Keywords
Cisplatin and ginseng; Hepatotoxicity; Drug-metabolizing-enzymes; Oxidative-stress/Inflammation; Apoptosis.
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Cisplatin and ginseng; Hepatotoxicity; Drug-metabolizing-enzymes; Oxidative-stress/Inflammation; Apoptosis.
Cisplatin (CIS) is one of the potent cytotoxic antineoplastics that is extensively used against a variety of types of solid tumors and is approved by the FDA. The pharmacological target of cisplatin refers to its irreversible binding to DNA, which blocks its transcription and replication, leading to cell apoptosis. Additionally, CIS-induced reactive oxygen species (ROS) generation induces oxidative damage to several biomolecules, which ultimately disrupts the cell integrity (Ijaz, et al., 2020; Abd Rashid, et al., 2021). The clinical application of cisplatin is obstructed due to its resistance and undesirable pathological impact, prominently nephrotoxicity, hepatotoxicity, gastrointestinal toxicity, uterine toxicity, ototoxicity, and neurotoxicity (Abdel-Daim, et al., 2020; Hwang, et al., 2020; Gholampour, et al., 2022; Lin, et al., 2024). Cisplatin hazard toxicity on healthy cells is propagated via induction of ROS, pro-inflammatory cytokines, DNA deterioration, caspase activation, mitochondrial dysfunction, and apoptosis. Therefore, banning cisplatin’s toxic effects is one of the important issues within the treatment plan (Dasari, et al., 2022).
The propagation of free radicals induces the up-regulation of nitric oxide synthase, NADPH oxidase, and cyclooxygenases, which manifestly alter the mitochondrial functions. Overproduction of ROS and RNS (reactive nitrogen species) can modify the redox system enzymes, which in turn negatively disrupt the cell’s functions (Mohamed and Badawy, 2019). Oxidative stress overproduction, with the reduction in the antioxidant defense system is implicated in cisplatin-induced mitochondrial toxicity and liver injury (Dasari et al., 2022). Moreover, cisplatin upregulates the expression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) and apoptotic pathway via the activation of the apoptotic markers (caspase, tumor protein 53, Bax) and the suppression of the anti-apoptotic markers as Bcl-2 (Abd Rashid et al., 2021).
Drug-metabolizing enzymes (DMEs) are responsible for the metabolism of exogenous xenobiotics (environmental pollutants, pesticides, carcinogens, drugs, and food toxicants) and endogenous compounds (steroids, bile acids, and prostaglandins). DMEs are generally classified into two main phases. Phase I covers oxidation-reduction and hydrolytic enzymes such as cytochrome P450 (CYP), flavin-containing monooxygenases, and monoamine oxidases. Phase II involves conjugative enzymes, including sulfonation, glucuronidation, methylation, acetylation, glutathione-S-transferases, and amino acid conjugation (Kaur, et al., 2020; Al-Hamadani, et al., 2020; Zeng, et al., 2022). One of the mechanisms by which cisplatin generates ROS is the destruction of the cytochrome P450 system, which leads to excessive production of ROS and oxidative stress (Zhou, et al., 2022).
Previous studies reported that a combination with complementary medicine can offer a satisfying solution for boosting the chemotherapeutic impact of cisplatin and minimizing its hazardous effects (He, et al., 2022). Ginseng (GIN) is one of the herbal remedies that has numerous bioactive pharmacological properties, such as antitumor, neuroprotective, cardiovascular-protective, hepatoprotective, antivirus, anti-allergic, antiangiogenetic, antiapoptotic, antidiabetic, antioxidant, anti-aging, and immunoregulatory effects (Zhou, et al., 2020; Ji, et al., 2022, Wu, et al., 2025). Ginseng and its constituents have a therapeutic role against COVID-19 by targeting inflammasomes and boosting immune system functions (Yi, 2022). Additionally, it has a hepatoprotective effect through detoxifying H2O2-induced hepatotoxicity in HepG2 cells and maintains the cellular redox status (Choudhry, et al., 2019).
The pharmacological effect of ginseng is associated with ROS scavenging through the induction of the Nrf2 pathway and heme oxygenase-1 (HO-1). The antioxidant impact of ginseng might be mediated through the activation of different cellular signaling pathways involving the heme oxygenase system. Also, GIN is used in cancer therapy programs due to its ability to protect against chemotherapy-induced damage (Carota, et al., 2019).
To the best of our knowledge, this study is among the first to comprehensively investigate the protective role of ginseng against cisplatin-induced hepatotoxicity with a particular focus on the modulation of hepatic drug-metabolizing enzymes together with oxidative stress, inflammatory responses, apoptosis, and histopathological alterations. Understanding the mechanisms underlying cisplatin-induced hepatotoxicity and identifying safe protective agents remain important research priorities for improving the therapeutic outcomes of chemotherapy.
Accordingly, GIN can be prescribed as an adjuvant with cisplatin therapy. Hindering liver damage by cisplatin is our goal in this research. Therefore, the current study was designed to assess the therapeutic capability of ginseng as an antioxidant agent against the hepatoxicity induced by the chemotherapeutic agent’ cisplatin. We evaluate specific markers that are responsible for liver injury, which include: DMEs, oxidative stress, inflammation, apoptosis, histopathology, and immunohistochemistry.
Cisplatin and Panax ginseng were obtained from Mayne Pharmaceuticals-UK and Pharco Pharmaceuticals-Egypt, respectively. The treated doses of cisplatin (4 mg/kg BW/week) and Panax ginseng (100 mg/kg BW/day) were chosen according to Jangra et al. (2016) and Voces et al. (2004), respectively. All other chemicals applied in this study were of analytical grade. Additionally, all reagents were employed at concentrations recommended by the manufacturer’s instructions.
Forty male Sprague-Dawley rats weighing from 160 to 180 g were obtained from the Faculty of Medicine, Alexandria University, Egypt. The research was achieved according to the Guide for the Care and Use of Laboratory Animals (International Council for Laboratory Animal Science, ICLAS) and was approved by the local ethical guidelines of the Institutional Animal Care & Use Committee (IACUC), Alexandria University, Egypt (AU14–200922–3-3) and all the methods were performed according to the guidelines and regulations of the same Committee. Also, the study was implemented according to the ARRIVE guidelines (Percie du Sert, et al., 2020). Additionally, the study was conducted following the Basic & Clinical Pharmacology & Toxicology policy for experimental and clinical studies (Tveden-Nyborg, et al., 2023).
Animals were housed and kept in a 12-hour light/dark cycle at room temperature. Food and water were available ad libitum. Randomly, all the experimental animals were divided into 4 groups (10 animals in each group). Group 1 served as control; group 2 was treated orally with Panax ginseng (100 mg/kg BW/day). Group 3 was injected (intraperitoneally) with cisplatin at a dose of 4 mg/ kg BW/week. Group 4 (combined group) received an oral dose of Panax ginseng (100 mg/kg BW/day) in addition to an intraperitoneal dose (cisplatin 4 mg/kg BW/week). The animals were treated once a week with cisplatin and daily with Panax ginseng for three months.
Animals were euthanized following anesthesia induced by isoflurane inhalation (approximately 2% delivered in a desiccator chamber). Adequate depth of anesthesia was confirmed by the absence of the pedal withdrawal reflex and loss of response to external stimuli. Euthanasia was completed by maintaining isoflurane exposure until respiratory arrest occurred, followed by cardiac puncture for blood collection. All procedures were conducted in accordance with the recommendations of the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals. Following anesthesia, blood was drawn from the heart and centrifuged for 10 minutes at 860 xg. Serum samples were obtained and stored at −80 °C for laboratory tests. Liver samples were obtained and divided into 2 parts. The first part was rinsed in cold saline solution and homogenized (10%, w/v) in 0.1 M phosphate-buffered (pH 7.4). The supernatant was obtained by centrifugation at 5000 rpm for 15 min. and stored at −80 C for biochemical assay. The second part was used for histopathological examination.
