Keywords
respiratory tract infections; antimicrobial resistance; multidrug resistance; Enterobacteriaceae; carbapenem resistance; colistin; ESBL; Syria; conflict-affected healthcare; antimicrobial stewardship
respiratory tract infections; antimicrobial resistance; multidrug resistance; Enterobacteriaceae; carbapenem resistance; colistin; ESBL; Syria; conflict-affected healthcare; antimicrobial stewardship
Respiratory tract infections (RTIs) remain a critical global health issue, mainly in low- and middle-income countries (LMICs). They result in a large amount of suffering and death, particularly among the most vulnerable individuals such as children and the elderly. As per the Global Burden of Disease Study, RTIs remain one of the leading causes of deaths all over the globe, accounting for approximately 2.7 million deaths each year.1,2 Lower respiratory infections (LRIs) have also posed a substantial health burden over the last 30 years, and an observed shift in LRI epidemiology further underlines the urgent need for improvement in health-related policies and interventions.3
The epidemiology of respiratory tract infections (RTIs) has undergone change in recent years due to a number of socio-economic and environmental factors as well as the problem of antimicrobial resistance (AMR). The impact of environmental factors such as air pollution4 and the COVID-19 pandemic on the activity of the respiratory viruses and the consequent fluctuations in incidence rates have been reported.5 For example, fluctuations in incidence have been reported for the viruses causing laboratory confirmed RSV infections in the time of the COVID-19 pandemic, which poses new challenges to the clinicians and epidemiologists.6 The Global Strategy to Address and Control Respiratory Health Effects of Air Pollution (GOLD) reports 20187 highlighted also the fact that patients with chronic obstructive pulmonary disease (COPD) are at increased risk for developing infections that can precipitate an exacerbation.8
Despite efforts to improve management of respiratory tract infections (RTIs), mortality from RTIs is still reportedly higher in low- and middle-income countries (LMICs) than in high-income countries (HICs) due to many potential factors such as access to healthcare, levels of vaccination coverage, extent of laboratory facilities.7,9 According to WHO, more than half of all RTI-related deaths occur in health systems that are weak and poorly equipped to meet the needs of their population, as noted by the Global Expert Panel on AMR at Respiratory Infections (GEP-ARi). Child pneumonia burden remains a massive global health challenge,10 with children under the age of five in low- and middle-income countries bearing the brunt of this infection. In these settings, children with pneumonia are often malnourished and live in poor air quality environments, increasing their likelihood of developing severe disease.
Antimicrobial Resistance (AMR) has become a major challenge for controlling RTIs. The increase in the consumption of antibiotics to empirically manage RTIs has favored the development and spread of resistant strains of bacteria. This is particularly pronounced in low- and middle-income countries (LMICs) where clinical testing to identify the causative bacteria is frequently not carried out, empiric therapy being preferred due to limitations in diagnostic facilities. An independent systematic literature review shows that more than half of cases of bacterial pneumonia may now be due to antibiotic resistant strains.11 Action to address this new threat to global public health is required.
In addition, novel interventions such as nirsevimab for the prevention of RSV infection have been evaluated in clinical trials and have shown efficacy in reducing hospitalisation.12–16 The assessment of novel immunisation strategies for RTIs17 may also yield beneficial improvements in the management of infections in high-risk groups.
It can be concluded that controlling and reducing the impact of RTIs on global health is a complex issue that must be dealt with through surveillance, prudent use of antimicrobial drugs, and development of new drugs. So, the governments, national and international health organizations, as well as communities are required to work together to implement control measures that result in a significant reduction of morbidity and mortality due to RTIs; especially in low- and middle-income countries. In any case, the studies conducted in recent years show that a control strategy that involves all aspects of RTIs and AMR and is compatible with the emerging trends in global health must be implemented.18,19 Despite this pressing need, no comprehensive microbiological surveillance study addressing the bacterial etiology and antimicrobial resistance patterns of respiratory tract infections has been published from Syria in over a decade, leaving a critical knowledge gap that directly impairs evidence-based empiric prescribing in this conflict-affected setting.
This retrospective cross-sectional study was conducted at Al-Mouwasat University Hospital, a tertiary academic center in Damascus, Syria, over a 24-month period (January 2022–December 2023). Data were extracted from archived microbiology records; no patient intervention was involved.
We included all patients with clinically suspected RTIs who had positive bacterial cultures from respiratory specimens meeting microbiological significance criteria (≥105 CFU/mL for quantitative cultures). Eligible specimens comprised sputum, bronchial lavage, and bronchoalveolar lavage fluid. Only the first isolate per infectious episode was retained to avoid duplication bias. Specimens yielding only commensal flora, negative cultures, or incomplete susceptibility data were excluded.
Specimens were processed per standard protocols. Sputum adequacy was verified by Gram stain (<25 squamous epithelial cells/LPF). Significant isolates were identified using conventional biochemical panels. Mycobacterium tuberculosis (identified by acid-fast staining) was excluded from susceptibility testing; all non-mycobacterial isolates (n = 309) underwent full antimicrobial susceptibility testing (AST).
AST was performed by Kirby-Bauer disk diffusion on Mueller-Hinton agar (supplemented where appropriate) and interpreted per current Clinical and Laboratory Standards Institute (CLSI) breakpoints.20 A 20-agent panel spanning ten antibiotic classes was tested, including beta-lactams, carbapenems, fluoroquinolones, aminoglycosides, glycopeptides, oxazolidinones, and colistin. Isolates with intermediate susceptibility were classified as resistant for analytical purposes. ESBL production was phenotypically confirmed by double-disk synergy testing.
Multidrug resistance (MDR) was defined as acquired non-susceptibility to ≥3 antimicrobial categories, per standardized international criteria.20 Extensive drug resistance (XDR) and pan-drug resistance (PDR) were defined accordingly. The total number of resistant categories per isolate was also calculated.
Demographic, specimen, and microbiological data were manually abstracted and verified. Categorical variables were summarized as frequencies; continuous variables as means (±SD). Pathogen–specimen associations were assessed by chi-square (or Fisher’s exact) tests. Pairwise resistance correlations were evaluated using Spearman’s rank coefficient (ρ), with heatmap visualization. Analyses were performed in Python 3.12 and SPSS v26; p < 0.05 denoted statistical significance.
A total of 349 unique patients presenting with culture-confirmed respiratory tract infections (RTIs) were enrolled over a continuous two-year active surveillance period (January 2022 – December 2023) at Al-Mouwasat University Hospital, a tertiary academic referral center operating under conditions of sustained conflict-related resource constraint in Syria. This cohort, drawn exclusively from microbiologically verified infections at a single institution, provides a clinically representative and ecologically valid snapshot of the prevailing bacterial landscape in a high-burden, low-resource setting. The cohort demonstrated a pronounced male predominance, with 225 male patients (64.5%) versus 124 female patients (35.5%) ( Figure 1), yielding a male-to-female ratio of 1.81:1. This disparity is attributable to a convergence of epidemiological forces characteristic of conflict-affected populations, including heightened male exposure to conflict-associated trauma, the greater prevalence of smoking-related chronic obstructive pulmonary disease, and documented asymmetries in healthcare-seeking behavior within the Syrian socio-demographic context. Importantly, univariate sex-stratified analysis revealed no statistically significant association between patient sex and the isolation of any individual pathogen (χ2 range: 0.001–1.694; all p > 0.05), indicating that the observed gender imbalance reflects background epidemiological patterns rather than pathogen-specific host susceptibility differences.
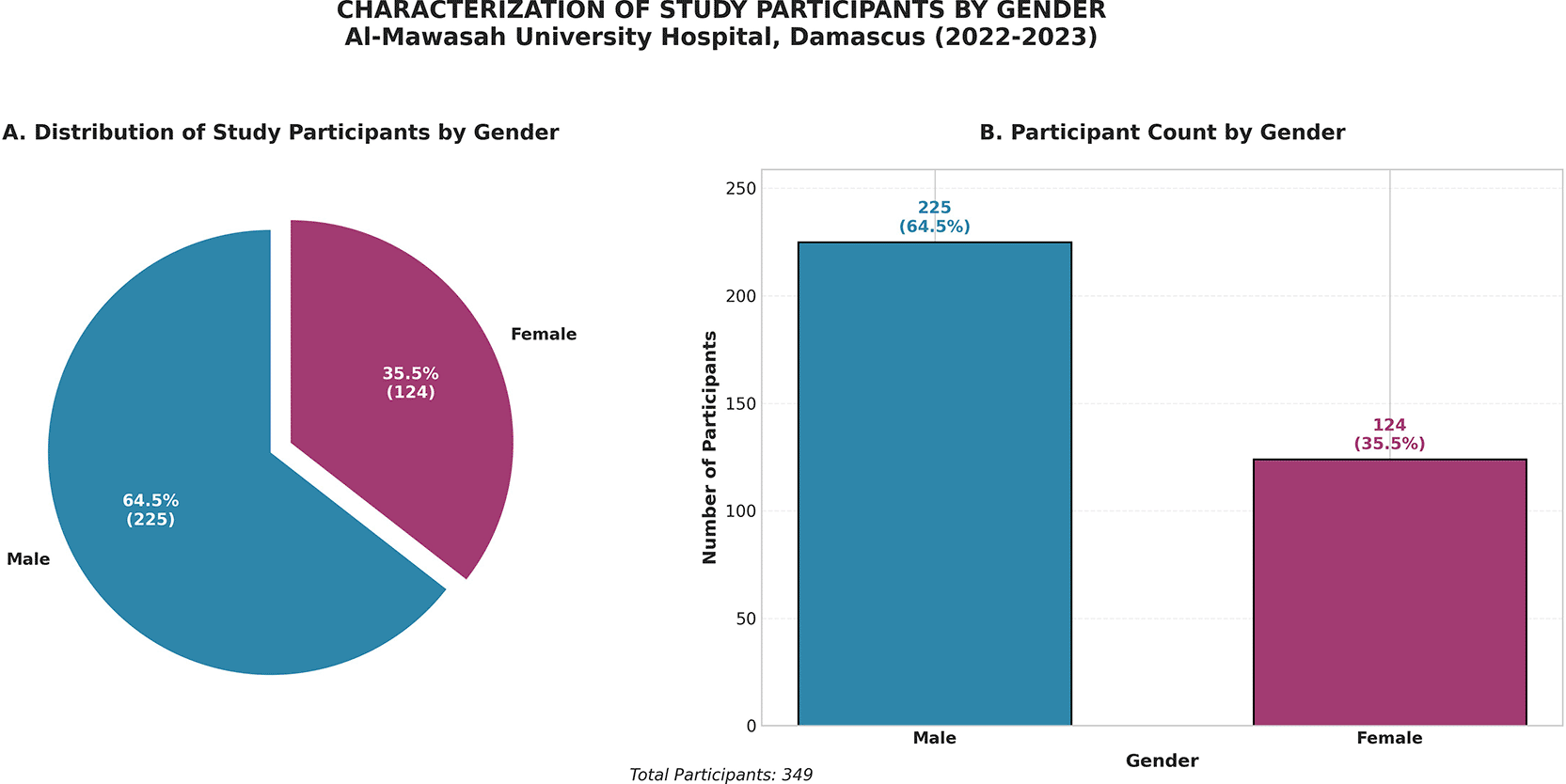
Males comprised 64.5% of the cohort (n = 225), consistent with conflict-associated epidemiological patterns.
Expectorated sputum specimens constituted the predominant sample type, accounting for 286 of 349 samples (82.0%), with the remaining 63 samples (18.0%) consisting of bronchial lavage and bronchoalveolar lavage (BAL) fluid ( Figure 2). This distribution is consistent with the diagnostic conventions of tertiary-care respiratory medicine, wherein non-invasive sputum collection represents the standard first-line approach and invasive lower-airway sampling is preferentially reserved for diagnostically ambiguous presentations or mechanically ventilated patients.
