Keywords
Chromobacterium violaceum; Poultry infection; Bacterial isolation; PCR identification; Mice model; Pathogenicity; Immunological response; Zoonotic potential
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Chromobacterium violaceum; Poultry infection; Bacterial isolation; PCR identification; Mice model; Pathogenicity; Immunological response; Zoonotic potential
Chromobacterium violaceum is a beta-proteobacterium that presents smooth-surfaced violet-colored colonies. It is a non-sporing bacillus and a facultative anaerobe (Kumar, 2012). It is motile because of a single centrally placed polar flagellum, one to four lateral flagella, and surface-associated pili (Ravichandran et al., 2018). C. violaceum is oxidase-positive and catalase-positive (Antony et al., 2013). It is predominantly prevalent, especially in tropical and subtropical areas (Batista and Da Silva, 2017). C. violaceum infections in humans are primarily linked with cases that involve patients with weakened immune systems. This is a normal bacterium found in water and soil. However, the most common entry into the bloodstream and subsequent systemic infections it induces occur through weakened skin, e.g., wounds or cuts, through which the bacterium can enter from contaminated surfaces or water reservoirs (Alisjahbana et al., 2021). There have been some case reports of complications due to C. violaceum, including bacterial hemophagocytic syndrome, brain abscesses, conjunctivitis, chronic granulomatosis, diarrhea, endocarditis, meningitis, neutropenic sepsis, osteomyelitis, pneumonia, puerperal sepsis, and urinary tract infections (Lin et al., 2016). It infects various species in mammals, including pigs, sheep, dogs, buffaloes, and monkeys (Liu et al., 2011).
Innate immunity is activated as a reaction to infection, developing granulomas in the liver. Granulomas consist of clusters of immune cells that the immune system has brought in to “wall off” the infection. The chemokine expression pattern of the granulomas is significant in the organization of the immune response (Amason et al., 2024). Neutrophils are considered one of the first lines of defense against C. violaceum infection. However, neutrophils may be insufficient to eliminate the pathogen in isolation. Repairing the infection will require a stronger immune response (Harvest et al., 2023). Granulomas are structured complexes by which immune cells (macrophages, lymphocytes) are walled off in reaction to a persistent pathogen such as C. violaceum. In contrast to humoral immunity employed in pathogen neutralization, cellular immunity is a host defense barrier for infection control and containment (Harvest et al., 2023). The bacteria also produce violacein, a purple antimicrobial pigment. There is also evidence to support the possibility that violacein can modulate host immune response to infection because violacein has immunostimulatory activity based on the stimulation of immune and inflammatory response pathways (Venegas et al., 2019; Lafta & Sadeq, 2024). Infection of poultry with C. violaceum is not uncommon, and neither its clinical presentation nor its immune responses to infection are fully understood. These immune mechanisms are central to formulating efficient vaccines and therapeutic approaches to combat and prevent transmission of C. violaceum in poultry targets. The main objective of the present study is to isolate Chromobacterium violaceum from poultry and investigate its immune response in mice.
Samples were collected from various poultry farms in Baghdad, yielding 50 specimens from two-week-old chickens that showed signs of diarrhea. These samples were carefully transported in a cold container to the laboratory within two hours. Each fecal sample was placed in a sterile test tube that contained 1 g of the sample, 10 ml of normal saline, and 0.1 ml of the sample suspension. This mixture was then inoculated onto MacConkey agar (HiMedia, India) and Salmonella Shigella agar (HiMedia, India). The inoculated media were incubated at 37°C for 24 to 48 hours—the Vitek 2 compact system was used for diagnosing the bacteria and antibiotic sensitivity test.
DNA extraction
Transfer 1 to 2 ml of cultured cells into a 2 ml tube. Centrifuge the cells at 13,000 rpm for 1 minute to pellet them. DNA extraction was performed using the G-spin kit (Intron Biotechnology, cat. no. 17045). Electrophoresis was conducted to analyze the DNA fragments post-extraction and to assess the results of the PCR interaction in the presence of standard DNA, allowing for the identification of the product size of the PCR interaction on the Agarose gel.
Purity and concentration of DNA
The concentration and purity of DNA are assessed through absorbance measurements of micro-volume samples utilizing the Nano Drop spectrophotometer (Nabi/Korea). Purity is evaluated using the 260/280 ratio, with optimal purity falling between 1.80 and 2.00.
Primers
The primers were lyophilized and dissolved in ultrapure ddH2O to create a stock solution with a final 100 pmol/μl concentration. This stock solution was stored at -20°C to prepare a working concentration of 10 pmol/μl. The working primer solution was made by combining 10 μl of the stock with 90 μl of ddH2O, resulting in a final volume of 100 μl. The primers were sourced from IDT (Integrated DNA Technologies, USA). Target size for amplification was 1250 bp using the primer sets of 16S rRNA 5'-AGAGTTTGATCCTGGCTCAG-3' and 5'-TACGGTTACCTTGTTACGACTT-3' (Mohammed & Al-Samarraae, 2021).
PCR Amplification Analysis
PCR amplification was performed in a 25 μl volume in the presence of iNtRON's Maxime PCR PreMix Kit that included Taq polymerase, PCR buffer, gel loading buffer, and dNTPs. The mixture consisted of 1.5 μl template DNA, one μl each primer at a concentration of 10 pmol, and 16.5 μl distilled water. Amplification was performed using a Mastercycler (Eppendorf, Germany). The PCR process began with the initial denaturation at 94°C for five minutes. Subsequently, it proceeded with a denaturation at 94°C for 45 seconds, an annealing at 56°C, an extension at 72°C for one minute, and a final extension at 72°C for seven minutes. The products of PCR amplification of 16S rRNA forward and reverse primers were separated on agarose gel electrophoresis (1.5% agarose, 5 volts/cm2) in 1x TBE buffer for 1 hour and 30 minutes. The molecular value standard was a DNA marker (100 bp) of Bioneer (Korea), and a gel documentation system to observe.
The PCR products were analyzed through 2% agarose gel electrophoresis and visualized under ultraviolet light (302 nm), after staining with either ethidium bromide or Red Stain. Macrogen Korea carried out gene sequencing. A homology search was performed utilizing the Basic Local Alignment Search Tool (BLAST), accessible through the National Center for Biotechnology Information (NCBI) at (http://www.ncbi.nlm.nih.gov) and via the BioEdit program.
The C. violaceum Killed Whole Cell Antigen (CKA) was constituted as described in reference (Haq et al., 2022). Bacteria were cultivated on nutrient agar at 37°C for 48 hours, then exposed to Gram staining and microscopic examination for their morphological integrity. The bacteria were washed with PBS (pH 7.2) three times after 3000 rpm centrifugation for 20 minutes and treated with a 0.3% formalin PBS (pH 7.2) solution. The combination was incubated at 37°C for one to two hours and then stored overnight at 4°C. Following three more washes using PBS (pH 7.2), the bacterial suspension was centrifuged for the second time at 3000 rpm for 20 minutes to prepare the CKA. The colony-forming units of the CKA were enumerated by the McFarland tube method, and protein concentration was quantified based on reference (Kielkopf et al., 2020).
Aloe Vera extract was obtained from a local supplement manufacturing company (Alemad Factory) and prepared at a concentration of 100 mg/ml.
Twenty-four Swiss mice weighing 14-15 grams were randomly divided into eight groups. The first group was immunized with a subcutaneous injection of 1.5 × 109 CFU/ml of the CKA antigen. The second group was immunized with the same amount of CKA antigen combined with 100 mg/ml of aloe vera subcutaneously. As a negative control, the third group was given subcutaneously 1 milliliter of PBS (pH 7.2). On day 14 post-immunization, the antigen's initial and second booster doses were administered. After immunization, serum samples were collected on day 21 to measure IgG and IL-6 levels using the Elabscience enzyme-linked immunosorbent assay (ELISA) kit tailored for mice (Elabscience, China), following the manufacturer's protocol. Optical density was measured at a wavelength of 450 nm (Socimed Sarl, France), and the concentrations of IgG and IL-6 were determined through interpolation from a standard curve, with results expressed in pg/mL. A pilot study was conducted to determine infectious dosages for C. violaceum using the McFarland tube method, preparing three dilutions (1.5 × 108, 3.0 × 108, and 6.0 × 108 CFU/ml) with a total of 24 mice divided into three groups of eight. Group 1 was infected orally with 1.5 × 108 CFU (0.1 ml), Group 2 received 3.0 × 108 CFU (0.1 ml orally), and Group 3 was also infected with 3.0 × 108 CFU (0.1 ml orally). Clinical symptoms and mortality rates were observed over three days post-infection, confirming that the infectious dose was 1.5 × 108 CFU/ml. Mice infected with C. violaceum at 21 days post-immunization (1.5 × 108 CFU/ml, orally) were monitored for clinical signs every 6 to 8 hours over a week. Ten days after infection, the serum was taken and the mice euthanized for histopathological examination of the vital organs; liver, kidney, spleen, and intestine were collected and fixed in a 10% formalin solution immediately after extraction (Snyder et al., 2022).
The Statistical Packages of Social Sciences (SPSS) program was used to detect the effect of different groups and periods on study parameters. LSD-Least significant difference (two-way) was used to compare the means significantly in this study (George & Mallery, 2019).
The analysis of fifty fecal samples from diarrhea cases revealed that seven isolates, representing 14%, tested positive, as indicated in Table 1.