The total hepatic contents of the cytochrome P450, cytochrome b5, NADPH-cytochrome C-reductase, amidopyrine N-demethylase, and aniline 4-hydroxylase were estimated by the methods of Omura and Sato (1964), Williams and Kamin (1962), Nash (1953), and Kato and Gillette (1965), respectively.
According to the manufacturer’s instructions, total RNA was isolated from liver tissues using an RNeasy Plus kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The purity and integrity of RNA were determined spectrophotometrically at 260/280 nm. Reverse transcription was performed to convert RNA to cDNA using a reverse transcriptase kit (Maxime RT PreMix kit, iNtRON, Korea). The amplification of CYP1A1, CYP2E1, CYP2D6, and CYP3A4 genes was performed by quantitative PCR assay using Rotor-Gene SYBR® Green with low rox qPCR Kit (Enzynomics, Korea). Specific primers (Life Technologies, Prague, Czech Republic) were used to determine the mRNAs of CYP1A1, CYP2E1, CYP3A4 (CYP3A1 in rats) and CYP2D6. The sequences of the primers used are presented in Table 1. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. The resulting data were analyzed by the ΔΔCt method.
Malondialdehyde (MDA) was assayed as a lipid peroxidation index in liver homogenate. In which thiobarbituric acid-reactive substances (TBARS) were measured at 532 nm using 2-thiobarbituric acid (2,6-dihydroxypyrimidine-2-thiol; TBA; Sigma-Aldrich, Cat. No. T5500, St. Louis, MO, USA). An extinction coefficient of 156,000 M−1 Cm−1 was used for calculation (Esterbauer and Cheeseman, 1990). Nitric oxide (NO) was measured using the Griess reaction (Sosroseno et al., 2008). In which about 150 μl of liver extract was mixed with 150 μl Griess reagent (Sigma-Aldrich, Cat. No. G4410), and absorbance was recorded at 540 nm. Superoxide dismutase (SOD; EC 1.15.1.1) activity was measured according to Marklund and Marklund (1974). In which 20 μl tissue supernatant and 10 μl pyrogallol (Sigma-Aldrich, Cat. No. P3130) were added to one ml buffer reagent. The inhibition in pyrogallol autooxidation by SOD was evaluated at 480 nm. The activity of Glutathione peroxidase (GPX, EC 1.11.1.9) was assessed using reduced glutathione (GSH; Cat. No. G4251) with DTNB (Sigma-Aldrich, Cat. No. D8130) as a substrate, and the activity was recorded at 480 nm (Paglia and Valentine 1967). Glutathione S-transferase (GST; EC 2.5.1.18) activity was measured at 310 nm using P-nitrobenzylchloride (Sigma-Aldrich, Cat. No. N7880) as substrate according to the method of Habig et al. (1974). GST induces the conjugation reaction with glutathione in the mercapturic acid synthesis. The catalase enzyme (CAT; EC 1.11.1.6) catalyzes H2O2 substrate (Sigma-Aldrich, Cat. No. H1009) into H20. Hydrogen peroxide (Sigma-Aldrich, Cat. No. H1009) was used as the substrate. CAT activity in liver extract was evaluated spectrophotometrically at 240 nm by measuring the rate of H2O2 degradation by catalase. (Xu et al., 1997). Reduced glutathione (GSH) content was determined, and the method utilized metaphosphoric acid (Sigma-Aldrich, Cat. No. 102433) for protein precipitation and 5,5′-dithiobis (2-nitrobenzoic acid) (DTNB; Sigma-Aldrich, Cat. No. D8130) for color development and its density was measured at 412 nm. GSH was determined according to the method of Jollow et al. (1974). Glutathione reductase (GR; EC 1.8.1.7) activity was measured according to Goldberg and Spooner (1983). In which glutathione reductase catalyzes the reduction of oxidized glutathione by NADPH. The activity was recorded at 340 nm, monitoring NADPH Sigma-Aldrich, Cat. No. N7505) oxidation and oxidized glutathione reduction in EDTA phosphate buffer. The activity of xanthine oxidase (XO) was measured using the commercial EnzyChromTM Xanthine Oxidase Assay Kit (Bioassay System; catalogue no. EXOX-100; Barcelona, Spain) according to the manufacturer’s protocol. Total antioxidant capacity (TAC) was assayed according to the manual instructions of the Biodiagnostic Kit (catalogue no. TA 25 13; Biodiagnostic Co., Giza, Egypt).
The levels of inflammatory molecules, including Tumor Necrosis Factor-alpha (TNF-α; catalogue number: ER1393), and Interleukin 6 (IL-6) in the liver extract were estimated according to instructions described in the commercial ELISA kit (Ray Biotech, Canada; ELR-IL6–1-RB). Transforming growth factor (TGF-β), and necrosis factor kappa-B (NF-κB) were assayed by using an ELISA kit (Active Motif co. Carlsbad, CA 92008 USA). While Cyclooxygenase-2 (COX-2; Cat. No.: CSB-E13399) was measured according to the kit instructions purchased from Cusabio Biotech Co, China. All the sample volumes and concentrations were applied as recommended by the manufacturer’s instructions.
Tumor suppressor gene p53 (p53), was assayed by using an ELISA kit (Active Motif co., 1914, Palomar Oaks Way, Suite 150, Carlsbad, CA 92008 USA). Caspase-3 was assayed by using the method according to Gen Script USA Inc (Cat. No. L00289).
The activities of serum and liver aspartate transaminase (AST) were determined using the commercial kit (Biosystems S.A., Barcelona, Spain, Cat. No.12531). Alanine transaminase (ALT) activities were evaluated using the commercial kit (Biosystems S.A., Barcelona, Spain, Cat. No.12533), and alkaline phosphatase (ALP) activities were measured using a commercial kit (Biosystems S.A., Barcelona, Spain, Cat. No.11592). Acid phosphatase (ACP) activities were assessed using a commercial kit (Biosystems S.A., Barcelona, Spain, Cat. No.11548), and lactate dehydrogenase (LDH) activities were analyzed by kit (Biosystems S.A., Barcelona, Spain, Cat. No.11580). The activities of the gamma-glutamyl transaminase (GGT) enzyme were evaluated with a commercial kit (Biosystems S.A., Barcelona, Spain, Cat. No.11584). Stored serum samples were analyzed for total protein (TP; Cat. No.11500) and albumin (Cat. No.12547) using commercial kits from Biosystems S.A., Barcelona, Spain, according to the manufacturer’s instructions. All sample volumes and concentrations were applied as recommended by the manufacturer’s instructions. All assays were performed according to the manufacturer’s instructions, using recommended sample volumes and concentrations.
Liver specimens were fixed in 10% formalin solution for 48 hours, then treated with 70% alcohol and xylene. The samples were divided into slices with a thickness of 3–6 μm using a rotary microtome after being immersed in paraffin. For the histological analysis, the liver slices were stained with Hematoxylin and Eosin (H&E) and examined under a microscope (Scheuer & Chalk, 1986).
Immunohistochemical staining was performed to reveal the expression of p53, Bax, and Bcl2 proteins in the liver. Sections using avidin-biotin complex (ABC) (Elite–ABC, Vector Laboratories, CA, USA; Cat. No. PK-6101). In apoptotic p53 proteins, sections were incubated with an anti-rabbit p53 monoclonal antibody (dilution 1:80; DAKO Japan Co., Ltd., Tokyo, Japan, Cat. No. M3629). In pro-apoptotic Bax expression, liver sections were incubated with Bax polyclonal rabbit antibody (dilution 1:100; Code A3533, Dako Denmark). In anti-apoptotic Bcl2 protein expression, sections were incubated with anti-rabbit Bcl-2 monoclonal antibody (dilution 1:2000; DAKO Japan Co, Ltd, Tokyo, Japan). The slides were counterstained with hematoxylin and cover-slipped using a permanent mounting media, and the prepared sections were examined using a research microscope (Kos and Matkovich, 2014; Tousson, et al., 2014; He, et al., 2016).
