made to enhance the clarity of the study. The Introduction has been expanded to provide a more detailed background on sepsis-induced kidney injury, and repetitive statements have been removed to improve readability. Vehicle control clarification, vehicle control group (10% DMSO in saline) showed results comparable to the CLP group, indicating no significant effect on the studied markers. Thus, the protective effects are attributed to montelukast. Histological images have been updated, with some replaced by higher-resolution versions, scale bars added, and standardized arrows used instead of hand-drawn annotations to improve interpretability. The role of montelukast in reducing KIM-1 levels has been clarified and linked to the ERK1/2–STAT3–KIM-1 signaling pathway. Minor editing changes have also been made throughout the manuscript to improve readability and overall presentation.
To read any peer review reports and author responses for this article, follow the "read" links in the Open Peer Review table.
Sepsis is a life-threatening condition caused by infection and characterized by systemic organ dysfunction. It is driven by an uncontrolled immune response, cytokine storm, and oxidative stress, which can culminate in multiple organ failure, eventually leading to death.1–3 Sepsis is primarily triggered by microorganisms, such as bacteria, viruses, and fungi, and can affect various organs, including the heart, kidney, liver, and central nervous system.4,5 Acute Kidney Injury is a common outcome associated with the clinical scenario of sepsis.6 The pathogenesis of AKI in sepsis is multifaceted. Among these contributing factors, oxidative stress and inflammation are pivotal etiological agents of septic AKI.7 Sepsis-associated acute kidney injury originates from intricate, heterogeneous mechanisms that culminate in renal injury. These mechanisms may arise directly from the infectious agent and the corresponding host immune response, or may represent indirect ramifications of sepsis or its therapeutic intervention. Various pathophysiological mechanisms may interact and participate in AKI in patients with sepsis, including systemic and renal inflammation, macrocirculatory anomalies, microcirculatory dysfunction, metabolic reprogramming, mitochondrial impairment, and dysregulation of the renin-angiotensin-aldosterone system (RAAS).8 Inflammation, recognized as a critical element in sepsis, appears to significantly influence the pathogenesis of SA-AKI. Pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) can stimulate toll-like receptors (TLRs). Among these receptors, TLR-2 and TLR-4 are expressed on the surface of tubular epithelial cells within the renal system.9 The engagement of TLR-2 and TLR-4 initiates a cascade of inflammatory responses, which is marked by the secretion of pro-inflammatory cytokines, including IL-1α, IL-6, IL-8, and TNF-α.10 In parallel, sepsis induces profound alterations in cellular energy metabolism, leading to metabolic reprogramming of immune cells. This adaptive shift, characterized by a transition from mitochondrial oxidative phosphorylation to glycolysis, supports the increased energy demands of the inflammatory response during the early phase of sepsis. However, sustained metabolic reprogramming ultimately promotes mitochondrial dysfunction, oxidative stress, and subsequent organ injury.11 Both macrovascular and microvascular dysfunction contribute to the development of SA-AKI. While hypotension-induced macrocirculatory impairment reduces renal perfusion and glomerular filtration, increasing evidence highlights the critical role of microcirculatory dysfunction.12 In particular, inflammatory signaling, oxidative stress, endothelial damage, and microthrombus formation.13 In addition, the mitigation of blood flow in the capillaries is also accompanied by capillary leakage and early renal interstitial edema.12 Cellular injury in SA-AKI involves multiple forms of regulated cell death, including apoptosis, necroptosis, pyroptosis, and ferroptosis, all contributing to tubular damage and renal dysfunction. Apoptosis is a caspase-dependent, relatively controlled process,14 whereas necroptosis15 and pyroptosis16 are highly inflammatory and amplify tissue injury. Ferroptosis, an iron-dependent form of cell death driven by lipid peroxidation and oxidative stress, has emerged as a potential contributor to renal damage in SA-AKI.17 The interplay among these pathways highlights the complexity of renal injury during sepsis. In response to cellular stress, adaptive processes such as autophagy play essential roles in maintaining cellular homeostasis. Autophagy facilitates the removal of damaged organelles, thereby limiting cellular injury and promoting survival during the early stages of sepsis.18 This process is regulated by autophagy-related genes, including Atg5 and Atg7, which have already been documented to be involved in the development of kidney disease,19 and interact with signaling pathways such as mTOR, sirtuins (SIRT1, SIRT3, and SIRT6), and NF-κB in SA-AKI.20 Apart from autophagy, efficient efferocytosis is required for the timely clearance of dying cells to prevent excessive accumulation of DAMPs and sustained inflammatory signaling.21 In this context, apoptosis inhibitor of macrophages (AIM) binds to IgM, preventing its renal excretion. However, during AKI, AIM dissociates from IgM, is filtered through the glomeruli, and binds to KIM-1 in renal tissue, thereby promoting SA-AKI through efferocytosis.22 Among the molecular pathways implicated in SA-AKI, NF-κB signaling plays a pivotal integrative role in coordinating inflammatory, immune, and stress responses.23 Under normal circumstances, inflammation is beneficial to the host and typically self-limiting. However, uncontrolled inflammation can lead to persistent tissue damage, leading to acute or chronic inflammatory diseases.24 Furthermore, the TLR4/NF-κB signaling pathway is regarded as a central pro-inflammatory signaling cascade, known as the switch to initiate and stop inflammation.25 Pro-inflammatory cytokines may activate NF-κB, amplifying the initial inflammatory signal, and trigger a vicious cycle that exacerbates the inflammatory response and further damages the microcirculation of the body.26
The current investigation was conducted using a cohort of 40 albino Swiss mice, with a weight spectrum of 25–30 g and an age range of 8–12 weeks. All euthanizing procedures were conducted under a combination of ketamine- and xylazine-based anesthesia, with diligent efforts made to mitigate individual distress. The experimental methodologies and protocols employed in this study were approved on August 29, 2024, under reference number (20553) from the ethics committee for the care and utilization of laboratory animals.
