Introduction
Signal transduction systems are often modeled as networks of biochemical kinetic equations implemented as continuous-time dynamical models using differential equations1,2. If we regard a subset of species as inputs, and make sure that the system always converges to equilibrium values by using weakly reversible equations3–5, we may transform these models into a set of matrices fulfilling the role of input-output transfer functions, i.e. a mapping from sustained input signal levels to steady-state concentrations for all target species6. Protein signaling functions (psfs) are a systemic generalization of individual dose-response functions, which are usually described by Hill equations7. In contrast to Hill equations, which are not available for enzymatic reactions, which only calculate relative concentrations, and which only work for one reaction in isolation, the psf system calculates enzymatic and complex formation reactions in a complex systemic environment using absolute concentrations6. In addition, the reaction times to equilibrium are calculated as delay values, and the dynamic shape (“transients”) is also available for further analysis.
In this paper, we want to ask the question of optimality of signal transduction. From an evolutionary standpoint, we assume that any biological signal transduction system is constructed with optimized efficiency of signal transmission. Furthermore, we assume that cells have the ability to adapt to perturbations of protein concentrations and changes in extracellular signaling by reinstating signal transmission efficiency. In the following, we investigate this question using a biologically realistic system – beta-adrenergic signaling in a submembrane compartment of a mouse embryonic fibroblast for a single input scenario, focusing on a selected target species as relevant output or actuator of the system (Figure 1A).
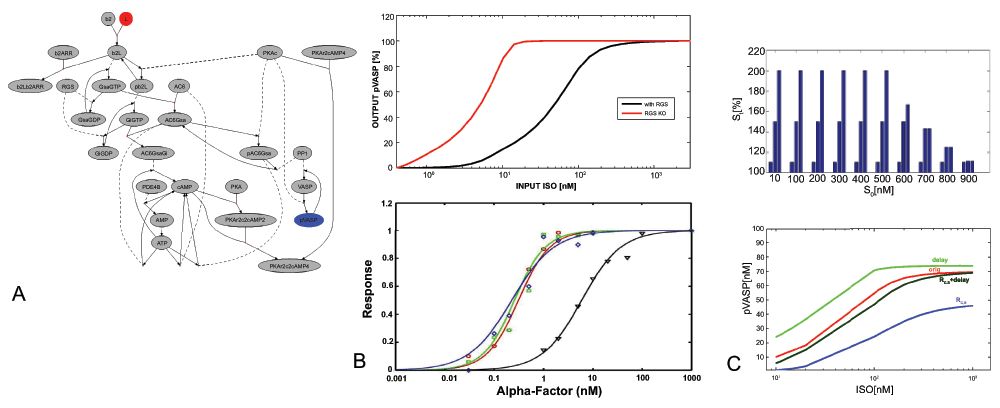
Figure 1.
A. Biochemical Reaction System with Selected Input (red) and Output (blue) B. (Top) ISO-pVASP transfer function (with RGS KO, red) in the MEF model11 (Bottom) Experimental Response to RGS KO for GPCR signal-response in a yeast model10 C. (Top) Distribution of Extracellular Signals Used to Calculate Signal Transmission. From a baseline signal S0 [10nM–900nM] the extracellular signaling level St rises to 110%,150% and 200%, but not above 1µM. (Bottom) Transfer Functions for ISO → pVASP. Shown is the original model, optimization by delay, by RC,S, and by both.
Experimental analysis of signal transduction systems has shown fold-change responses to changes in input8,9. Accordingly, input-output transfer functions usually follow the shape of hyperbolic (saturating) curves, which are equivalent to sigmoids for logarithmically scaled input6. In Figure 1B we show the effect of a knock-out (KO) for a RGS protein in an experimental assay in yeast10, and compare this with the effect in the model system11. We see that the effects of the RGS KO on dose-response signaling efficiency are robust across very different cellular systems.
Usually, when we use a computational model to investigate perturbations, we only study the effects as reflected in the simulation. By utilizing optimization in terms of signaling efficiency, we can make the system itself adjust to the perturbation. In this way, we are studying signal transduction as a self-organizing system, which uses objective functions to adapt. This basic idea is extremely powerful, and could be used with different kinds of constraints on parameters, reaction times, etc. and with different, multiple input scenarios for larger systems. To explore this question further is of significant importance in assessing cellular health and functioning.