All strains of C. violaceum manifested as black colonies on SS agar due to their ability to produce H2S after 24 hours (Figure 1). On MacConkey agar, the isolates appeared as pink colonies due to lactose fermentation (Figure 1) and non-producing pigmentation on nutrient agar (Figure 1). Additionally, all isolates tested positive for oxidase and catalase.
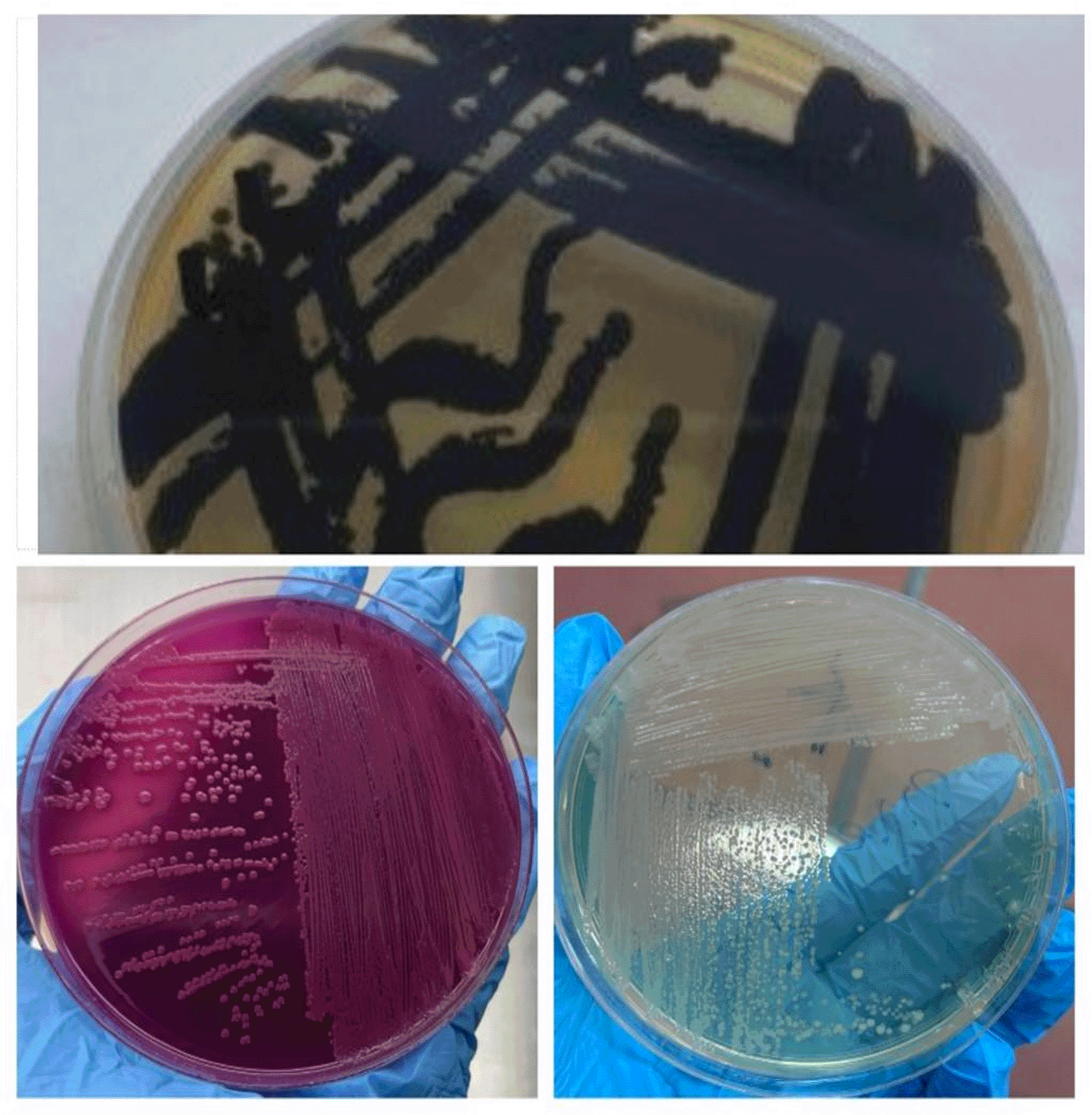
The Vitek 2 Compact system was employed to identify and assess the antimicrobial susceptibility of C. violaceum, yielding a 98% probability of identification, as illustrated in Table 2.
When standard DNA is present, the PCR interaction's outcome allows for the product size differentiation resulting from the PCR process on the Agarose gel (Figure 2).

The concentration of C. violaceum DNA was measured at 42.5 ng/ml, with a purity value of 1.937. One isolated strain was examined to identify the presence of 16S rRNA genes. As a result, the strain showed a positive amplification of 1250 bp on an agarose gel, while no amplification was detected in the negative control (Figure 3).
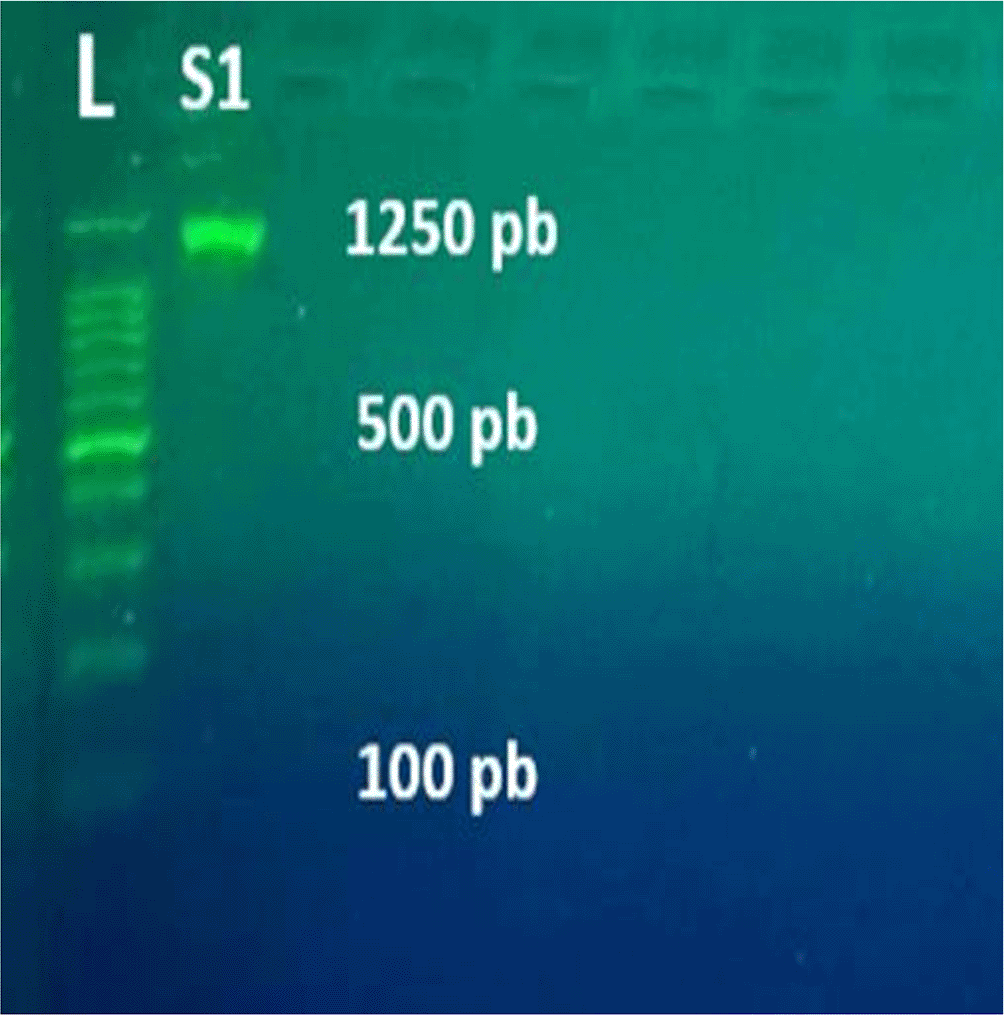
L: represents the DNA ladder (100 bp). S1: C. violaceum.
The NCBI website (http://www.ncbi.nlm.nih.gov) was used to analyze the sequences. There was 98% homology between the C. violaceum reference strains and the isolated strains in GenBank (Accession No. MW282912.1) Figure 4 Table 3.

The results regarding IL-6 levels across the various groups (G1 and G2) indicated no significant differences (P<0.05) at 21 weeks post-immunization, with measurements of 34.98 ±0.97 pg/ml for G1 and 35.37 ±0.54 pg/ml for G2 when compared to the control negative group, which had a level of 30.93 ±0.62 pg/ml. Furthermore, Group 3 (control negative) demonstrated a significantly elevated IL-6 concentration (P<0.05) ten days following infection with C. violaceum, reaching 945.06 ±9.87 pg/ml. Additionally, both G1 and G2 showed significant increases in IL-6 levels (P<0.05) ten days post-infection, measuring 287.54 ±13.75 pg/ml and 302.11 ±14.68 pg/ml, respectively, as detailed in ( Table 3) and (Figure 5).
