Keywords
ceftriaxone; adaptative humoral immune response; Artemis; Tetanus;Toxin vaccination
ceftriaxone; adaptative humoral immune response; Artemis; Tetanus;Toxin vaccination
The β-lactam antibiotics act by inhibiting the synthesis of the peptidoglycan layer of bacterial cell walls. However, the increasing resistance to these drugs especially among Gram-negative bacteria has been a concern for public health worldwide and challenged the manufacturer for generation of news β-Lactam antibiotics (e.g., penicillins, cephamycins, carbapenems, monobactams and cephalosporins).1–3 Each new antibiotic was developed to increase the number of bacterial species covered by antimicrobial activity and to bypass resistance mechanisms. The main resistance mechanism is β-lactamase enzymes, divided into four classes, active site serine β-lactamases (classes A, C and D) and metallo-β-lactamases (class B). For example, Ceftriaxone, a β-lactam antibiotic of the third generation cephalosporins, known to be a competitive inhibitor of Artemis can act as an inhibitor on most of the β lactamases which are enzymes exhibiting different activities.4 Also, we know that currently five generations of cephalosporins (e.g., cefotaxime, ceftriaxone, ceftazidime …), have been made available to infectiologists and that Ceftriaxone is one of the most frequently used antibiotics in clinical practice to treat lung and urinary tract infections with pathogenic bacteria.3,5–7
It has been demonstrated that Ceftriaxone is a competitive inhibitor of exonucleases of the human metallo-β-lactamase super family and that there are 18 metallo β lactamase-encoding genes in the human genome.8,9
Artemis, a protein which belongs to this family and extensively expressed in cells, is involved in DNA repair and in immune response development.10 It seems that Artemis doesn’t have any β-lactam degradation activity despite the fact that there is a common catalytic site with other metallo-β-lactamases and that Ceftriaxone is blocking the catalytic activity in vitro (Data not shown). Artemis is also an endonuclease playing a key role in development of B and T lymphocytes during the early stage of maturation in their G1 phase cell cycle.11 It’s also a protein phosphorylated by DNA-dependent protein kinase catalytic subunit (DNA-PKcs) and acting to open DNA hairpin intermediates generated during V(D) J and class switch recombination.12,13 On the immune level, the strength of this response in mammals is based on a process of combinatorial assembly of the variable parts of the antigen receptors. More particularly by the rearrangement of the exons of the variable parts of the heavy chains of immunoglobulins and of the receptors on the variable (V), divergent (D) and junction (J) fragments by a mechanism of “cut and paste” of the DNA. This process, called V(D) J recombination first assembles two segments, a D to a J then a V segment joins the rearranged DJ segment.14 These unique, specifically generated antigen receptors can thus recognize a very wide range of molecules. In some exceptionally rare radiation sensitive patients, the gene encoding the Artemis protein has been found to be defective. The defective Artemis protein can no longer cleave the ends of the hairpin, causing the blockage of V(D) J recombination leading to a reduction or even loss of maturation of B and T cells.15,16 This phenomenon is known under the name Severe Combined Immunodeficiency (SCID) in humans and mice leading to transient impaired immune response to pathogens.11,17,18
Recently, Chapman and colleagues reported a retrospective analysis of a cohort of children aged 6 to 24 months regarding antibiotic prescriptions and vaccine antibody measurements relative to diphtheria-tetanus-acellular pertussis, inactivated polio, haemophilus influenzae type b, and pneumococcal conjugate vaccines.19 They concluded that, for several vaccines, antibiotic use in children below 2 years of age is associated with lower vaccine-induced antibody levels. The normalized antibody level was significantly reduced according to the number of antibiotic prescriptions (including Ceftriaxone) compared to children with no antibiotic prescriptions (control) and was more pronounced when the antibiotic was injected before vaccination.
According to our previously published hypothesis and the observation of the decrease in vaccine-induced antibodies in children receiving β-lactam antibiotics such as ceftriaxone, this molecule could inhibit the exonuclease activity of Artemis, leading to transient immunodeficiency.9,19 Given the recognized affinity of beta-lactam antibiotics with beta-lactamases, we hypothesize that the Artemis protein would be inhibited transiently upon contact with an antibiotic of this particularly family. Thus, the maturation of B lymphocytes may be inhibited, slowed or delayed until the effects of Ceftriaxone wear off. We can speculate that the Artemis protein is unable to act to open the DNA hairpin intermediates during V(D) J segment rearrangements as suggested in schema of Figure 1. To address this issue, we used a mouse model to study the impact of Ceftriaxone treatment on the immune response to tetanus toxin.