Methods
Example system: GPCR signaling in a submembrane compartment
Figure 1A shows the example system, a submembrane compartment with a GPCR (G-protein coupled receptor) pathway from a mouse embryonic fibroblast, with ISO as input (extracellular ligand to β(2)-adrenergic receptors) and the phosphorylation of a protein VASP as output. This model was implemented as an ODE model with 23 reactions and 27 molecular species, derived from 12 initial concentrations (cf. Table 1, Table 2). The parameters were adapted to experimental biological data (not shown11). In this subsystem, the central cAMP response often follows a plateau curve, i.e. a rise to steady-state, but cAMP transients which are typically observed in cytoplasm may also occur11. The dose-response transfer functions were derived as in6.
Table 1. Kinetic rates of the sample system, adapted from Sabio-Rk25 and Brenda26, adjusted to experimental cAMP time-series data11.
| kon | koff | kcat |
|---|
| b2 + L ↔ b2L | 0.0003 | 0.1 | |
| b2L + PKAc ↔ b2LPKAc → pb2L + PKAc | 0.00026 | 1 | 5.4 |
| pb2L → b2L | | | 0.1 |
| GsaGDP + b2L ↔ b2LGsaGDP → GsaGTP + b2L | 0.006 | 0.8 | 0.2 |
| GsaGTP + RGS ↔ RGSGsaGTP → GsaGDP + RGS | 0.0008 | 1.2 | 16 |
| GiGDP + pb2L ↔ pb2LGiGDP → GiGTP + pb2L | 1.2 | 0.8 | 16 |
| GiGTP + RGS ↔ RGSGiGTP → GiGDP + RGS | 1.2 | 0.8 | 16 |
| GsaGTP + AC6 ↔ AC6Gsa | 0.00385 | 3 | |
| GiGTP + AC6Gsa ↔ AC6GsaGi | 0.00385 | 10 | |
| AC6Gsa + PKAc ↔ AC6Gsa_PKAc → pAC6Gsa + PKAc | 0.00026 | 1.5 | 30.4 |
| pAC6Gsa + PP1 ↔ pAC6PP1 → AC6Gsa + PP1 | 0.0026 | 3 | 54 |
| ATP + AC6Gsa ↔ AC6Gsa_ATP → cAMP + AC6Gsa | 6e-05 | 10 | 80.42 |
| ATP + pAC6Gsa ↔ pAC6Gsa_ATP → cAMP + pAC6Gsa | 6e-05 | 10 | 8.042 |
| ATP + AC6 ↔ AC6_ATP → cAMP + AC6 | 0.0001 | 120 | 0.142 |
| cAMP + PDE4B ↔ PDE4BcAMP → AMP + PDE4B | 0.03 | 77.44 | 19.36 |
| 1PKA + 2cAMP ↔ 1PKAr2c2cAMP2 | 3.5e-08 | 0.06 | |
| 1PKAr2c2cAMP2 + 2cAMP ↔ 1PKAr2c2cAMP4 | 2.7e-07 | 0.28 | |
| 1PKAr2cAMP4 + 2PKAc ↔ 1PKAr2c2cAMP4 | 8.5e-08 | 0.05 | |
| VASP + PKAc ↔ VASP PKAc → pVASP + PKAc | 0.00026 | 1.5 | 30.4 |
| pVASP + PP1 ↔ pVASPPP1 → VASP + PP1 | 0.0026 | 3 | 54 |
| AMP → ATP | | | 1 |
| ATP + AC6GsaGi ↔ AC6GsaGi_ATP → cAMP + AC6GsaGi | 6e-05 | 10 | 1 |
| b2L + bARR ↔ b2LbARR | 0.0006 | 0.1 | |
Table 2. Initial concentrations (in nM) of species in the sample system.
| PDE4B | 200 | b2 | 100 |
| bARR | 500 | PKA | 500 |
| cAMP | 100 | AC6 | 1000 |
| GsaGDP | 200 | GiGDP | 200 |
| RGS | 100 | VASP | 200 |
| PP1 | 100 | ATP | 1e+06 |
Objective function
A biological signal transduction system is defined by its state variable vector x, the set of all kinetic rate and initial concentration parameters.