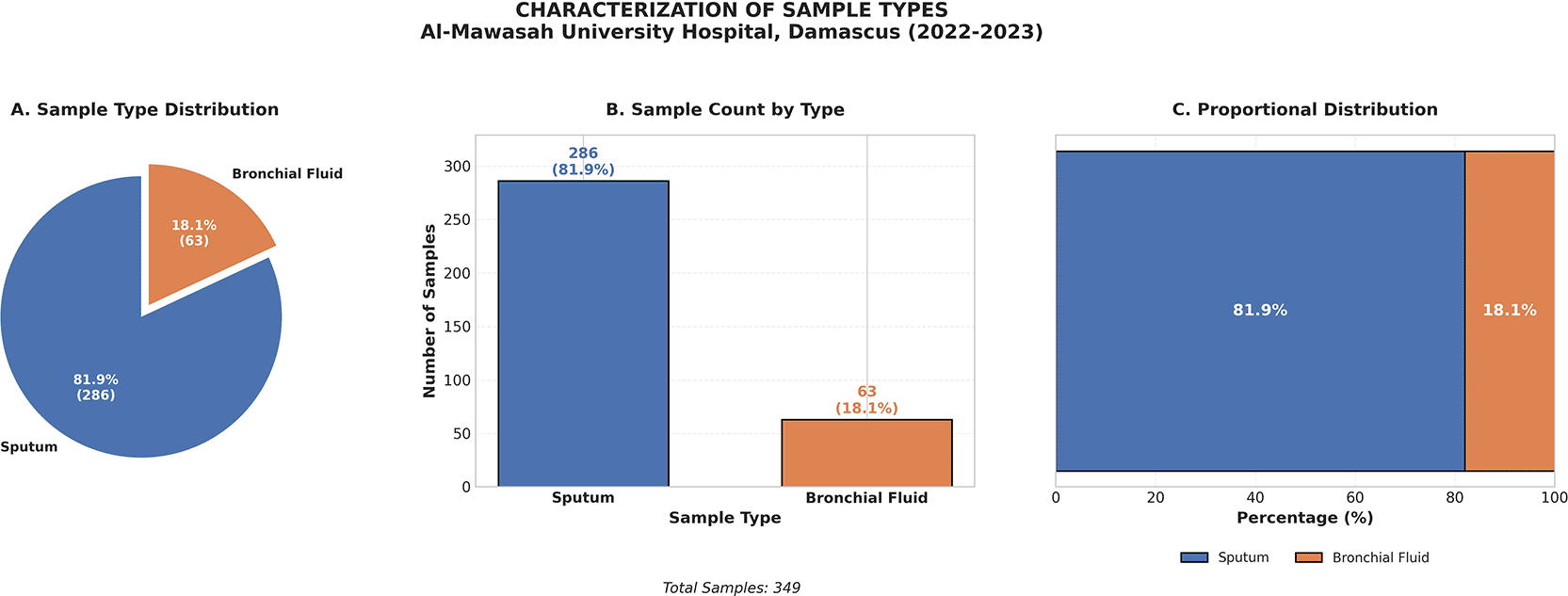
Expectorated sputum (82.0%) represented the primary diagnostic sample; bronchial and bronchoalveolar lavage (18.0%) was reserved for clinically complex cases.
Microbiological analysis of the 349 culture-positive specimens identified twelve distinct bacterial species or genera, with a clear hierarchical pattern dominated by Gram-negative Enterobacteriaceae. The complete pathogen distribution is presented in Table 1 and illustrated in Figures 3 and 4.
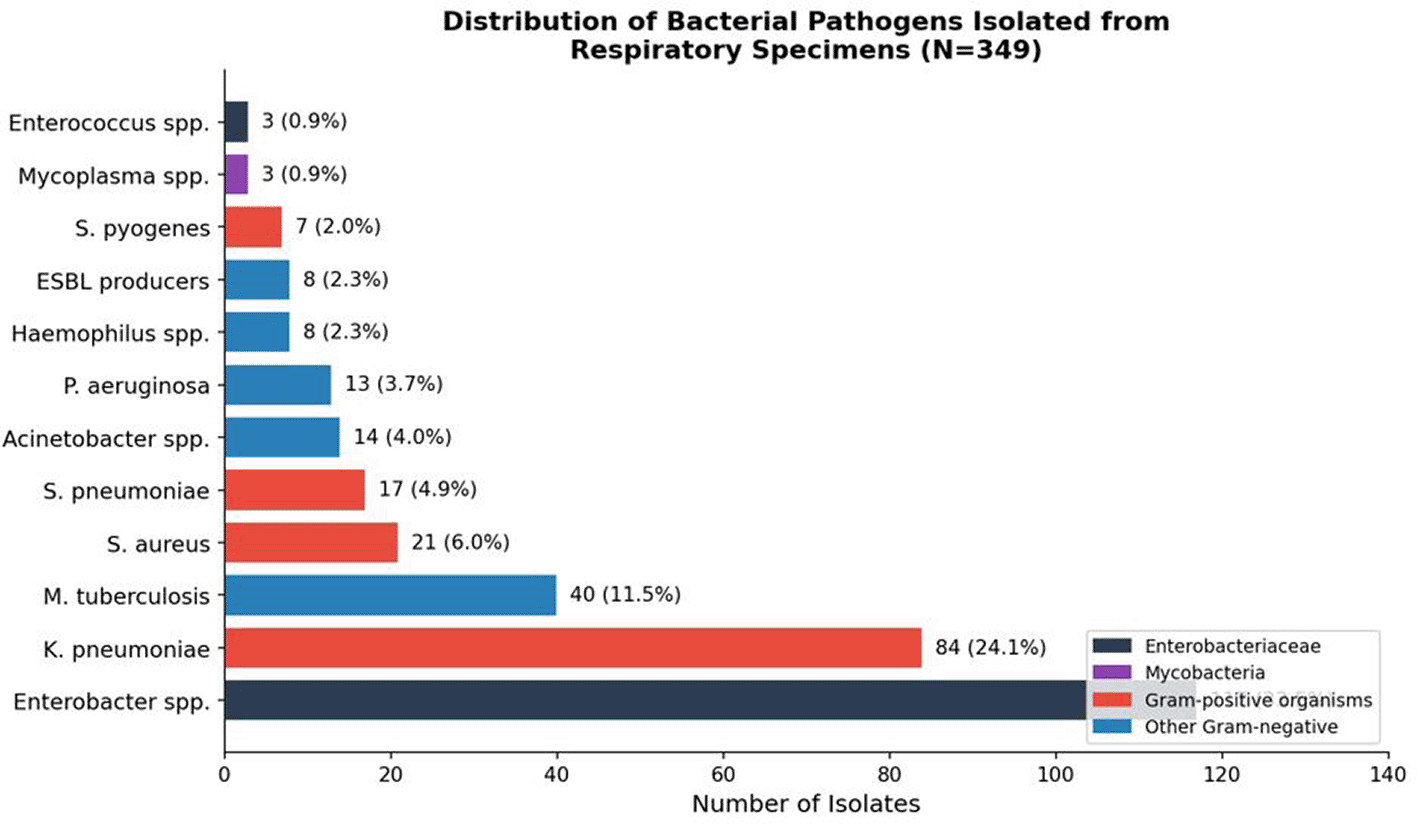
Bars are color-coded by organism class: dark blue, Enterobacteriaceae; purple, Mycobacteria; red, Gram-positive organisms; light blue, other Gram-negative pathogens. Percentages represent proportion of total cohort.
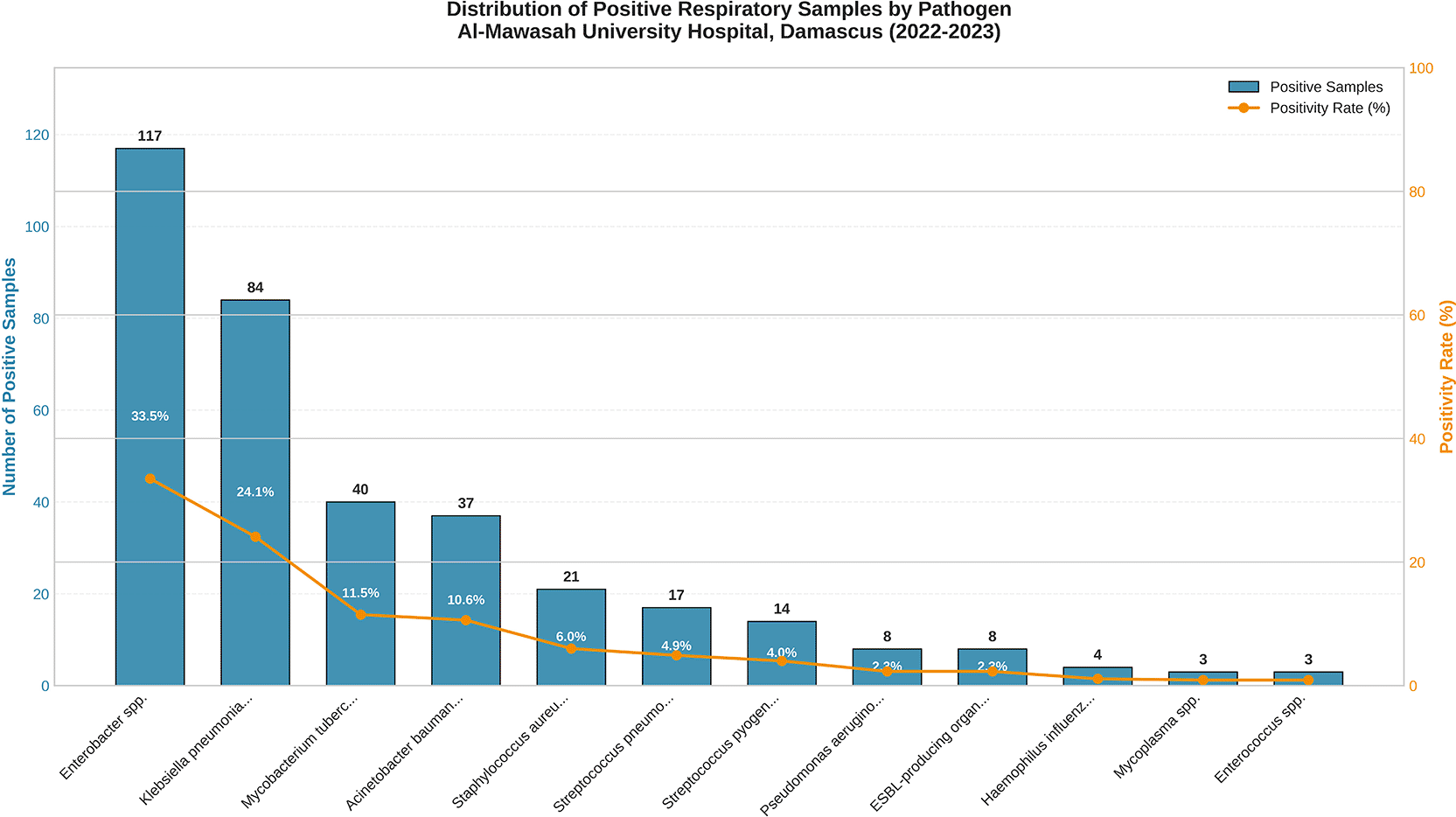
Enterobacteriaceae collectively accounted for 201 isolates (57.6%), a proportion markedly exceeding those reported from high-income country surveillance networks, where atypical and Gram-positive pathogens typically dominate community-acquired pneumonia etiology. Enterobacter spp. emerged as the single most prevalent organism (n = 117; 33.5%), a finding that departs substantially from Western epidemiological norms and reflects the converging pressures of prolonged hospitalization, immunosuppression from comorbid conditions, prior broad-spectrum antibiotic exposure, and the systematic erosion of infection control infrastructure intrinsic to conflict-affected healthcare systems. Klebsiella pneumoniae ranked second (n = 84; 24.1%), a pathogen increasingly recognized as a sentinel species for the global emergence of carbapenem-resistant Enterobacteriaceae. Mycobacterium tuberculosis was identified in 40 specimens (11.5%), a prevalence substantially exceeding global incidence estimates for Syria (~127 per 100,000 population) and reflecting both the referral function of a tertiary-care center and the epidemiological consequences of prolonged conflict, mass population displacement, and the near-total disruption of national tuberculosis control programs. All M. tuberculosis isolates were excluded from antimicrobial susceptibility testing (AST), which was performed exclusively on the remaining 309 non-mycobacterial isolates. Gram-positive pathogens were considerably less prevalent. Staphylococcus aureus (n = 21; 6.0%) and Streptococcus pneumoniae (n = 17; 4.9%) constituted the most frequently isolated Gram-positive species, followed by Streptococcus pyogenes (n = 7; 2.0%) and Enterococcus spp. (n = 3; 0.9%). The relative underrepresentation of S. pneumoniae compared to high-income country data likely reflects the preponderance of healthcare-associated and hospital-acquired infections in this tertiary cohort. Eight isolates (2.3%) were phenotypically confirmed as extended-spectrum beta-lactamase (ESBL) producers,21 a conservative estimate representing a minimum burden given the possibility that routine screening may not have captured all producers.
Antimicrobial susceptibility testing was performed on all 309 non-mycobacterial isolates using a standardized panel of twenty antimicrobial agents spanning ten pharmacological classes, with interpretation according to Clinical and Laboratory Standards Institute (CLSI) breakpoints. The resistance landscape was profoundly concerning across essentially all pathogen–antibiotic combinations, with the exception of colistin, which maintained meaningful—if diminishing—activity against most Gram-negative organisms. A heatmap summarizing overall resistance rates across major pathogens is presented in Figure 5.