The results obtained from biochemical, molecular, and histopathological analyses are summarized in Tables and Figures.
Liver drug metabolizing enzymes (cytochrome P450, cytochrome b5, amidopyrine N-demethylase, aniline 4-hydroxylase, and NADPH cytochrome C-reductase) of male rats treated with ginseng extract, cisplatin, and their combination groups are exhibited in Figure 1. Administration of cisplatin showed a significant (p < 0.05) increase in the activities of liver cytochrome P450, NADPH cytochrome C-reductase, amidopyrine N-demethylase, and aniline 4-hydroxylase, while significantly (p < 0.05) decreased the activity of cytochrome b5 compared to the control values. Ginseng therapy significantly decreased the induction of cytochrome b5 induced by cisplatin alone while significantly improving the activities of liver cytochrome P450, NADPH cytochrome C-reductase, amidopyrine N-demethylase, and aniline 4-hydroxylase compared with cisplatin ( Figure 1).
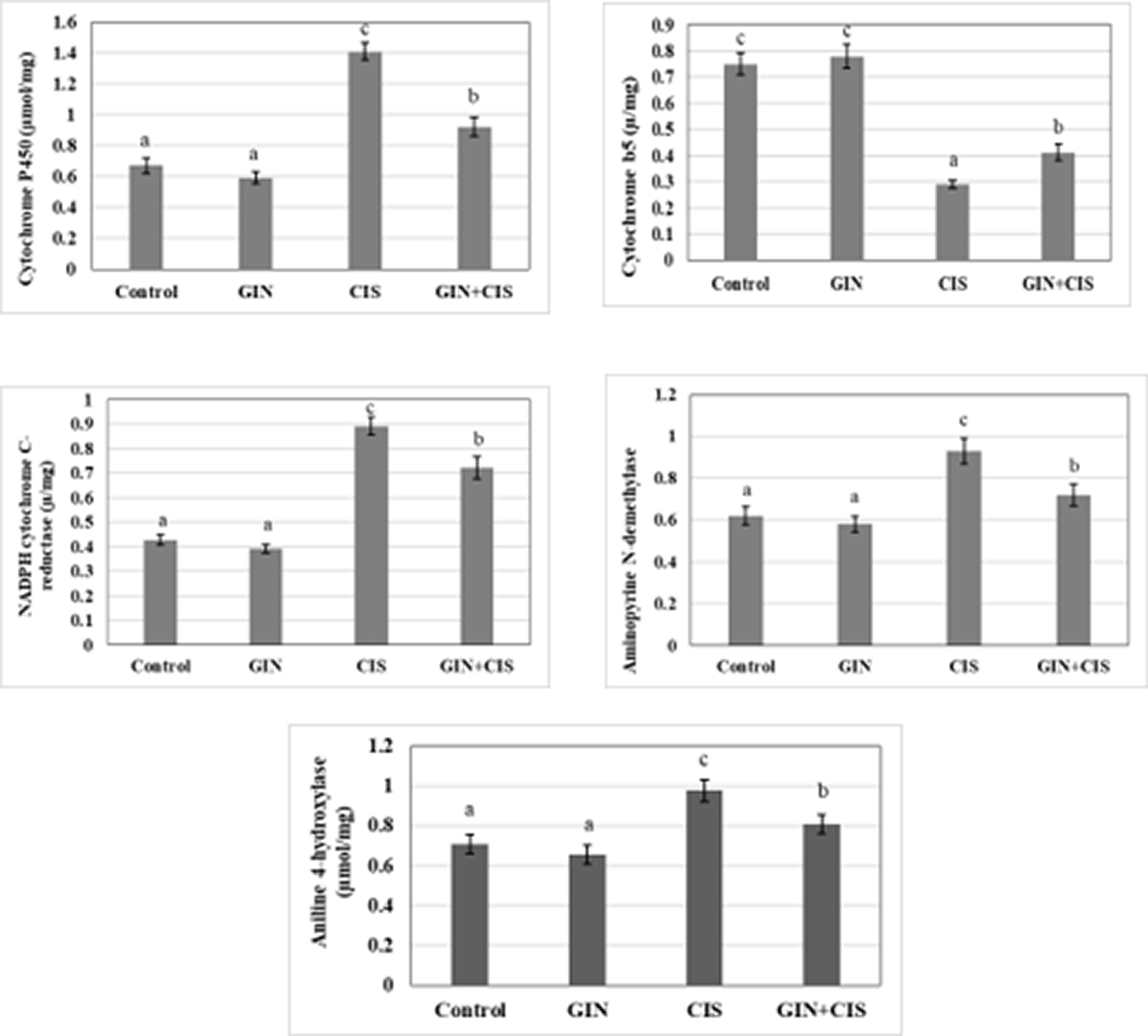
Values are expressed as means ± SD with different superscript letters (a, b, c) of significance at P < 0.05. Mean with letter (a) is the lowest value of significance while mean with (c) is the highest value of significance. Means with the same letters are not significantly different at p < 0.05.
Figure 1 illustrates the relative gene expression levels of hepatic cytochrome P450 enzymes in different experimental groups. Cisplatin treatment significantly altered the expression of several CYP isoforms compared with the control group, whereas co-administration of ginseng partially restored their expression toward normal levels, suggesting a modulatory effect of ginseng on hepatic drug-metabolizing enzymes.
Figure 2 manifested the changes in the gene expression of hepatic phase I enzymes (cytochrome P1A1, cytochrome P2E1, cytochrome P2D6, and cytochrome P3A4) of male rats treated with cisplatin, ginseng, and their combination. Rats treated with cisplatin alone caused a significant (p < 0.05) induction in cytochrome P1A1, cytochrome P2E1, and cytochrome P3A4, while cytochrome P2D6 significantly (p < 0.05) decreased compared to the control group. In comparison to rats treated with cisplatin alone, animals given ginseng with cisplatin in the combination group displayed a significant (p < 0.05) decrease in cytochromes P1A1, cytochrome P2E1, and cytochrome P3A4, while cytochrome P2D6 considerably (p < 0.05) increased compared to the cisplatin group.