The current study was done with a total of 40 albino Swiss mice, aged 8-12 weeks and weighing 25-30 g, obtained from the College of Science, University of Kufa. These animals were housed in the animal house within their cages under 12:12 light: dark cycles with 25°C room temperature, 60-65% humidity, and free access to food and water ad libitum. All animal procedures were conducted based on the guidelines of the Kufa University and were approved by the Institutional Animal Care and Use Committee (IACUC). The experimental protocol was reviewed and approved by the Research Ethics Committee, Faculty of Pharmacy, Kufa University under approval number (20553 on August 29, 2024). All efforts were made to minimize animal suffering, encompassing the use of appropriate anesthesia (xylazine of (20 mg/ml) and ketamine of 100 mg/ml), and animals were monitored closely throughout the study. Humane endpoint and euthanasia protocols were strictly followed through cardiac puncture following intraperitoneal injection of thiopental sodium (50 mg/kg) to induce deep anesthesia to ensure a painless and ethical procedure.
In the current study, CLP used in earlier studies, including those performed by Refs. 27, 28, was employed to induce sepsis in animals. It is introduced by making a midline incision measuring 1.5 cm while the subject is under general anesthesia, wherein xylazine of (20 mg/ml) and ketamine of 100 mg/ml are mixed (2:1) and administered.29 The cecum was ligated, situated below the ileocecal junction, and punctured twice with a cutting cannula to inflict kidney organ damage during the first 24 h of acute sepsis. Subsequently, the puncture hole was used to squeeze a tiny amount of fecal material from behind the perforation site. Thereafter, the anterior abdomen was sutured to prevent leakage. Sham mice were subjected to the same procedures, except that CLP was not performed.
Mice were classified into the subsequent four distinct groups (n=10):
Sham group: Evidently, mice exhibited no apparent signs of disease.
CLP cohort: Mice belonging to this cohort experienced a Cecal Ligation and Puncture surgical procedure.
Vehicle group: Murine classified within this category was administered an equivalent volumetric measurement of the solvent dimethyl sulfoxide (DMSO) intraperitoneally; CLP was performed after 1 h, and then the animals were euthanized after 24 h.
Montelukast group: Mice affiliated with this group received montelukast 20 mg/kg intraperitoneally30; CLP was performed after 1 h, and then the animals were euthanized after 24 h.
After 24 h, the mice were euthanized under anesthesia and blood was collected using the direct heart puncture method. Blood was left in a gel tube rack for approximately 20 min to allow for clot formation, after which it was centrifuged at 10,000 rpm for approximately 10 min. Then, the supernatant was retained at -20°C for sequences of biochemical assessment.31 The right kidney tissues were fixed in 10% formaldehyde for histological investigation (H&E) and IHC analysis,32 whereas the left kidneys of different groups were collected and divided into two sections. The first section was homogenized in cold phosphate-buffered saline (PBS) (pH 7.4), and the supernatants were used for ELISA. The remainder of the tissue (1/3) was immersed in the TRIzol reagent for gene expression measurement. Until analysis, both the tissue homogenate supernatants and the TRIzol-treated tissue samples were kept at -80°C.33
After fixation with 4% paraformaldehyde for 24 h, the right kidney was embedded in paraffin. H&E and periodic acid-Schiff (PAS) reagents were used to stain 4 μm thick sections cut from the renal tissue wax blocks. Tissue damage was scored as follows: 0, no harm; 1, 0–25; 2, 25–50; 3, 50–75; 4, >75%.34
BUN and Cr concentrations were measured using a Biolis colorimetric assay kit (Biolis, Tokyo, Japan).
ELISA kit instructions (Sunlong, China) were followed while processing tissue samples kept in a refrigerator at -80°C for evaluation of TNF-α, IL-1β, F2-isoprostane, and caspase-3, in addition to measurement of serum KIM-1 levels. Absorbance was measured at 450 nm using a microplate reader to create a standard curve and determine the concentration.
Paraffin-embedded blocks of the renal sections were studied after xylene deparaffinization; the rehydration step of these sections was performed by decreasing the alcohol concentration. Then, blockade of Peroxidase activity was blocked with H2O2, while protein blockers were used for non-specific binding sites, primary antibodies against NF-κB (mouse NF-κB antibody, 1:200) were incubated at 4°C overnight, followed by the addition of a secondary antibody (biotinylated antibody) done at 37°C for 30 min. HRP was added at the same time. Finally, each slide was treated with the chromogen (100μl/slide) for 15 min. All sections were counterstained with hematoxylin. Positive immunostaining was visualized as brown granules contained in the cytoplasm.35
Total RNA was extracted using TRIzol reagent, according to the manufacturer’s instructions. We used an ABI 7500 real-time PCR machine and the Power SYBR Green PCR master mix to run real-time PCR in triplicate on cDNA produced by a reverse transcription reaction. Table 1 displays the primer sequences used, with GAPDH as the internal control. The 2^-∆∆Ct technique was used to determine the relative expression levels of target genes.