We hypothesize that an efficient signal transmission would maximize the response coefficient RC,S (the response of species C to input S) defined as
with concentration change of target C and input S from baseline (t=0) to signal time t. For RC,S, values < 1 show signal loss, with 1 for perfect transmission, and values > 1 showing signal amplification. We may also optimize for the slope s of the sigmoid at half-maximum concentration. This is equivalent to maximizing RC,S, provided that the input signal remains entirely between the upper and lower boundaries of the sigmoid (Figure 1C). By optimizing for s, additional to RC,S, we may force the system to implement a switch-like function instead of a more linear function. However, shifting the sigmoid function to the left or to the right is more important as the slope in our models.
In addition, we optimize for reaction time (delay to steady-state). Steady-state is defined, pragmatically, as relative change of less than 2% over 100s. The delay (dS) is computed for 90% (EC90) of steady-state. We may now define an aggregate objective function:
to select the system state variables that minimize delay and maximize response.
In addition to signal transmission from extracellular concentration changes onto steady-state concentrations, such as they typically occur for temporally integrating proteins like transcription factors, we also look at signal transmission by transients, i.e. peak concentration, in response to extracellular signals. In this case, we minimize the delay to peak value, maximize the response at peak value, and minimize the response at steady-state value.
Results
Delay vs. efficiency trade-off
The computation of signal transmission efficiency will be explained here for a single input-output pathway of cAMP/PKA-mediated transmission in a cellular membrane compartment. It is clear that a complex signaling system may have several inputs, and a large number of outputs or target proteins, and this is especially the case for cAMP-mediated signaling. Nonetheless we will focus on the simple case here to explain the basic principle. The input is a membrane receptor, a G-protein coupled receptor (GPCR), β(2)AR, which is activated by an extracellular pharmacological agonist ISO (isoproterenol); the output is a membrane protein, VASP, which promotes actin filament elongation. Activation of the receptor selectively inhibits VASP by phosphorylating VASP to pVASP via protein kinase A (PKA) activation. In Figure 1B the original ISO/pVASP transfer function is shown, which we use here for further optimization. The system was trained for a signal distribution between 10nM and 1 µM ISO adjusting both kinetic rate and concentration parameters (Figure 1A). The optimization method used is a simplex algorithm12. Figure 1B shows the transfer function before and after adjustment, maximizing for RC,S, dS or the combined function f. The results are summarized in Table 3. We see that optimizing for the response coefficient alone shifts the function to the right to better cover the input range. Optimizing for the delay alone shifts it to the left, speeding up signals in the lower range (which are slower). Both objectives together (equally weighted) are fairly close to the original, biologically validated curve, with an improved RC,S.
Table 3. Reaction times (dS) and response efficiency (RC,S) in a biological system and under optimization.
| RC,S | dS |
|---|
| original | 0.272 | 423 |
| optimized for RC,S
| 0.41 | 594 |
| optimized for dS
| 0.11 | 169 |
| optimized for f | 0.34 | 481 |
In general, biochemical reactions are faster at higher substrate concentrations, but the relative concentration change in response to an increase in the enzyme or the binding partner is less. This is a fundamental trade-off between delay and transmission efficiency that may define an optimal operating range for a signaling system and be of relevance in disease processes6. The results obtained with this experiment are simple, intuitive and encourage continuing to explore the basic idea.
Evolutionary learning of kinetic rate parameters
In principle, we may use all parameters in a system, concentrations or kinetic rates, to maximize signal transmission. But the evolution of protein structure and interactions shows that it is fine-tuning of molecular kinetic parameters which is subject to evolutionary learning, while concentrations are often regulated adaptively in each cell. Mass-action kinetics approximate molecular kinetic parameters, even though there are significant sources of uncertainty, such as the stochastics of molecular interactions. In the following, we explore the idea that kinetic rate learning operates on evolutionary time-scales, and that biologically attested signal transduction pathways contain reaction rates which are optimal in terms of signal transmission efficiency. We use known concentration ranges, specific by cell type, together with kinetic rate optimization.