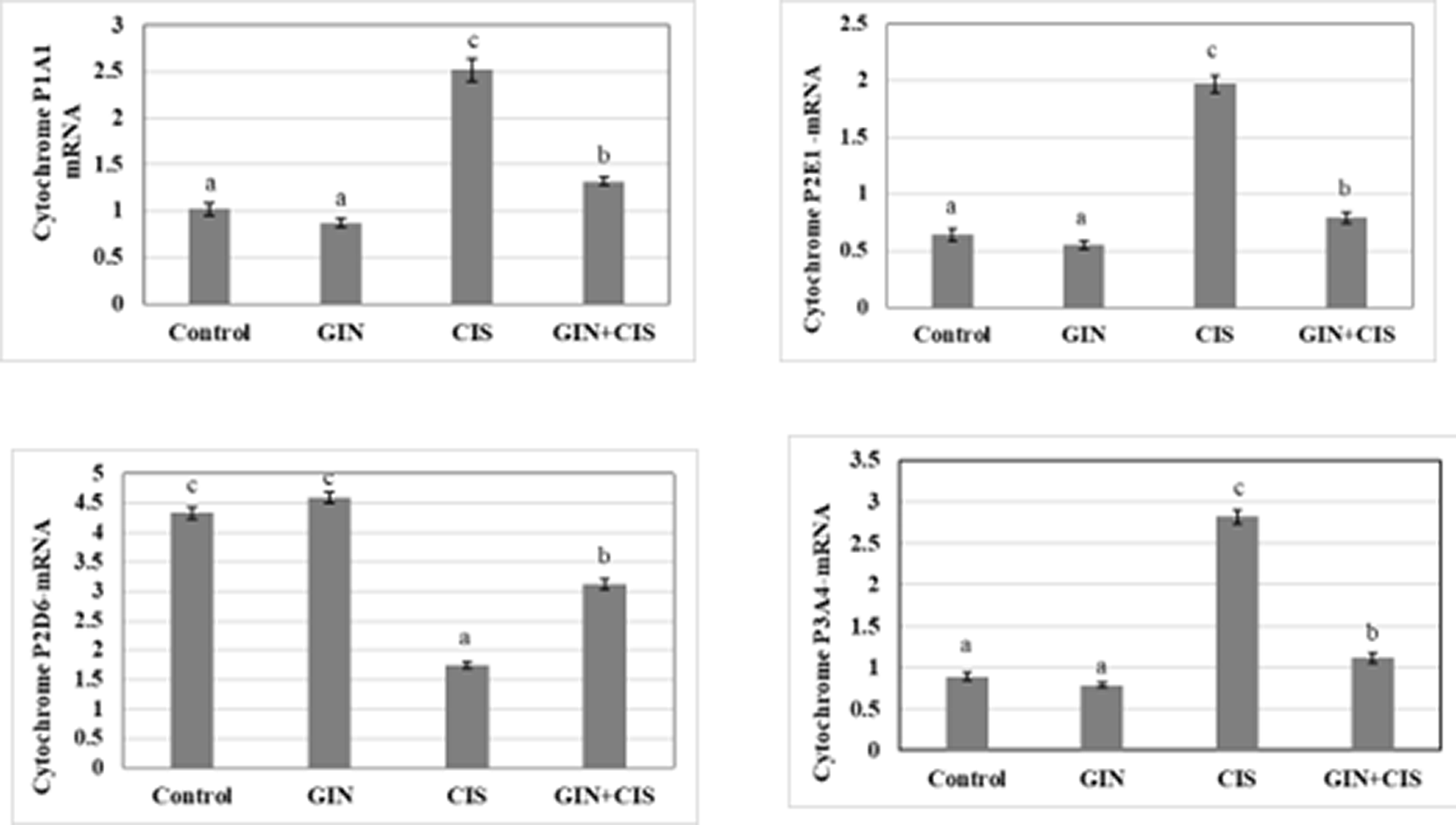
Values are expressed as means ± SD with different superscript letters (a, b, c) of significance at P < 0.05. The mean with letter (a) is the lowest value of significance while the mean with (c) is the highest value of significance. Means with the same letters are not significantly different at p < 0.05.
Figure 2 shows the changes in hepatic inflammatory markers among the experimental groups. Cisplatin administration markedly increased inflammatory mediators compared with the control group, while treatment with ginseng significantly attenuated these elevations, indicating its anti-inflammatory potential.
In the liver tissue, cisplatin triggered the oxidative stress markers. Cisplatin considerably (p < 0.05) induced the prooxidant indicators via the increase in the levels of TBARS, Nitric oxide, and xanthine oxidase, and decreased the activity of antioxidant parameters (GST, CPX, CAT, SOD, GR, GSH, and TAC) compared with the control group. Ginseng alone showed improvement in all antioxidant parameters and decreased the oxidative stress markers. The group treated with ginseng and cisplatin showed an increase in the reduction in all antioxidants and a decrease in the induction of the oxidative stress parameters (TBARS, NO, and XO) compared with cisplatin ( Table 2).
Cisplatin treatment triggered the production of inflammatory molecules. Cisplatin significantly (p < 0.05) enhanced the synthesis of TNF-alpha, IL-6, COX-2, TGF-β, and NF-κB levels. However, ginseng alone showed an insignificant decrease in all inflammatory markers, indicating that it had an anti-inflammatory impact. The presence of ginseng with cisplatin reduced the induction in each of the inflammatory parameters ( Figure 3).
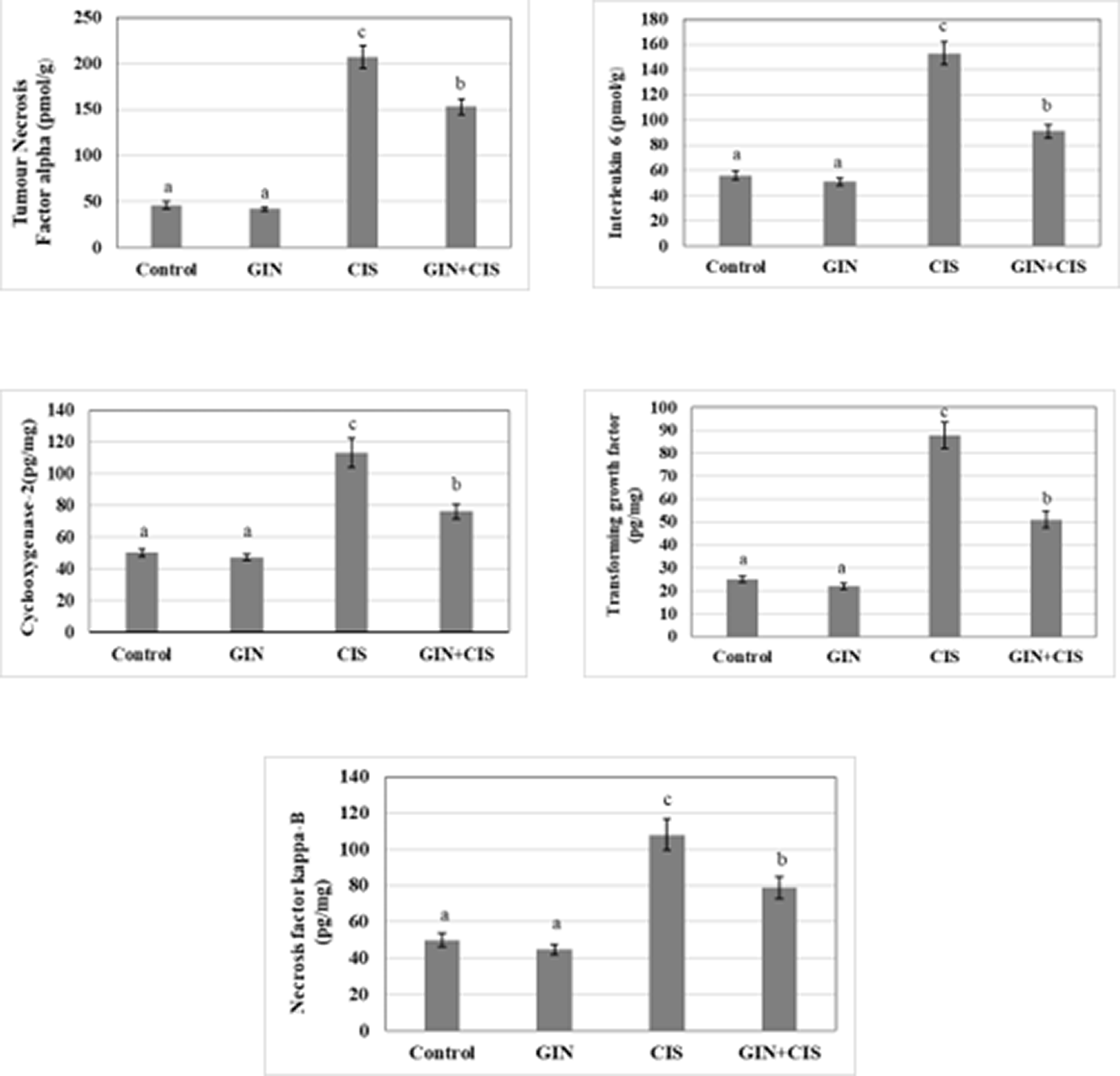
Values are expressed as means ± SD with different superscript letters (a, b, c) of significance at P < 0.05. Mean with letter (a) is the lowest value of significance while mean with (c) is the highest value of significance. Means with the same letters are not significantly different at p < 0.05.
As presented in Figure 3, cisplatin significantly increased apoptotic markers in liver tissues, reflecting enhanced hepatocellular apoptosis. In contrast, co-treatment with ginseng markedly reduced these apoptotic changes, suggesting a protective anti-apoptotic effect.