This study utilized version 9.3.1 of GraphPad Prism for statistical analysis https://www.graphpad.com/demos/. One-way analysis of variance (ANOVA) was used to examine differences across groups. Subsequently, the Bonferroni method for multiple comparisons was used to conduct post-hoc tests. All tests were deemed statistically significant when P was less than 0.05. All data are presented as the mean ± SEM.
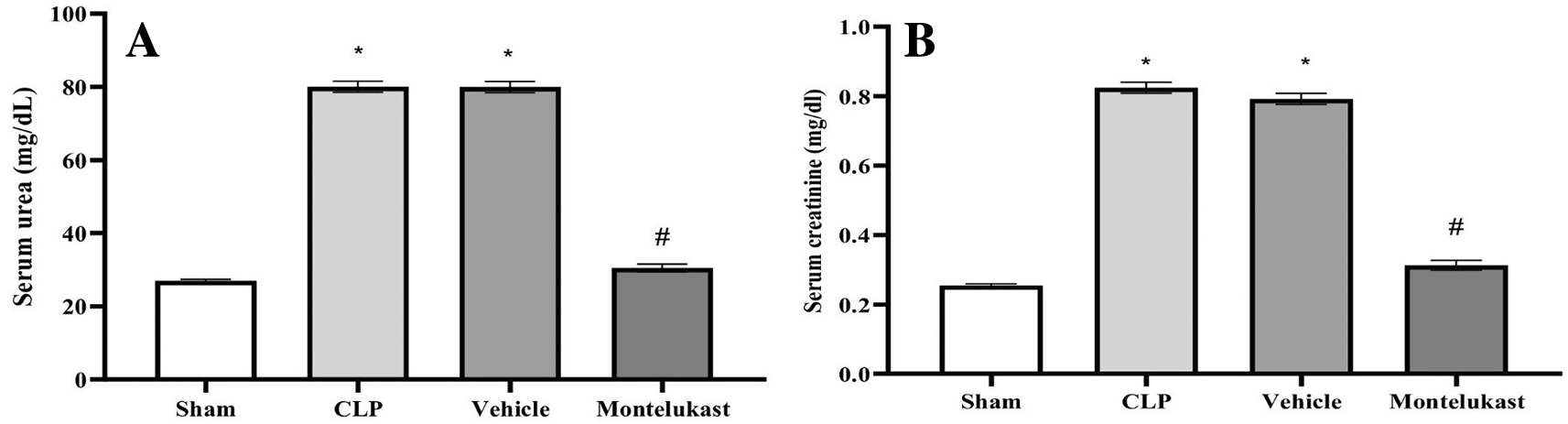
The results indicated that the CLP and vehicle cohorts had remarkably elevated serum BUN ( Figure 1A) and Cr ( Figure 1B) levels, in contrast to the sham cohort. Additionally, the BUN and Cr levels in the montelukast group were significantly lower than those in the CLP or vehicle cohorts.
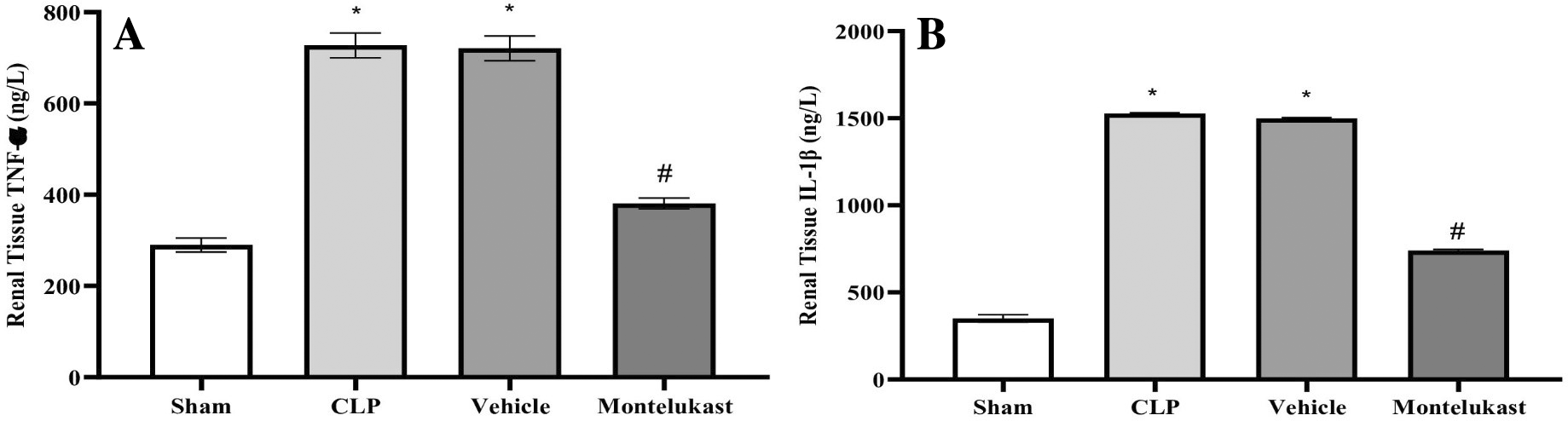
According to these findings, the CLP and vehicle cohorts exhibited a notably heightened level of TNF-α ( Figure 2A) and IL-1β ( Figure 2B), which was in stark contrast to the sham cohort. Furthermore, TNF-α plus IL-1β concentrations in the montelukast cohort were significantly lower than those in the CLP or vehicle cohorts.