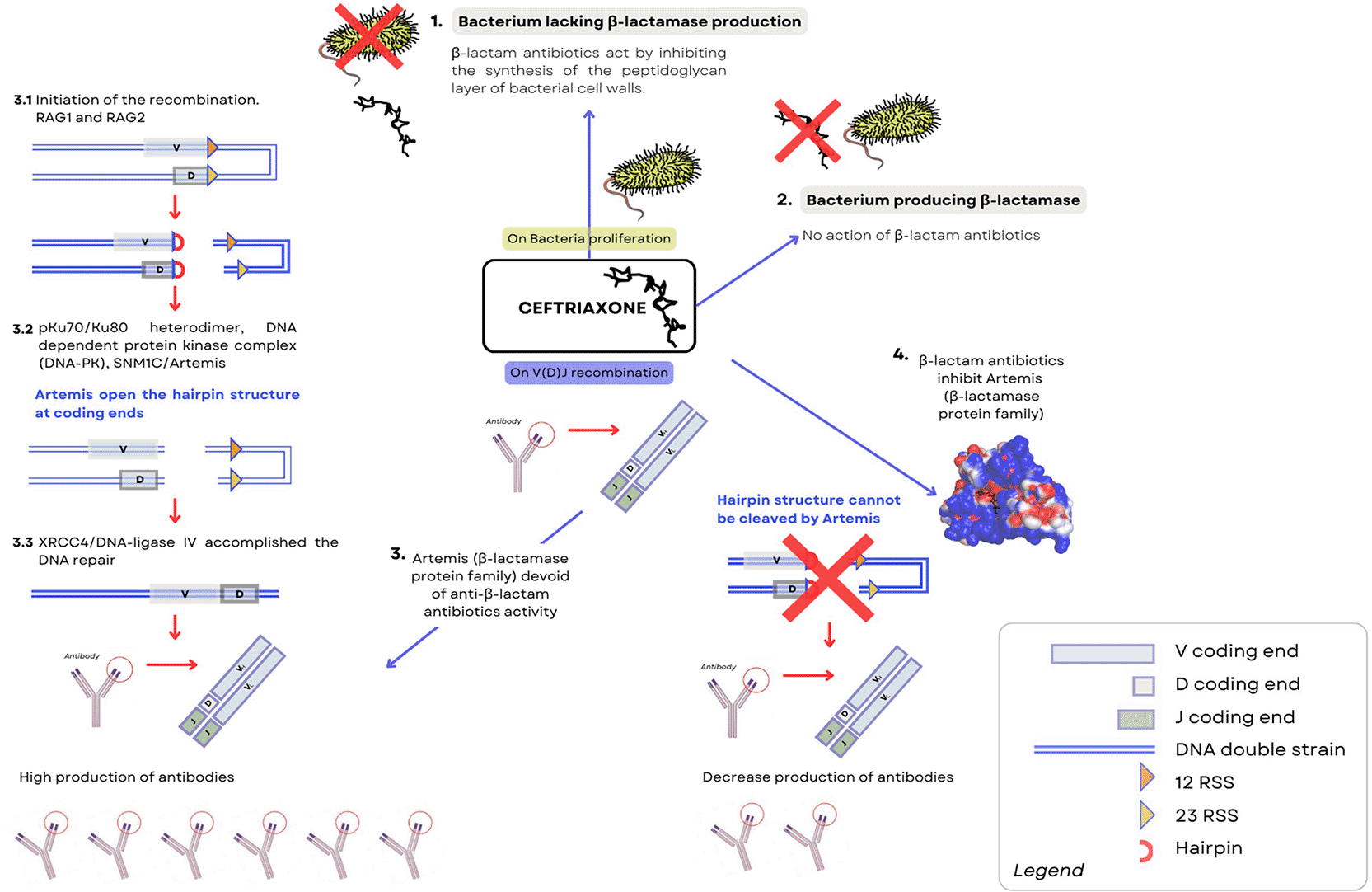
A. Hypothesis of the effect of Ceftriaxone on the proliferation of Bacteria (1 & 2).
1. If some bacteria cannot produce beta-lactamases, beta-lactam antibiotics (such as Ceftriaxone, CFTX) can inhibit the synthesis of the peptidoglycan layer of the cell walls of these bacteria. These bacteria are called “sensitive to beta-lactam antibiotics”.
2. On the other hand, if bacteria capable of producing beta-lactamases hydrolyze beta-lactam antibiotics, then these bacteria become “resistant to antibiotics from the beta-lactam family”, and therefore to Ceftriaxone.