To explore the parameter space, we drew 1662 values for all kinetic rate parameters (kon, koff, kcat) from a distribution of 20% to 500% of the original parameters, and calculated RC,S and dS for the corresponding models, relative to an improved signal distribution from 1nM to 1µM (Figure 2A). We find that there are parameter combinations which greatly improve efficiency of the signal transmission function. The basic distribution of a uniform low value of RC,S for fast dS and a wide variability in RC,S above a certain threshold in dS is robust against different types of signaling input (cf. also Figure 3A). This may, however, be highly dependent on the reaction network that underlies the transfer function. We have not further explored this question. To test for robustness of these systems against variability of concentration, we repeated experiments for 100 systems with 20% variation of original concentrations, which corresponds to generally accepted noise levels (cf.13,14). As expected, this low variation did not significantly affect the quality of a set of kinetic rates (Supplementary table).
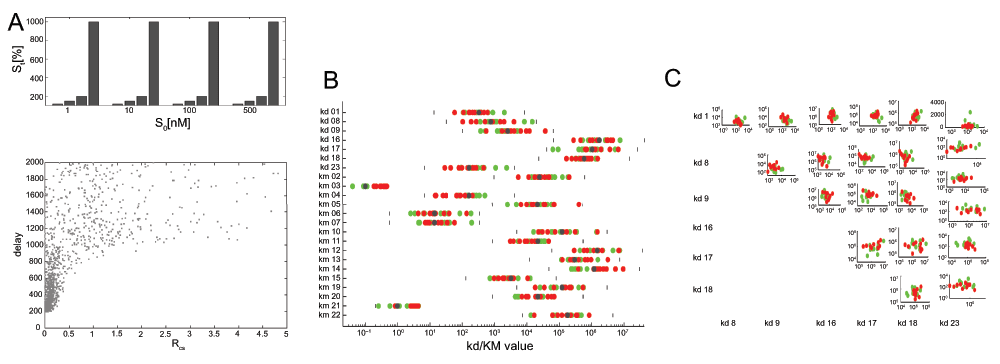
Figure 2.
A. Distribution of systems according to R and d values. ~ 1600 different systems states were randomly generated with k-parameter variations (20–500%), RC,S and d values were measured with the signal set shown on top. B. Kd and KM parameters for high efficiency (red, f < 2) and low efficiency (green, f > 5) systems. C. Cross-correlation of Kd values.
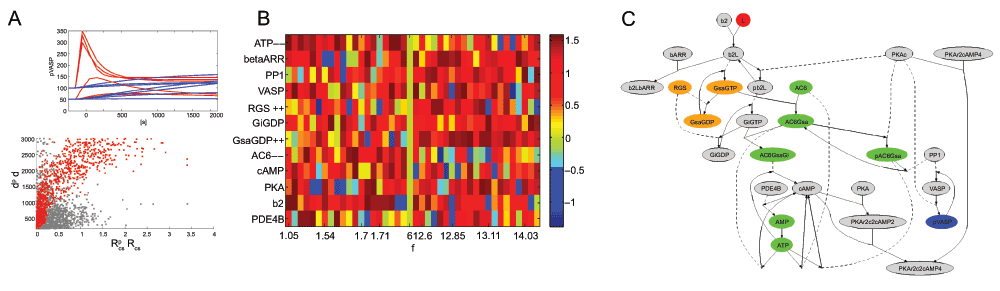
Figure 3.
A. (top) Dynamics of selected systems with low (ftransient < 3, blue) or high (ftransient > 10, red) propensity for a transient response. (bottom) Distribution of concentration parameters according to dp, (grey) or RC,S, d (red). B. Concentration changes in response to optimizing for transients, with plateau signaling (left) and strong transients (right), sorted by ftransient. C. Concentration shifts (orange=high, green=low) in the biochemical reaction network for a high transient system, as averaged from B. Conforming to intuition, we see that the earlier, driver complex (G-protein) is high for transients, which increase the on-slope for cAMP production.