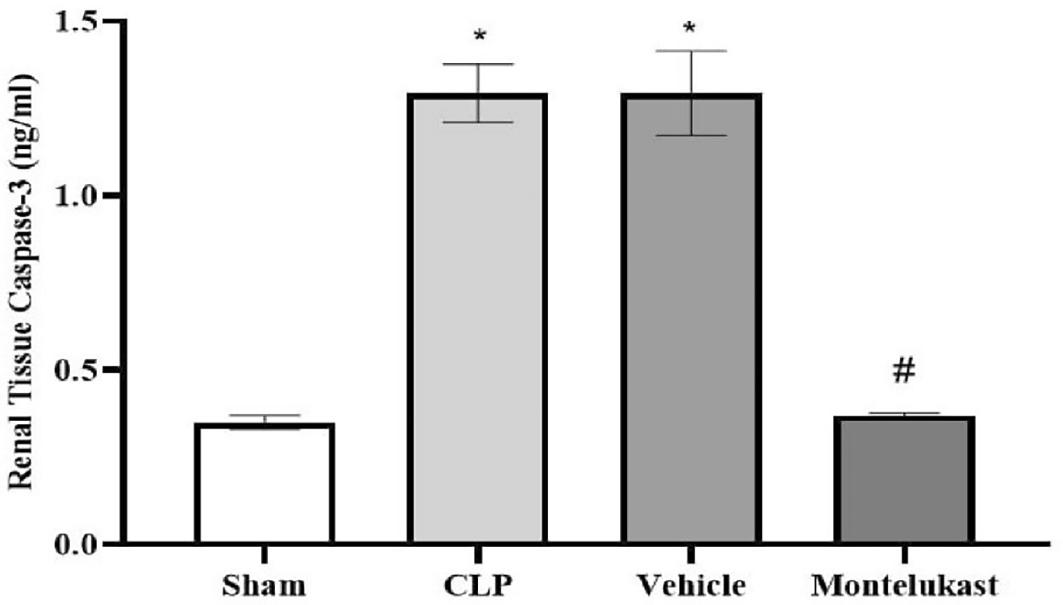
The results indicated that caspase-3 tissue levels in the CLP and vehicle cohorts were remarkably higher than those in the sham cohort. Additionally, the montelukast group had significantly lower levels of caspase-3 than the CLP or vehicle groups, Figure 3.
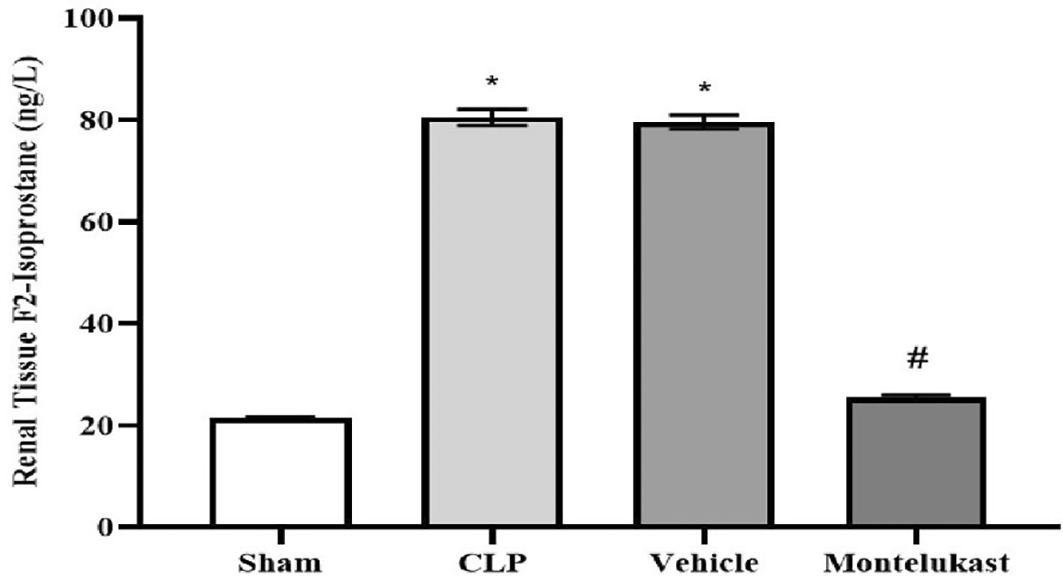
The results indicated that CLP and vehicle cohorts had a significantly higher concentration of F2-isoprostanes than the sham cohort. Additionally, the cohort administered montelukast exhibited significantly reduced concentrations compared to the CLP or vehicle cohorts, Figure 4.