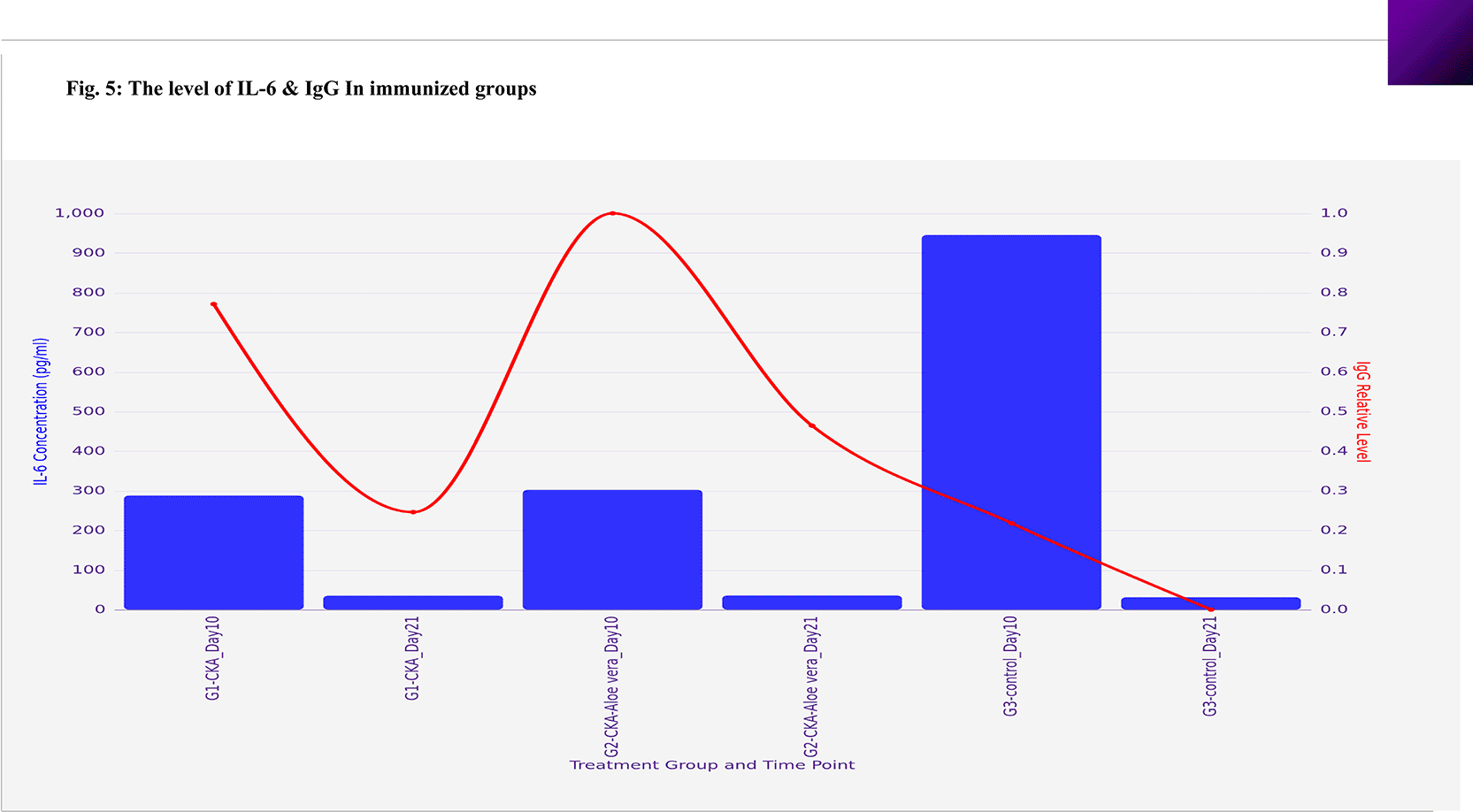
The findings related to IgG levels among the different groups (G1 and G2) revealed significant variations (P<0.05) at 21 weeks after immunization. Specifically, G1 presented an average of 34.98 ±0.98 pg/ml, while G2 showed a higher average of 55.43 ±0.49 pg/ml, in contrast to the negative control group, which recorded a level of 11.93 ±0.70 pg/ml. Additionally, Group 2 exhibited a notably increased IL-6 concentration (P<0.05) ten days post-infection with C. violaceum, reaching 105.61 ±2.15 pg/ml. In comparison, G1 attained a level of 84.19 ±1.60 pg/ml, whereas G3 reached only 32.39 ±1.16 pg/ml, as outlined in (Table 4) and (Figure 5).
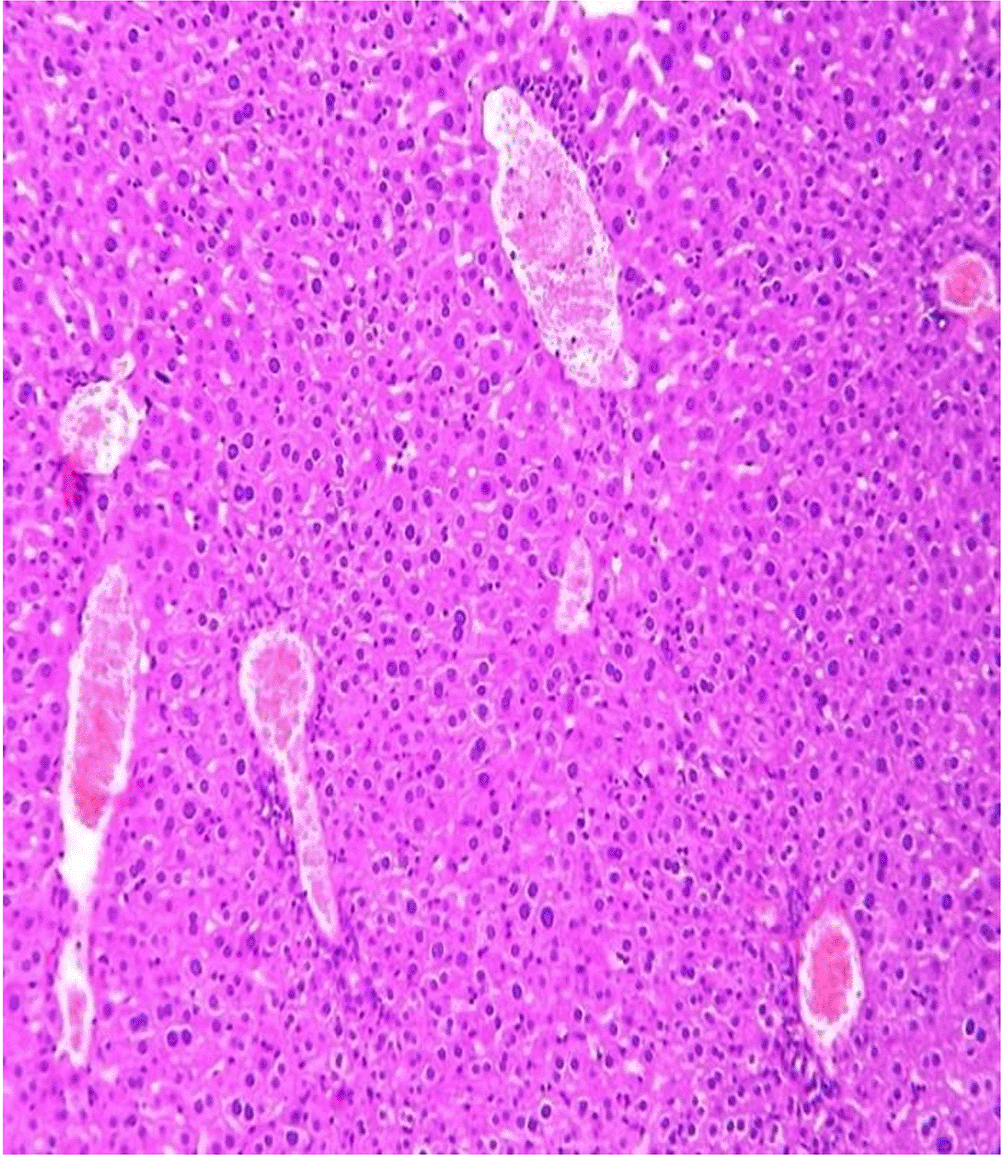
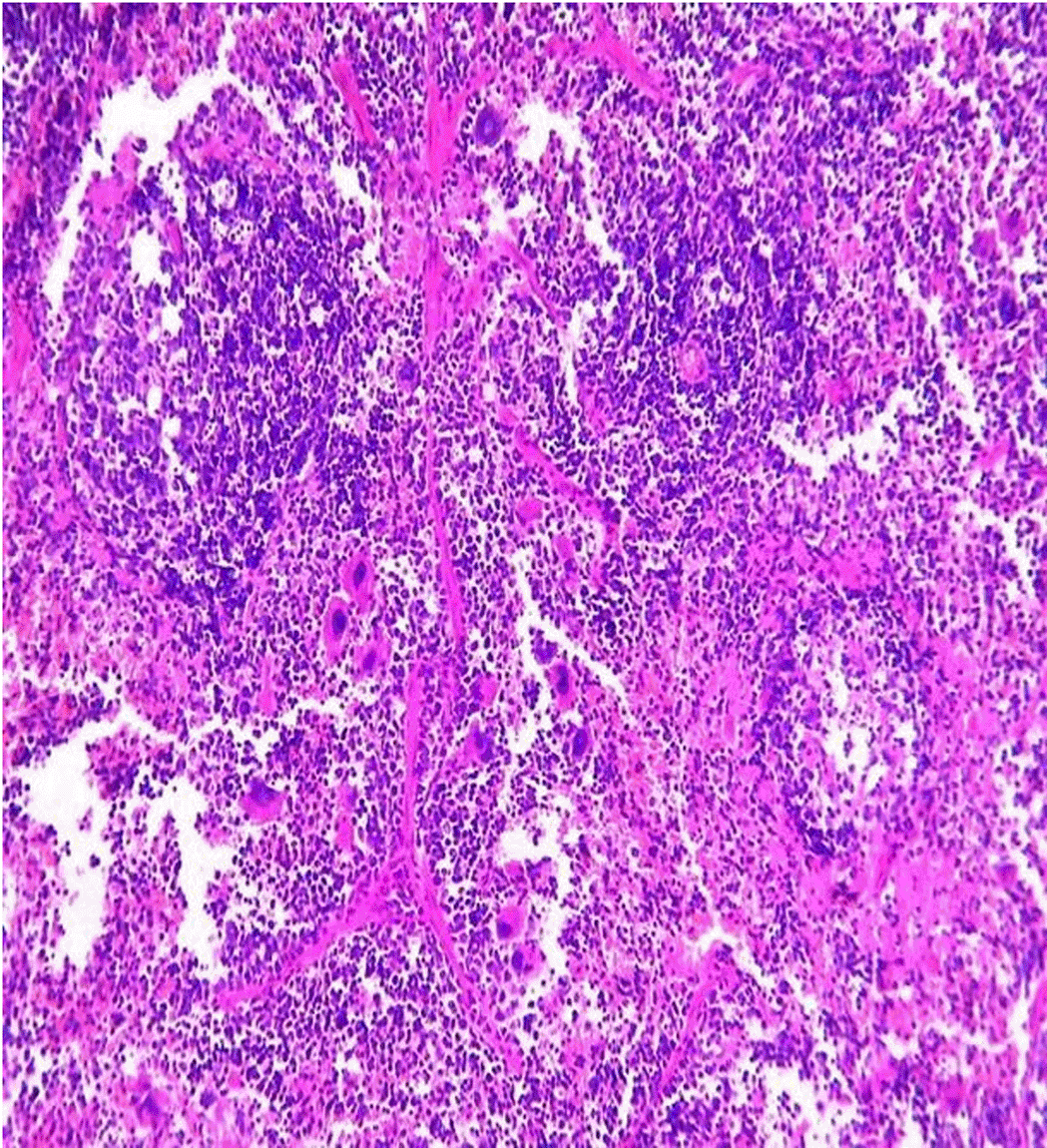
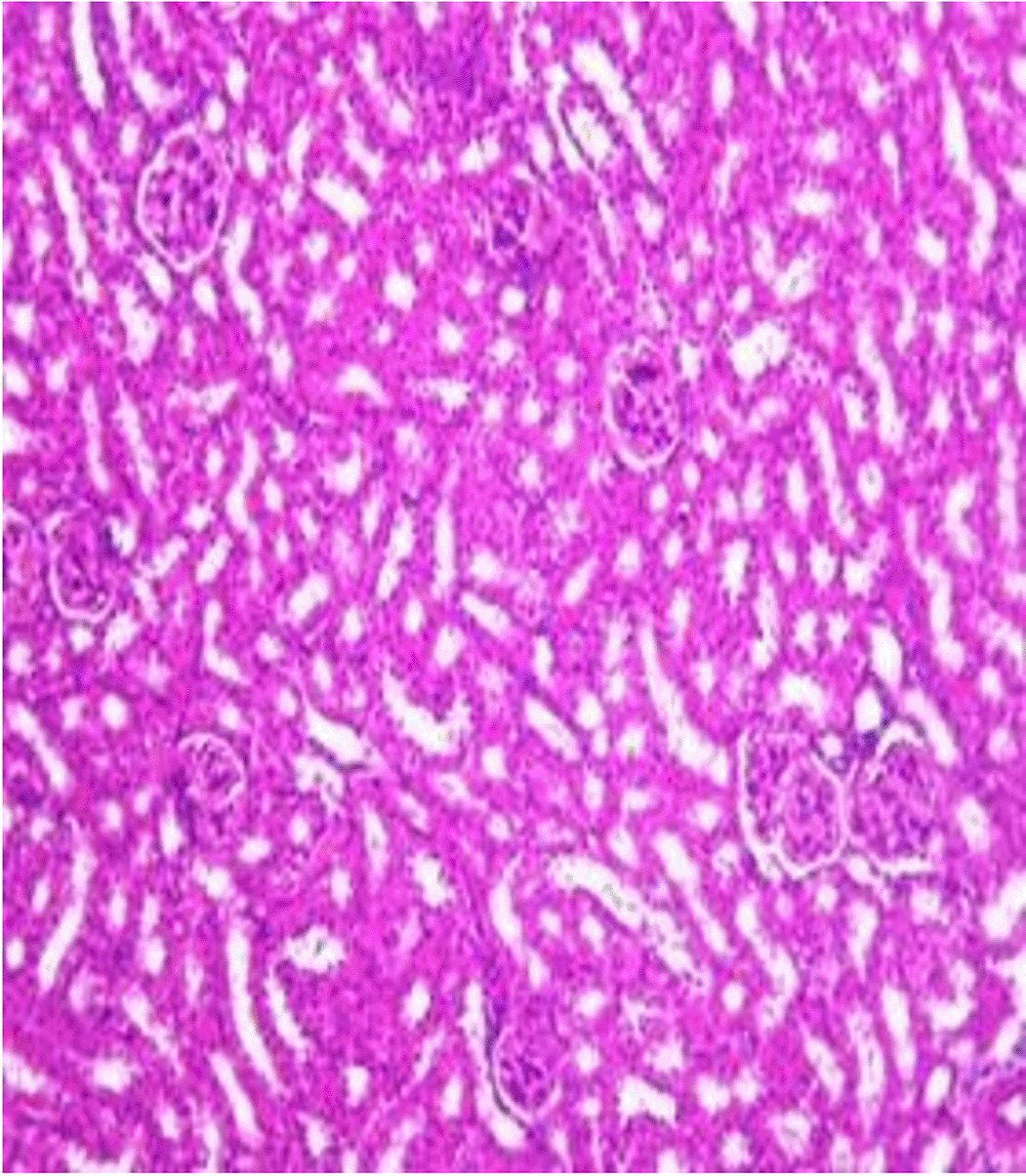
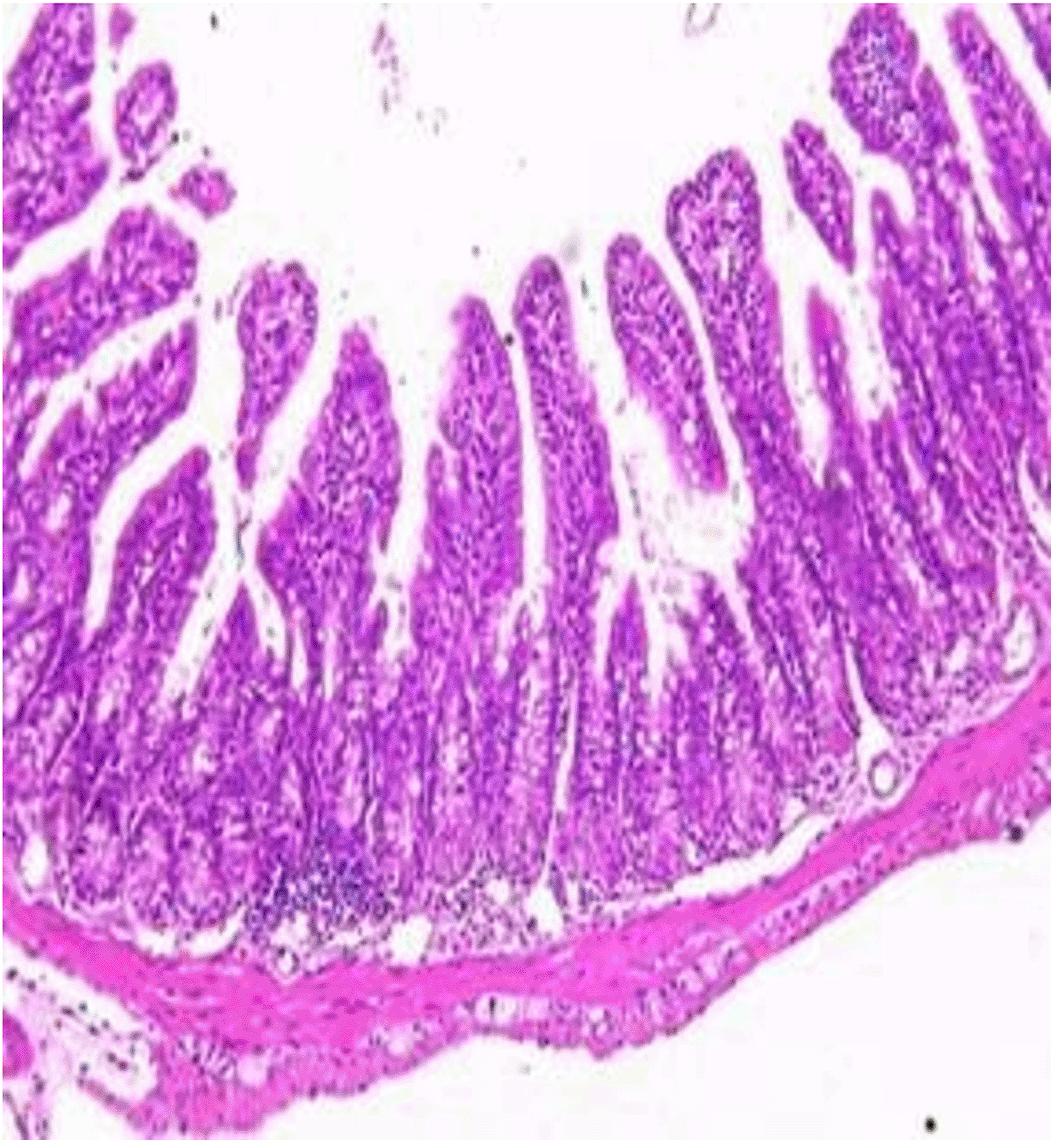
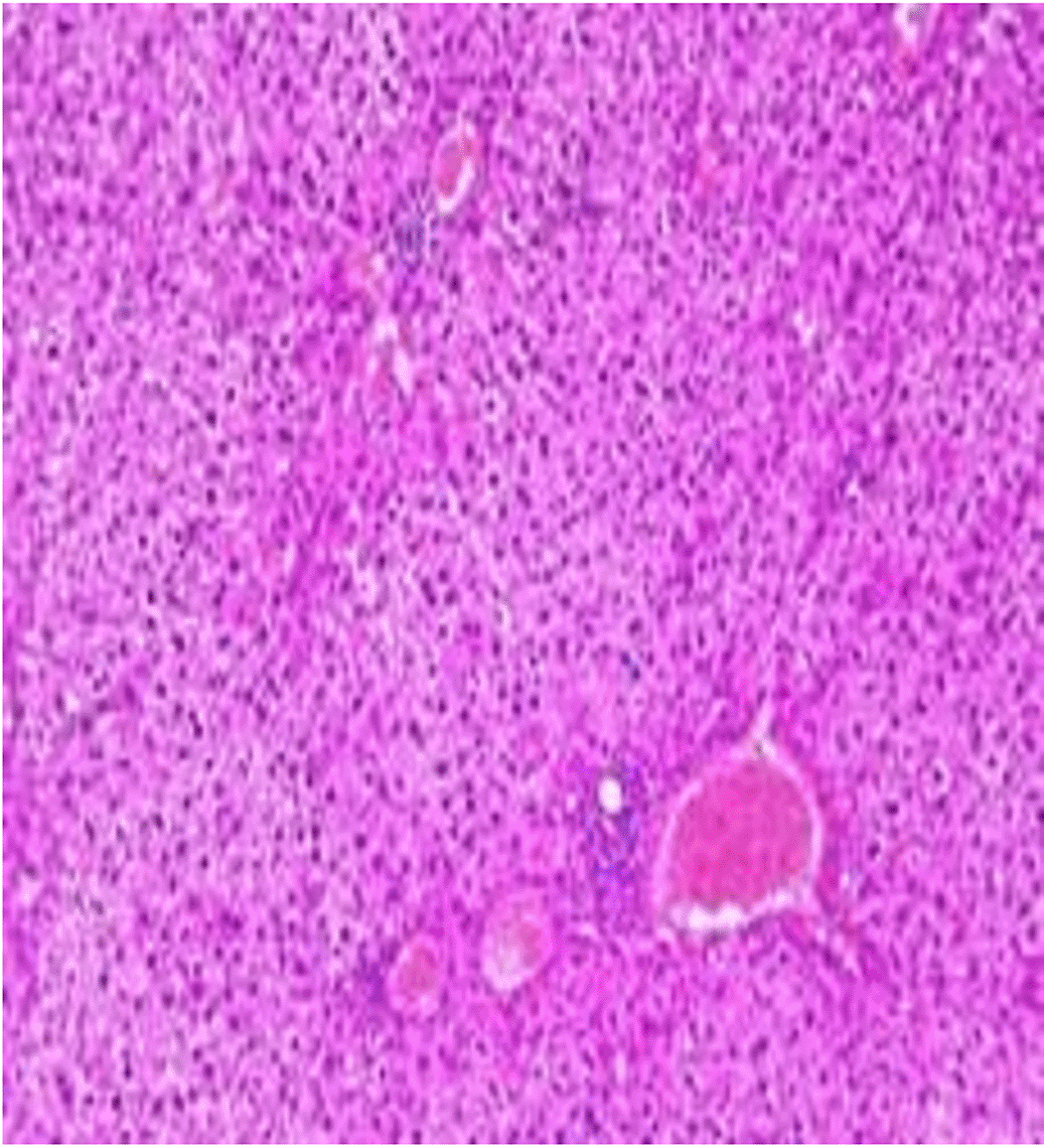
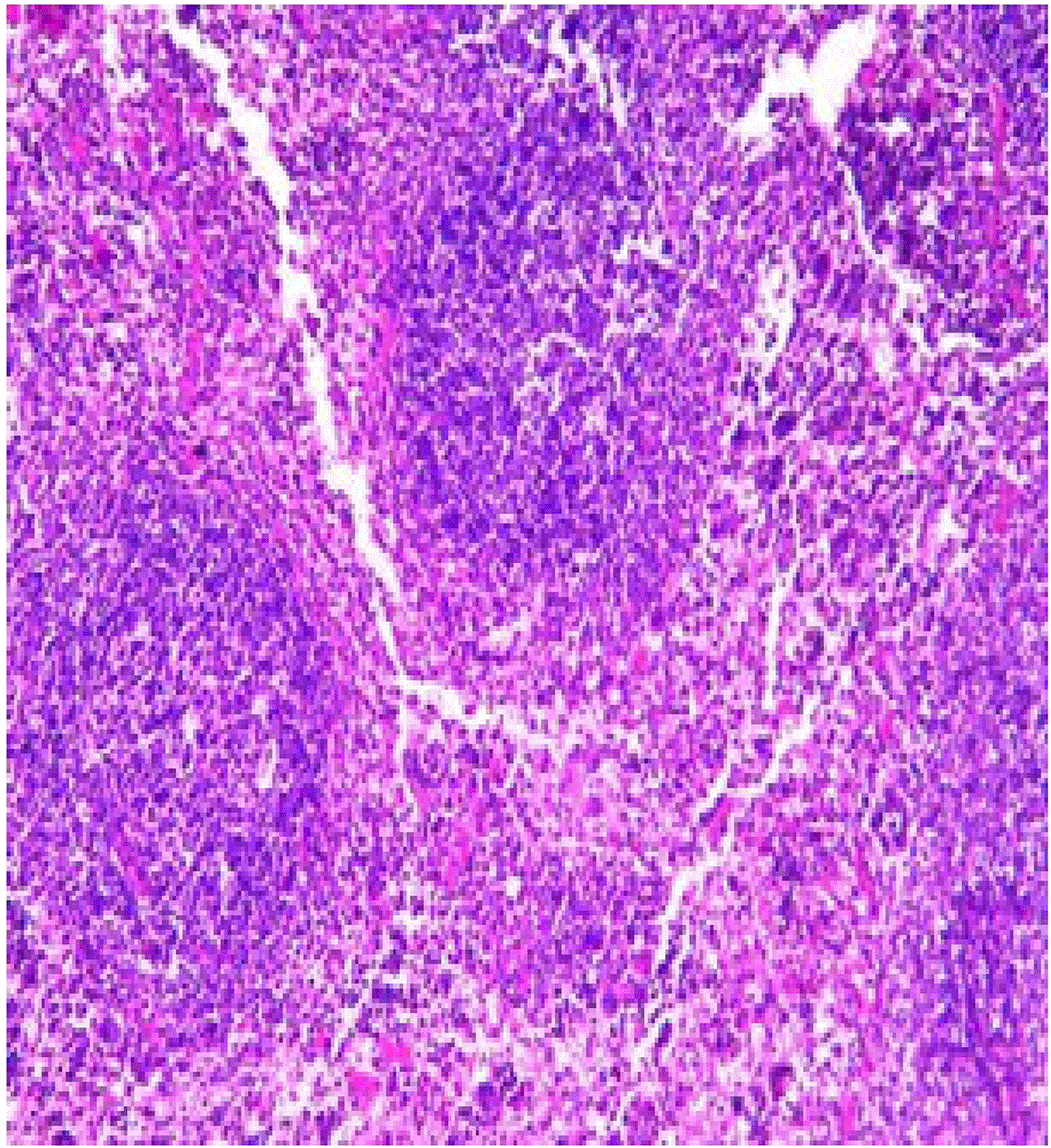
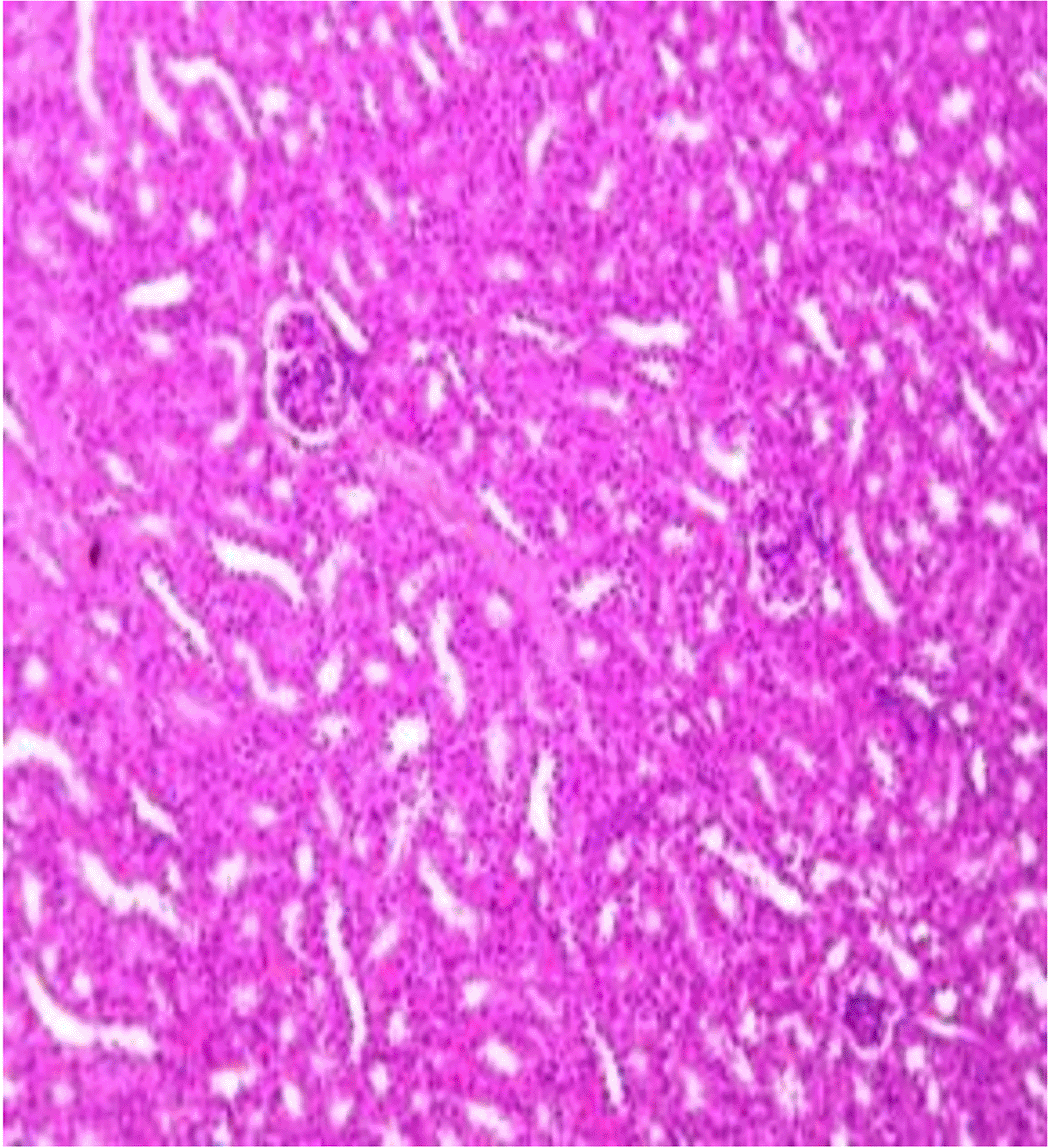
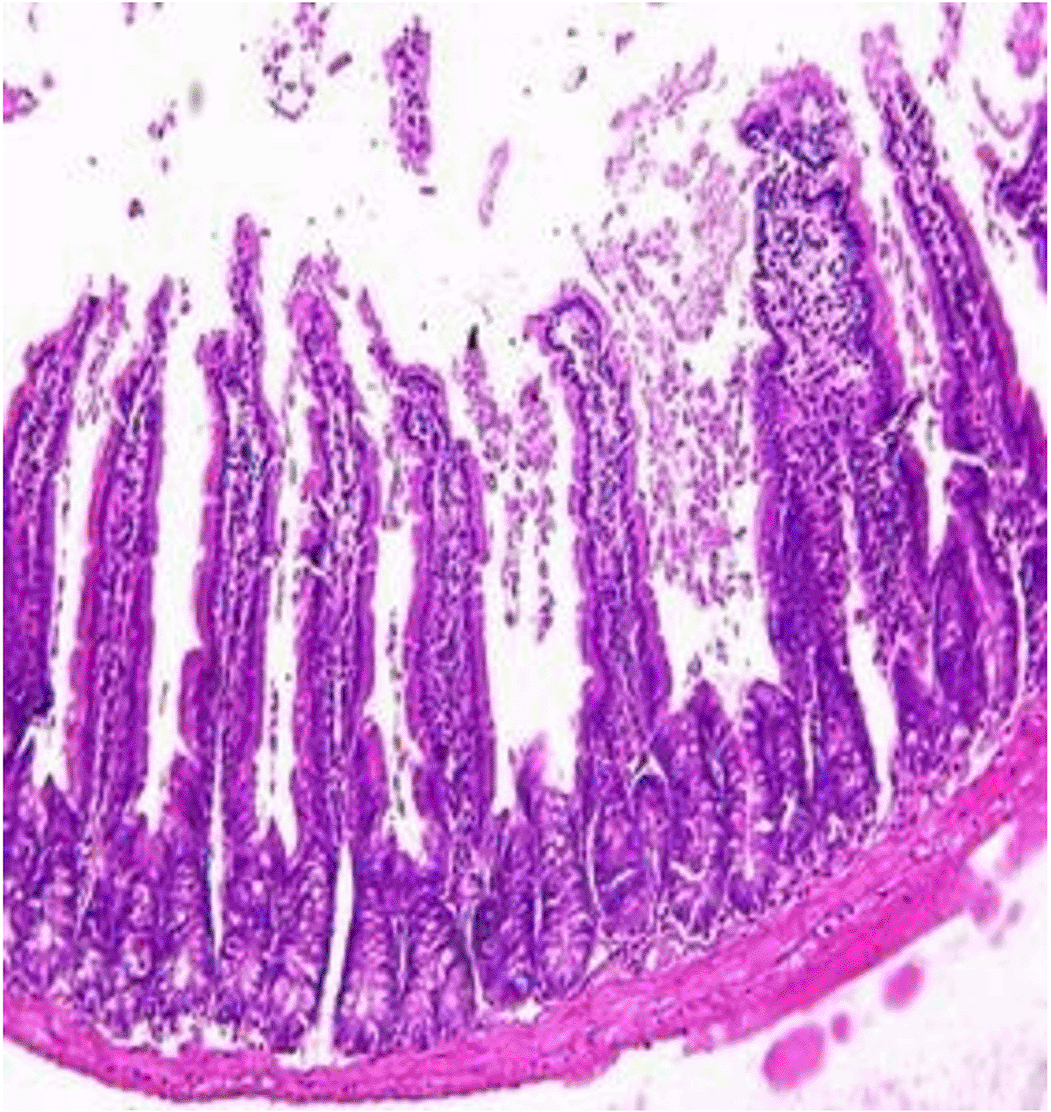
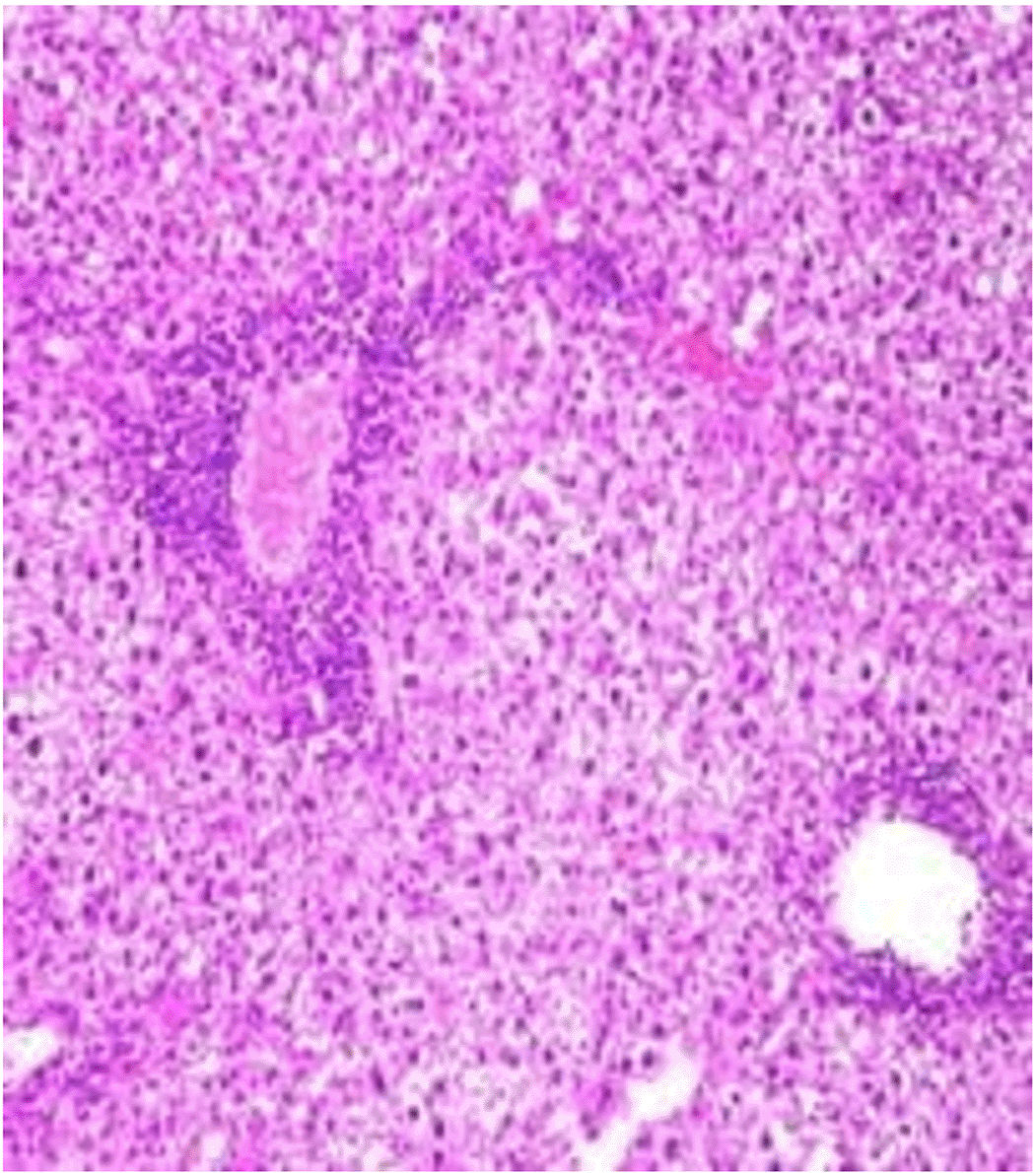
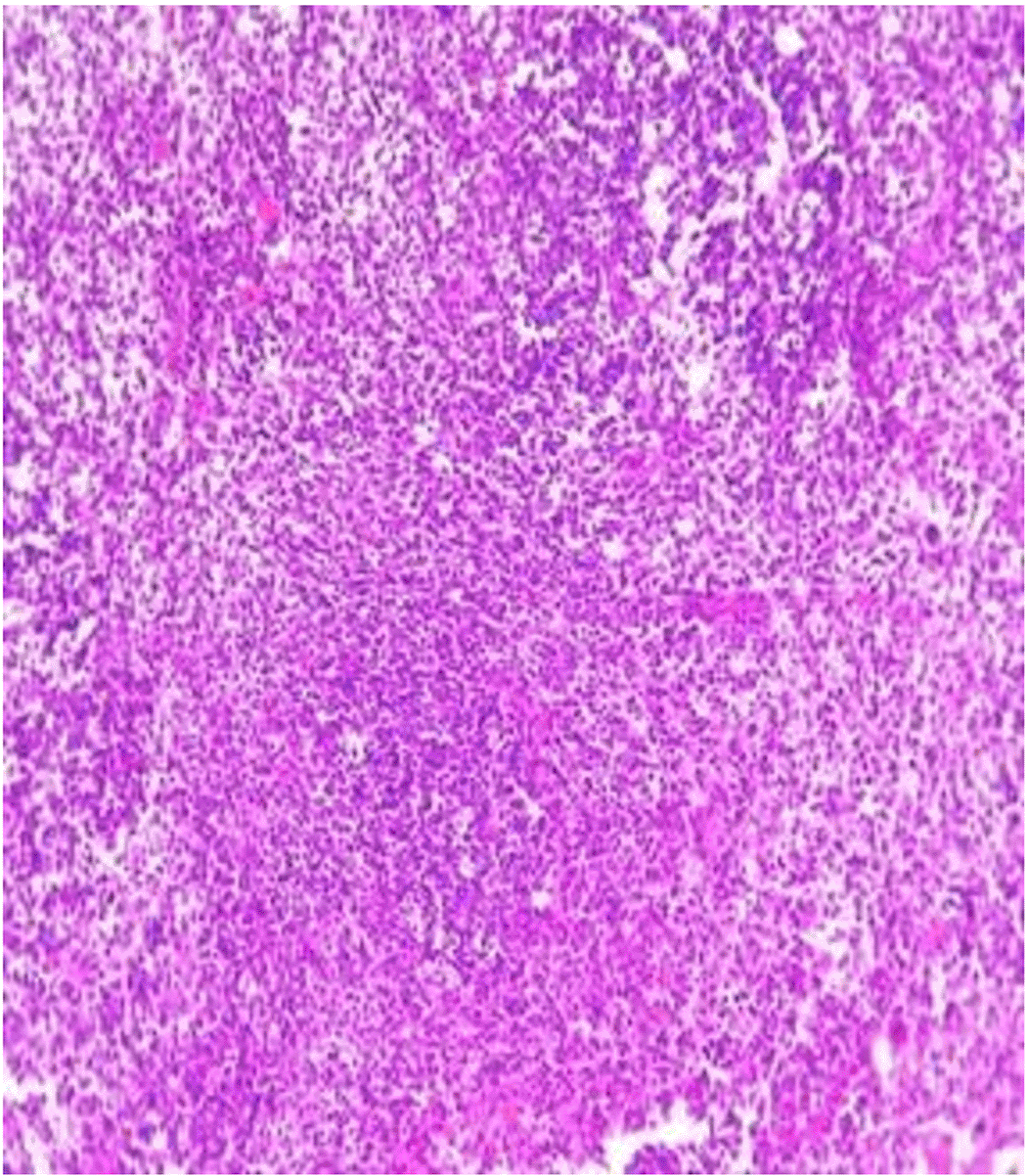
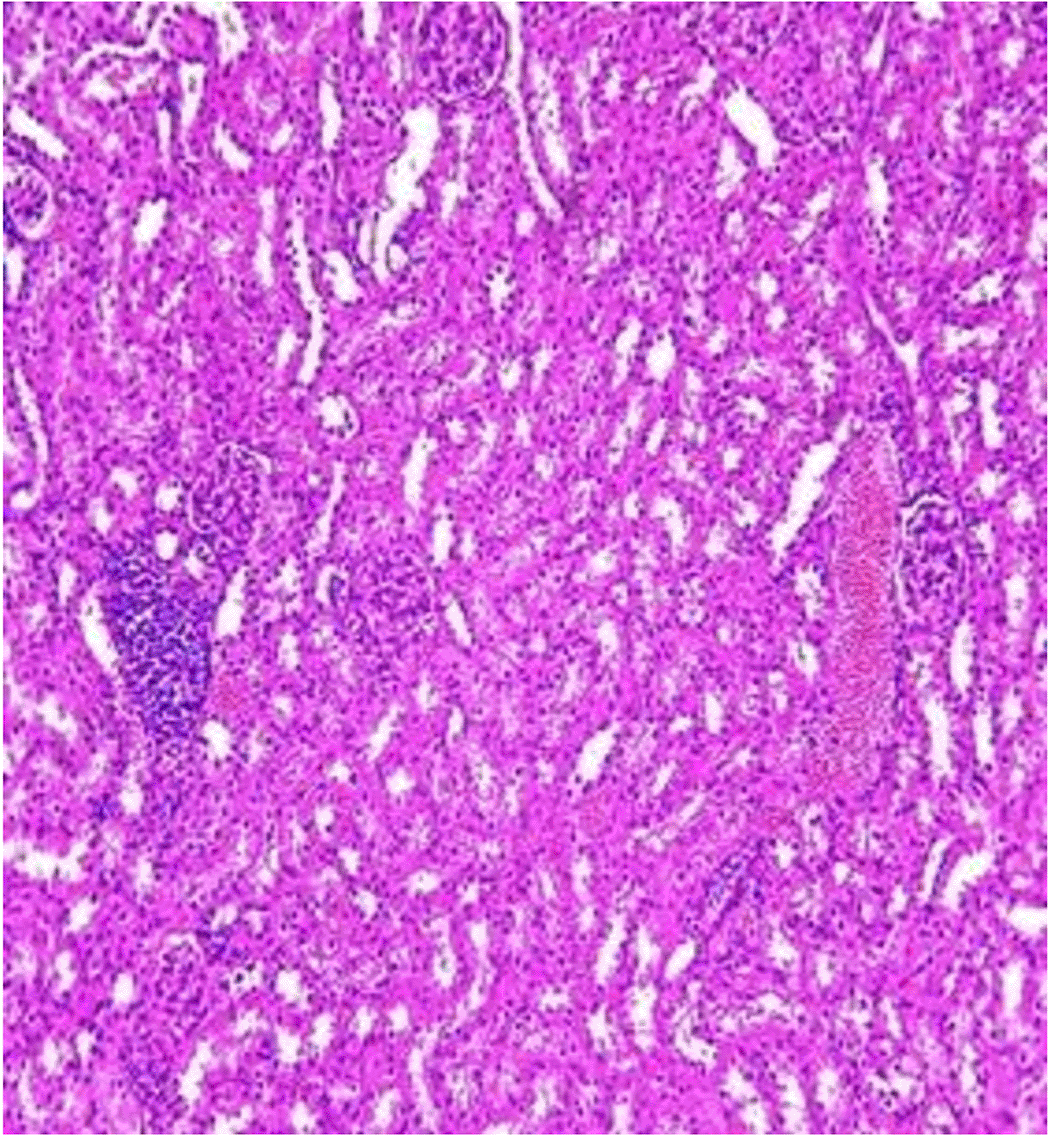
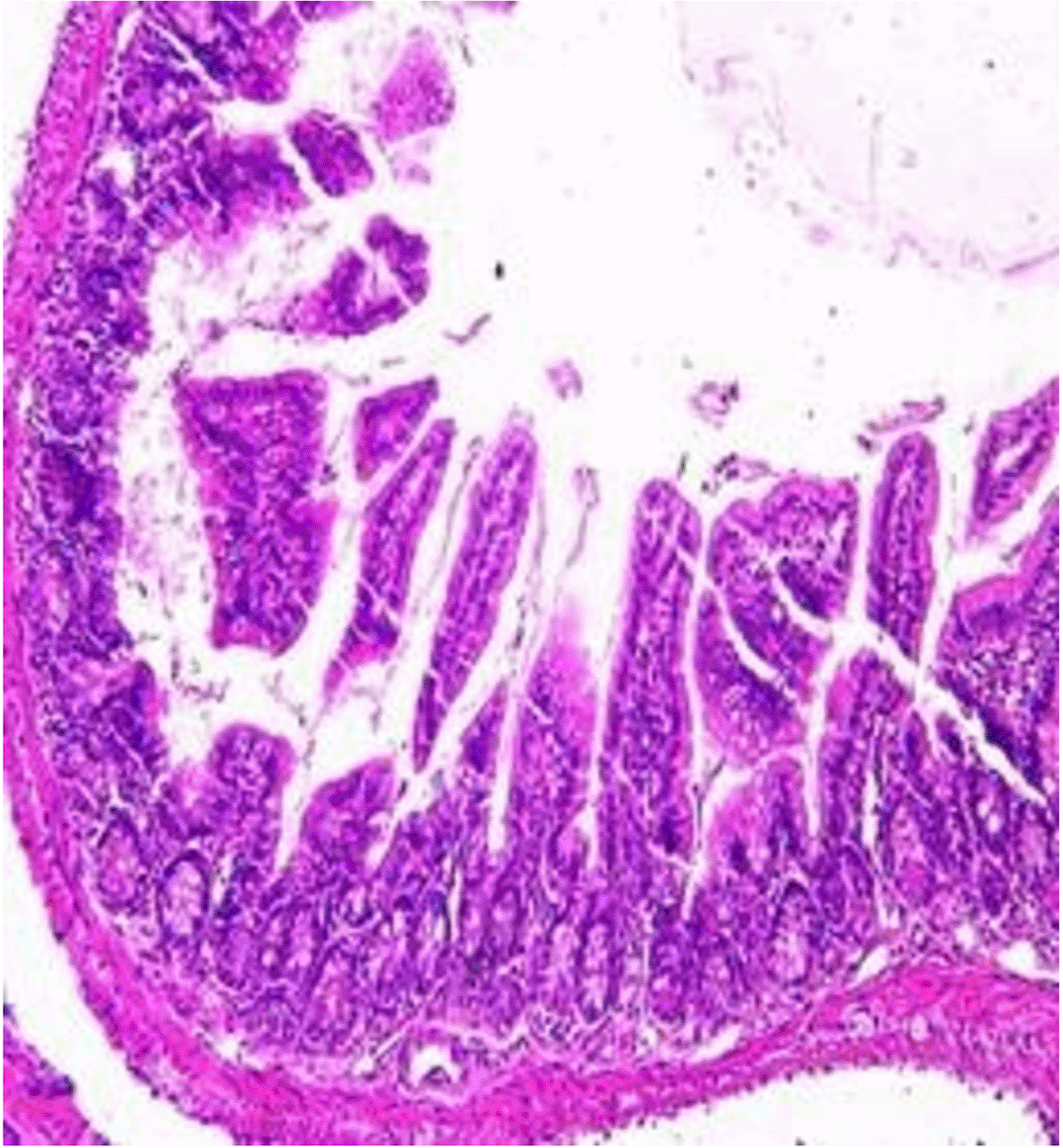
Histopathological examination after 10-day post-challenge showed that all groups under study were affected with different histopathological changes: The G1 CKA: the liver section revealed normal parenchyma with well-conserved hepatic cords and sinusoids, normal liver histology, no pathological lesions identified (Figure 6). The spleen section