We further analysed the parameter combinations with different signal transmission efficiency. In Figure 2B and C, we distinguish low and high efficiency signal transmission. Interestingly, we find, with respect to signal efficiency of the transfer function, that all parameters are “sloppy” (allow a wide variation), there are no “stiff” (low variation) parameters15. Standard deviation for all parameters in our case is similar (Figure 2B). Since it has been argued that optimization to experimental data yields reactions which allow more variance than others, as an indication of their influence on the signal transmission pathway, this analysis seems to contradict this effect. Possibly, these results pertain mostly to parameter variation that results from matching a networked system with many species to selected time-series data for only few species, which may behave differently from general optimization.
Co-regulation of protein concentration as an adaptive response to perturbation
Co-regulation of protein expression in cellular systems is important in disease progression and often a problem in targeted interventions. Here we are exploring the question of self-organization of protein concentration after a perturbation that reduces one protein to only 10% of its previous concentration. Keeping kinetic rates fixed, all concentrations in the system are allowed to adjust until optimality of signal transmission and delay is reinstated. There is a number of interesting observations here (cf. Figure 4A), which relate to the biological reality. For instance, reducing PDE4B causes much regulation in other proteins, but it is almost never targeted. In contrast, reducing PP1 has little effect, but PP1 is frequently responsive to other proteins. Reducing PKA, RGS and AC6 leads to widespread down-regulations, to maintain sensitivity of signaling, but reducing the receptor beta-2 leads to up-regulation.
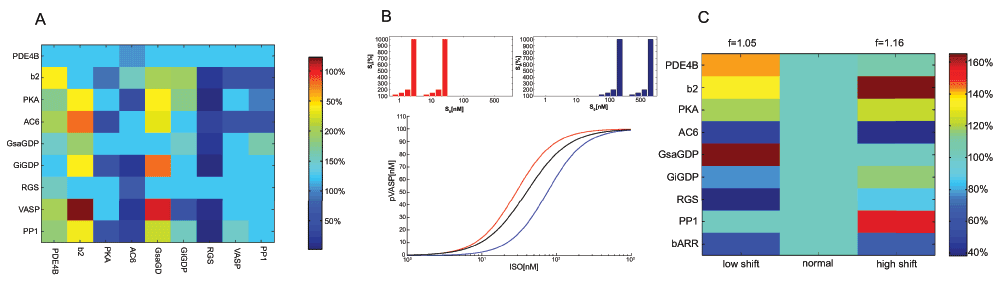
Figure 4.
A. Concentration changes in response to total protein concentration reduction. Selected concentrations were reduced to 10% (shown on the x-axis). Learning was applied until f values were improved (shown on top for each experiment). Colors show the relative adjustment of all concentrations on the y-axis. B. Transfer function response to shifts in input signal range (shown on top). C. Concentrations changes in response to extracellular signal shift. Matrix is constructed as in A.
There are many individual adaptations which can be interpreted to maintain the sensitivity and responsivity of the small molecule cAMP. The results show that protein regulation is highly sensitive to positions and roles of individual proteins in the reaction network in transmitting the signal. This problem may also be amenable to a more principled mathematical analysis16. Another idea would be to rank reaction systems defined by kinetic parameters as in Figure 2 by how well they adapt to perturbations.
In our example, the quality of the readjustment is not always the same, but the simple optimization scheme that we use may easily produce suboptimal solutions (local minima). Concentration changes may be caused by genetic up- or down-regulation, secretion and re-uptake, increased degradation, RNA interference, etc. and are therefore not easy to model from the standpoint of mechanistic biological modeling, which would need modules for all biologically attested processes. A unified perspective by a set of constraints and a set of objectives, such as has been envisaged here, may lead to better predictive results and may also be used as a guiding principle in constructing mechanistic models. Since there are intricate biological processes of adjusting concentrations, we may assume that in the cell optimal solutions are more easily found.