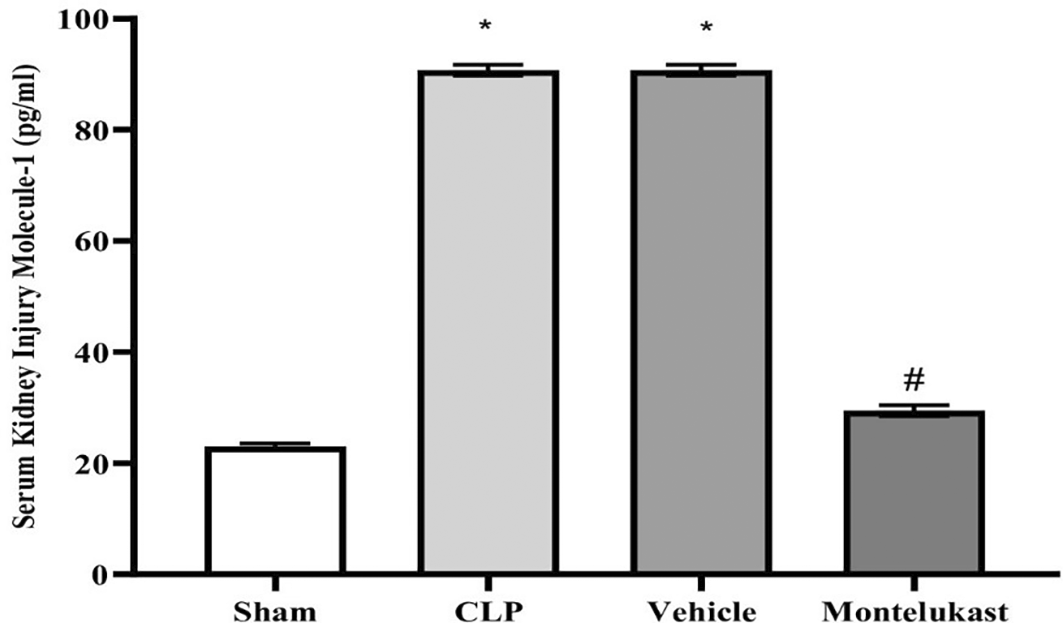
The results indicated that the CLP and vehicle cohorts had remarkably higher serum KIM-1 levels than the sham cohort. Additionally, the KIM-1 levels in the montelukast group were significantly lower than those in the CLP or vehicle groups, Figure 5.
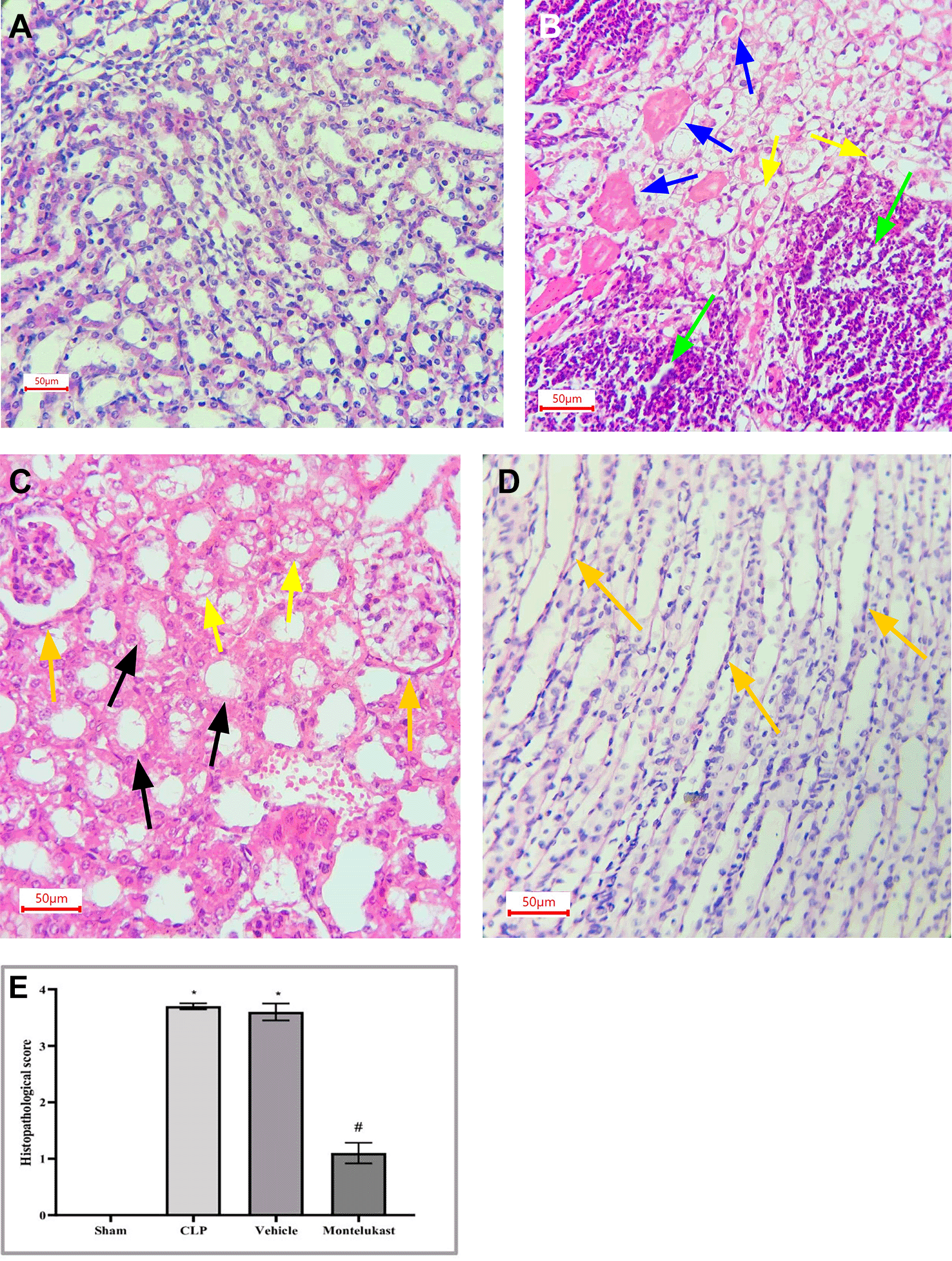
As illustrated in Figure 6, significant pathological alterations were observed in both CLP and vehicle cohorts. Nevertheless, renal injury induced by CLP were markedly ameliorated by pretreatment with montelukast.