B. Hypothesis on recombination of the V(D) J segment (3 & 4).
3. Under normal conditions during an immune response, Artemis, a protein from the beta-lactamase family, can initiate the recombination cascade of V(D) J fragments by cleaving the hairpin structure. In this case, a high production of antibody response can be observed.
4. On the contrary, if Ceftriaxone can bind and block the activity of the SNM1C/Artemis protein as we hypothesized in C. Melenotte et al.,9 then the hairpin structure could not be cleaved and a reduction and/or delay in antibody production could be observed.
We show that the humoral response to tetanus toxin is impaired by Ceftriaxone treatment.
The experimental procedures have been validated by the Institutional Committee for the Protection and Use of Animals CEEA-14 of the University of Aix-Marseille, France and approved by the French ministry of research under file reference 201905211839232/APAFIS Number 20772. The mice were housed in specific pathogen-free conditions in group cages while respecting the number of animals per cage in the European Union EU0210 - MEPHI RITMES IHU laboratory - under approval number E1305509 after an acclimatization period of at least 7 days to avoid animal stress. Igloo-type enrichment and nesting material have been added to each cage. Daily monitoring and control of the animals was carried out by the experimenter even on public holidays. To minimize confounding factors, each cage and each mouse is processed in the same order at each stage of substance administration or blood sampling. Each cage is then returned to its original location. We used 77 BALB/cByJ male mice aged 6 to 8 weeks supplied by the Charles River Laboratory (BP 0109 L’Arbresle, 69592 Lyon, France). The health certificate for the animals complied with the specifications for the animals ordered. The experimenter, the designer, and the zoological technician were aware of the distribution of the groups at the different stages of the experiment.
Eighteen to twenty mice per group were treated with a 100 microliters intraperitoneal injection of Ceftriaxone (1g, powder for injection, VIATRIS, 1 rue de Turin, 69007 Lyon, France under reference J01DD04) at a concentration of 50 mg/kg or with 0.9% Sodium Chloride (NaCl), an isotonic saline solution for injection daily for 5 days. Ceftriaxone was prepared according to the manufacturer’s instructions at the concentration indicated above in 0.9% NaCl (From AGUETTANT, Quai Louis Aulagne, 69190 Saint-Fons, France, Code ATC B05XA03) and based on results described by Majhi et al.20
Mice received a single dose of DTP (diphtheria-tetanus-polio) vaccine purchased from the Sanofi-Winthrop manufacturer (product REVAXIS, ATC code J07CA01, Sanofi Winthrop Industrie, 83 avenue Raspail, 94250 Chantilly, France; batches R3J398V, R3J391V, T3C215V). Two different vaccine administration protocols (Conditions A or B, see below) were studied and for each condition, mice were divided into groups receiving either the antibiotics ceftriaxone or a solution of isotonic saline (0.9% NaCl) as a control. Condition A: immunization before the start of antibiotics (or control) treatment (Groups 1C & 1D/ 20 mice/ Ceftriaxone and Groups 1A & 1B/ 19 mice/ 0.9% NaCl – controls), and Condition B: immunization after the end of antibiotics (or control) treatment (Groups 2G & 2H/20 mice/Ceftriaxone and Groups 2E & 2F/18 mice/0.9% NaCl – controls), as illustrated in Figure 2. Each group (under each condition) was compared: 1- to other groups in the same condition, 2-to groups in the other condition and 3- to placebo (NaCl) control groups. We excluded older mice to retain only the groups of animals exhibiting a strong immune response. We considered all animals from all groups (insufficient sample size for testing). The sole reason for the reduction in sample size (n) stems from the number of samples where the quantities were insufficient, as shown in particular by the decrease in n (number of mice) in Figures 3 and 4.
