Keywords
FN3K, FN3KRP, glycation, deglycation, glucosepane, Maillard reaction, fructosamine, ketosamine
FN3K, FN3KRP, glycation, deglycation, glucosepane, Maillard reaction, fructosamine, ketosamine
Nonenzymatic reactions between glucose and proteins (Maillard reactions) are an unavoidable feature of life. These reactions have mostly adverse effects on proteins by inhibiting enzymes, interfering with receptor function, and crosslinking structural proteins such as collagen.1,2 Traditionally, these reactions were thought to be totally nonenzymatic with no mechanisms to reverse or control them. This changed in the year 2000 with the discovery of Fructosamine-3-Kinase (FN3K)3,4
This enzyme phosphorylates the first stable intermediates of the Maillard reaction, protein-bound fructosamines, to fructosamine-3-phosphates. Since these phosphoesters are intrinsically unstable, this reaction leads to decomposition of the glycation adducts, regeneration of an unmodified protein, and concomitant production of 3-deoxyglucosone.4
When FN3K was isolated in our laboratory, it copurified with another closely related protein that has an identical MW but a slightly lower isoelectric point (Fig. 1).4 At the time, this second protein was assumed to be an isozyme of FN3K with the same substrate specificity, namely D-fructosamine. Subsequent work revealed, however, that this enzyme, renamed Frucose-3-Kinase-Related Protein (FN3KRP), is quite distinct from FN3K because it does not phosphorylate D-fructosamines. Extensive work by Van-Schaftingen et al. has shown that FN3KRP phosphorylates a number of other Amadori products on the C3-hydroxyl. These include D-erythrulosamine, D-ribulosamine and D-psicosamine.5 While none of these ketosamines, nor their aldose precursors, are found in cells, Van Schaftingen et al. have suggested and documented ways in which they can be produced from erythrose-4-phosphate and ribose-5-phosphate by a combination of Maillard reactions of the phosphorylated metabolites and dephosphorylation of the resulting Amadori products by specific phosphatases.6,7