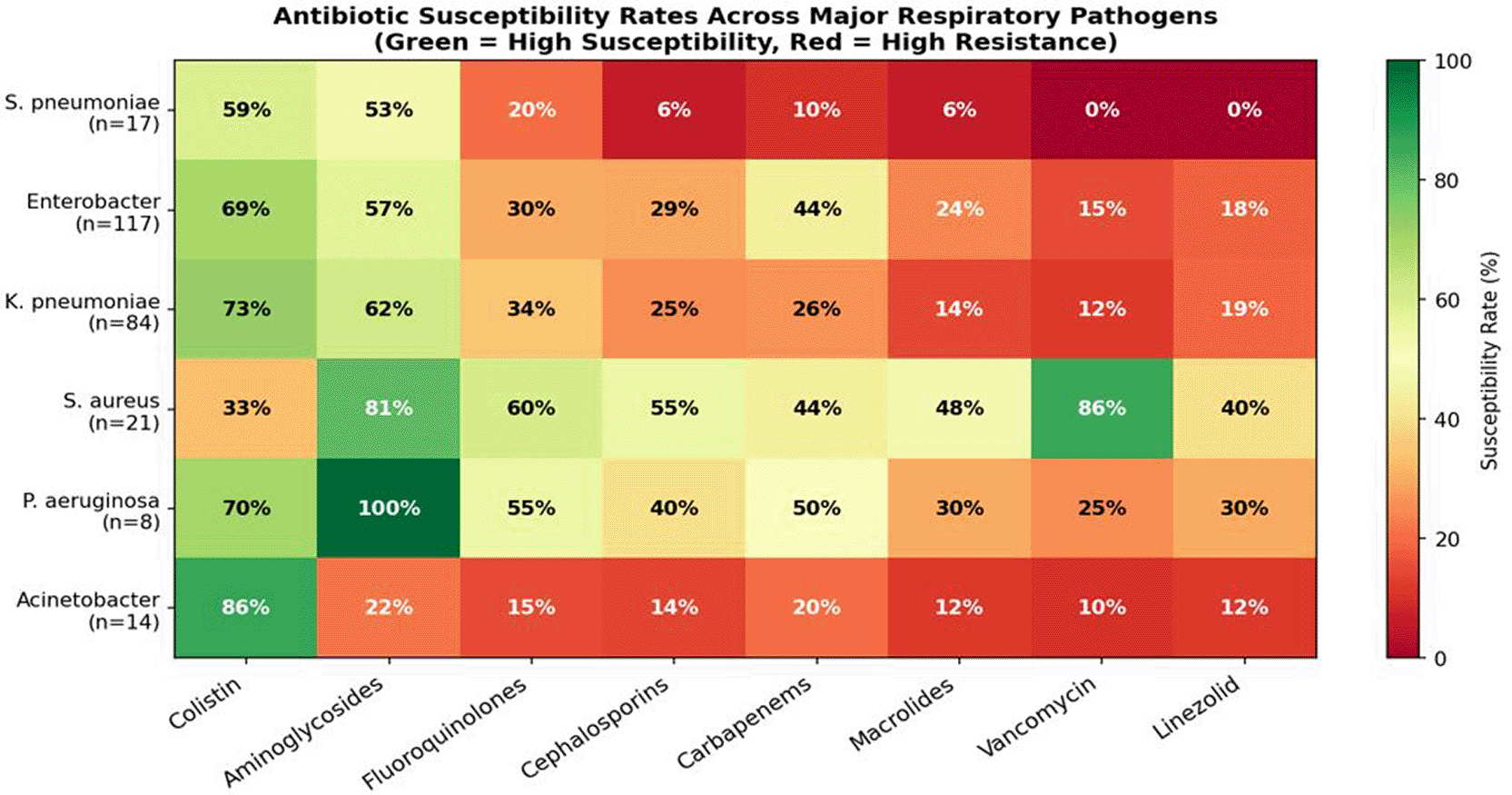
Color intensity reflects resistance rate: dark red indicates near-universal resistance; green indicates retained susceptibility. Colistin (COL) consistently maintains the lowest resistance rates across Gram-negative organisms.
Streptococcus pneumoniae (n = 17)
The susceptibility profile of S. pneumoniae in this cohort represents arguably the most extraordinary single finding of this investigation, constituting a departure from global surveillance benchmarks of exceptional magnitude. Universal resistance was documented to vancomycin (17/17; 100%) ( Table 2; Figure 6) and near-universal resistance to linezolid (16/17; 94.1%) — both agents representing the primary and secondary pharmacological pillars of treatment for beta-lactam-intolerant pneumococcal disease in contemporary practice globally. Macrolide resistance was similarly near-total (azithromycin/erythromycin/clarithromycin: 16/17; 94.1%). The practical consequence of this resistance profile is the therapeutic nullification of the two most established salvage regimens for serious pneumococcal infections in essentially all isolates from this collection.
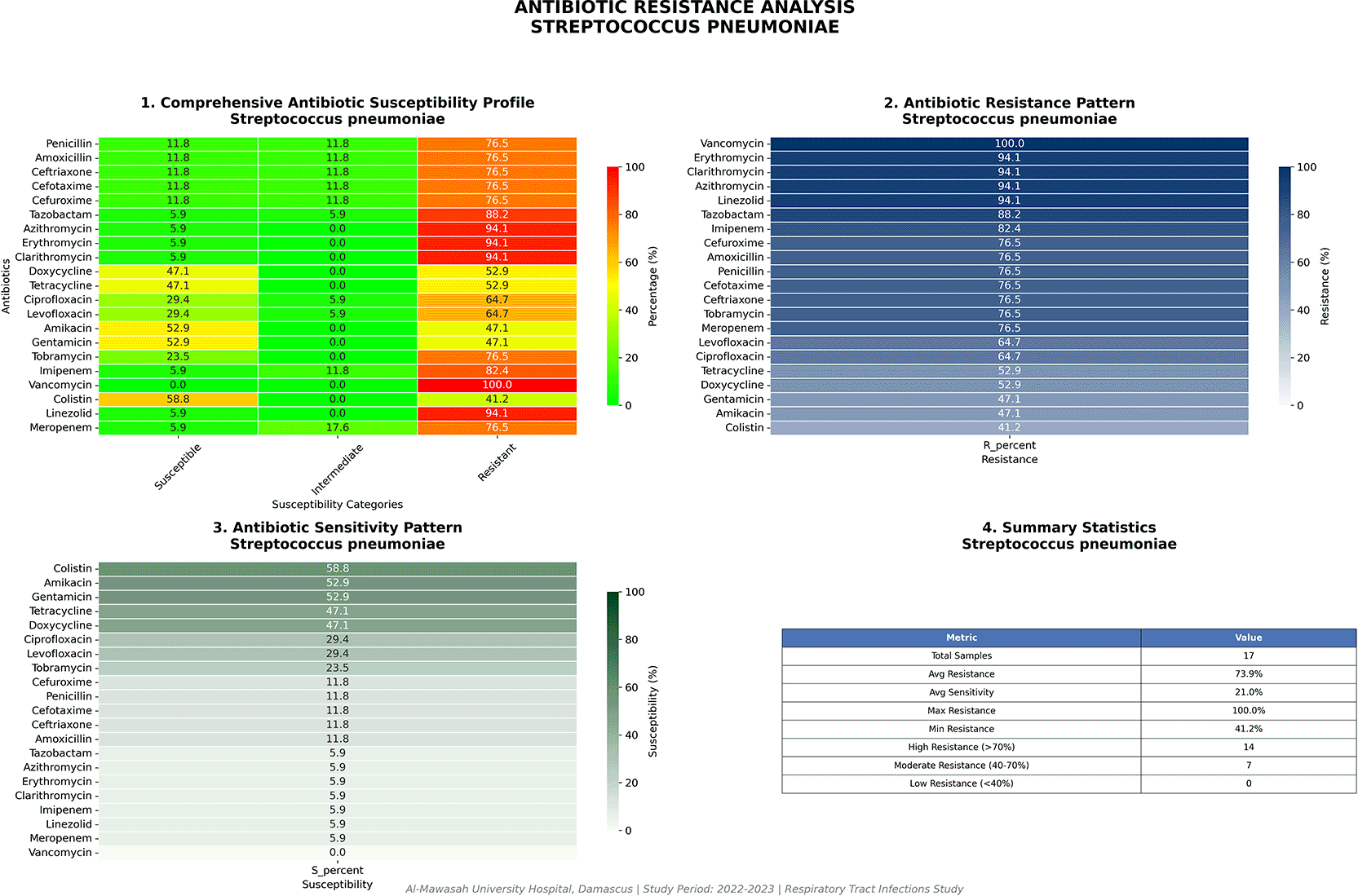
Near-complete loss of activity across vancomycin, linezolid, and macrolides contrasts sharply with global surveillance data.
Residual activity was confined exclusively to colistin (10/17 susceptible; 58.8%) and the aminoglycosides amikacin and gentamicin (9/17; 52.9%) — antibiotic classes with well-recognized pharmacodynamic limitations in the pulmonary compartment and not conventionally indicated as primary therapy for pneumococcal pneumonia. These findings mandate urgent molecular characterization (PCR-based resistance gene profiling; whole-genome sequencing) and confirmatory susceptibility testing by reference broth microdilution, as discussed in detail later.
Enterobacter spp. (n = 117)
As the single most prevalent pathogen in this cohort, the resistance profile of Enterobacter spp. carries disproportionate clinical weight in defining the therapeutic landscape at this institution. The detailed susceptibility data are presented in Table 3, with the corresponding graphical representation in Figure 7. Colistin retained the highest susceptibility rate at 69.2% (81/117), followed by aminoglycosides at 57.3% (67/117). Crucially, even these figures — representing the best available options — indicate that approximately one-third and two-fifths of isolates, respectively, are unresponsive to these last-resort and secondary agents.
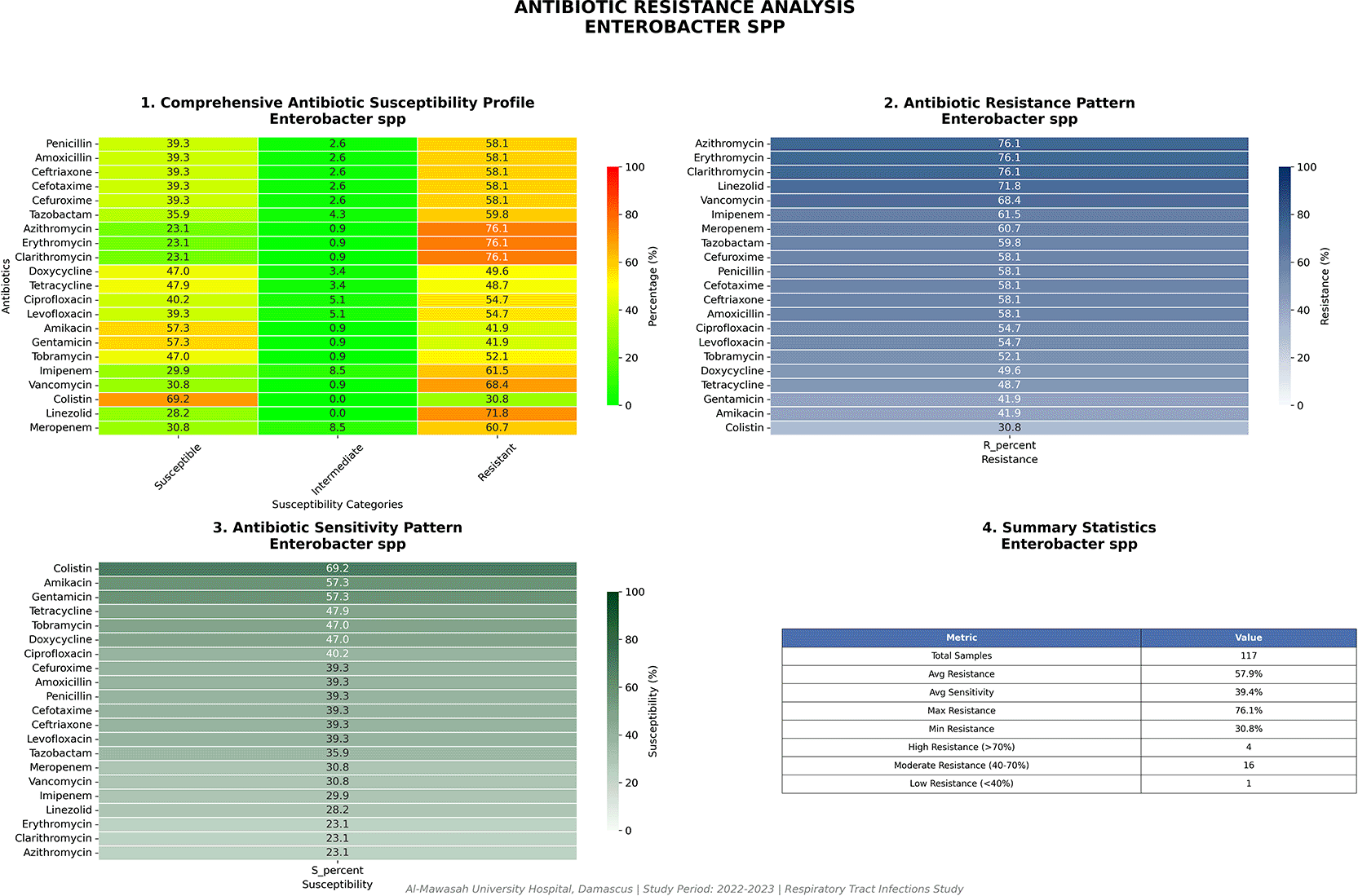
Colistin and aminoglycosides represent the principal active agents; resistance to carbapenems (60.7%) and fluoroquinolones (70.1%) severely constrains therapeutic options.