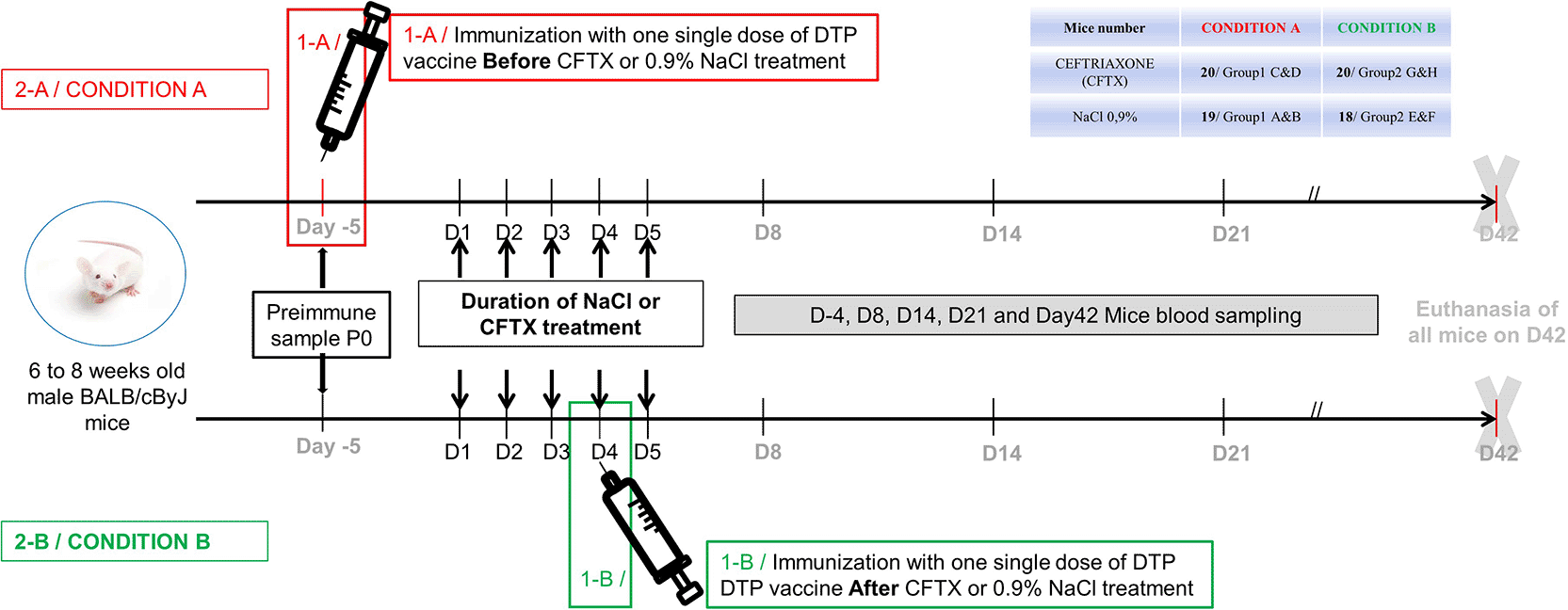
20 male BALB/cByJ mice were immunized by intraperitoneal injection of DTP Vaccine four days before antibiotic treatment during 5 days (Red/ 1-A/Condition A). 20 male BALB/cByJ mice were immunized by intraperitoneal injection of DTP Vaccine four days after beginning of antibiotic (Green/1-B Condition B) treatment during 5 days. Blood from mice was collected at Day-5 for preimmune control, and at Day8, D14, D21and D42, final blood collection.
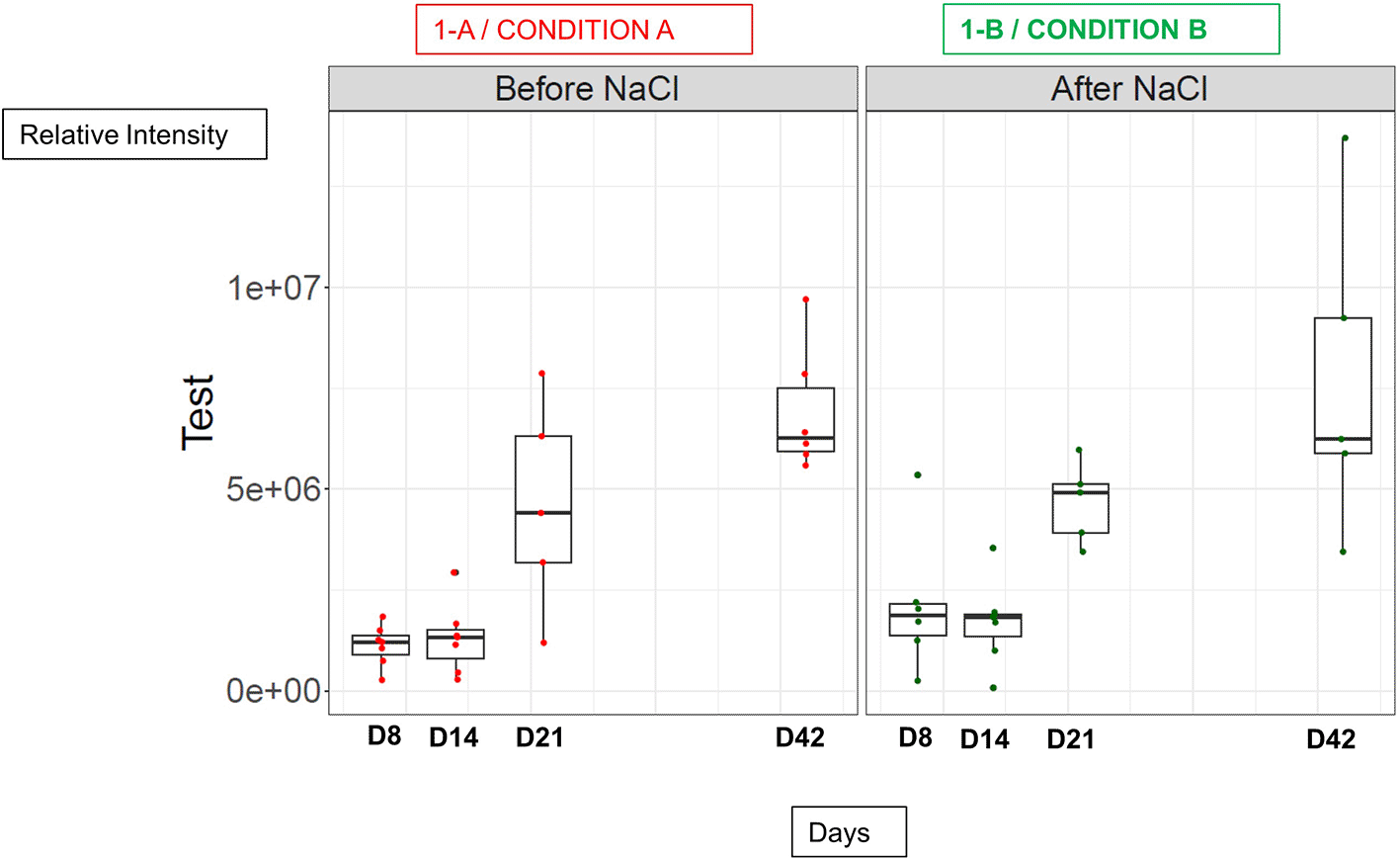
A- DTP Immunization before 0.9% NaCl treatment (As Condition A), B- DTP Immunization after 0.9% NaCl treatment (As Condition B). Between 5 to 7 serum mice per day on each treated group was quantified as described in M&M. The median of each mice group by days (D8, D14, D21 and D42) was calculated and plot of each serum value is represented. *** p value= 1.03e-05.
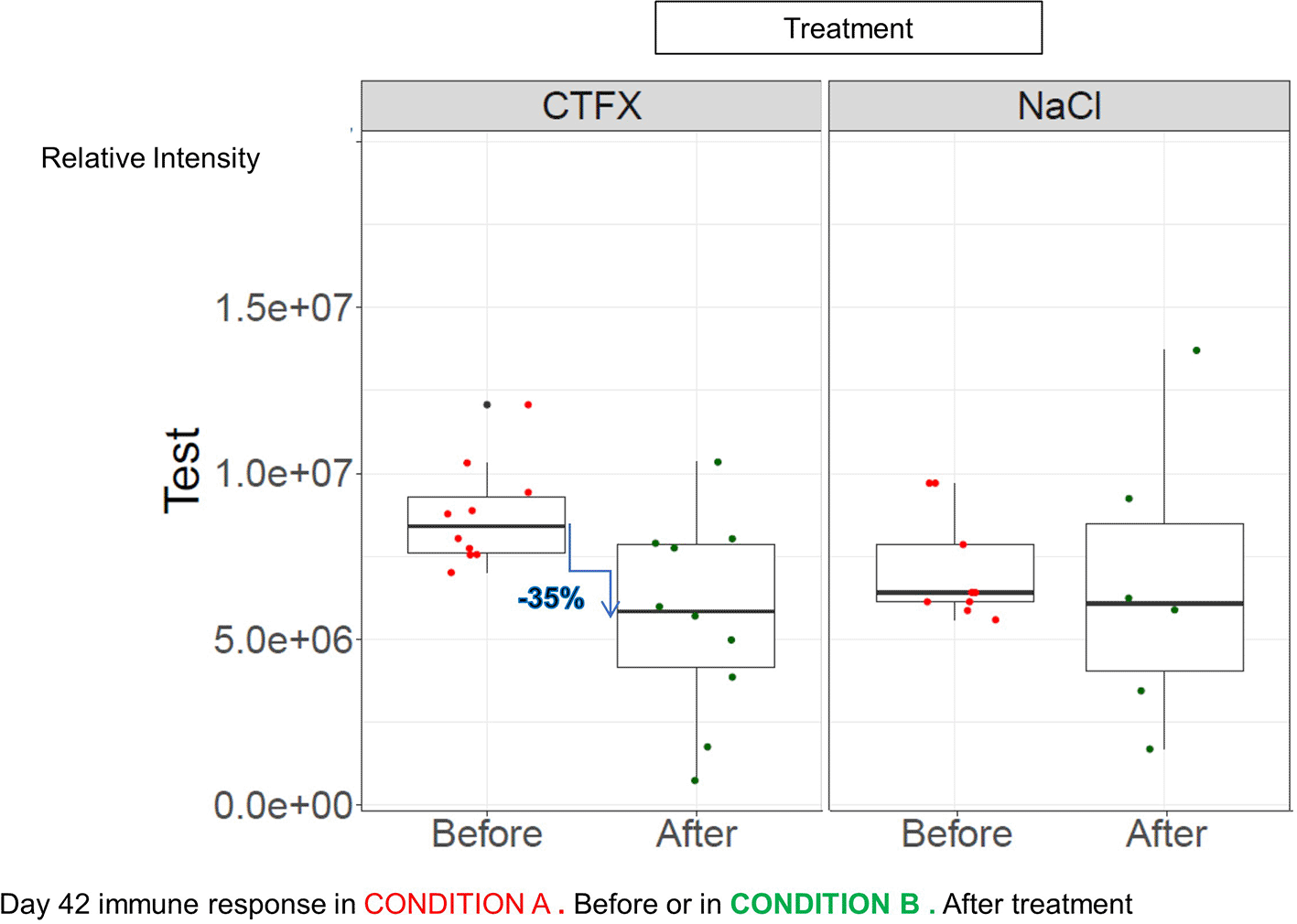
Mice were vaccinated with DTP vaccine before or after Ceftriaxone (CFTX) treatment (n = 10) versus 0.9%NaCl as “control”/NaCl (n = 9 and 6). An Anova test give us a significant effect between both treatment (p = 1.31e-10). Condition A or B gave us also a significant p value (p = 0.00167).
We selected the commercial three-component DTP vaccine because it is a vaccine commonly used in humans and tested on mice before obtaining marketing authorization.21 The dose of vaccine used was 150 microliters by intraperitoneal injection (with a 25G needle of 16 