showed the presence of well-formed granulomas—collections of epithelioid histiocytes (elongated, pale cytoplasm) often surrounded by lymphocytes and increased vasculature with some fibrous septae (Figure 7). The kidney section showed histologically normal renal cortex, and no apparent pathology was present in this section (Figure 8). The intestinal section showed intact villous architecture with slight development of subepithelial (Gruenhagen's) spaces at the villus tip, extremely mild mucosal injury (Figure 9). The G2 CKA- Aloe vera: the liver section showed mild portal inflammation with lymphocytic infiltrates in and around portal tracts, no significant lobular inflammation or hepatocyte necrosis (Figure 10). The spleen section revealed Lymphoid hyperplasia or possible lymphoma and dense lymphocytic infiltrate (Figure 11). The kidney section showed no obvious lesions such as necrosis, inflammatory cell infiltration, fibrosis, or glomerular sclerosis (Figure 12). The intestinal section showed moderate subepithelial space formation and epithelial lifting to the sides of villi (Figure 13). The G3 Control negative (infected = C+): the liver section showed dense infiltrate of lymphocytes in the portal area extending into the periportal area, and interface hepatitis is seen (piecemeal necrosis), scattered lobular inflammation, and mild hepatocyte ballooning (Figure 14). The spleen section showed preserved follicular hyperplasia, tubules appeared dilated with cytoplasmic vacuolization, and moderate lymphocytic