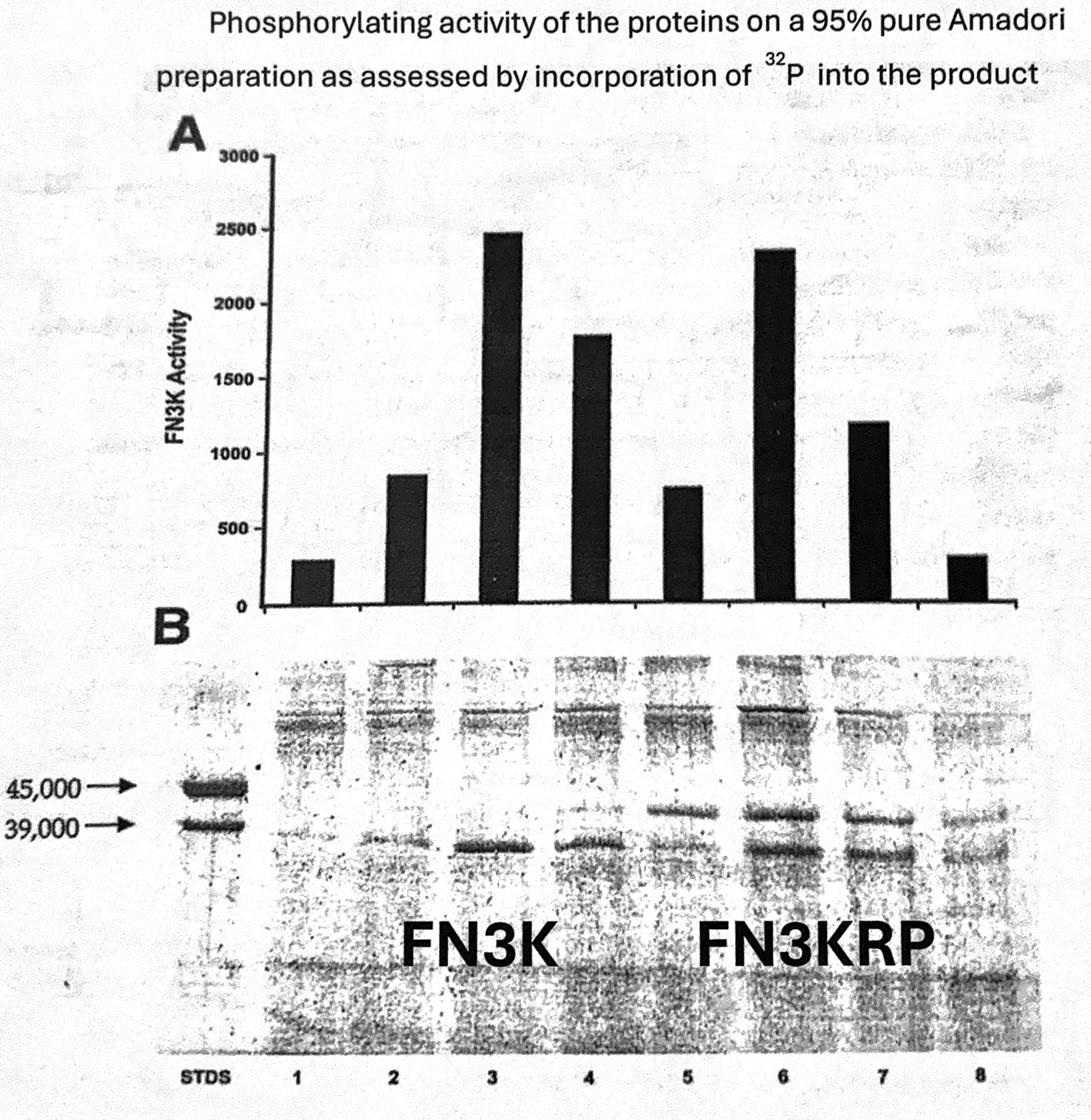
The assay was conducted by measuring the incorporation of 32P into the product from 32P ATP. B) SDS PAGE of fractions from an isoelectric-focusing gel showing the IEF isoforms of FN3K and FN3KRP corresponding to the activity profile shown in panel A.
Subsequent to Van Schaftingen’s work, using recombinant FN3KRP, I was able to show that, in addition to the compounds identified above, FN3KRP also phosphorylates glucitolamines such as N-methylglucamine (meglumine).8 However, unlike the other substrates, this phosphorylation occurs on the C4-hydroxyl. This was a surprising result, the significance of which was not apparent until a recent closer re-examination of data on purified FN3K, as shown in Fig.1. As can be seen in figure, phosphorylating activities are evident with both FN3K and FN3KRP, even though the substrate used here is nominally just fructosamine. The activity of FN3K is easy to understand; however, the source of FN3KRP activity is not clear.
The most likely explanation for the phosphorylating activity of FN3KRP is that while the substrate used was nominally fructosamine, it was not completely homogeneous. As assessed at that time, it was only 95% pure but it probably also contained a compound (or compounds) that would be a substrate for FN3KRP. Given that the radioassay for FN3K used in these experiments contained saturating concentrations of fructosamine (1 mM vs. Km ~ 25 μM), it is likely that the FN3KRP substrate, even if present as only a small fraction in this preparation, would be phosphorylated sufficiently (by FN3KRP) to be detected.