mm length), i.e. a concentration of 6 IU of TT/mouse. This dose was calculated based on data from Huh DH, et al. and the dose tests carried out in our laboratory to have a sufficiently detectable signal with the Immunochromatographic test we chose after a single injection of antigens (Data not shown).22 The active substances in each 0.5 ml dose of vaccine are: - Purified diphtheria toxoid not less than 2 IU* - Purified tetanus toxoid (TT) not less than 20 IU* - Inactivated poliomyelitis virus (produced in Vero cells) --Type 1: 40 antigen D units** --Type 2: 8 antigen D units** --Type 3: 32 antigen D units** * IU is an international unit for measuring vaccine potency ** the antigen unit expresses the amount of antigen present in the vaccine dose.
Intermediate and pre-immune blood samples were taken using a submandibular lancet. A final blood sample was taken intracardially after anesthesia and euthanasia of each mouse. Each blood sample was centrifuged at 800 rpm at 4°C during 15 minutes and sera samples were then stored at -30°C until used for analysis.
As described in the May 2019 Project Authorization Application (PAA), general anesthesia is achieved by intraperitoneal (IP) injection of a mixture of ketamine (90 mg/kg) and xylazine (10 mg/kg) in mice. Ketamine was marketed under the name Imalgene 1000 by VIRBAC (13 rue LID, 05617 Carros, France) and xylazine under the name ROMPUN 2% by BAYER HealthCare SAS - Animal Health (1 rue Claude Bernard, 59000 Lille, France). The