Benslimane Y and Harrington L. The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.12688/f1000research.6664.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Department of Molecular Biology, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada 2Department of Biochemistry, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada 3Department of Medicine, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
Fluorescence microscopy can be used to assess the dynamic localization and intensity of single entities in vitro or in living cells. It has been applied with aplomb to many different cellular processes and has significantly enlightened our understanding of the heterogeneity and complexity of biological systems. Recently, high-resolution fluorescence microscopy has been brought to bear on telomeres, leading to new insights into telomere spatial organization and accessibility, and into the mechanistic nuances of telomere elongation. We provide a snapshot of some of these recent advances with a focus on mammalian systems, and show how three-dimensional, time-lapse microscopy and single-molecule fluorescence shine a new light on the end of the chromosome.
Oskar Heimstädt, who built the first fluorescence microscope, ended his 1911 paper with the following perspective: “If and to what degree fluorescence microscopy will widen the possibilities of microscopic imaging only the future will show”1. More than a century later, fluorescence microscopy has proven transformative in our ability to illuminate almost all aspects of cellular biology. One of the more recent frontiers in fluorescence microscopy is the resolution of biological phenomena at the single molecule level, called single molecule fluorescence2. Biological processes have evolved to be inherently heterogeneous, transient and dynamic3, and therefore difficult to track. Single molecule microscopy often permits a birds-eye view of ephemeral and complex mechanisms. This review will focus on selected recent advances in high-resolution microscopy including, but not limited to, single-molecule fluorescence microscopy, that have enlightened our understanding of chromosome ends and the enzyme that replenishes them.
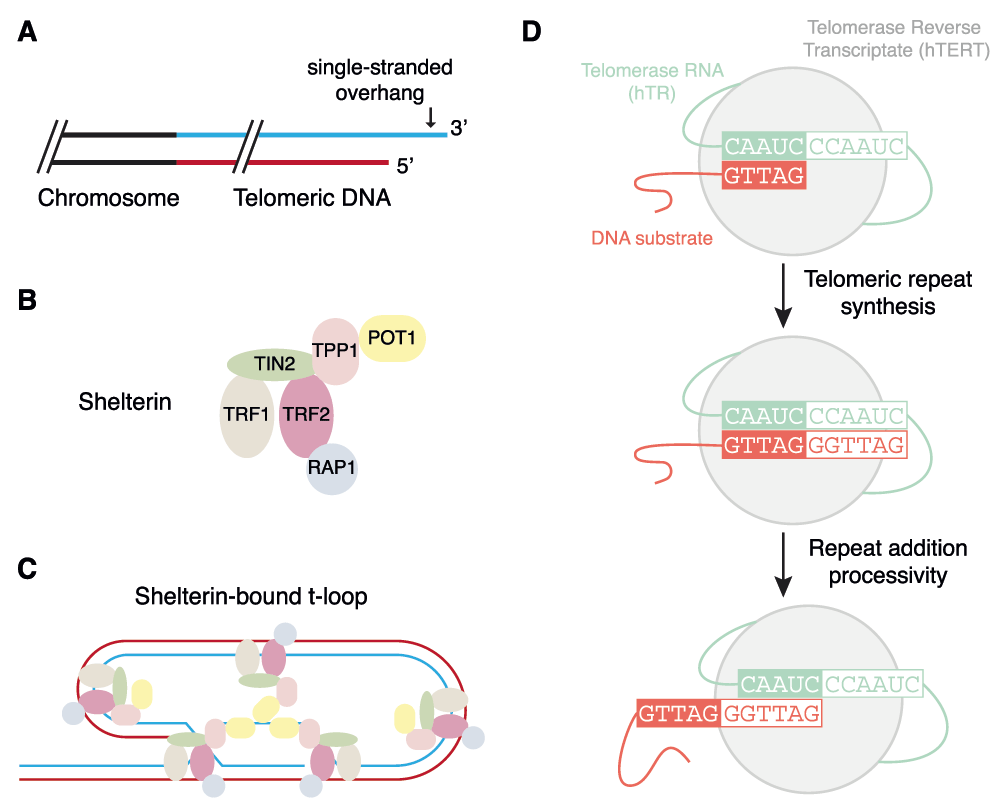
Many organisms must contend with the vulnerability of linear chromosome ends to enzymes that degrade, rearrange, or incompletely replicate DNA. This susceptibility to breakage, and their distinct “knob-like” appearance under the light microscope, made them an early target of study by scientists such as Barbara McClintock and Hermann Müller4. We now appreciate that telomeres are a highly specialized nucleoprotein structure whose maintenance is critical to genome stability (Figure 1A–C)5,6. Replenishment of the G-rich sequences that comprise the telomeres is carried out by telomerase, whose core components are a reverse transcriptase and an integral RNA that provides the telomere template (Figure 1D)7.