As shown in Figure 4, exposure to cisplatin remarkably (p < 0.05) induced apoptosis and cell damage via activating the apoptotic factors caspase-3 and the p53 protein. Co-administration of ginseng reduced the levels of liver p53 and caspase-3 compared to the cisplatin group, restoring these aberrant parameter values.
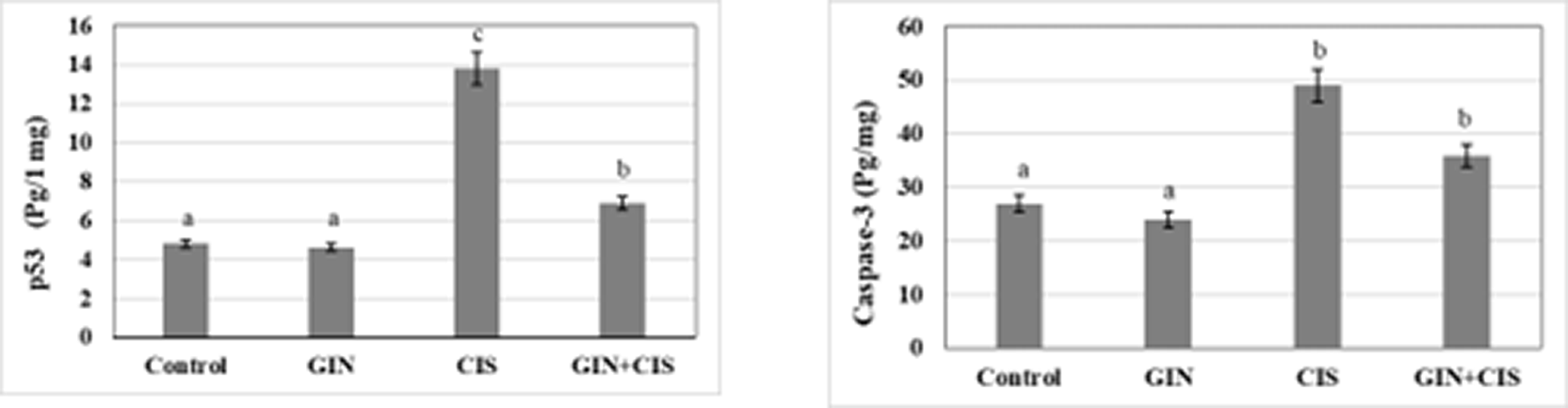
Values are expressed as means ± SD with different superscript letters (a, b, c) of significance at P < 0.05. Mean with letter (a) is the lowest value of significance while mean with (c) is the highest value of significance. Means with the same letters are not significantly different at p < 0.05.
Figure 4 presents representative histopathological sections of liver tissues from the different experimental groups. The control group showed normal hepatic architecture, whereas the cisplatin-treated group exhibited marked structural alterations and hepatocellular damage. Co-administration of ginseng markedly improved hepatic histological features and reduced tissue injury.
Treatment with cisplatin resulted in significant (p < 0.05) elevations in the liver function tests (AST, ALT, LDH, ACP, ALP, and GGT) in serum, and suppressed their activities in liver homogenates, and reduced the levels of serum total protein and albumin. The presence of ginseng with cisplatin significantly improved these enzymes and alleviated the levels of total protein and albumin, reversing the liver toxicity induced by cisplatin ( Tables 3, 4).
Figure 5 (A – D) shows photomicrographs of liver sections stained with HE in different groups. Liver sections of the control group (A) and ginseng group (B) showed the normal structure of hepatocytes and central vein. The liver section of rats treated with cisplatin (C) revealed mild to moderate vacuolated hepatocytes, marked cellular infiltration, and congestion in central veins. The liver section of rats treated with cisplatin with ginseng (D) revealed little tissue injury, with only mild cellular infiltration and no congestion in the central veins.
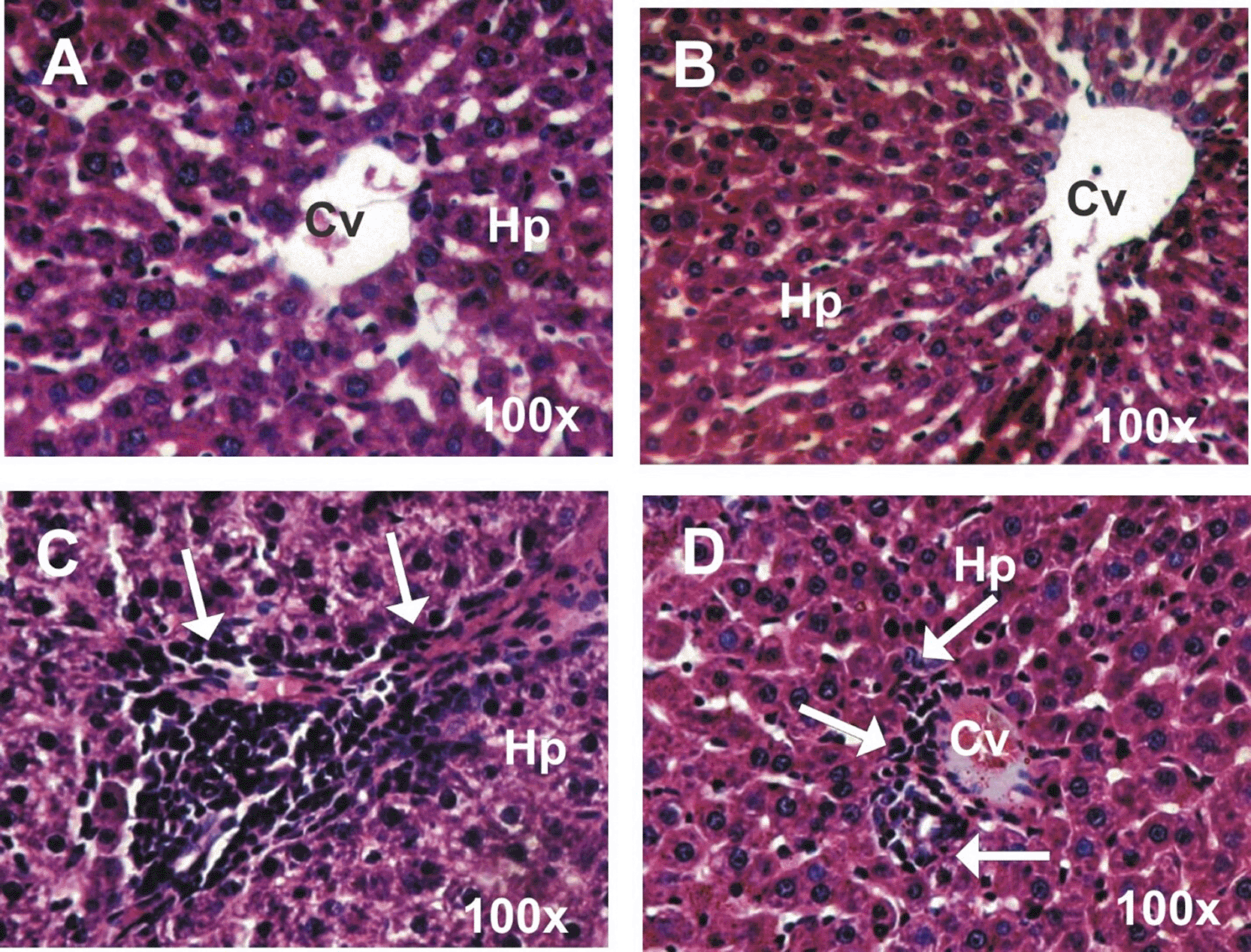
A&B: Liver sections of the control group (A) and ginseng group (B) showing normal structure of hepatocytes (Hp) and central vein (Cv). C: The liver section of the treated rats with cisplatin (C) revealed moderate vacuolated hepatocytes (Hp), marked cellular infiltration (arrows), and congestion in central veins (Cv). D: Liver section in treated rats with Cisplatin and ginseng (D) revealed a little tissue injury with only mild cellular infiltration (arrows).