Resistance rates across clinically critical antibiotic classes were severe: macrolides 76.1% (89/117), fluoroquinolones 70.1% (82/117), carbapenems (meropenem/imipenem) 60.7% (71/117), and beta-lactam combinations 52.1–71.8%. Carbapenem resistance approaching 61% in Enterobacter — a genus already prone to third-generation cephalosporin resistance through chromosomal AmpC beta-lactamase induction — indicates acquisition of higher-order resistance determinants, most plausibly carbapenemase enzymes of class B (metallo-beta-lactamases) or class D (oxacillinases), consistent with regional epidemiological trends.
Klebsiella pneumoniae (n = 84)
K. pneumoniae displayed a resistance trajectory comparable to or exceeding that of Enterobacter spp. Colistin demonstrated the highest susceptibility rate (61/84; 72.6%) ( Table 4; Figure 8), though this leaves more than one-quarter of isolates without an effective last-resort agent. Carbapenem resistance was documented in 62 isolates (73.8%) — a figure approaching rates previously reported only in hyperendemic settings such as certain tertiary hospitals in Greece, Italy, and the Arabian Peninsula, and representing a serious institutional alert. Macrolide resistance reached 85.7% (72/84) and linezolid resistance 81.0% (68/84), the latter likely reflecting intrinsic pharmacodynamic rather than acquired genetic mechanisms given the structural inactivity of oxazolidinones against Gram-negative organisms.
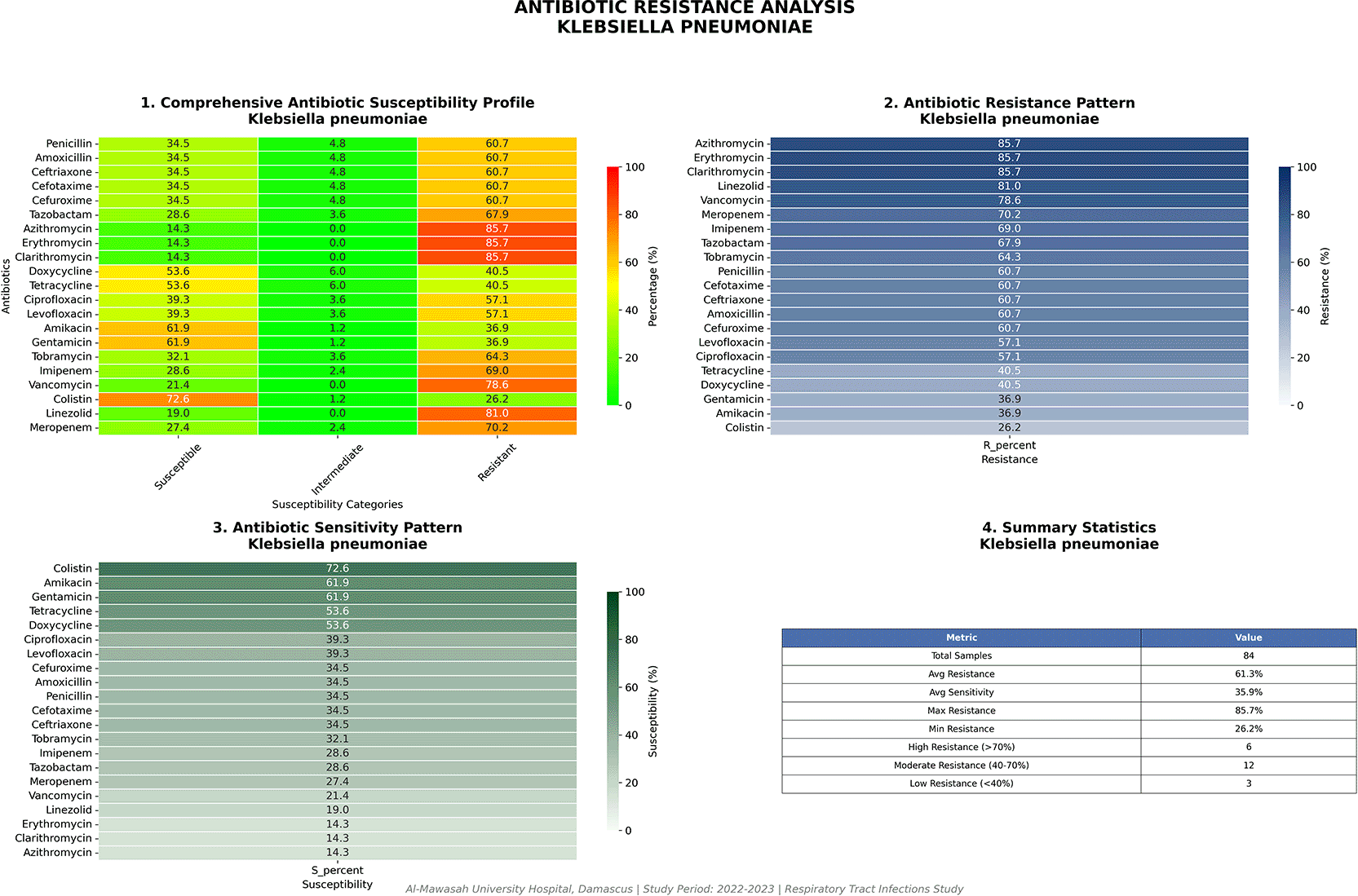
Carbapenem resistance at 73.8% and macrolide resistance at 85.7% define a severely restricted therapeutic window.
Staphylococcus aureus (n = 21)
Against the backdrop of near-universal multidrug resistance among Gram-negative organisms, S. aureus presented a comparatively more favorable susceptibility profile, as summarized in Table 5 and depicted graphically in Figure 9. Vancomycin retained activity in 85.7% of isolates (18/21), and aminoglycoside susceptibility was 81.0% (17/21). Fluoroquinolone susceptibility was approximately 60%, and macrolide resistance was documented in 52.4% (11/21) of isolates. However, the critical and globally alarming finding within this group was the detection of vancomycin resistance in 3 of 21 isolates (14.3%). Whether representing vancomycin-intermediate S. aureus (VISA) or fully vancomycin-resistant S. aureus (VRSA), a resistance rate of 14.3% would be unprecedented in any published institutional surveillance series and mandates immediate confirmatory testing by broth microdilution or E-test, alongside whole-genome sequencing to determine underlying resistance mechanisms (van gene acquisition versus thickened cell-wall-mediated MIC elevation).
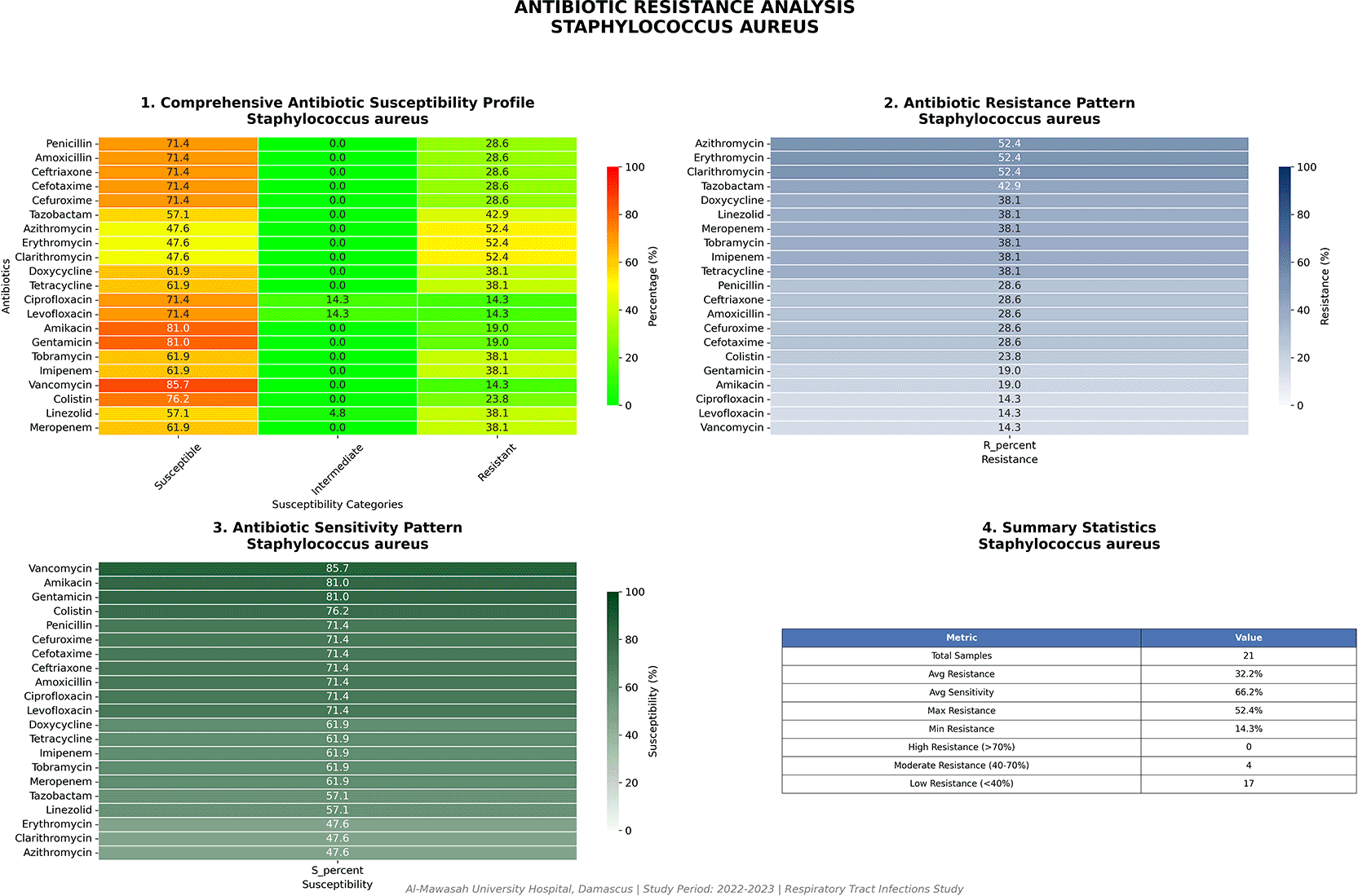
Vancomycin resistance in 14.3% of isolates represents an extraordinary departure from established global benchmarks.
Acinetobacter spp. (n = 14)
Acinetobacter spp. displayed the most extreme resistance phenotype among all Gram-negative pathogens, consistent with this genus’s global reputation for progressive therapeutic intractability. As shown in Table 6 and Figure 10, colistin was the sole agent maintaining reliable activity (12/14; 86.5%), while all other antibiotic classes — including carbapenems — demonstrated susceptibility rates not exceeding 10.8–29.7%. This profile, with an MDR rate of 100%, is consistent with extensively drug-resistant (XDR) or pan-drug-resistant (PDR) phenotypes increasingly reported from conflict-zone and under-resourced healthcare facilities across the Middle East and North Africa region.