Figure 1. An overview of telomeres and telomerase.
(A) In many organisms, chromosome ends terminate in a single-stranded, G-rich overhang preceded by up to several kilobase pairs of double-stranded G-rich DNA. (B) Telomeres are capped by a six-subunit complex called shelterin. (C) Shelterin (particularly TRF2) promotes the formation of a higher order telomeric loop (T-loop) structure that serves to mask telomeres from the deleterious fates associated with a free DNA end5,6. (D) The catalytic cycle of the core telomerase enzyme, comprised of a protein (TERT) and RNA (hTR)7.
Zooming in on telomeres
Fluorescence microscopy has revolutionized our ability to probe the length, location, and recombination of telomeres in vivo through the application of fluorescently labeled peptide nucleic acids that bind tightly and specifically to telomeric DNA8–10. These techniques have uncovered interesting distinctions in the three-dimensional (3D) localization of mammalian telomeres in normal cells versus cancer cells11,12. Time-lapse microscopy has also revealed increased mobility of telomeres upon induction of a DNA break13 as well as unexpected long-range telomere associations in telomerase-negative cells after DNA breakage14. In the budding yeast Saccharomyces cerevisiae, fluorescent tagging of several components of telomerase have also provided considerable insight into the temporal and spatial dynamics of telomerase recruitment in living cells15–18.
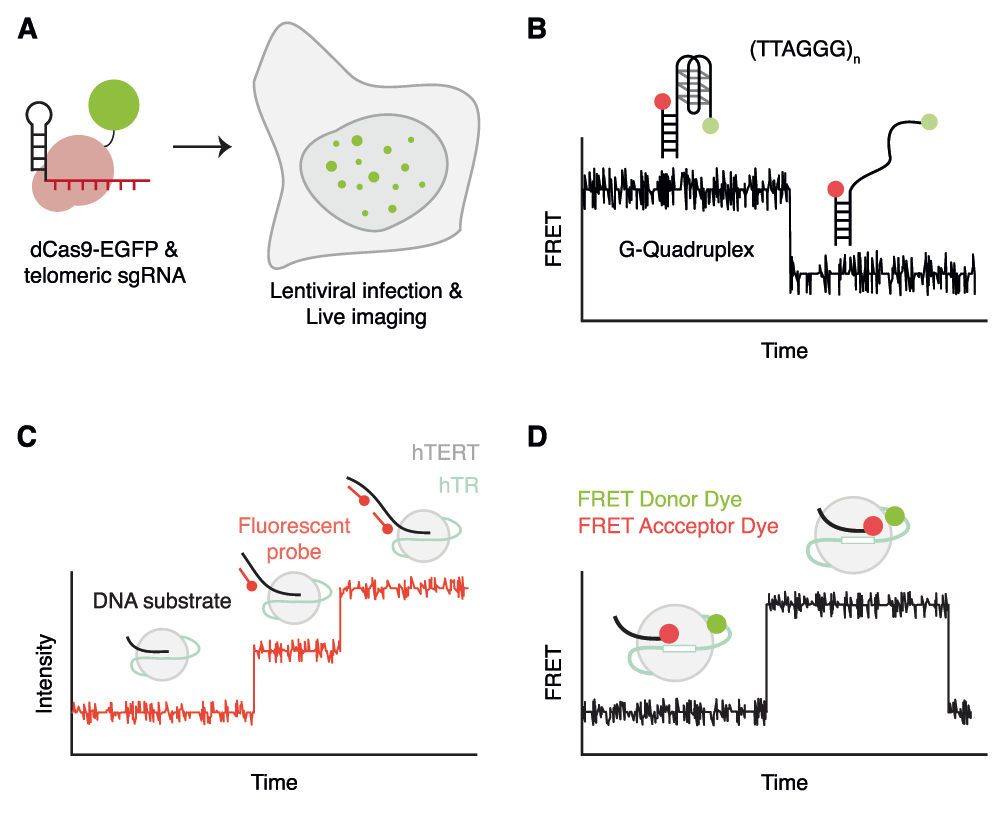
Fluorescence microscopy is also being combined with other leading-edge technologies to directly target specific genomic regions, including the telomere. Using the CRISPR/Cas system in which Cas9 can be guided to specific genomic loci using a small RNA19, Chen and colleagues targeted an enhanced green fluorescent protein (EGFP)-tagged catalytically inert Cas9 specifically to telomeres20. Telomere-recruited Cas9 demonstrated a punctate pattern that spatially overlapped with that of the telomeric DNA binding protein, telomeric-repeat binding factor 2 (TRF2). Furthermore, EGFP intensity correlated linearly with telomere intensities obtained using fluorescence in situ hybridization (FISH) (Figure 2A)20. This method could also be employed to visualize Cas9 directed to a single genomic locus20. Technological advances such as these may soon permit the ability to explore the spatial and dynamic localization of telomeres and telomerase in living mammalian cells to a similar extent as has been explored in budding yeast.