As shown in Figure 7, the CLP and vehicle groups had a lower ΔCT than the sham group, indicating a significant increase in the NF-κB p65 mRNA expression. Additionally, there was a substantial ΔCT surge in the montelukast group compared to the CLP or vehicle groups, representing a decrease in NF-κB p65 mRNA expression.
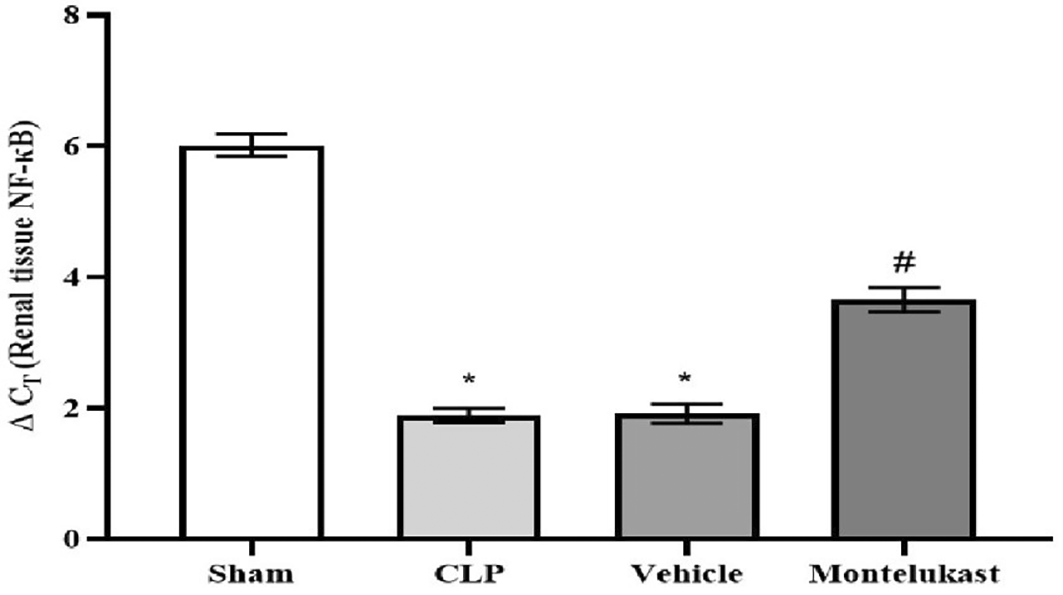
*p<0.001, vs. Sham group; #p<0.001, vs. CLP or vehicle group, CLP: Cecal ligation & puncture; ΔCT: delta cycle threshold; NF-κB: nuclear factor kappa B.