Optimal signal transmission depends on the signaling level
From the standpoint of disease modeling, an unusual protein concentration may be an adaptive response where a still functioning cell in a dysfunctional external signaling environment struggles to keep signal transmission efficient to support cellular function. In such a case, targeting this protein by pharmacological intervention will lead to co-regulation on other proteins. In general, as well as in real biological signaling systems, it is the localization of the input signaling range and the distribution of signals that the system transmits which are important for optimization. If we select the signal set that we optimize for in the right way, the input range will move towards the loglinear range, i.e. between the lower and upper input boundaries (Figure 4B). As a result, a number of internal concentrations will change. Figure 4C shows the relative concentration changes that result from a shift in input range. Interestingly, the low shift requires a strong reduction in RGS, and upregulation for PDE4B, among other effects. GsaGDP (the activating G protein) is strongly increased, and GiGDP (the inhibitory G protein) is decreased. In the high shift, GiGDP and PP1 (the phosphatase which decreases pVASP) are stronger, and the beta-2 receptor density is much increased. In the future, it will be interesting not only to calculate these effects based on different optimization measures, and in multiple input-output scenarios, but also to compare this with attested cases of biological adaptation. By reverse engineering, we may infer an optimization measure from a sufficient number of attested co-regulations. For instance in addiction, protein co-regulation as a form of sensitization is well-attested17, but also in cancer where intercellular signaling (e.g. by cytokines18) is affected. Gene expression data may then be mined not only for evidence of the mechanics of genetic regulatory pathways but also for evidence of shifts in extracellular signaling, which cause altered protein expression.
Role of transients
The appearance of a transient vs. a plateau signal (or even a dampened oscillation) in response to sustained signaling depends on the construction of the biochemical reaction network (negative feedback interactions) and its parameters. We show that we can also train a system for the appearance of a transient response to a sustained signal. This means to search for high response at peak, low delay to peak, but also a low response at steady-state (see Section “Objective function” and Figure 3A). This requirement of invariance for the steady-state is sometimes considered a form of “robustness” or “homeostatic regulation” of the cellular response19,20, and signaling by transients is widely regarded as an important mechanism. Here we found that concentration adjustment is quite sufficient to acquire a switch from transient to plateau response, with a high variability of response shape dependent on concentration, but also on the size of the signal (Figure 3A). In Figure 3B we show the distribution of concentrations that achieve high or low propensity for transients, as indicated by ftransient. We focus on concentrations with fairly uniform up- or down-regulation to create an experimental graph in Figure 3C. The simplicity of Figure 3C undermines the notion, as proven by the random search optimization, that deviations from up- or down-regulation for individual concentration may not be irrelevant noise, but rather part of a guided adaptation process that has many different solutions. We may have to consider this complexity to understand concentration adjustments in biological cells.
By using different objective functions for each target, it will be quite possible to combine different goals for targets in a multiple-output signaling system. Interestingly, a self-organizing cellular signaling system may also incorporate adaptive controls as objective functions, in particular when ion channels or membrane transporters are targeted. In this case, the goal of the system is to respond to an input (e.g. ISO at beta-2 receptors) such that the magnitude of another input such as calcium (ion channels as targets) is controlled. Signal transmission efficiency for such a target is not to be optimized to a maximum value, but instead to the appropriate ratio between the inputs. We have not followed up on this idea, but it may be worth further investigation as an example of natural computation for adaptive control.
Discussion
The idea to look for parametric optimization as the basis of realistic cellular properties has been applied with success to metabolic fluxes21. In that case, optimization of growth is usually regarded as the single objective function. Here we use another objective function, signal transmission efficiency, to study the adaptive response of a signaling system to perturbation in concentrations. This equals maximization of concentration change in a target species in response to input signal. Optimization of growth and optimization of signal transmission are therefore related.
Signal transmission in a cellular system may have multiple functions: transcription factor activation, which may be related to the cell cycle, to morphological change or to adjustment of protein concentrations, cytosolic kinase/phosphatase activation with multiple cellular targets, membrane protein activation such as ion channels or receptors, etc. We assume that signal transduction has been optimized by evolution, and that concentration adjustment exists to maintain effective signal transmission. We have shown how to optimize a single-input single-output system for both speed and signal efficiency due to the basic properties of kinetic equations6. It is easy to extend the present discussion to optimize for multiple outputs in parallel, and a system could also be optimized for a number of I/O functions. In that case, other measures, such as mutual information, may also be employed as objectives. We used optimization in a two-step process: (a) in evolutionary learning, in order to find kinetic rates for estimated concentrations (b) in cellular adaptation, in order to re-calibrate the model in response to perturbations in concentrations, changes in extracellular signaling, or to effect a transient vs. plateau-like response. This also means that we have transitioned from a biologically defined system that is built bottom-up from available parameters and data to a self-organizing, learning system that adjusts to changes on evolutionary or individual time-scales.