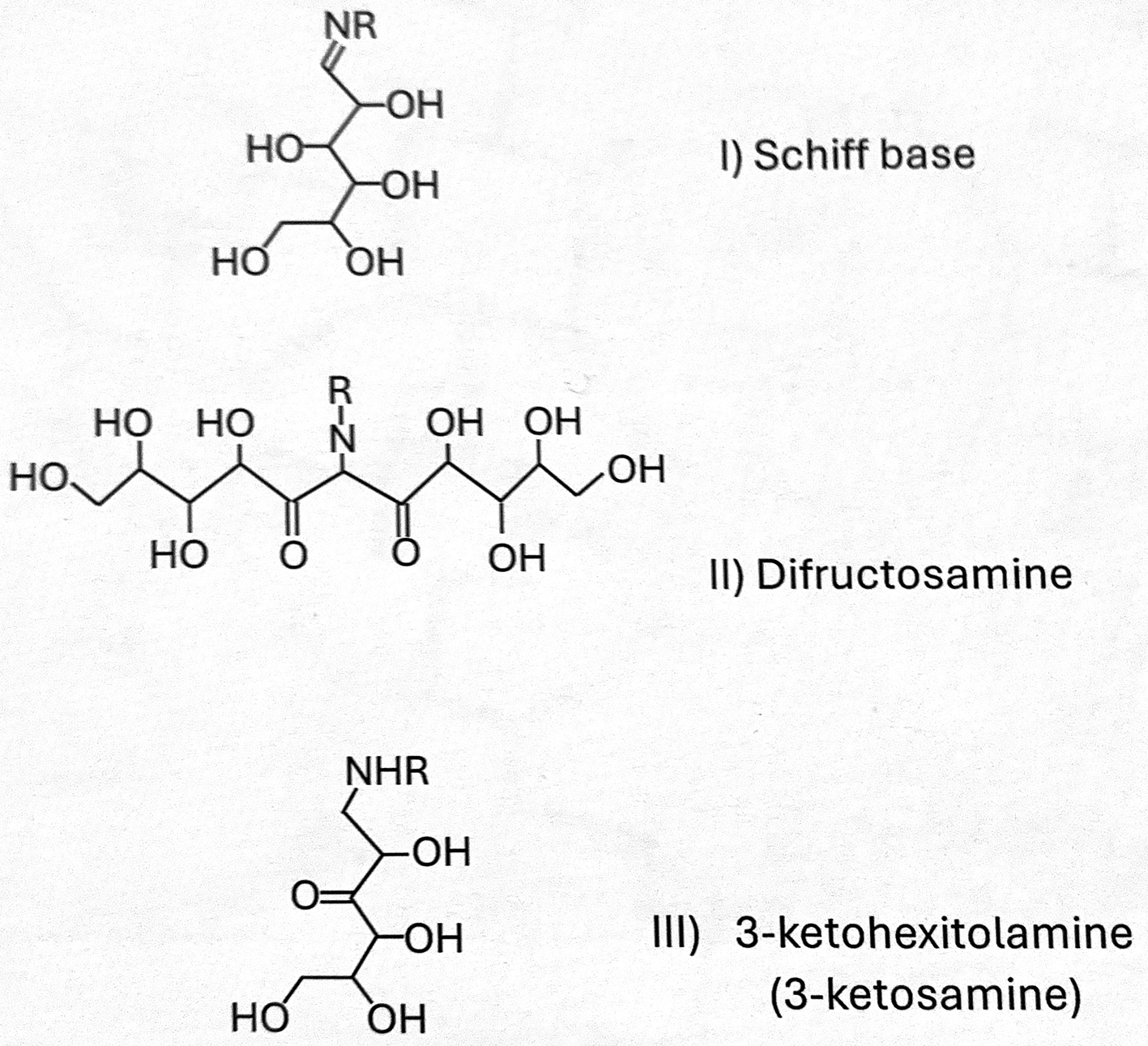
Several candidates for the identity of this “contaminant” substrate are shown in Fig. 2, including the following:
I) A Schiff base of the amine with glucose
II) A di-fructosamine adduct
III) A glycation intermediate downstream of fructosamine, namely 3-keto-D-glucitolamine.