Figure 8 illustrates the intensity of positive NF-κB p 65 staining in renal tissue, in addition to negative staining, as follows:
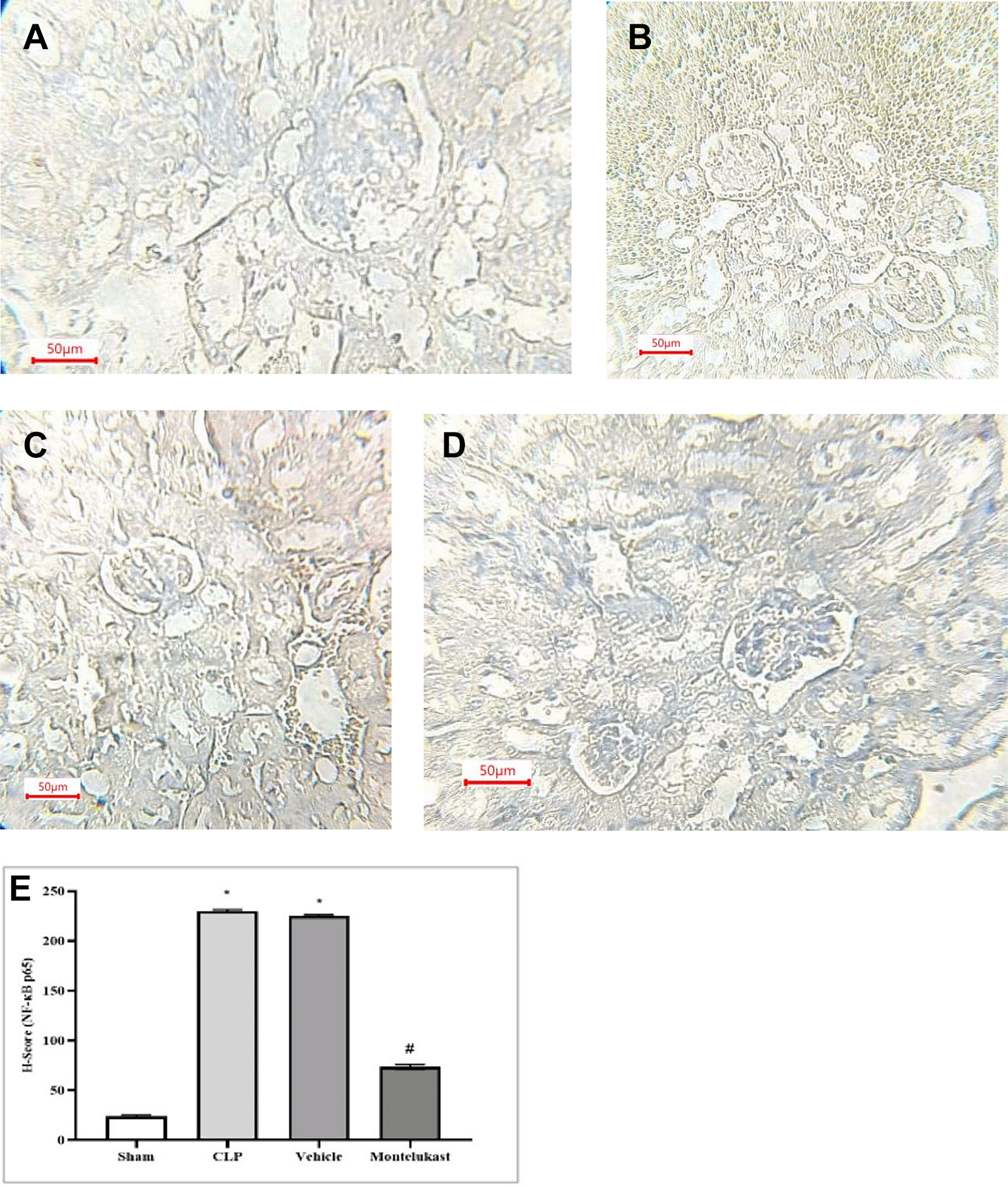
Despite the widespread recognition of the clinical importance of SA-AKI, its pathophysiological mechanisms remain controversial, and there is a lack of unified diagnostic and therapeutic standards.36 Sepsis is characterized not only by hyperinflammation but also by a complex immunosuppressive state. Several experimental and clinical investigations have shown that the immunosuppressive state induced by sepsis is typified by decreased antimicrobial effector functionalities, thereby increasing vulnerability to infections.37 Immunosuppression associated with sepsis is multifaceted and is believed to arise from compromised cytokine production and reduction in the phagocytic capabilities of myeloid cells.38 The present investigation revealed that the tissue concentration of the pro-inflammatory cytokine TNF-α was markedly elevated in the CLP cohort compared to that in the ostensibly healthy cohort. This investigation corroborates previous findings,39 which demonstrated that in sepsis models, the concentrations of proinflammatory cytokines, particularly TNF-α, were elevated in CLP mice and that increased concentrations of TNF-α correlated with mortality outcomes. In addition, this result was consistent with a previous study40 showing that renal ischemia-reperfusion injury (RIRI) is a major cause of AKI, characterized by significant inflammation that exacerbates tissue damage. Additionally, the level of tissue TNF-α was significantly lower in the montelukast group than that in the CLP group. Our results were confirmed by an investigation conducted by Khodir et al., which provided evidence suggesting that montelukast may confer a protective effect on the heart in the context of LPS-induced cardiac injury in rat models.41 These findings imply that the anti-inflammatory effects of montelukast may be ascribed to the attenuation of pro-inflammatory mediators.42 Furthermore, our study revealed a notable increase in the tissue levels of IL-1β in the CLP cohort compared to the sham group. This work concurs with a prior study conducted by Ibadi et al. in exploring the lung damage induced by CLP, and found that the CLP procedure caused a significant surge in pro-inflammatory cytokine levels, IL-1β.43 Another study investigated the effects of resveratrol on IRI in rats. They found that the level of IL-1β became altered (increased significantly) in ischemic rats.44 On the other hand, the present study, regarding the influence of montelukast on the concentration of IL-1β, demonstrates that the levels of tissue IL-1β were considerably diminished in the montelukast cohort, in contrast to CLP. Our study is in agreement with those visualized by Ref. 42 and found that compared to the LPS cohort, management with montelukast lessened the surge in serum IL-1β concentration. In addition, the current study found that the CLP and vehicle groups had significantly higher tissue levels of caspase-3 than the sham group. Similar results showed that the sepsis group had higher caspase-3 levels than the normal physiological state.45 Moreover, this work concurs with a prior study that examined renal damage induced by RIRI. They found that RIRI caused a significant surge in the kidney marker of apoptosis, caspase-3, compared to the sham group.46,47 Additionally, in the present study, concerning the effect of montelukast on the level of caspase-3, the concentration of renal caspase-3 was remarkably lowered within the montelukast cohort, contrary to the CLP group. To the best of our knowledge, no prior study has investigated the pharmacological effects of montelukast on caspase-3 during sepsis. This finding could be ascribed to the ability of montelukast to reduce the levels of NO and renal HO-1. Montelukast effectively mitigated elevated concentrations of ET-1, MCP-1, and TNF-α. These effects were associated with a reduction in caspase-3 expression in the kidneys, confirming its anti-apoptotic activity.48 Moreover, the current investigation found that the CLP and vehicle cohorts had considerably higher kidney tissue concentrations of F2-isoprostane than the sham cohort. This study supports another work that established a notably higher concentration of the oxidative stress marker (F2-Isoprostane) within the sepsis cohort than in the sham cohort.49 In the present study, regarding the impact of montelukast on F2-isoprostane concentration, it was observed that montelukast markedly reduced the concentration of F2-isoprostane within kidney tissue compared to the CLP group. Additionally, our work is compatible with,50 which examined whether montelukast possesses the potential to conserve lung function during instances of polymicrobial sepsis. The results of this study showed that