Figure 2. Selected recent advances in single-molecule microscopy that illuminate telomeres and telomerase.
(A) The use of a fluorescently tagged Cas9 and a guide RNA specific to telomeres to measure telomere dynamics and length in live cells20. (B) The ability of single-molecule Förster resonance energy transfer (smFRET) to measure the dynamics of G-quadruplex folding27,30–36,38. (C) The application of fluorescent probes complementary to telomeric substrates to measure the elongation properties of telomerase47. (D) The application of FRET to assess the intermolecular proximity of the DNA substrate and the RNA subunit of telomerase during telomere synthesis48.
The telomere terminus ends in a 3’ G-rich overhang of variable length that can invade the upstream double-stranded telomeric DNA to form a structure called a T-loop21. Super-resolution microscopy, specifically stochastic optical reconstruction microscopy (STORM), has recently enabled an unprecedented visualization of T-loops that are crosslinked and purified from murine cells22 (Figure 1B,C). The loss of T-loops occurred specifically upon loss of the shelterin subunit TRF2, and not other shelterin components, which provides an elegant demonstration that it is the T-loop structure that protects ends from the deleterious molecular events observed when TRF2 is removed from telomeres22,23.
Telomeric DNA sequences can also form in vitro intra- or inter-parallel structures called G-quadruplexes, and evidence is accruing to suggest their existence at telomeres and at interstitial G-rich regions24. Single-molecule Förster resonance energy transfer (smFRET) has been applied to the molecular dynamics of G-quadruplex formation in vitro25–28 (Figure 2B). The smFRET approach has several advantages that permit a high-resolution view of G-quadruplex dynamics. Firstly, this technique can resolve different conformations using FRET signal intensities and Gaussian curve fitting29. Secondly, individual single-molecule traces reveal dynamic switching between states in real time. Hidden Markov modeling can then extract the dwell time of each molecule, which is a reflection of the stability of each state29. Applying smFRET to the analysis of shelterin binding to telomeric substrates in vitro, several groups have investigated the intricate relationship between G-quadruplexes and POT1/TPP1 (protection of telomere 1/Pot1-interacting protein TINT1-PTOP-PIP1)30,31. Furthermore, this technique also revealed the binding and unfolding of G-quadruplexes by proteins such as RAD51, WRN, BLM, RecQ and RPA32–36. Recently, the use of smFRET combined with magnetic tweezers spectroscopy37 has been used to measure the thermodynamic properties of G-quadruplex folding38. These findings provide important insights into the dynamics of telomeric DNA structure in a more native nucleoprotein context.