Matching models to experimental time-series data is an under-constrained problem that often yields large ranges of suitable parameters15,22. Analysing signals and targets to find optimal transmission parameters may be used to further constrain and investigate the parametric space. Signal transmission efficiency in a biological system can be measured directly23,24 and be compared to what is theoretically possible given a set of equations. During evolution, new protein subtypes develop with a different set of interactions and regulations. This corresponds to an adjustment of the available set of reactions, a type of structural learning to overcome the bottlenecks that are a result of tightly specified molecular kinetics and limited adjustment of concentrations.
Competing interests
No relevant competing interests disclosed.
Grant information
The author(s) declared that no grants were involved in supporting this work.
Supplementary table
Table 4. For 21 systems with RC,S > 0.5 and dS < 800, we re-calculate RC,S and dS < 800 from 20 variations on concentration values within a 20% interval.
Shown are the original, the mean and std values.
| System no | Basic RC,S
| Mean RC,S
| Std RC,S
| Basic dS
| Mean dS
| Std dS |
|---|
| 1 | 0.578 | 0.582 | 0.0311 | 757 | 779 | 41.8 |
| 2 | 0.623 | 0.623 | 0.0419 | 723 | 722 | 35.4 |
| 3 | 0.575 | 0.575 | 0.0483 | 732 | 740 | 34.89 |
| 4 | 0.782 | 0.792 | 0.0571 | 749 | 768 | 50.45 |
| 5 | 0.609 | 0.622 | 0.099 | 578 | 590 | 46.82 |
| 6 | 0.609 | 0.615 | 0.033 | 728 | 737 | 37.05 |
| 7 | 0.612 | 0.628 | 0.0636 | 659 | 663 | 18.43 |
| 8 | 0.672 | 0.666 | 0.061 | 658 | 662 | 25.36 |
| 9 | 0.703 | 0.715 | 0.049 | 782 | 812 | 44.66 |
| 10 | 0.655 | 0.651 | 0.0511 | 761 | 760 | 29.3 |
| 11 | 0.576 | 0.551 | 0.0698 | 649 | 638 | 44.94 |
| 12 | 0.54 | 0.56 | 0.108 | 664 | 669 | 20.91 |
| 13 | 0.694 | 0.695 | 0.043 | 692 | 695 | 22.5 |
| 14 | 0.504 | 0.501 | 0.0312 | 726 | 731 | 35.70 |
| 15 | 0.522 | 0.526 | 0.0594 | 620 | 621 | 24.49 |
| 16 | 0.8 | 0.806 | 0.0369 | 726 | 742 | 50 |
| 17 | 0.693 | 0.7 | 0.0341 | 717 | 724 | 24.03 |
| 18 | 1.24 | 1.29 | 0.221 | 761 | 779 | 59.1 |
| 19 | 0.651 | 0.63 | 0.0512 | 668 | 666 | 11.64 |
| 20 | 0.529 | 0.5 | 0.045 | 777 | 762 | 43.25 |
| 21 | 0.537 | 0.534 | 0.044 | 752 | 754 | 33.42 |
Faculty Opinions recommendedReferences
- 1.
Bhalla US, Iyengar R:
Emergent properties of networks of biological signaling pathways.
Science.
1999; 283(5400): 381–387. PubMed Abstract
| Publisher Full Text
- 2.
Hoops S, Sahle S, Gauges R, et al.:
COPASI--a COmplex PAthway SImulator.
Bioinformatics.
2006; 22(24): 3067–3074. PubMed Abstract
| Publisher Full Text
- 3.
Deng J, Jones C, Feinberg M, et al.:
On the steady states of weakly reversible chemical reaction networks. 2011; 1–20. Reference Source
- 4.
Akle S, Dalal O, Fleming RMT, et al.:
Existence of positive steady states for mass conserving and mass-action chemical reaction networks with a single terminal-linkage class. Report SOL, 2011; 3. Reference Source
- 5.
van der Schaft A, Rao S, Jayawardhana B:
On the Mathematical Structure of Balanced Chemical Reaction Networks Governed by Mass Action Kinetics. 2011. Publisher Full Text
- 6.