mice were then euthanized by IP injection of pentobarbital sodium (CEVA Santé Animale, 8 rue de Logroño, 33500 Libourne, France) or DOLETHAL containing pentobarbital (100 mg/kg) (VETOQUINOL, 10 rue du Docteur Deubel, 70200 Lure).
We particularly followed the anti-tetanus response because it was demonstrated that in male mice of different age (the youngest and the oldest mice only), the cellular and humoral immune responses are not altered unlike those against diphtheria toxin.23 Tetanus antitoxin sera were analyzed by Immunochromatographic test with the “TQS TETANOS QUICK STICK” kit, referenced # TE-20B/S-GZ-20B from Eurobio Scientific (7 av. De Scandinavie, ZA de Courtaboeuf, 91940 Les Ulis, France). Following the manufacturer’s procedure, twenty microliters of each serum were dispensed in the well of the stick and three drops of the diluent provided by the kit was immediately added to the sample. The results are read after no ten minutes by scanning the stick using the “Image Scanner 3” (GE Healthcare, Buc, Yvelines, France). Scanned sticks were formatted for alignment by mouse batch and treatment group using Adobe Photoshop CS3 software. Quantification of each stick was calculated by measuring band intensities using the “Image Quant TL Version 7.0 analysis software (GE Healthcare, Buc, Yvelines, France). In Figures 2 and 3, the values on the y axis represent the relative intensity of the band (area) of the pink T Test line quantified by the software.