infiltration in interstitial areas (Figure 15). The kidney section revealed Diffuse lymphocytic infiltration, focal necrosis, and congestion in zones, glomerular structure observed to be mildly disturbed, severe damage to parenchymal tissue evident (Figure 16). The intestinal section revealed massive epithelial lifting and few villi with denuded tips, demonstrating advancing mucosal injury (Figure 17)
Chromobacterium violaceum is involved in opportunistic infections among humans and other animals (Hammerschmitt et al., 2017); however, it is also found in avian hosts, especially chickens. A critical literature review indicates that there have been no reports of isolating C. violaceum from chickens, even from diarrhea in feces. Thus, this study is the first report on isolating C. violaceum from chicken diarrhea cases. The results show that among 50 fecal samples of chickens presenting with diarrhea, 7 C. violaceum isolates were obtained. These results are in concordance with other research where this bacterium has been isolated from other household animals; it has been found in visceral abscesses, pleuropneumonia, and septicemia in pigs (Liu et al., 1989), sheep (Carrasco et al., 1996), dogs (Crosse et al., 2006), cattle (Ajithdoss et al., 2008), and horses (Hammerschmitt et al., 2017). The bacterium was separated to the Genus level by the routine methods, whereas Vitek2 and PCR were employed to differentiate the species. This study's result detected black and non-pigmented colonies on SS agar. This