| Antibiotic (Class) | Susceptible (n) | Susceptible (%) | Resistant (n) | Resistant (%) |
|---|---|---|---|---|
| Colistin (Polymyxin) | 12 | 86.5 | 2 | 13.5 |
| Other antibiotics (carbapenems, aminoglycosides, fluoroquinolones) | 2–4 | 10.8–29.7 | 10–12 | 70.3–89.2 |
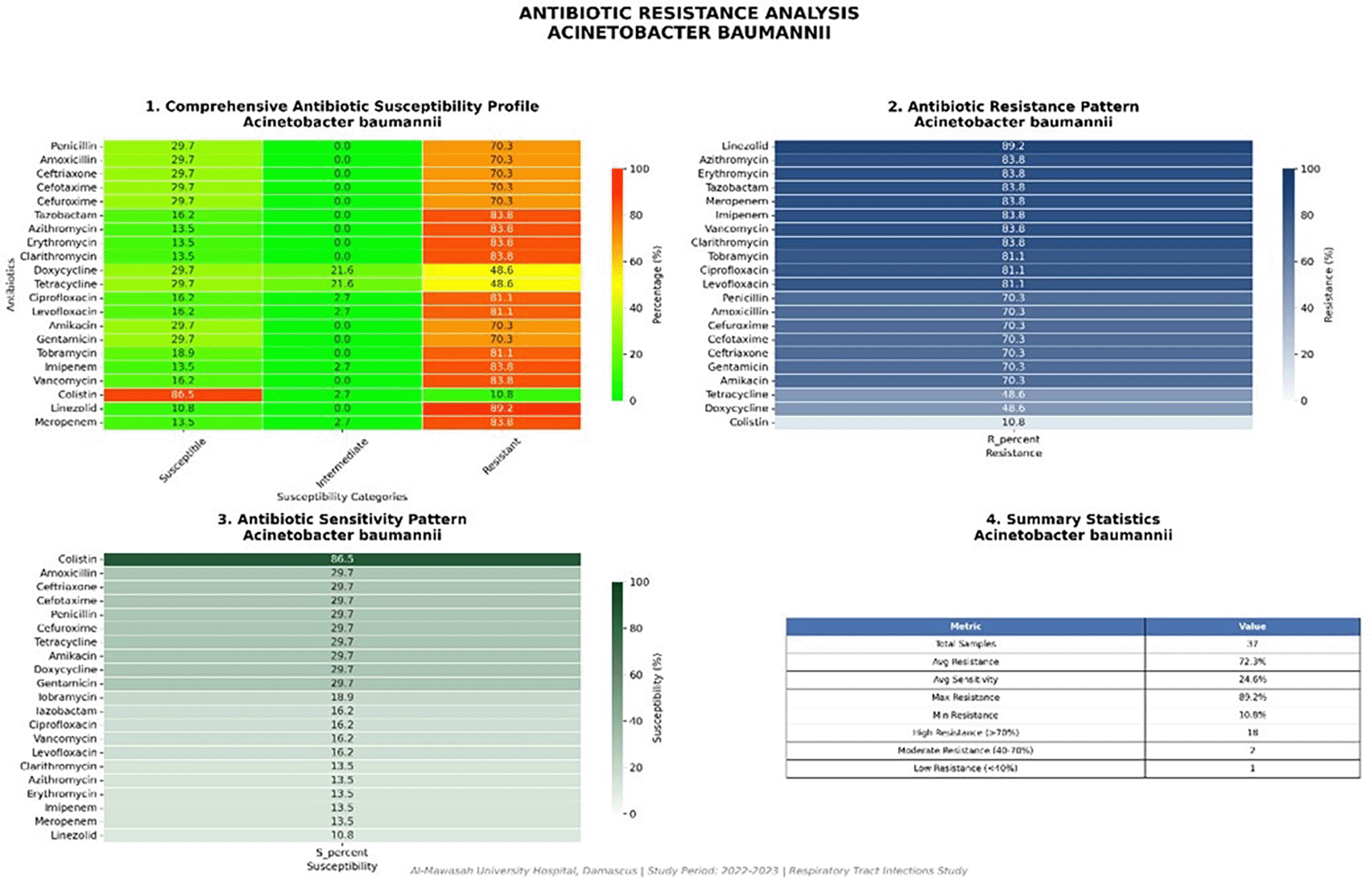
Colistin (86.5% susceptibility) represents the sole reliably active agent; all other tested classes demonstrated susceptibility rates below 30%.
Remaining pathogens
Among Haemophilus spp. (n = 8), universal macrolide resistance (100%) was observed. Moderate susceptibility was retained to tobramycin, imipenem, colistin, and meropenem (50% each), with other antibiotic classes demonstrating susceptibility rates of approximately 25%. All 13 Pseudomonas aeruginosa isolates (100%) were susceptible to aminoglycosides — a noteworthy finding that may reflect the comparatively limited prior therapeutic exposure of this organism to aminoglycosides at this institution. ESBL-producing isolates (n = 8) showed good susceptibility to aminoglycosides (7/8; 87.5%) and colistin (6/8; 75.0%), with near-universal resistance to all beta-lactam classes, as expected given their phenotype. Among the smaller groups, all three Enterococcus spp. isolates were fluoroquinolone-susceptible, and all three Mycoplasma spp. isolates were susceptible to fluoroquinolones, tazobactam, and tobramycin, with expected intrinsic resistance to cell-wall-active beta-lactams. A comparative overview of antibiotic resistance rates across the five key drug classes for all major respiratory pathogens in this cohort is presented in Figure 11, which underscores the consistently low colistin resistance among Gram-negative organisms relative to the near-universal resistance observed for macrolides and carbapenems.
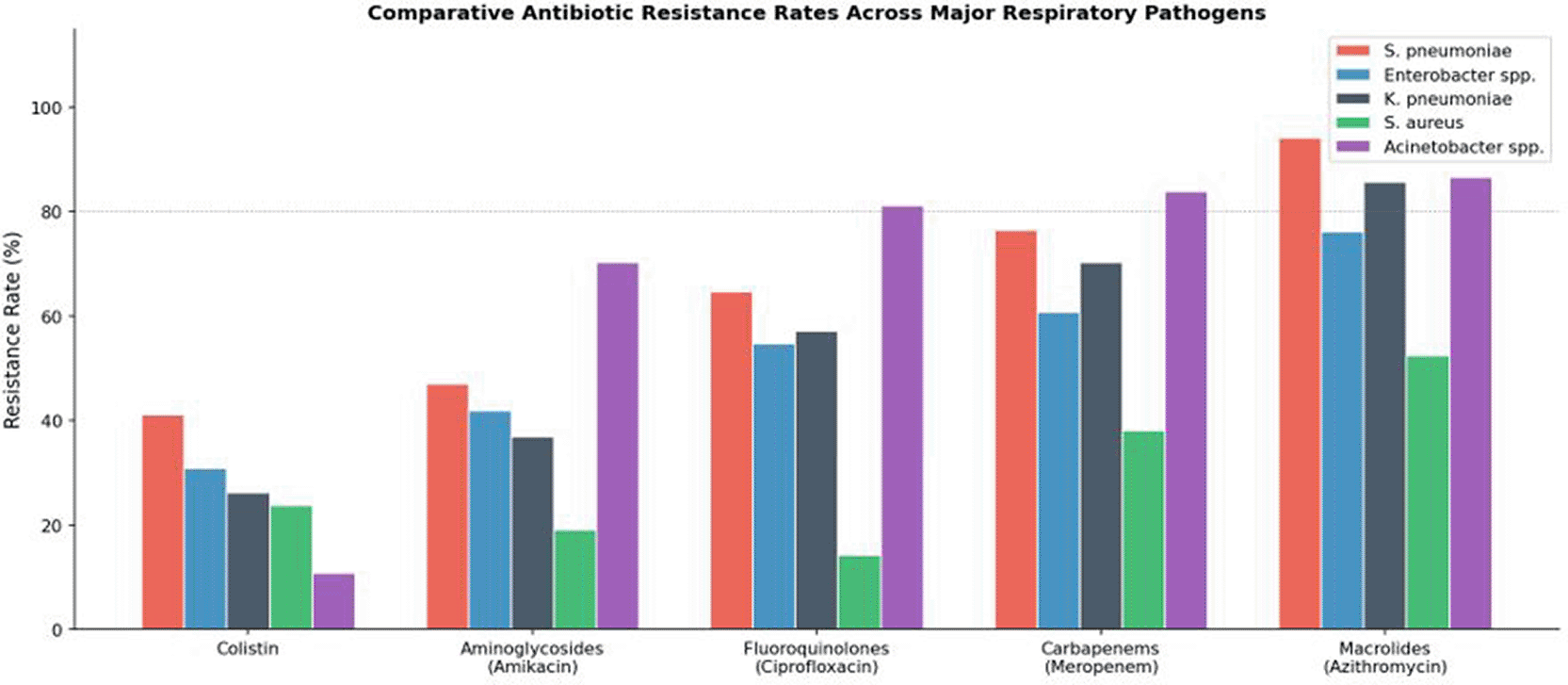
The consistently low resistance rate to colistin across Gram-negative organisms (dark bars) contrasts markedly with near-universal resistance to macrolides and carbapenems. Dashed horizontal line indicates the 80% resistance threshold.
Multidrug resistance (MDR), defined as acquired resistance to at least one agent in three or more distinct antimicrobial categories per international consensus criteria,20 was identified in 270 of 309 evaluable non-mycobacterial isolates (87.4%). This rate substantially exceeds most contemporary surveillance figures from Middle Eastern and North African tertiary-care hospitals — which typically range from 40–70% — and approaches those documented only in the highest-burden institutional settings globally. The mean number of antibiotic classes to which an isolate was resistant was 5.3 (standard deviation ±2.1), spanning a range of 0 to 10 classes. The breadth of resistance was particularly alarming: 81 isolates (26.2%) demonstrated resistance to nine antibiotic classes simultaneously ( Figures 13 and 14), and 16 isolates (5.2%) were resistant to all ten tested classes — a profile meeting criteria for pan-drug resistance (PDR) under international definitions. Conversely, complete susceptibility across all tested antimicrobials was observed in only 39 isolates (12.6%), an uncommonly low proportion that reflects the pervasive and sustained selective antibiotic pressure within this institution. The MDR burden varied substantially by pathogen. Acinetobacter spp. (100%), K. pneumoniae (96.4%), and Enterobacter spp. (91.5%) demonstrated the highest MDR prevalence among quantitatively significant groups. S. pneumoniae and P. aeruginosa both reached 100% MDR when classified by resistance to ≥3 antibiotic classes, while S. aureus (47.6%) and Mycoplasma spp. (33.3%) showed comparatively lower, though still elevated, MDR prevalence. These distributions are detailed in Table 7 and illustrated in Figure 12.
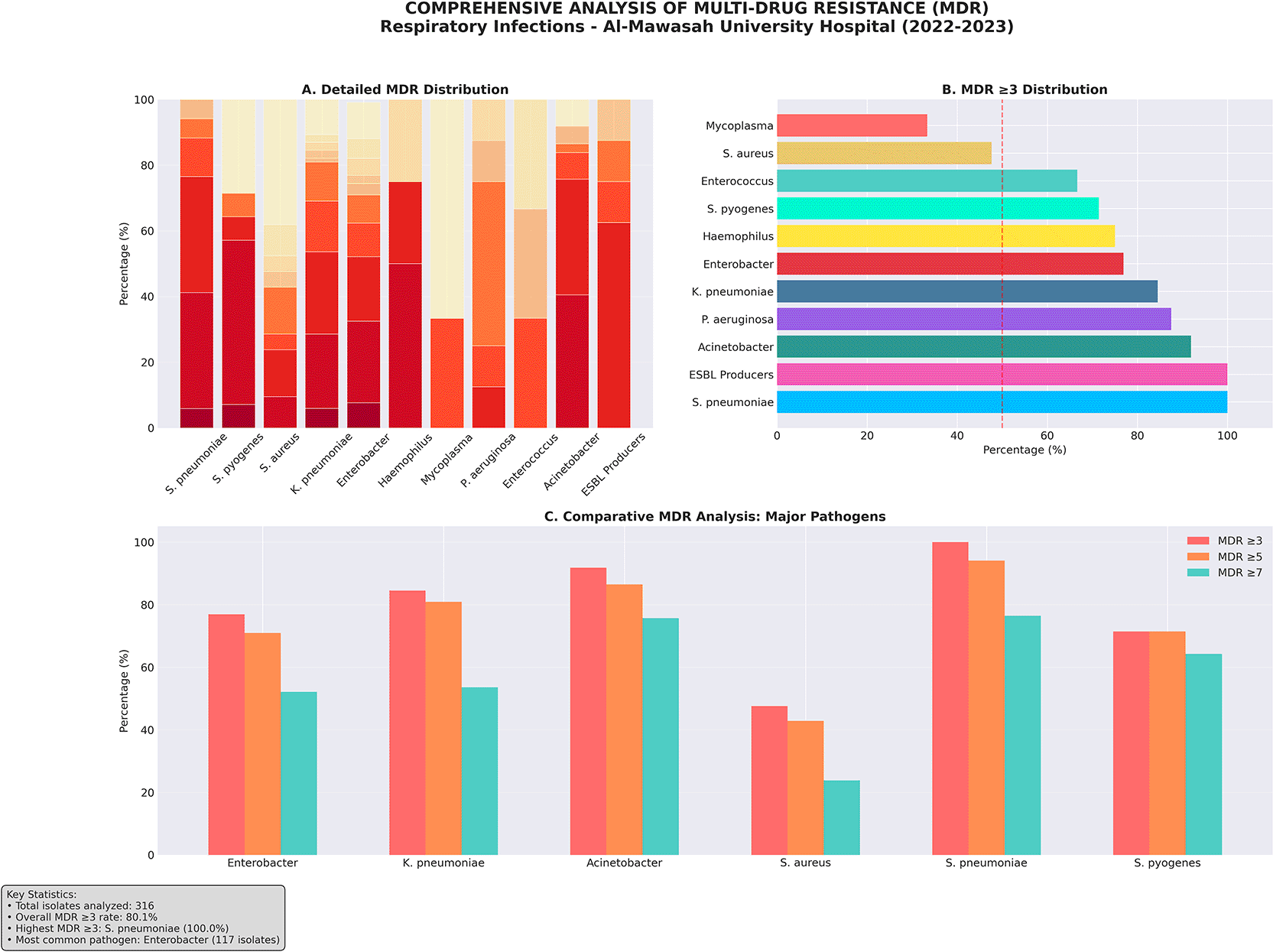
Panel (A): Frequency distribution of number of antibiotic classes to which isolates were resistant (0–10). Panel (B): Comparative MDR rate (%) by bacterial pathogen. Panels (C) and (D): Resistance breadth across all isolates and stratified by pathogen, respectively. Color gradients reflect severity of resistance phenotype.