Scheler G:
Transfer functions for protein signal transduction: application to a model of striatal neural plasticity.
PLoS One.
2013; 8(2): e55762. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 7.
Barlow R, Blake JF:
Hill coefficients and the logistic equation.
Trends Pharmacol Sci.
1989; 10(11): 440–1. PubMed Abstract
| Publisher Full Text
- 8.
Ferrell JE Jr:
Signaling motifs and Weber's law.
Mol Cell.
2009; 36(5): 724–7. PubMed Abstract
| Publisher Full Text
- 9.
Buijsman W, Sheinman M:
Efficient fold-change detection based on protein-protein interactions. 2012. Reference Source
- 10.
Yi TM, Kitano H, Simon MI:
A quantitative characterization of the yeast heterotrimeric G protein cycle.
Proc Natl Acad Sci U S A.
2003; 100(19): 10764–9. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 11.
Blackman BE, Horner K, Heidmann J, et al.:
PDE4D and PDE4B function in distinct subcellular compartments in mouse embryonic fibroblasts.
J Biol Chem.
2011; 286(14): 12590–601. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 12.
Teukolsky SA, Vetterling WT, Flannery BP, et al.:
Section 10.5. Downhill Simplex Method in Multidimensions. In: Numerical Recipes: The Art of Sci. Computing, 2007. Reference Source
- 13.
Sigal A, Milo R, Cohen A, et al.:
Variability and memory of protein levels in human cells.
Nature.
2006; 444(7119): 643–6. PubMed Abstract
| Publisher Full Text
- 14.
Newman JR, Ghaemmaghami S, Ihmels J, et al.:
Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise.
Nature.
2006; 441(7095): 840–846. PubMed Abstract
| Publisher Full Text
- 15.
Gutenkunst RN, Waterfall JJ, Casey FP, et al.:
Universally sloppy parameter sensitivities in systems biology models.
PLoS Comput Biol.
2007; 3(10): 1871–78. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 16.
Steuer R, Waldherr S, Sourjik V, et al.:
Robust signal processing in living cells.
PLoS Comput Biol.
2011; 7(11): e1002218. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 17.
Robison AJ, Nestler EJ:
Transcriptional and epigenetic mechanisms of addiction.
Nat Rev Neurosci.
2011; 12(11): 623–637. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 18.
O’Shea JJ, Murray PJ:
Cytokine signaling modules in inflammatory responses.
Immunity.
2008; 28(4): 477–87. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 19.
Shinar G, Feinberg M:
Structural sources of robustness in biochemical reaction networks.
Science.
2010; 327(5971): 1389–91. PubMed Abstract
| Publisher Full Text
- 20.
Kitano H:
Towards a theory of biological robustness.
Mol Syst Biol.
2007; 3: 137. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 21.
Edwards JS, Ibarra RU, Palsson BO:
In silico predictions of Escherichia coli metabolic capabilities are consistent with experimental data.
Nat Biotechnol.
2001; 19(2): 125–30. PubMed Abstract
| Publisher Full Text
- 22.
Secrier M, Toni T, Stumpf MP:
The ABC of reverse engineering biological signalling systems.
Mol BioSyst.
2009; 5(12): 1925–1935. PubMed Abstract
| Publisher Full Text
- 23.
Cheong R, Rhee A, Wang CJ, et al.:
Information transduction capacity of noisy biochemical signaling networks.
Science.
2011; 334(6054): 354–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 24.
Brennan MD, Cheong R, Levchenko A:
Systems biology. How information theory handles cell signaling and uncertainty.
Science.
2012; 338(6105): 334–5. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 25.
Wittig U, Kania R, Golebiewski M, et al.:
SABIO-RK--database for biochemical reaction kinetics.
Nucleic Acids Res.
2012; 40(Database issue): D790–6. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 26.
Scheer M, Grote A, Chang A, et al.:
BRENDA, the enzyme information system in 2011.
Nucleic Acids Res.
2011; 39(Database issue): D670–76. PubMed Abstract
| Publisher Full Text
| Free Full Text
Comments on this article Comments (0)