Detection and distribution of P53 immunoreactivity in the liver of the different groups were presented in Figure 6. Liver sections of the control and ginseng groups showed faint positive reactions for P53 ( Figures 6A and B, respectively). In contrast, moderate to strong positive reactions for P53 were detected in the liver of treated rats with cisplatin ( Figure 6C). On the other hand, rats treated with cisplatin and ginseng showed mild to moderate positive reactions for P53 in their liver sections. ( Figure 6D).
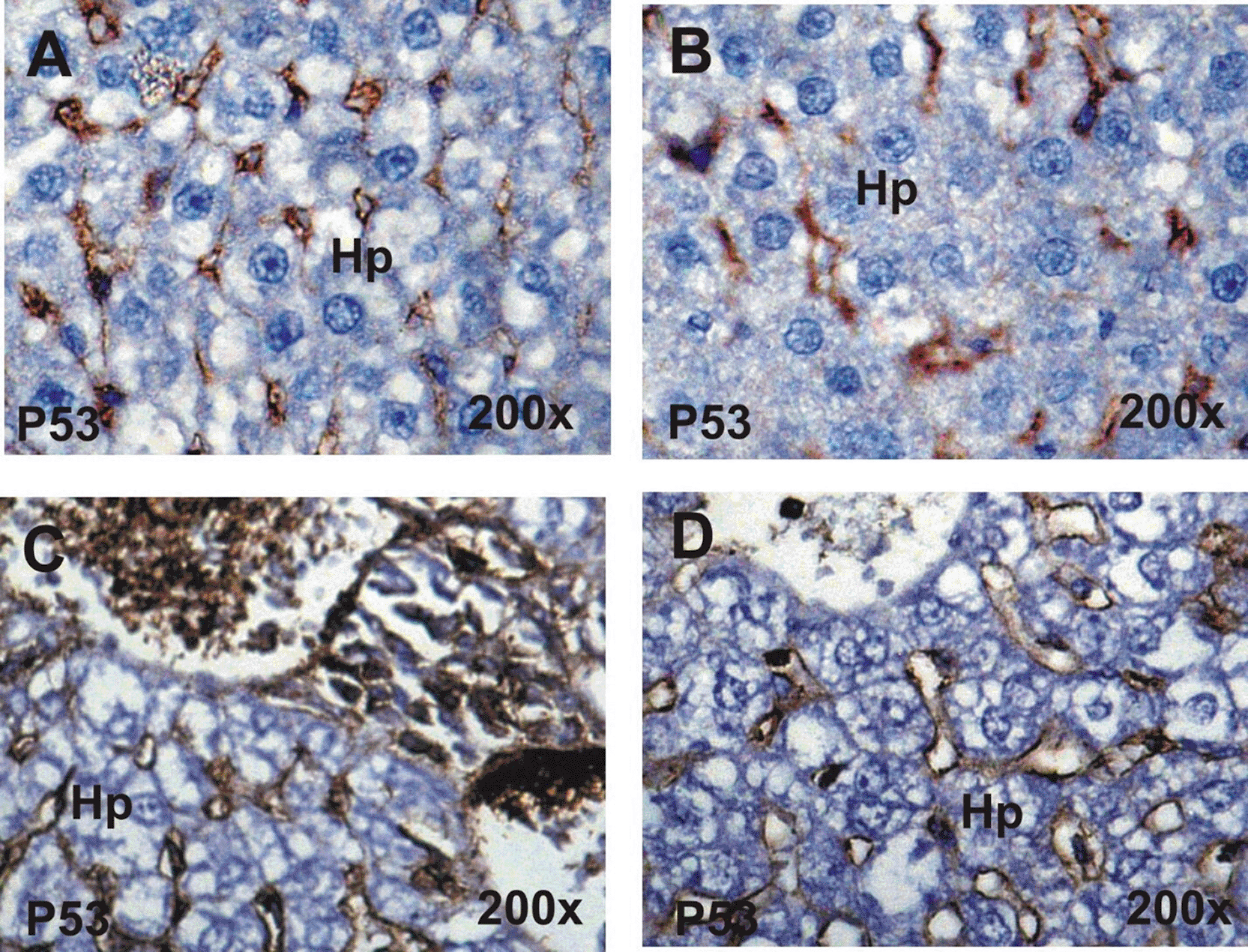
A&B: Faint positive reaction for P53 in control and ginseng groups. C: Moderate to strong positive reaction for P53 in liver section on treated rat with cisplatin. D: Mild to moderate positive reaction for P53 in post-treated rats with cisplatin and ginseng.
The detection and distribution of Bax immunoreactivity in the liver of the different groups were revealed in Figure 7 (A-D). Liver sections in the control and ginseng groups showed faint positive reactions for Bax ( Figures 7A and 8B, respectively). Oppositely, moderate positive reactions for Bax were detected in the liver of treated rats with cisplatin ( Figure 7C). Mild positive reactions for Bax were detected in liver sections in post-treated rats with cisplatin and ginseng ( Figure 7D).
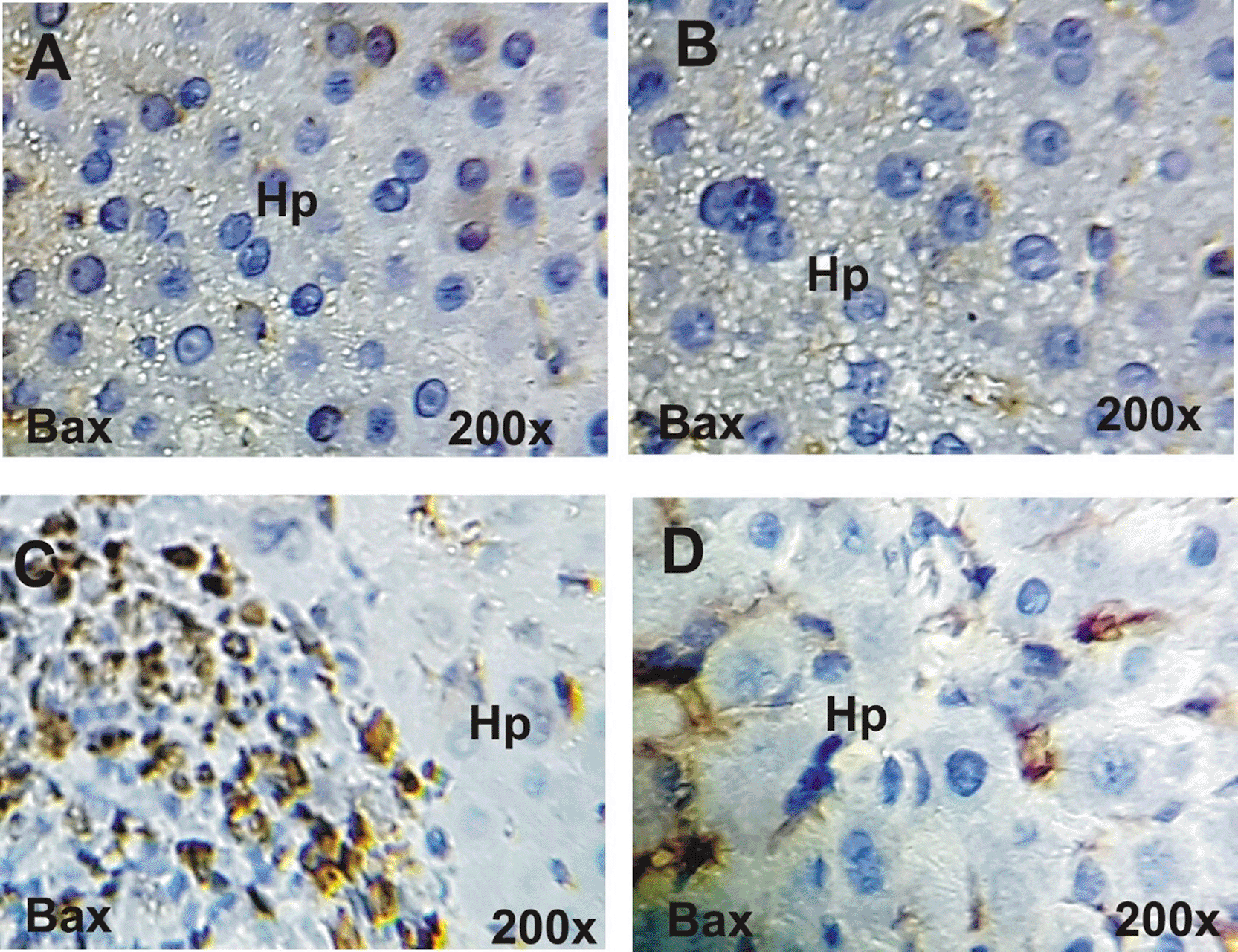
A&B: Faint positive reaction for Bax in control group (A) and ginseng group (B). C: Moderate positive reaction for Bax in liver section on treated rat with cisplatin (C). D: Mild positive reaction for Bax in treated rats with cisplatin and ginseng (D).