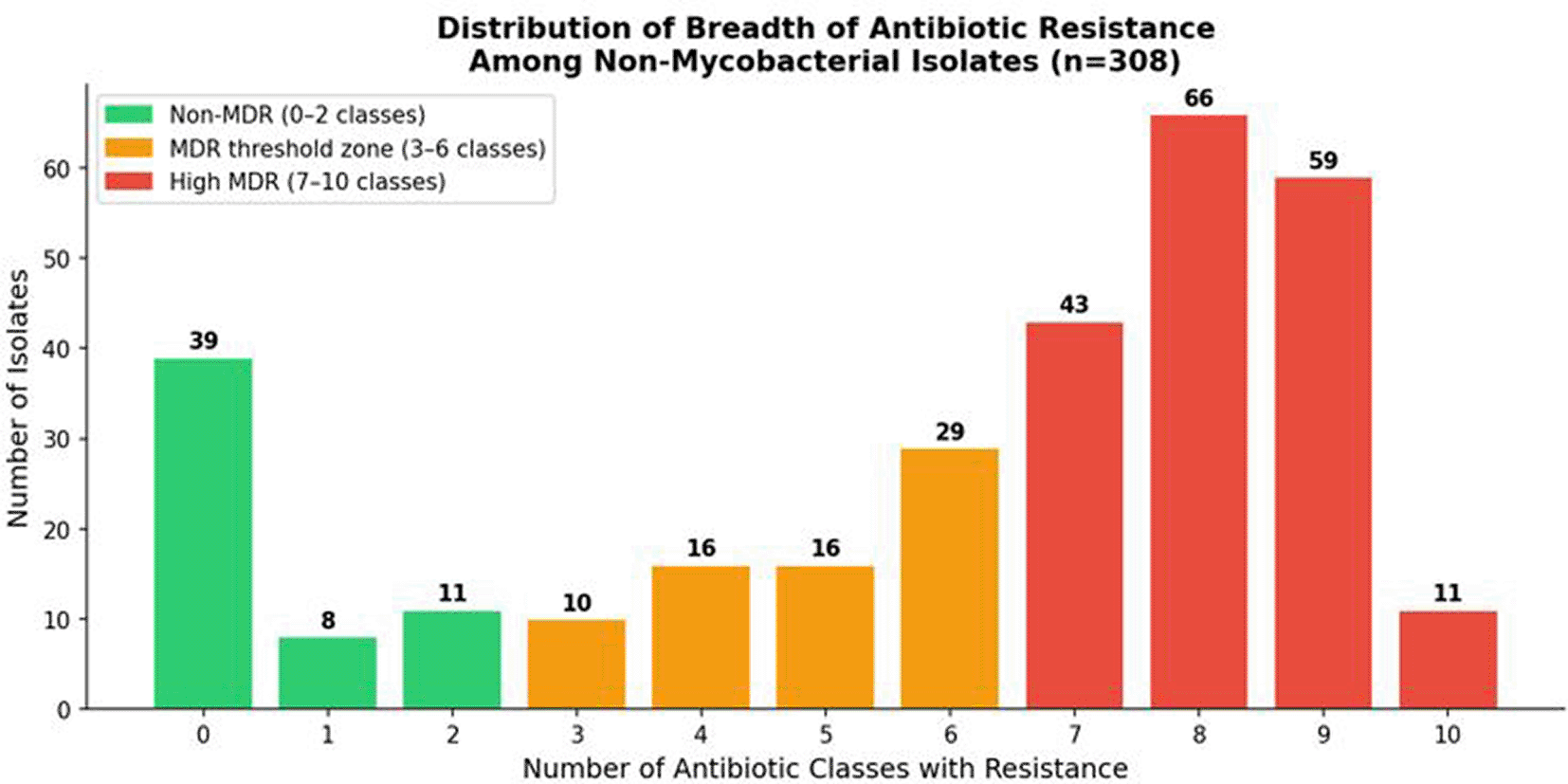
Color coding: green = non-MDR; amber = MDR zone; red = high MDR (≥7 classes). The pronounced rightward skew reflects the extreme degree of resistance selection within this cohort.
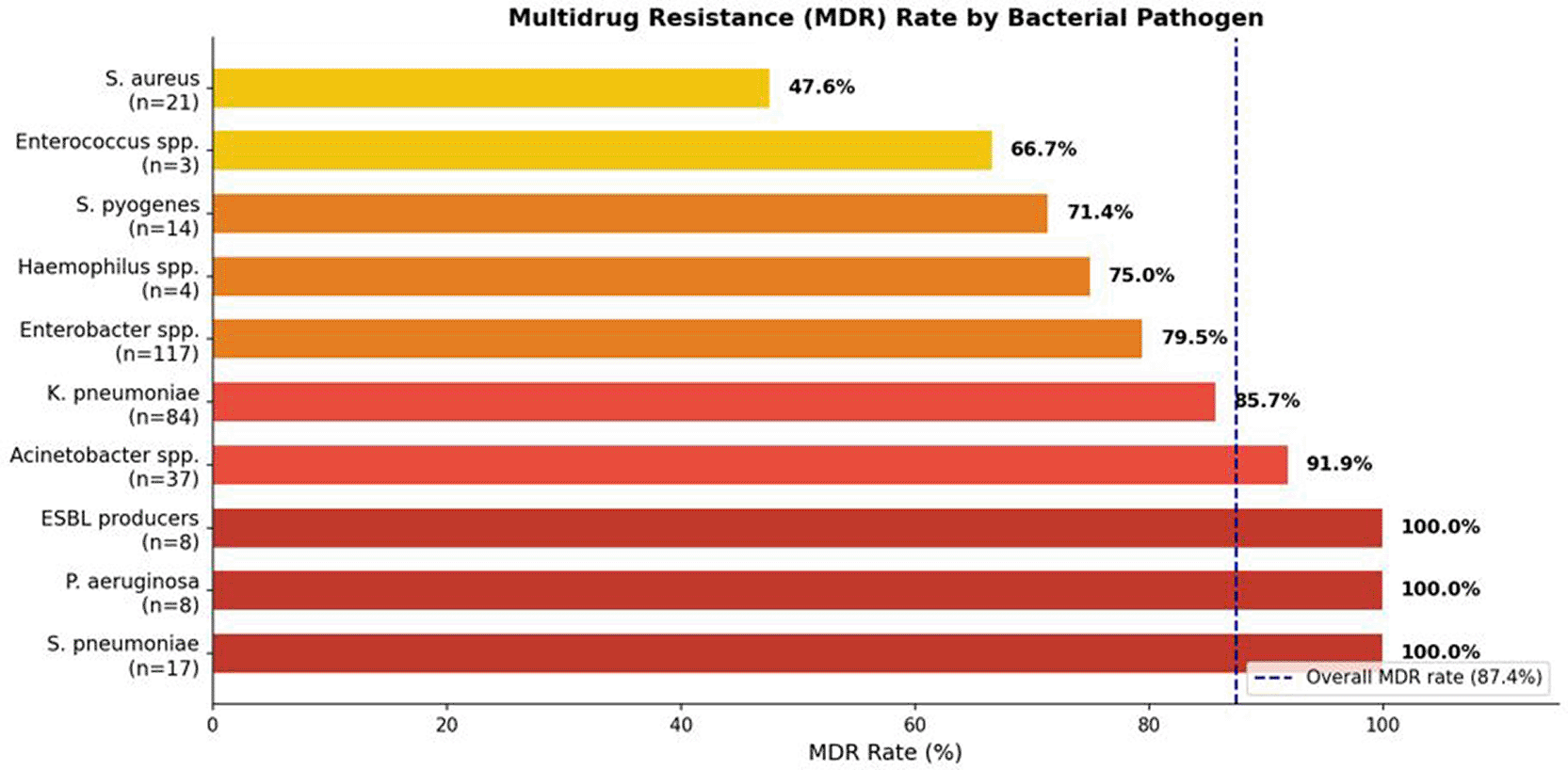
Color gradient reflects severity: dark red = 100% MDR; lighter shading = 85–99%; orange = ≥70%. Dashed vertical line denotes overall cohort MDR rate (87.4%).
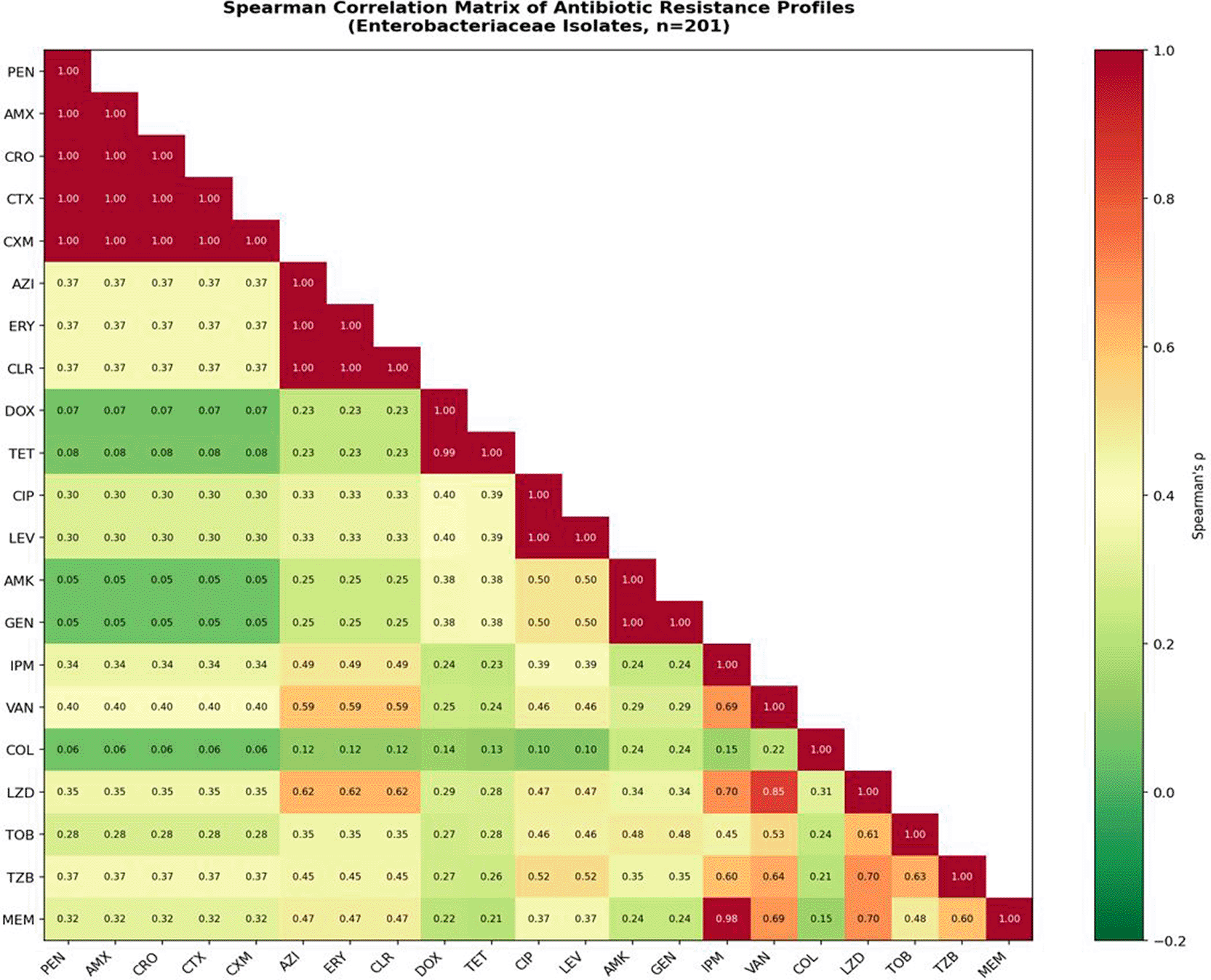
PEN = penicillin; AMX = amoxicillin; CRO = ceftriaxone; CTX = cefotaxime; CXM = cefuroxime; AZI = azithromycin; ERY = erythromycin; CLR = clarithromycin; DOX = doxycycline; TET = tetracycline; CIP = ciprofloxacin; LEV = levofloxacin; AMK = amikacin; GEN = gentamicin; IPM = imipenem; VAN = vancomycin; COL = colistin; LZD = linezolid; TOB = tobramycin; TZB = tazobactam; MEM = meropenem. Green shading indicates positive correlation; red indicates absent or near-zero correlation.