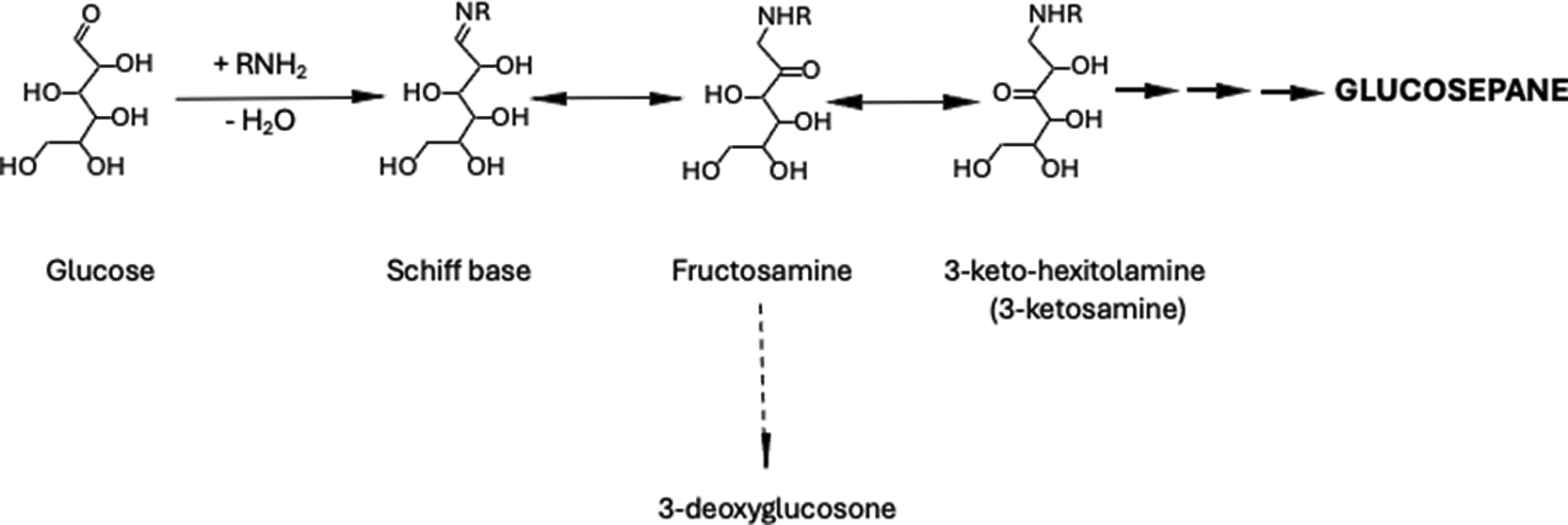
I propose that the physiological substrate of FN3KRP is 3-keto-D-glucolamine (3-ketosamine) produced from fructosamine by migration of the C2-carbonyl to the C3 position (Fig. 3), The product of this reaction, 3-ketosamine-4-phosphate, is intrisically unstable and decomposes to regenerate an unmodified protein along with a concomitant production of a tricarbonyl, 5,6-dihydoxy-2,3-dioxohexanal (Fig. 4).
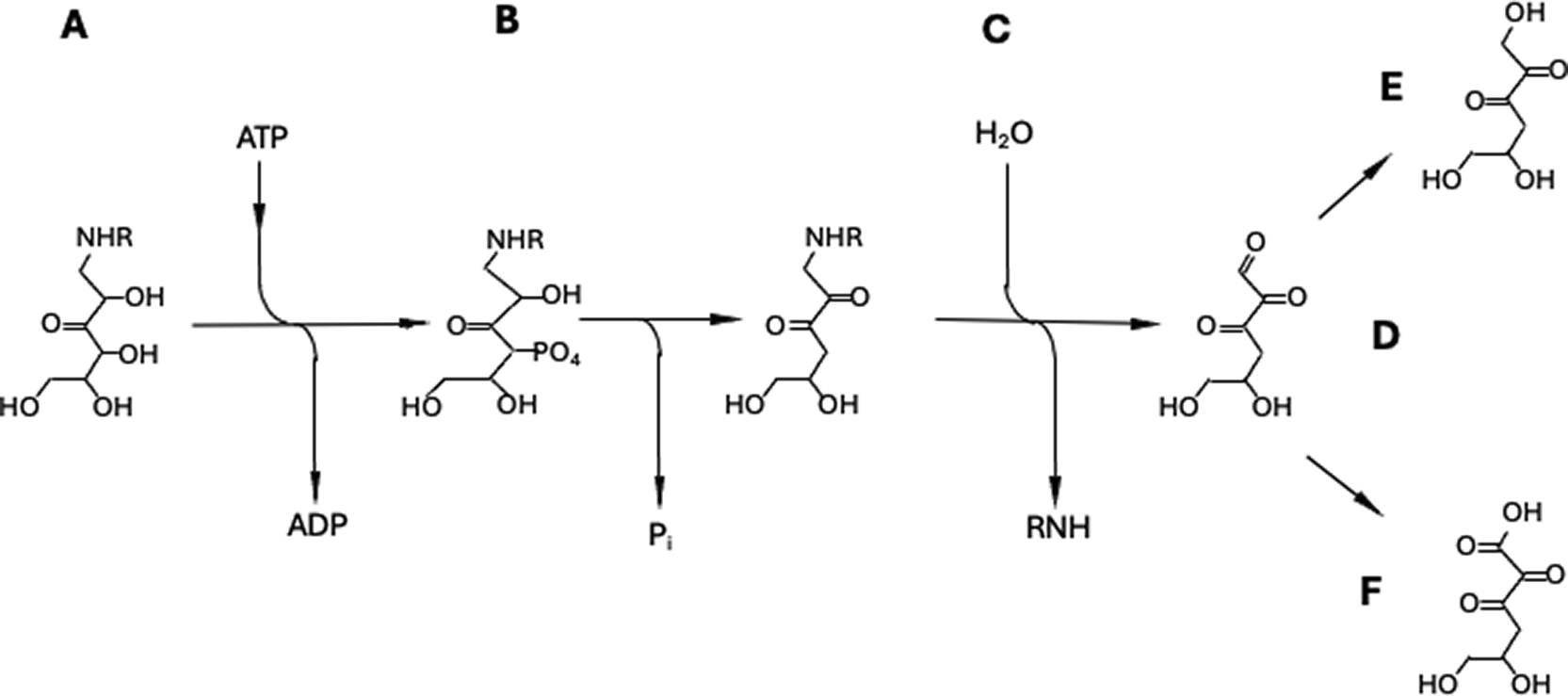
A) 3-ketosamine is phosphdrylated by FN3KRP to 3-ketosamine-4-phosphate. B) Due to its structure, 3-ketosamine-4-phosphate decomposes by-elimination of phosphate to 4-deoxy-3-ketosamine. C) In a reverse Schiff reaction, 4-deoxy-3-ketosamine dissociates to an unmodified amine (protein) along with the production of a tricarbobyl, 4,5-dihydroxy-2,3-dioxohexanal (compound D). This tricarbonyl group is then reduced to 4-deoxyglucosone (compound E) and/or oxidized to 4-deoxy-2,3-dioxogluuconic acid (compound F).
A) Based on the work of Lederer et al.,9,10 it is clear that during the Maillard reaction of glucose with an amine, the carbonyl moieties of the sugar migrate rapidly along its backbone from C2 to C3 and onwards. Thus, it is very likely that in an Amadori preparation, synthesized by condensation of an amine with glucose, some ketosamines with C3, C4, C5, and C6 carbonyls will be present. Since 3-ketosamine (−3-ketohexitolamine) is a glycation intermediate directly downstream of fructosamine, it is a good candidate as a substrate for FN3KRP and would be phosphorylated at C4.
B) Given the ability of FN3KR to phosphorylate N-glucitolamines on the C4 hydroxyl, its suggested function as a 4-kinase of 3-ketosamines is consistent with the fact that its close homolog, FN3K, acts as a 3-kinase not only with fructosamines but also with glucitols and glucitol amines.8,11–13