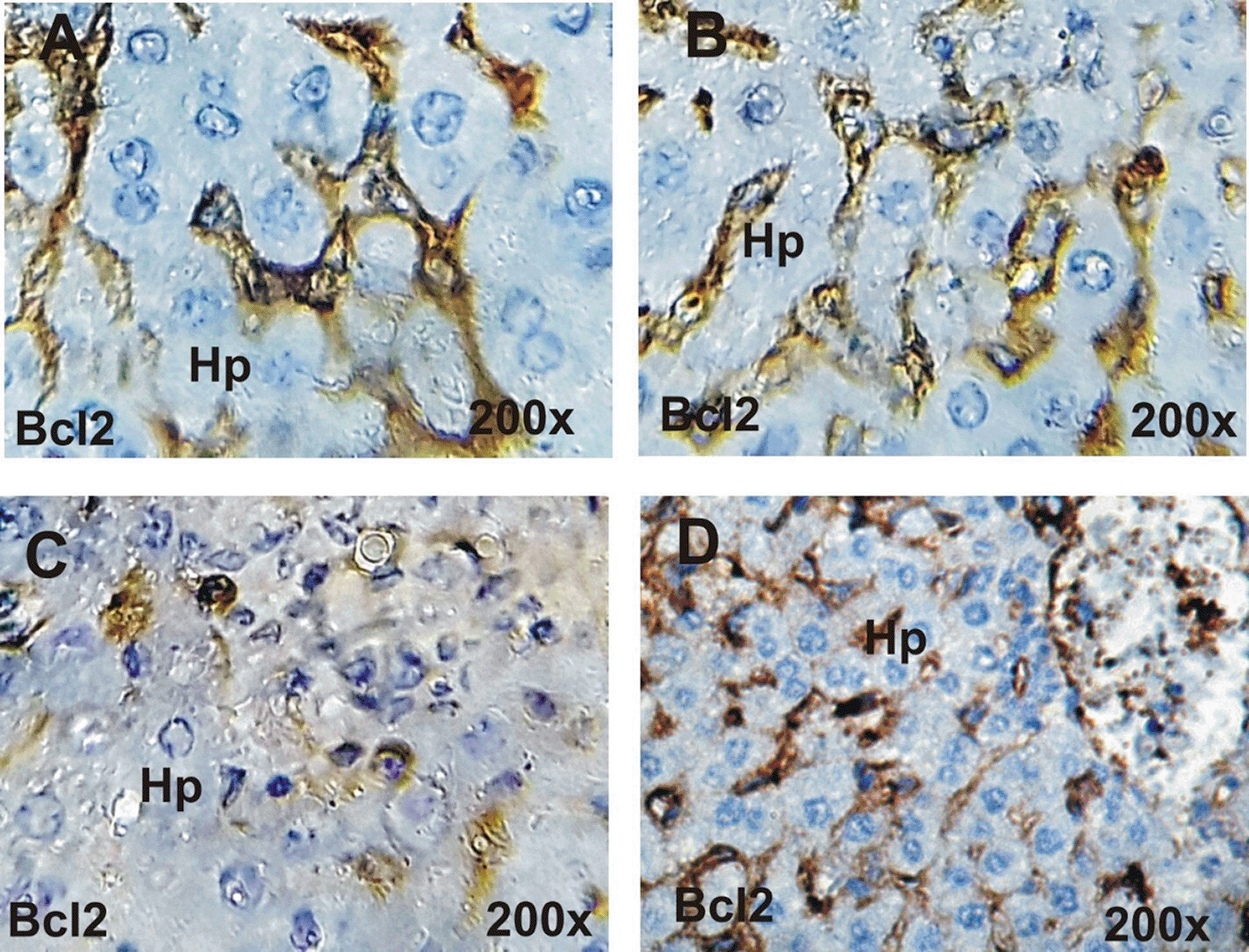
A&B: Strong positive reaction for Bcl2 in control group (A) and ginseng group (B). C: Faint positive reaction for Bcl2 in the liver section on treated rats with Cisplatin (C). D: Moderate positive reaction for Bcl2 in treated rats with Cisplatin and ginseng (D).
Anti-apoptotic Bcl2 protein immunohistochemistry in the liver: The Bcl2 immunoreactivity in the different groups was revealed in Figure 8 (A-D). Liver sections in the control and ginseng groups showed strong positive reactions for Bcl2 ( Figures 8A and 8B, respectively). While faint positive reactions for Bcl2 were detected in the liver of treated rats with cisplatin ( Figure 8C). On the other hand, moderate positive reactions for Bcl2 were detected in liver sections in post-treated rats with cisplatin and ginseng ( Figure 8D).
The present study provides experimental evidence supporting the protective role of ginseng against cisplatin-induced hepatotoxicity.
Cisplatin, a common chemotherapeutic drug, causes liver damage by inducing oxidative stress, inflammation, mitochondrial dysfunction, hyperplasia, portal fibrosis, hepatic cord disturbance, DNA damage, and apoptosis (Taghizadeh et al., 2021; Abbasnezhad et al., 2022). Ginseng has shown promise in protecting the liver from chemotherapy-induced toxicity (Wan et al., 2021; Chen et al., 2022; Dasari et al., 2022). This study aimed to uncover the specific mechanisms by which ginseng mitigates cisplatin-induced liver injury.
Liver drug-metabolizing enzymes (phase I) contain several enzymes, including cytochrome-P450, NADPH-cytochrome C reductase, and cytochrome b5, which metabolize several carcinogens and xenobiotics into less/or more effective intermediates (Hassan, et al., 2012). Cytochrome P450 (CYP) enzymes, found primarily in the liver, metabolize most drugs. These enzymes play a crucial role in drug metabolism and can influence drug-drug interactions (Sonawane et al., 2019; Lee et al., 2002). Cytochrome P450 enzymes can be disrupted by cisplatin, leading to increased ROS production and cellular damage. These disruptions can contribute to cisplatin-induced toxicity and cell death. (Li et al., 2021; Quintanilha et al., 2017; Nan et al., 2019).
The result revealed that cisplatin significantly impaired liver drug-metabolizing enzymes, affecting xenobiotics, carcinogens, and drug metabolism. NADPH-cytochrome c reductase, a key enzyme, stimulates hydroxylation reactions and drug metabolism (Finn et al., 2008). Various compounds can modulate its activity, influencing drug metabolism rates. Cytochrome b5, as a component of this system, plays roles in fatty acid metabolism and apoptosis. Cytochrome b5 levels are associated with drug metabolism, disposition, and therapeutic efficacy. Microsomal amidopyrine N-demethylase and aniline 4-hydroxylase serve as biomarkers for drug metabolites and liver toxicity (Tousson et al., 2020; Li et al., 2021).
Cisplatin disrupts the cytochrome system, leading to oxidative stress and mitochondrial dysfunction. These effects contribute to cisplatin’s toxicity and can cause apoptosis, tissue damage, and liver failure (Tolouian et al., 2023). Therefore, using CYP450 inhibitors may help prevent cisplatin-induced cellular damage.