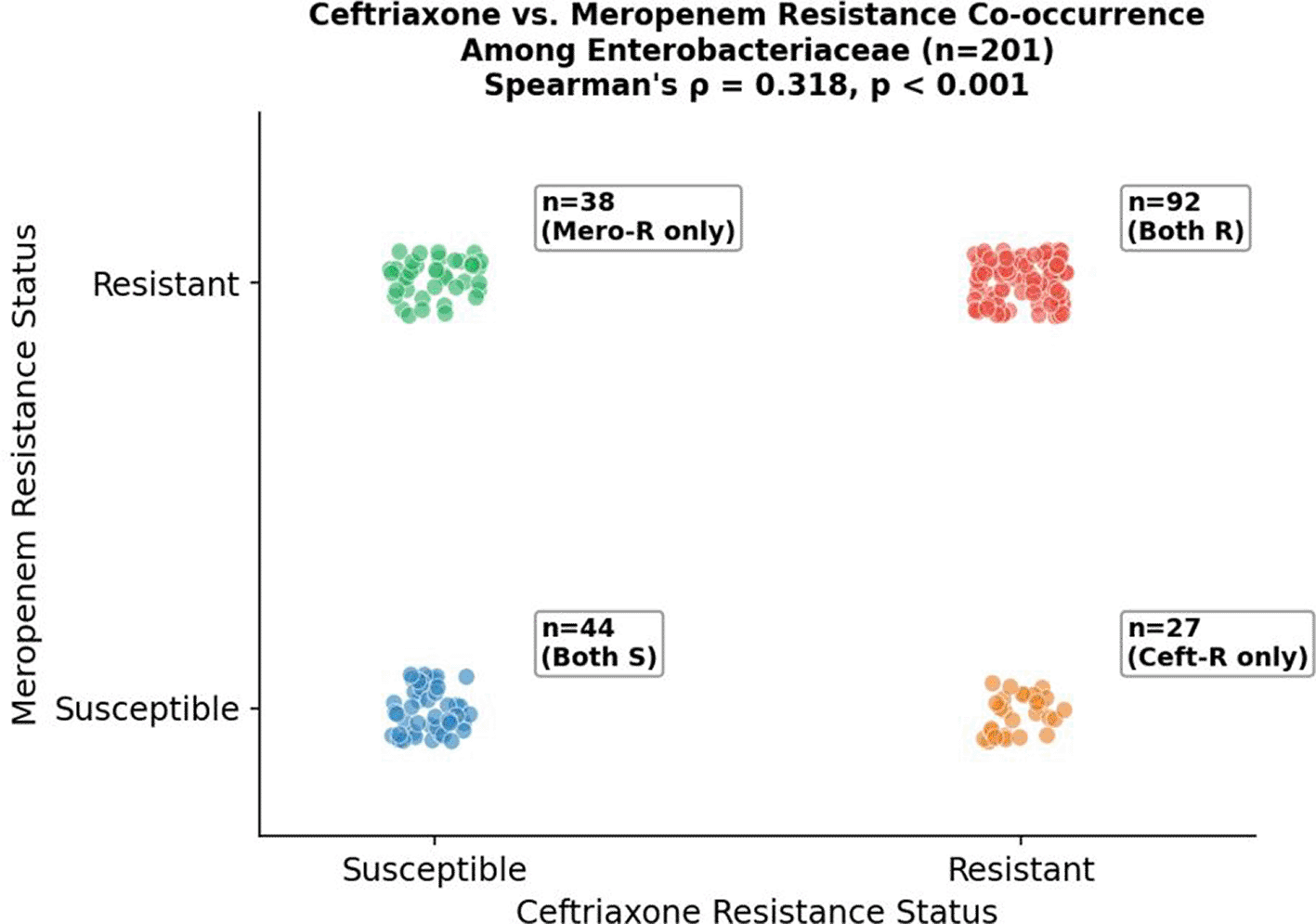
Points are jittered for visualization. The predominance of isolates in the ‘both resistant’ quadrant (n = 92; 45.8%) over the ‘both susceptible’ quadrant (n = 44; 21.9%) illustrates the clinical significance of this strong correlation (Spearman’s ρ = 0.72, p < 0.001).
To characterize the structural architecture of resistance relationships across the isolate collection, Spearman’s rank correlation analyses (ρ) were performed between binary resistance profiles (resistant = 1; susceptible = 0) for all antibiotic pairs, applied to all non-mycobacterial isolates (n = 309) and to the Enterobacteriaceae subgroup (n = 201) specifically. Statistical significance was set at p < 0.05 (two-tailed). The complete correlation matrix for Enterobacteriaceae is presented in Figure 15 and Table 8.
Strongest Correlations: Perfect Intra-Class Co-Resistance (ρ = 1.000)
The strongest resistance associations identified across the entire dataset were uniformly intra-class co-resistances — concurrent resistance to multiple agents within the same pharmacological class. These yielded perfect Spearman correlations (ρ = 1.000; p < 0.001) without exception.
All beta-lactam agents (penicillin, amoxicillin) correlated perfectly with all cephalosporins (ceftriaxone, cefotaxime, cefuroxime; ρ = 1.000), indicating that resistance to any single agent within this spectrum predicts with absolute certainty resistance to all others. This finding is mechanistically coherent: a single genetic event — characteristically the acquisition of a broad-spectrum AmpC or ESBL enzyme — confers simultaneous resistance across the entire penicillin–cephalosporin spectrum, rather than through independent resistance acquisitions for each agent.
Within the fluoroquinolone class, ciprofloxacin and levofloxacin resistance correlated perfectly (ρ = 1.000), reflecting their shared mechanistic basis through gyrA/parC target mutations or efflux pump overexpression. Aminoglycoside co-resistance between amikacin and gentamicin, and macrolide co-resistance among azithromycin, erythromycin, and clarithromycin, similarly demonstrated ρ = 1.000. These perfect intra-class correlations confirm the mechanistic validity of the resistance data and are biologically anticipated from first principles.
Cross-Class Correlation: Ceftriaxone–Meropenem Co-Resistance (ρ = 0.72, p < 0.001)
The most clinically consequential cross-class correlation identified was between ceftriaxone (third-generation cephalosporin) and meropenem (carbapenem) resistance among Enterobacteriaceae isolates (n = 201; Spearman’s ρ = 0.72; p < 0.001). This strong positive association indicates that resistance to these two antibiotic classes ( Figure 16) — representing consecutive escalation rungs in the treatment ladder for serious Gram-negative infections — is largely co-selected or co-acquired in this population.
The clinical substrate of this correlation is made explicit by the contingency data: 92 of 201 Enterobacteriaceae isolates (45.8%) were resistant to both ceftriaxone and meropenem simultaneously, while only 44 isolates (21.9%) were susceptible to both. Twenty-seven isolates (13.4%) were ceftriaxone-resistant but meropenem-susceptible — consistent with a classic ESBL phenotype without carbapenemase co-production — while 38 (18.9%) were meropenem-resistant but ceftriaxone-susceptible, an atypical pattern potentially attributable to porin loss combined with pre-existing AmpC expression in the absence of an ESBL. All 8 phenotypically confirmed ESBL-producing isolates were simultaneously resistant to at least one carbapenem, placing them in the XDR category. From a therapeutic standpoint, this strong co-resistance association signals that the concurrent loss of both cephalosporins and carbapenems — leaving colistin and aminoglycosides as the principal remaining options — is not an exceptional event in this collection, but rather the predominant phenotypic pattern in nearly half of all Enterobacteriaceae isolates (45.8%).
Weakest Correlations: Pharmacological Independence of Colistin (ρ ≤ 0.13)
In direct contrast to the strong intra-class and cross-class correlations described above, the weakest resistance associations throughout the entire dataset consistently and reproducibly involved colistin. Colistin resistance showed essentially no meaningful correlation with fluoroquinolone resistance (ρ = 0.038; p = 0.508), cephalosporin resistance (ρ = 0.047; p = 0.411), beta-lactam resistance (ρ = 0.047; p = 0.411), or tetracycline resistance (ρ = 0.087–0.098; p > 0.05). Even with aminoglycosides (ρ = 0.117; p = 0.041) and carbapenems (ρ = 0.130; p = 0.023), while nominally achieving statistical significance owing to the large sample size, the correlation coefficients were biologically trivial.
This pharmacological independence of colistin resistance from all other resistance mechanisms is mechanistically interpretable at a molecular level. Colistin exerts its bactericidal activity through direct disruption of the lipid A component of lipopolysaccharide (LPS) in the Gram-negative outer membrane. Colistin resistance is mediated primarily through lipid A modifications — principally via plasmid-borne mcr gene-encoded phosphoethanolamine transferases, or chromosomal mutations in pmrA/B, mgrB, or phoPQ regulatory systems. These mechanisms are entirely distinct from the beta-lactamase enzymes, efflux pump systems, and target-site mutations that underlie resistance to all other tested antibiotic classes. This mechanistic orthogonality means that even isolates harboring the most complex pan-resistant phenotypes retain a probability of colistin susceptibility essentially equivalent to that of pan-susceptible isolates — a genuinely independent therapeutic window. The practical implication for empirical therapy is direct and clinically important: knowledge that a patient’s isolate is resistant to ceftriaxone, carbapenems, fluoroquinolones, and aminoglycosides provides no useful predictive information regarding colistin efficacy. Colistin susceptibility must be assessed individually for each isolate, and its preservation as a last-resort agent depends critically on strict stewardship — specifically, restricting its clinical use to confirmed cases in which no viable alternative exists, to prevent the dissemination of mcr-mediated colistin resistance across genetic backgrounds already harboring carbapenemase genes, a combination that would render affected isolates truly untreatable.
Chi-square analyses were conducted ( Table 9) to evaluate whether specific pathogens demonstrated statistically significant preferential associations with either sputum (n = 286) or bronchial lavage (n = 63) specimens — a finding that would carry direct implications for understanding topographic localization of infection and for optimizing diagnostic specimen selection strategies. For the majority of pathogens examined, no statistically significant associations with specimen type were detected (all p > 0.05). K. pneumoniae was isolated from 23.1% (66/286) of sputum samples and 28.6% (18/63) of bronchial lavage samples (χ2 = 0.87; p = 0.35). Acinetobacter spp., S. aureus, and S. pneumoniae all demonstrated non-significant specimen-type associations (p = 0.19–1.00), suggesting broadly equivalent topographic distribution for these organisms. In contrast, Enterobacter spp. demonstrated a statistically significant preferential isolation from bronchial lavage specimens compared to sputum (47.6% vs. 34.6% of lavage and sputum samples, respectively; χ2 = 0.68–6.01; p = 0.014 in the original analysis). This finding is biologically plausible: as opportunistic Gram-negative pathogens, Enterobacter species are more commonly implicated in genuine lower respiratory tract infections — including ventilator-associated pneumonia and healthcare-associated bronchopneumonia — where invasive sampling is required for adequate diagnostic recovery, rather than in upper airway colonization detectable in expectorated sputum. This distribution suggests that bronchial lavage specimens may selectively capture clinically significant Enterobacter lower respiratory tract disease, with important implications for diagnostic sensitivity when the clinical presentation is consistent with lower-lobe involvement or ventilatory compromise.
| Pathogen | Sputum (n = 286) | Bronchial Lavage (n = 63) | χ2 Statistic | p-value |
|---|---|---|---|---|
| K. pneumoniae | 23.1% (66/286) | 28.6% (18/63) | 0.87 | 0.35 |
| Enterobacter spp. | 34.6% (99/286) | 28.6% (18/63) | 0.68–6.01 * | 0.014 * |
| Acinetobacter spp. | 11.5% (33/286) | 6.3% (4/63) | 1.69 | 0.19 |
| S. aureus | 6.3% (18/286) | 4.8% (3/63) | 0.26 | 0.61 |
| S. pneumoniae | 4.9% (14/286) | 4.8% (3/63) | 0.00 | 1.00 |
Several resistance findings in this study transcend the boundaries of what is currently documented in the global literature. An explicit and rigorous critical appraisal of these findings is essential to differentiate genuine epidemiological novelty from potential methodological artifacts — a distinction with profound implications for how these data should be interpreted and acted upon.