A) Testing this hypothesis should be straightforward, requiring only the synthesis of an Amadori product using glucose and an amine and the use of recombinant KN3KRP produced in HEK193 cells.14,15 Since FN3KRP does not phosphorylate fructosamines, a reaction of the recombinant enzyme with high concentrations of the Amadori product (~ 50 mM) is likely to result in the phosphorylation of a small amount of substrate present in the preparation. The 1H decoupled 31P NMR spectrum of the reaction mixture revealed several peaks attributed only to the phosphorylated substrate. In the proton-coupled 31P NMR spectrum, these peaks split into doublets with a coupling constant of ~10 Hz.
B) Secondary tests of the hypothesis will involve determination of the decomposition rates of the phosphorylated substrate(s) and identification of byproducts that should include tricarbonyl (5,6-dihydroxy-2,3-dioxohexanal) as well as its reduced and oxidized metabolites, 4-deoxyglucosone and 4-deoxy-2,3-dioxogluconic acid (Fig.4).
Reservations about the hypothesis
There is little doubt that the Maillard reaction between an amine and glucose produces substrate(s) for FN3KRP. However, even if that compound is not 3-ketosamine, identifying the physiological substrate of FN3KRP is still worthwhile.
Implications
If my hypothesis proves correct, then, at the very least, it will provide some new insights into the process of intracellular Maillard reactions and their control. Moreover, given the number of recent publications linking FN3KRP to a number of interesting phenotypes,16–21 the potential impact of this finding may prove to be substantial.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)