is consistent with Sivendra et al.'s (1975) study, in which pigmented and non-pigmented colonies of C. violaceum water isolates were detected. The VITK2 test was 98% likely, whereas C. violaceum was 98% identical with GenBank MW282912.1 for the presence of the 16s rRNA gene. The findings concur with research by Ibrahim et al. (2020). Isolate C. violaceum VITK2 was the one with a 99.9% matching rate. In the meantime, PCR analysis also indicated the 16S rRNA gene sequence to be 96.58%-99.31% identical to C. violaceum ATCC 12472, C. violaceum NBRC 12614, and C. violaceum JCM 1249 strains of other species within the same genus.
The current study showed that IL-6 levels in all the immunized groups were not statistically different (P<0.05) at the 21st week of immunization. There was an increase in ten days post-infection with C. violaceum. In contrast, there was a difference (P<0.05) ten days post-infection with C. violaceum in the negative control group. These actions can be justified by the induction of inflammatory cytokines and inhibition of the macrophage and neutrophil microbicidal functions and neutralization of IL-6 in infected test animals that could lead to the reduction of mortality (Mohammed, 2025c; Ulich et al., 1991). From the findings, the IgG antibodies in all the immunized groups had an increased antibody titer (P<0.05) 21 days post-immunization. There was also an increase in the antibody level in all groups vaccinated ten days after bacterial infection relative to the control. All these