Shining a light on telomerase
The telomerase reverse transcriptase, TERT, is able to synthesize new telomeric DNA, one nucleotide at a time, by virtue of an integral telomerase RNA that contains a short telomere-complementary sequence (Figure 1D). Although TERT shares several features in common with other viral reverse transcriptases, one unique aspect is its ability to repeatedly copy the same template for many cycles in an iterative process termed repeat addition processivity (RAP)39,40. Although much information has been gleaned using standard biochemical techniques regarding the complex DNA-protein, RNA-protein and protein-protein interactions that contribute to telomerase RAP (for a few current examples, see 41–46), it is only recently that single-molecule fluorescence has been brought to bear on the elongation properties of telomerase in vitro47. Hwang and colleagues immobilized immunopurified telomerase from cell extracts on a surface and employed a fluorescently labeled probe complementary to telomeric DNA to obtain a digital readout of telomerase activity in real time (Figure 2C). Their results suggested that active telomerase can exist in two dynamic states defined as an initial activation period followed by an extension period in which telomere elongation was visualized as a step-wise increase in fluorescence47. In addition, they found that TPP1-POT1 stimulated the elongation rate and overall product length47, consistent with previously described properties of TPP1-POT1. Although the nature of the fluorescent probe binding (which comprises the sequence 5′-CCCTAACCCTAACCC-3′) precludes single base resolution, this technique promises to provide an unprecedented, single-molecule view of the intricacies of the telomerase elongation cycle.
In another smFRET approach, the distance between the region 5’ of the RNA template of telomerase and a DNA substrate was followed during telomeric repeat synthesis (Figure 2D)48. Using different combinations of regular and chain-terminating nucleotides, information about the position of the DNA oligonucleotide substrate was registered at single-base pair resolution during elongation. While the substrate appears in a compact conformation after initial binding by telomerase, after a round of DNA synthesis the substrate realigns with the template via Watson-Crick base pair interactions. The aforementioned study by Parks and colleagues suggests that DNA dissociation and realignment is not the limiting step, and that an additional conformational adjustment is necessary after DNA:RNA repositioning to re-acquire a catalytically competent state48. This study illustrates the power of non-linear Gaussian curve fitting and hidden Markov modeling to extract the dwell time of the different FRET states from individual traces, which in turn permits a very precise dissection of the catalytic mechanism, albeit only over the short distances in which FRET can be observed.
smFRET has also been used to evince the real-time dynamics of folding of the pseudoknot domain with the telomerase RNA49–51. It has also been applied to examine the assembly and activity of the Tetrahymena thermophila telomerase RNP51–53 and, more recently, the role of the N-terminal domain of T. thermophila TERT in the stabilization of short RNA:DNA hybrids during telomerase catalysis54. In addition to valuable insights into telomerase catalysis, these techniques might also permit a precise elucidation of the mechanism-of-action of chemical modulators of telomerase activity, as well as an in vivo determination of human telomerase component stoichiometry, as was recently demonstrated for budding yeast telomerase18.
Future perspectives
In the year in which fluorescence microscopy was first described, Arthur Brisbane offered the sage advice: “Use a picture. It’s worth a thousand words.”55. We have reached a technological watershed in biology that will enable an unprecedented single-molecule and high-resolution view of the inner workings of many cellular machines. As we have illustrated here with but a few selected examples, fluorescence microscopy can be applied in many different ways to different problems, but key advances are the ability to dissect individual events instead of ensemble, population-based outputs, and to permit dynamic measurements in living cells, in real time. How far we have come from seeing telomeres as a cytogenetic “knob”, and how far we have yet to come.
Competing interests
The authors declare that they have no competing interests.
Grant information
The author(s) declared that no grants were involved in supporting this work.
Acknowledgments
We thank Paul Maddox, Tracy Bryan, Michael Stone, and Raymund Wellinger for comments and constructive input on the review.
Faculty Opinions recommended
References
1.
Heimstadt O:
Das Fluoreszenzmikroskop.
Z Wiss Mikrosk.
1911; 28: 330–7.
2.
Haustein E, Schwille P:
Single-molecule spectroscopic methods.
Curr Opin Struct Biol.
2004; 14(5): 531–40. PubMed Abstract
| Publisher Full Text
3.
Pross A:
On the emergence of biological complexity: life as a kinetic state of matter.
Orig Life Evol Biosph.
2005; 35(2): 151–66. PubMed Abstract
| Publisher Full Text
4.
Blackburn EH:
A History of Telomere Biology. In: de Lange T, Lundblad V, Blackburn EH, editors. Telomeres. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. 2006; 1–19. Reference Source
5.
de Lange T:
How shelterin solves the telomere end-protection problem.
Cold Spring Harb Symp Quant Biol.
2010; 75: 167–77. PubMed Abstract
| Publisher Full Text
6.
de Lange T:
Shelterin: the protein complex that shapes and safeguards human telomeres.