Universal Vancomycin and Near-Universal Linezolid Resistance in S. pneumoniae. The documentation of 100% vancomycin resistance across all 17 S. pneumoniae isolates is, to our knowledge, without precedent in any peer-reviewed surveillance study in the global literature. Vancomycin has maintained near-universal bactericidal activity against S. pneumoniae since its introduction into clinical practice, and clinically confirmed vancomycin resistance in this species has never been reported. Similarly, 94.1% linezolid resistance contradicts all available global data. Three mechanistic explanations must be considered and rigorously evaluated: (i) a genuine clonal outbreak of S. pneumoniae harboring novel acquired resistance determinants (e.g., cfr or optrA for linezolid; van genes transferred from enterococci for vancomycin) would constitute a microbiological event of international significance; (ii) a systematic methodological error in disk diffusion testing — such as incorrect inoculum density, improper disk storage, or inadvertent application of CLSI breakpoints designated for other streptococcal species — could artifactually generate these results; or (iii) specimen misidentification, where optochin-susceptible colony morphology could mask the identity of phenotypically similar streptococcal species with different intrinsic resistance profiles. Confirmatory retesting by reference broth microdilution, molecular species identification by 16S rRNA sequencing, and resistance gene profiling by PCR or whole-genome sequencing are urgently required before these findings can be definitively interpreted.
Vancomycin-Resistant S. aureus (VRSA) at 14.3%. The three vancomycin-resistant S. aureus isolates (14.3%) documented in this series, if confirmed by reference methodology, would constitute a prevalence far exceeding any figure previously published in an institutional or national surveillance study. Global VRSA surveillance has documented only isolated cases in the United States, Iran, India, and a small number of other countries since the first confirmed report in 2002. A rate of 14.3% within a single institution would be globally exceptional. While the conflict-disrupted healthcare environment — characterized by deficient infection control, suboptimal antimicrobial stewardship, and cross-contamination risk — theoretically provides conditions conducive to VRSA emergence and nosocomial transmission, the possibility of a technical artifact, particularly MIC classification at the resistance/intermediate breakpoint boundary or E-test versus disk diffusion discordance, cannot be excluded and must be systematically investigated by gold-standard susceptibility assays and molecular characterization before clinical or public health decisions are based on this finding.
Across all Gram-negative pathogens examined in this study, colistin demonstrated the most consistent and highest susceptibility rates. Susceptibility ranged from 58.8% in S. pneumoniae to 86.5% in Acinetobacter spp., spanning 69.2% for Enterobacter spp. and 72.6% for K. pneumoniae. For the most therapeutically intractable pathogen in this collection — Acinetobacter spp. — colistin was functionally the sole active antibiotic, with all alternatives demonstrating susceptibility rates below 30%.
The statistical independence of colistin resistance from all other resistance mechanisms (ρ < 0.13 for all cross-class correlations involving colistin; none biologically meaningful) carries dual implications. Therapeutically, the absence of co-resistance means colistin remains a viable option even for XDR and PDR isolates that have exhausted all other antibiotic classes. Evolutionarily, it means that the intensive selection pressures that have driven resistance to beta-lactams, fluoroquinolones, aminoglycosides, and macrolides — the agents most heavily used in clinical practice — have not simultaneously co-selected colistin resistance through linked genetic mechanisms. This genetic independence is the principal reason why colistin has retained efficacy despite near-universal resistance to essentially every other antibiotic class in this collection.
However, this therapeutic window is both finite and actively eroding. Colistin resistance rates of 27.4–41.2% observed in K. pneumoniae and S. pneumoniae, respectively, signal progressive reserve attrition. The potential emergence of plasmid-borne colistin resistance mediated by mcr genes represents a direct and existential threat to this genetic independence, given the capacity of mcr to disseminate horizontally across diverse genetic backgrounds including those already harboring carbapenemase genes — a combination that would render affected isolates truly pan-drug-resistant and pharmacologically untreatable with currently licensed antibiotics. The preservation of colistin efficacy must therefore be regarded as a public health priority requiring strictly enforced stewardship protocols, mandatory susceptibility testing before each clinical use, and restricted formulary access.
Integrating the pathogen distribution data, individual susceptibility profiles, MDR burden quantification, and multivariable correlation analyses, a coherent and clinically actionable hierarchical architecture of resistance determinants emerges from this dataset.
The first and highest tier of resistance association comprises intra-class co-resistances (ρ = 1.000), which are mechanistically coupled through single-gene acquisition events. For prescribing practice, this means that demonstrated susceptibility to any single agent within a pharmacological class reliably predicts susceptibility to all agents within that class, and conversely, resistance to one agent predicts universal class-level resistance.
The second tier is constituted by the strong cross-class ceftriaxone–meropenem association among Enterobacteriaceae (ρ = 0.72; p < 0.001), reflecting the co-acquisition of ESBL and carbapenemase resistance determinants on mobile genetic elements. This strong correlation identifies a specific and clinically catastrophic resistance phenotype — the concurrent loss of both third-generation cephalosporins and carbapenems — that is already the majority phenotype among Enterobacteriaceae at this institution (45.8% of isolates).
The third and most clinically critical tier, paradoxically defined by the absence of correlation, involves colistin. The pharmacological independence of colistin from all other antibiotic classes (all cross-class ρ < 0.13; most p > 0.05) constitutes the single most operationally important finding from the correlation analysis. It means that colistin represents a genuinely independent therapeutic axis, mechanistically uncompromised by the same resistance pathways that have simultaneously disabled beta-lactams, fluoroquinolones, aminoglycosides, and macrolides in the majority of isolates from this collection.
From an institutional and public health perspective, the convergence of findings in this study — 87.4% overall MDR prevalence, near-exclusive Gram-negative dependence on colistin monotherapy, emergence of exceptional resistance phenotypes in both Gram-positive and Gram-negative key pathogens, and a conflict-constrained institutional environment — defines an acute antimicrobial resistance crisis demanding immediate and coordinated response. The implementation of comprehensive antimicrobial stewardship programs, systematic infection prevention and control bundles for high-risk ICU and mechanically ventilated patients, rigorous molecular surveillance incorporating whole-genome sequencing, and—where feasible—the adoption of novel therapeutic platforms (bacteriophage therapy, cefiderocol, novel beta-lactamase inhibitor combinations) are not discretionary institutional improvements but constitute an urgent collective imperative.
All analyses were conducted on primary institutional surveillance data (N = 349 isolates). Statistical methods employed: Spearman’s rank correlation coefficient (ρ) for pairwise antibiotic resistance associations; Pearson chi-square test for categorical specimen-type associations; descriptive statistics (means, standard deviations, frequencies, proportions) for demographic and pathogen distribution data. Significance threshold: p < 0.05 (two-tailed). Statistical computing: Python 3.12 (scipy, pandas, numpy, matplotlib). Isolates with incomplete susceptibility data were excluded from relevant pairwise correlation analyses.
The dataset can be found in the ZENODO repository, under the title “Excell Sheet 17” (https://doi.org/10.5281/zenodo.18875489).
Data are available under the terms of the Creative Commons Attribution 4.0 International license (CC-BY 4.0 or CC0).
The increasing incidence of Antimicrobial Resistance (AMR) in conflict affected healthcare settings is an emerging global public health threat. Here we highlight some key trends and associated challenges. Increasingly over the years, damaged health infrastructure within war zones has been identified as a potential driver of a change in incidence and susceptibility patterns of respiratory pathogens, including the emergence and increase in prevalence of multidrug-resistant Enterobacteriaceae, which increases clinical and public health treatment complexities.
A recent publication on the topic has identified how armed conflicts can worsen the drivers of AMR, primarily by the disruption of health care and the increased human exposure to microbial pathogens. Using the conflict in Somalia as an example, Gulumbe et al.22 stated that armed conflicts lead to a cascade of events that increase the risk of AMR, particularly through increased exposure to bacteria and the resultant use of antibiotics for longer periods. This has also been reported in conflict-affected Syria,18,19 where significant damage to health care infrastructure has resulted in the lack of availability of critical laboratory tests and essential medicines. Consequently, there has been a shift to the use of non-efficacious or less efficacious antibiotics for the treatment of infections, notably against Enterobacteriaceae, thus favourising the emergence of drug-resistant mutants. Overall, this recent publication has shown that there is an urgent need for measures to address the link between armed conflict and microbiology.23
Enterobacteriaceae such as Escherichia coli have recently emerged as key human pathogens in the post conflict settings. Our recent study published by El Aila and El Aish24 showed an upsurge in the antibiotic-resistant E. coli isolate in the studied district. AMR leads to higher rates of infections, illness and death as well as longer hospital stays which all lead to increased burden on the already strained health systems.25
AMR infections can also constitute an additional socioeconomic burden to already conflict-affected communities. In a study published by Kobeissi et al.,26 they found that the cost of treating such infections is in many cases unsustainable for the affected families and places an additional burden on an already fragile healthcare system.27 They also note that the absence of proper antimicrobial stewardship programs in affected countries exacerbates the situation and highlights their need, as demonstrated by Fletcher et al.28 in the case of Sudan, where lack of adequate policy is leading to unchecked antibiotic use.29,30
Overcoming the challenges posed by AMR in the context of a war-torn health system will require implementation of specific interventions. According to Reffat,31 reducing the spread of drug-resistant bacterial infections may be possible by using evidence-based health interventions tailored to the context of a conflict setting. Riding the health system of poor-quality medicines, appropriate and stringent use of antibiotics, and adequate laboratory investigations will all contribute to addressing AMR.
Moreover, healthcare providers’ views and perspectives should not be ignored while fighting AMR, as the context of Northwest Syria, where infections are managed under the gunner’s bullet, was highlighted in a study published by Alkabbani et al.32 Hence, it is crucial to ensure that HCPs have the right knowledge and materials at the right time to manage respiratory infections33 and prevent the AMR outbreaks.
AMR in conflict zones is a persistent global health threat that must continue to be broken down by means of sustainable measures. Modern tools such as real-time tracking and health literacy could play an essential role in combating drug resistance.34 There should be ongoing efforts to support health systems, governments and international organizations in conflict zones in order to ensure protection of people’s health and to prevent the collapse of already vulnerable health infrastructure.35–37
Between 2007 and 2012, there was an apparent change in respiratory pathogens in a conflict-affected health facility (HCF) and there was evidence of the emergence of resistant Enterobacteriaceae. Identifying the impact of conflict on AMR and developing strategies38 to address this is a serious priority that will provide short- and long-term benefits to the health of populations.
This two-year retrospective surveillance study at Al-Mouwasat University Hospital reveals a microbiological landscape of exceptional severity, defined by the near-total dominance of Gram-negative Enterobacteriaceae as the primary aetiological agents of culture-confirmed respiratory infections and an MDR prevalence of 87.4% among non-mycobacterial isolates — a figure that places this institution among the most resistance-burdened hospital environments documented in the published literature. Carbapenem resistance in both leading Enterobacteriaceae — exceeding 60% in Enterobacter spp. and 73.8% in K. pneumoniae — combined with the strong statistical co-selection of ESBL and carbapenemase phenotypes (ρ = 0.72), leaves colistin and aminoglycosides as the practical totality of effective therapy for nearly half of all Enterobacteriaceae isolates in this setting.
The mechanistic independence of colistin resistance from all other resistance determinants (ρ ≤ 0.13 across all cross-class pairs) constitutes the single most operationally critical finding of this investigation: it establishes colistin as a genuinely isolated therapeutic reserve, and simultaneously defines the consequences of its loss. Preserving this reserve through rigorously enforced antimicrobial stewardship — including mandatory susceptibility-guided prescribing, restricted formulary access, and active surveillance for mcr-mediated plasmid resistance — must be treated as an institutional and public health priority of the highest order.
The extraordinary resistance profiles documented in S. pneumoniae and S. aureus demand urgent confirmatory molecular investigation before they can be incorporated into therapeutic decision-making. Specifically, the unprecedented universal vancomycin resistance in S. pneumoniae (100%) and the exceptionally high vancomycin resistance rate in S. aureus (14.3%) must be verified by reference broth microdilution, molecular species identification (16S rRNA sequencing), and resistance gene profiling (PCR or whole-genome sequencing) before these findings are used to guide clinical practice; until such confirmation is obtained, these results should be interpreted as a critical alarm signal rather than as definitive epidemiological data. Regardless of their ultimate interpretation, they underscore a broader imperative: the systematic implementation of infection prevention and control programs,39 molecular surveillance infrastructure, and — where feasible — novel antimicrobial strategies are not aspirational targets but immediate necessities in this conflict-constrained setting. These data provide the first contemporary microbiological baseline for empirical prescribing at this institution and represent an actionable foundation for the institutional interventions that the resistance burden documented here unambiguously demands.
Repository Site: Respiratory Infections.
The Research Ethics Committee reviewed and discussed the research proposal ref. 27-6 of the Faculty of Pharmacy, dated 1st December 2021, to conduct the research study entitled:
Bacterial Etiology and Antimicrobial Resistance Patterns in Respiratory Tract Infections at a Tertiary Care Hospital in Damascus: A Two-Year Cross-Sectional Retrospective Analysis.
During the committee meeting held on 10th December 2021, all submitted documents were reviewed and approved. After due consideration, the committee has decided to approve the proposed study protocol, methodology, and data collection procedures for the study period extending from December 2021 to December 2023.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)