The statistical analysis and Figures 3 and 4 result from the free R software used for statistical calculation and graphics. An Anova test was used to calculate p values for the analysis in Figure 3 to find out if there is a significant difference between the two conditions (before or after isotonic saline treatment) and a significance difference of time for the two conditions. For Figure 4, an Anova test gave a significant result for the condition effect and time, so a non-parametric Wilcoxon test was used to calculate p-values to confirm the significant effect.
Our goal was to evaluate whether Ceftriaxone alters DTP humoral immune response.
For this purpose, we decided to use an animal model to experimentally follow the antibody response against Tetanus Toxin in groups of DTP immunized mice before (CONDITION A), after (CONDITION B) antibiotics treatment or not submitted to any antibiotic treatment as a control group as shown in Figure 2.
Thus, we chose to examine the Tetanus Toxin immune response after DTP vaccine administration in mice treated with Ceftriaxone (n = 40) versus those treated with isotonic saline solution as control (n = 37). A group of mice was immunized before the start of Ceftriaxone treatment or with isotonic saline solution (n = 20/19, respectively). ( Figure 2A), and two groups were immunized after treatment with Ceftriaxone or isotonic saline solution (n = 20/18, respectively) ( Figure 2B). We collected the pre-immune blood samples five days before the start of the Ceftriaxone or saline treatment (Day-5) for each mouse. During the experiment, we then took blood from the mice at Day8, D14, D21 and D42 (the final blood collection).
We monitored the anti-tetanus response with the Tetanus Quick Sticks kit used in the context of a vaccination schedule or a suspicion of Clostridium tetani infection diagnosis in our laboratory. The advantage of this technique is that it is easy to use, consumes very little serum (or whole blood, 20 microliters) and that results are obtained quickly (less than 10 minutes). This kit is based on immunochromatography assay principle. A mixture of gold conjugates coupled with tetanus toxoid and BSA is fixed on the solid phase of the stick. The mouse serum sample to be tested is placed in the well of the stick, followed by a few drops of diluent. The migration of the diluent onto the stick carries the gold conjugate which forms a complex with the anti-tetanus immunoglobulins present in the sample. If samples are positives, complexes between gold conjugates and anti-tetanus immunoglobulins, stopped on the immobilized toxoid on the stick and appear in the stick window in lane T. The control band, C pink lane in the window, is formed by the capture of the excess of gold conjugates by the control reagent constituted of poly-L-Lysine based buffer ( Figure S1).
Before attempting to answer the question about the potential alteration of the immune response by Ceftriaxone treatment, we tested the efficacy of the immunological responses in mice immunized with the human DTP vaccine. The response dose was established at 6IU per mouse as described in M&M based on D.H. Huh et al. data.22 By immunizing the mice with the DTP vaccine at 6IU, we were able to detect anti-Tetanus Toxin immune response with the specific kit we chose: the object was not to protect them against tetanus but to measure whether they produced antibodies against an antigen under β-lactams family’s antibiotic treatment or not (Data not shown). Then, we followed the kinetics of the immune responses of immunized mice with the DTP vaccine before ( Figure 2A) and after ( Figure 2B) the isotonic saline treatment. This control treatment consisted of a five-day daily intraperitoneal injection of a 0.9% sodium NaCl solution. We observed in scanned sticks ( Figure S1) that, for the eight randomly chosen mice (four for each condition), no band appeared in the pre-immune serum P0 samples in the Test T line compared to the control C line which guaranteed that the analyzed sticks were valid. Then, from day 8 to day 42, we observed a band corresponding to the detection of anti-tetanus toxin antibodies in the serum of the majority of the mice. Notably, the intensity of signal gradually increased over time, particularly from day 14 to day 42 in the test line.
The Figure 3 presents the relative intensity of quantified bands of each stick from immunized mice serum in the two tested condition (A and B). We observed that, as generally observed in literature, the maximum immune response after a natural exposure to antigens or to one dose of vaccine, occurred around 4-6 weeks after injection, as we can see for D42 in our Figure 3.24 There was a significant difference in the immune response over time (from D8 to D42, last day test) similar for the two conditions A and B (p value= 1.03e-05).