Genes Dev.
2005; 19(18): 2100–10. PubMed Abstract
| Publisher Full Text
7.
Autexier C, Lue NF:
The structure and function of telomerase reverse transcriptase.
Annu Rev Biochem.
2006; 75: 493–517. PubMed Abstract
| Publisher Full Text
8.
Aubert G, Hills M, Lansdorp PM:
Telomere length measurement-caveats and a critical assessment of the available technologies and tools.
Mutat Res.
2012; 730(1–2): 59–67. PubMed Abstract
| Publisher Full Text
| Free Full Text
9.
Williams ES, Cornforth MN, Goodwin EH, et al.:
CO-FISH, COD-FISH, ReD-FISH, SKY-FISH.
Methods Mol Biol.
2011; 735: 113–24. PubMed Abstract
| Publisher Full Text
10.
Molenaar C, Wiesmeijer K, Verwoerd NP, et al.:
Visualizing telomere dynamics in living mammalian cells using PNA probes.
EMBO J.
2003; 22(24): 6631–41. PubMed Abstract
| Publisher Full Text
| Free Full Text
16.
Gallardo F, Laterreur N, Cusanelli E, et al.:
Live cell imaging of telomerase RNA dynamics reveals cell cycle-dependent clustering of telomerase at elongating telomeres.
Mol Cell.
2011; 44(5): 819–27. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
21.
Griffith JD, Comeau L, Rosenfield S, et al.:
Mammalian telomeres end in a large duplex loop.
Cell.
1999; 97(4): 503–14. PubMed Abstract
| Publisher Full Text
24.
Murat P, Balasubramanian S:
Existence and consequences of G-quadruplex structures in DNA.
Curr Opin Genet Dev.
2014; 25: 22–9. PubMed Abstract
| Publisher Full Text
40.
Peng Y, Mian IS, Lue NF:
Analysis of telomerase processivity: mechanistic similarity to HIV-1 reverse transcriptase and role in telomere maintenance.
Mol Cell.
2001; 7(6): 1201–11. PubMed Abstract
| Publisher Full Text
41.
Berman AJ, Akiyama BM, Stone MD, et al.:
The RNA accordion model for template positioning by telomerase RNA during telomeric DNA synthesis.
Nat Struct Mol Biol.
2011; 18(12): 1371–5. PubMed Abstract
| Publisher Full Text
| Free Full Text
42.
Qi X, Xie M, Brown AF, et al.:
RNA/DNA hybrid binding affinity determines telomerase template-translocation efficiency.
EMBO J.
2012; 31(1): 150–61. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Steczkiewicz K, Zimmermann MT, Kurcinski M, et al.:
Human telomerase model shows the role of the TEN domain in advancing the double helix for the next polymerization step.
Proc Natl Acad Sci U S A.
2011; 108(23): 9443–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
44.
Tomlinson CG, Moye AL, Holien JK, et al.:
Two-step mechanism involving active-site conformational changes regulates human telomerase DNA binding.
Biochem J.
2015; 465(2): 347–57. PubMed Abstract
| Publisher Full Text
45.
Wu RA, Collins K:
Human telomerase specialization for repeat synthesis by unique handling of primer-template duplex.
EMBO J.
2014; 33(8): 921–35. PubMed Abstract
| Publisher Full Text
| Free Full Text
46.
Dalby AB, Hofr C, Cech TR:
Contributions of the TEL-patch amino acid cluster on TPP1 to telomeric DNA synthesis by human telomerase.
J Mol Biol.
2015; 427(6 Pt B): 1291–303. PubMed Abstract
| Publisher Full Text
| Free Full Text
1
Department of Molecular Biology, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada 2
Department of Biochemistry, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada 3
Department of Medicine, University of Montreal, Institute for Research in Immunology and Cancer, Montreal, Quebec, Canada
Benslimane Y and Harrington L. The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.12688/f1000research.6664.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Stone M. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11521)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Stone M. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11521)
Bryan T. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11520)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Bryan T. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11520)
Wellinger R. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11519)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Wellinger R. Reviewer Report For: The lighthouse at the end of the chromosome* [version 1; peer review: 3 approved]. F1000Research 2015, 4(F1000 Faculty Rev):1427 (https://doi.org/10.5256/f1000research.7160.r11519)
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)