Ginseng significantly reversed cisplatin’s effects on liver drug-metabolizing enzymes, reducing oxidative stress and restoring enzyme activity. It normalized cytochrome P450, NADPH-cytochrome C-reductase, Amidopyrine N-demethylase, and Aniline 4-hydroxylase levels while increasing cytochrome b5 levels. These changes prevented cisplatin-induced hepatotoxicity and tissue damage.
Cytochrome (CYP) subforums have a critical function in detoxification and are involved in the metabolism of drugs and carcinogens. The overactivity of CYP enzymes can result in anticancer drug resistance, one of the most common issues during chemotherapy (Pathania, et al., 2018). Cisplatin disrupted cytochrome (CYP) enzyme function, leading to drug resistance. It induced CYP1A1, CYP2E1, and CYP3A4, while reduced CYP2D6. Cisplatin induces the overexpression of CYP2E1 enhancing hepatotoxicity via the accumulation of ROS and oxidative stress in vitro and in vivo. CYP1A1 inhibitors have anticancer effects and prevent cisplatin resistance. (Quintanilha, et al., 2017, Nan et al., 2019; Sonawane et al.,2019).
Ginseng, known for its antioxidant properties, can reverse cisplatin-induced abnormalities in cytochrome P450 enzymes. It reversed all the cisplatin abnormalities in cytochromes P1A1, P2E1, P3A4, and P2D6. This suggests its potential to mitigate cisplatin’s hepatotoxicity. Previous studies have shown ginseng’s ability to modulate cytochrome P450 expression and activity, supporting its potential therapeutic use. Also, licorice and rosmarinic acid can counter the expression of CYP1A1, CYP1A2, and CYP2E1, improving oxidative stress and protecting the liver from cisplatin-induced toxicity (Kim et al., 2016; Man et al., 2020; Zhou et al., 2022).
Cisplatin disrupts the liver’s antioxidant system, leading to oxidative stress and hepatotoxicity. This is evident by increased oxidative stress markers (MAD, NO, and XO) and decreased antioxidant levels (GST, GPX, CAT, SOD, GR TAC, and GSH). Cisplatin’s toxicity is linked to its ability to increase the production of free radicals, such as superoxide anion and hydroxyl radical, which cause oxidative damage and impair antioxidant defences (Fatima et al., 2016; Ma et al., 2017; Nam et al., 2019; Dasari et al., 2022).
Cisplatin disrupted liver biochemical parameters, increasing serum AST, ALT, LDH, ACP, ALP, and GGT while decreasing total protein and albumin. On the other hand, cisplatin suppressed the activities of these enzymes in the hepatocytes, indicating leakage of these enzymes from the liver cells, damaging them, which is confirmed by the histological study ( Figure 5). These changes indicate liver cell damage and impaired protein synthesis (Abdel-Daim et al., 2020). Cisplatin also induces oxidative stress by increasing ROS generation and suppressing antioxidants, contributing to liver injury (Choudhry et al., 2019; Ijaz et al., 2020).
Ginseng’s antioxidant properties effectively counteract cisplatin-induced oxidative stress and liver damage (Xu et al., 2018; Song et al., 2020; Zhou et al., 2020; Abdelfattah-Hassan et al., 2019). It restored antioxidant levels, protected liver tissue, and improved biochemical markers associated with liver function. These findings suggest ginseng’s potential to mitigate cisplatin-induced hepatotoxicity.
Inflammation is an important response in defense mechanisms; excessive induction of inflammatory molecules leads to severe diseases. Therefore, suppression of upregulated inflammatory mediators is pivotal in banning these diseases. Cisplatin induces inflammation by increasing TNF-α, IL-6, IL-1β, COX-2, TGF-β, and NF-κB levels. This, along with oxidative stress, contributes to hepatic damage. Elevated TNF-α, IL-1β, and IL-6 reflect fractions of inflammatory processes and oxidative stress and can serve as early diagnostic markers for cisplatin-induced liver injury (Eid and El-Shitany 2021).
Cisplatin activates NF-κB, leading to increased production of inflammatory cytokines (TNF-α, IL-6, IL-1β) and oxidative stress (Fatima et al., 2016; Salama et al., 2021; Lin, et al., 2022). Ginseng can target NF-κB and reduce inflammation, potentially protecting the liver (Yang et al., 2020; Nam et al., 2019; Domingo et al., 2022). In our findings, ginseng markedly reversed the elevation of TNF-α, IL-6, IL-1β, COX-2, TGF-β, and NF-κB, improving the hepatotoxicity of cisplatin, which may be related to its antioxidant capacity. Ginseng’s anti-inflammatory properties may help mitigate cisplatin-induced liver damage.
Cisplatin induces apoptosis by increasing ROS, activating p53 and caspase-3, and disrupting the Bax/Bcl-2 ratio (Taghizadeh et al., 2021; Abd Rashid et al., 2021). Our findings confirm these effects, showing increased p53 and Bax expression, decreased Bcl-2 expression, and evidence of apoptosis and hepatic cell death (Ma et al., 2017).
Ginseng restored caspase-3 and p53 levels, reduced Bax expression, and enhanced Bcl-2 expression, mitigating apoptosis and preventing liver damage. Its antioxidant properties may contribute to these effects. The histopathological screening revealed good ameliorations in liver structure and confirmed the hepatoprotective effect of ginseng against cisplatin-induced liver injury. Ginseng administration reduces the expression of caspase-3 by alleviating oxidative stress (Jung et al., 2017; Wan et al., 2021; Hu et al., 2019).
Although the present study provides important insights into the hepatoprotective effects of ginseng against cisplatin-induced toxicity, several limitations should be acknowledged. First, the study was conducted using an experimental animal model, which may not fully reflect the complexity of human physiological responses. Second, only a single dose of ginseng and cisplatin was investigated; therefore, dose-response relationships were not evaluated. In addition, the precise molecular pathways underlying the modulation of drug-metabolizing enzymes by ginseng require further mechanistic investigations. Future studies involving different doses, longer treatment durations, and clinical investigations are required to confirm the therapeutic potential of ginseng in patients receiving cisplatin therapy.
Further studies exploring the molecular signaling pathways involved in the hepatoprotective effects of ginseng may provide deeper insights into its potential therapeutic applications.
This study demonstrates that cisplatin induces liver toxicity by disrupting liver drug-metabolizing enzymes, elevating inflammatory markers, and causing oxidative stress. Ginseng administration mitigates these harmful effects effectively, suggesting its potential as a protective agent against cisplatin-induced liver damage. The findings highlight the importance of considering the impact of chemotherapeutic agents on liver function and the potential benefits of complementary therapies like ginseng. Future studies could delve deeper into the specific molecular pathways through which ginseng exerts its protective actions on the liver. Evaluate any potential drug-drug interactions that may occur when ginseng is used in combination with cisplatin or other medications. The present study provides compelling evidence that ginseng exerts a significant hepatoprotective effect against cisplatin-induced liver injury through modulation of drug-metabolizing enzymes and attenuation of oxidative stress, inflammation, and apoptosis. These findings highlight the potential of ginseng as a complementary therapeutic agent to reduce chemotherapy-associated hepatic toxicity and warrant further experimental and clinical investigations.
All experimental procedures followed institutional and national guidelines for the care and use of laboratory animals. The research was conducted according to the Guide for the Care and Use of Laboratory Animals (International Council for Laboratory Animal Science, ICLAS) and was approved by the local ethical guidelines of the Institutional Animal Care & Use Committee (IACUC), Alexandria University, Egypt (AU14–200922–3-3).
During the preparation of this manuscript, the authors utilized the generative AI tool Perplexity at the intermediate editorial stage to assist with phrasing and to explore alternative wording options aimed at enhancing the clarity, fluency, and readability of the text. Following the AI-assisted revisions, the authors thoroughly reviewed, modified, and approved all content to ensure accuracy and integrity.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)