results agree with findings made in (Oladejo and Adebolu, 2013), where they noted that the initial antibody response to the E. coli O157 antigen increased by the first dose and once again reached its peak after booster administration. On the heels, Al-Samarraae and Kareem (2018) reported a very marked rise in IgG titers (P<0.05) after 35 days of vaccination with sonicated Salmonella Typhimurium (KWCSA-ST) and Lactobacillus acidophilus (KWCSA-LBA). This was consistent with the findings of (Mohammed & Al-Samarraae, 2021), who reported that Citrobacter WCSA and LPS Ag-vaccinated rabbits induced elevated humoral (IgG) and cellular (DTH and IL-6) immune responses.
The histopathological results highlight the immunoprotected function of immunization in averting undue damage to tissue in infection: Group 1 (G1), which received the killed antigen alone, exhibited a disease, compared to the other groups, whose immune response was significantly more severe. This agrees with studies that show killed bacterial vaccines to cause immunity without causing excessive tissue damage (Cavallaro et al., 2020). Group 2 (G2), which was vaccinated with the killed antigen and Aloe vera, had heightened immunity with greater inflammation of the spleen and liver, but not excessive tissue damage. The immunomodulatory action of Aloe vera likely resulted in this increased immunity without inflicting so much destruction, evidenced by studies showing that Aloe vera stimulates immunity without exposing toxic side effects (Vijayalakshmi et al., 2015; Nasir et al., 2019). In Group 3, infected with C. violaceum and considered (C+), there were severe histopathological changes in the liver, kidney, spleen, and intestine. The tissue damage in specific organs reflects the inability of the immune system to contain the infection without prior immunization. The above observations highlight the role of immune priming (through vaccination or prior exposure) in preventing severe infection and associated organ injury. This highlights the importance of vaccine immunity and the need for pre-exposure immunity to effectively combat bacterial infections like C. violaceum (Zhao et al., 2020; Deng et al., 2017).
The study is the initial reported isolation of Chromobacterium violaceum from diarrheic poultry. The study demonstrates that immunization with antigens of C. violaceum, in combination with Aloe vera as an immunomodulator, increases humoral and cellular immunity, which confers protection against infection. The study shows that C. violaceum can potentially injure poultry health, and active immunization may significantly prevent severe infection and associated tissue damage. Additional research is recommended to study vaccination efficacy in larger poultry flocks and the bacterium's role in avian disease.
This study received local ethical permission through the local animal care and use committee at the College of Veterinary Medicine at the University of Baghdad (Number P-G\582,6\3\2025).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)