We also observed no significant difference in TT-response in both conditions: before (Condition A) or after (condition B) isotonic saline treatment from Day 8 to Day 42.
The date-to-date variability [antibodies were not measured daily] in anti-Tetanus Toxin antibody responses per mouse could be explained by animal-to-animal variability.
Despite the high value at D8 in condition B for two mice, the mean values remained very close to condition A. At Day 8, in condition B, the discrepancy between the minimum and maximum values and the high anti- Tetanus Toxin antibody responses can both be explained by the fact that, in this particular condition, the DTP injection took place just 5 days before the D8 blood collection. Conversely, in condition A, the injection of DTP was carried out twelve days before, which may explain a weaker anti-Tetanus Toxin antibody response.
By observing all the results obtained in Figures 3 and Figure S1, we decided to compare in the rest of the study, the anti-tetanus serological responses at D42 only, where the quantification signal was the highest.
Figure 4 summarizes all immune responses against tetanus under the two conditions in each group of mice for both treatments at D42. An Anova test give us a significant effect between both treatment (p = 1.31e-10). Condition A or B gave us also a significant p value (p = 0.00167). We then calculated p-values with a non-parametric statistic test and we observed a significant drop of 35% in terms of p-values between the mean values of the anti-tetanus immunoglobulins in the sera of the mice immunized after treatment with Ceftriaxone compared with those measured in the mice immunized before the same treatment (p value = 0.02881 for Wilcoxon test). The effect “before or after treatment” during isotonic saline was much less marked than during the Ceftriaxone treatment. In isotonic saline treatment there was no significant difference between the two conditions (p value = 0,5949 for Wilcoxon test).
In this study, we evaluated the effect of the anti-tetanus toxin antibody response in mice exposed to Ceftriaxone, an antibiotic from the beta-lactam family, under two conditions. Either one dose of DTP vaccine carried out before antibiotic treatment, or four days after the start of treatment. As a control, we tested the same conditions (before or after vaccination) under treatment with isotonic saline solution.
We confirmed that the major anti-tetanus immune responses in mice immunized before or after control treatment are maximal at Day 42/5-6 weeks. In our model, the humoral immune response against the Tetanus Toxoid after one immunization dose decreased after a five-day treatment with Ceftriaxone administered intraperitoneally. On the contrary, the humoral response of mice immunized before treatment with Ceftriaxone is not affected by the treatment compared to the humoral responses under isotonic saline solution. We can conclude that ceftriaxone interferes with the humoral immune response to an injection of DTP vaccine in our experimental animal model and these results could therefore confirm our initial hypothesis ( Figure 1).
However, there are other mechanisms of action described for β-lactam and non-β-lactam antibiotic that can influence humoral immune responses reported in the literature.25,26 In addition, it was reported that one β-lactamase inhibitors which is not known to have antibiotic properties has the capacity to inhibit B cells response.11,27 Moreover, the fact that an antibiotic not belonging to the β-lactam family (Erythromycin) ( Figure S2) tend to boost the humoral response instead of inhibits it, suggests that the action of Ceftriaxone on the humoral immune response occurs by acting on Artemis as we had initially postulated.9,25 This hypothesis is strongly consolidated by the fact of a progressive re-increase after a flagrant decrease in the humoral response at the time of serum collection on days 8, 14 and 21 after treatment with Ceftriaxone. We suggested the term reversible response, when the effect of the molecule dissipates, the humoral immune response continues but at a lower threshold.
Indeed, several publications report inhibition of the humoral response (refractory period) by certain antibiotics without clear arguments in favor of an underlying molecular mechanism.25,28 Our results open a new avenue to explore the complex interactions between antibiotic therapy and immune response. By extrapolating these results to human vaccination, the choice of the possible prescription of antibiotics (β-lactam versus non β-lactam) if the subject treated (particularly in early childhood must receive a vaccine injection during this period must be taken into account).19
Conceptualization: DR, Animal experimentation: MM. Bioinformatic and statistical analyses: MM. Data interpretation: MM, DR, and PP. Writing and original manuscript preparation: MM, CD and PP Review and editing: MM, DR, and PP Final manuscript validation: CD, and DR. Supervision: DR. All authors discussed the results and commented on the manuscript. All authors contributed to the article and approved the submission version.
No human samples or clinical data were used in this study. This study did not require ethical approval.
The animal experimentation project has been reviewed and approved by the Ministry of Higher Education and Research under the reference number APAFIS (Project authorization for the use of animals for scientific purposes) #20772. All animals handling and procedures were performed in accordance with the French Decree N° 2013–118, 7 February 2013 and with European law and in agreement with animal research.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)