mice subjected to sepsis exhibited a notable increase in the level of F2-isoprostane within lung tissues, in stark contrast to the sham cohort. Compared to the sepsis cohort, the administration of montelukast resulted in a significant reduction in F2-isoprostane levels in the lung tissue. This finding can be attributed to the montelukast inhibitory effect of the NF-κB/NLRP3 pathway in which NLRP3 can induce reactive oxygen species production.51 Furthermore, the current study showed that serum KIM-1 was remarkably higher in the CLP cohort than in the sham cohort. This work agrees with a previous study that confirmed that the levels of KIM-1 in Lyn mice that underwent sepsis were remarkably higher than those in sham mice.52 In addition, the current study aligns with another research effort that established the upregulation of inflammatory marker KIM-1 in both the RIRI and vehicle groups when compared to the sham group.46,47 In the present investigation, concerning the impact of montelukast on the concentration of KIM-1, it was observed that montelukast markedly reduced the concentration of KIM-1 in kidney tissue compared to the CLP group. To the best of our knowledge, this investigation represents an inaugural work that elucidates the influence of this pharmacological agent on renal KIM-1 level within the context of the CLP model of sepsis in mice. This observation may be attributed to the ability of montelukast to inhibit the ERK signaling pathway.53 Collier and Schnellmann posited that inhibition of ERK1/2 prevents renal sepsis-induced STAT3 phosphorylation, thereby attenuating KIM-1 level through the ERK1/2–STAT3–KIM-1 signaling pathway following renal injury.54 In addition, the present investigation revealed that the mRNA and protein expression of NF-κB p65 were notably elevated in the CLP cohort compared to the sham cohort. This work corroborates findings from a previous study, which indicated that the phosphorylation level of p65 within the renal tissue of the CLP cohort was significantly increased when juxtaposed with the sham cohort.55 In the present study, concerning the impact of montelukast on the expression of NF-κB p65, there was a notable reduction in NF-κB p65 expression within the renal tissue relative to the CLP cohort. This study is the first to clarify the effect of this pharmacological agent on renal NF-κB p65 expression in a murine model of CLP-induced sepsis. This result may be attributed to the ability of montelukast to mitigate IL-1β-induced NF-κB p65 phosphorylation, thereby inhibiting nuclear translocation and subsequent NF-κB activity related to gene expression.56 Moreover, the CLP and vehicle groups showed remarkable histopathological changes compared to the sham group. The renal tissue of the sham group mice had normal architecture, whereas the kidneys obtained from the mice in the CLP cohort exhibited signs of hemorrhage, severe inflammation, increased cytoplasmic eosinophilia, eosinophilic casts, and cytoplasmic vacuoles, as well as loss of brush border. These observations are consistent with those obtained by57 in their study on groups of CLP mice. They reported that CLP causes inflammation, necrosis, hemorrhage, and degeneration of kidney tissue. Additionally, the Montelukast group showed a significantly lower level of renal tissue injury than did the sepsis group. Our results agree with Khodir and colleagues’ study on rats to examine the renoprotective effects of Montelukast after CLP.58 This finding can be attributed to their antioxidant and anti-inflammatory properties. It is important to note that the potential influence of the vehicle (DMSO) was carefully controlled in this study. The vehicle group, which received 10% DMSO in saline via intraperitoneal injection under identical experimental conditions, did not show significant changes in inflammatory, oxidative, or apoptotic markers compared with the sepsis control group. This finding suggests that DMSO at this concentration did not exert a measurable biological effect under the present experimental conditions. Thus, the protective effects observed in this study are attributed to montelukast rather than the vehicle.
The use of a montelukast pretreatment protocol and post-treatment effects were not evaluated in the present study. Future studies are warranted to investigate the therapeutic (post-treatment) effects of this pharmacological agent to better reflect clinical scenarios.
Montelukast demonstrated significant renoprotective effects in septic mice with AKI by attenuating the release of inflammatory mediators through inhibition of NF-κB P65 expression. Additionally, it reduced oxidative stress, thereby improving renal outcomes. Furthermore, it mitigated the adverse consequences associated with apoptosis following renal injury by modulating key apoptotic factors.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)