Observation
Multi-species synchronous spawning of scleractinian corals is a feature reported from almost all speciose coral assemblages (Baird et al., 2009a; Baird et al., 2010; Raj & Edwards, 2010), including the Arabian Sea (Baird et al., 2014), and the Red Sea (Bouwmeester et al., 2011; Bouwmeester et al., 2014; Hanafy et al., 2010). A notable exception is at Eilat, on the Israeli coast, in the Gulf of Aqaba, where spawning is described as asynchronous with different species spawning in different seasons, on different months, and at different stages of the lunar cycle, with no overlap in spawning between species (Shlesinger & Loya, 1985; Shlesinger et al., 1998).
Here, we quantify the reproductive synchrony of Acropora corals in Dahab, on the Egyptian coast of the Gulf of Aqaba, 125 km south of Eilat, Israel (Figure 1). Among Red Sea reef habitats, fringing reefs in the Gulf of Aqaba support distinct coral assemblages with high cover and diversity of hard corals (DeVantier et al., 2000).
Figure 1.
Map of a the Red Sea, showing the location of previous work on scleractinian coral spawning in the Red Sea, and b the Gulf of Aqaba, showing Dahab, our study site.
The reproductive condition of 90 colonies from 15 Acropora species was assessed at two dive sites in Dahab, Um Sid (28° 25’14.16”N, 34° 27’27.52”E) and Eel Garden (28° 30’19.21”N, 34° 31’15.58”E), from the 2nd to the 4th of June 2014, a week before the full moon that month (Table 1). Acropora colonies at 1–10m depth were examined on snorkel by breaking 1–3 coral branches below the sterile apical zone to expose the developing oocytes. Colonies were recorded as mature when oocytes were visible and pigmented (Figure 2), immature when oocytes were visible and white, and empty when oocytes were too small to see with the naked eye or absent (following Baird et al., 2002). Colonies with mature oocytes are highly likely to spawn close to the night of the next full moon (in this case, the June full moon), whereas colonies with immature eggs are likely to spawn on or around a full moon one or two months later (in this case the July or August full moon). Colonies without oocytes have either already spawned or are unlikely to do so for at least three months.
Table 1. Percentage (%) of Acropora colonies with mature, immature, and no visible oocytes, on the 2–4 June 2014, in Dahab, Egypt, in the Gulf of Aqaba, Red Sea.
n: number of sampled colonies.
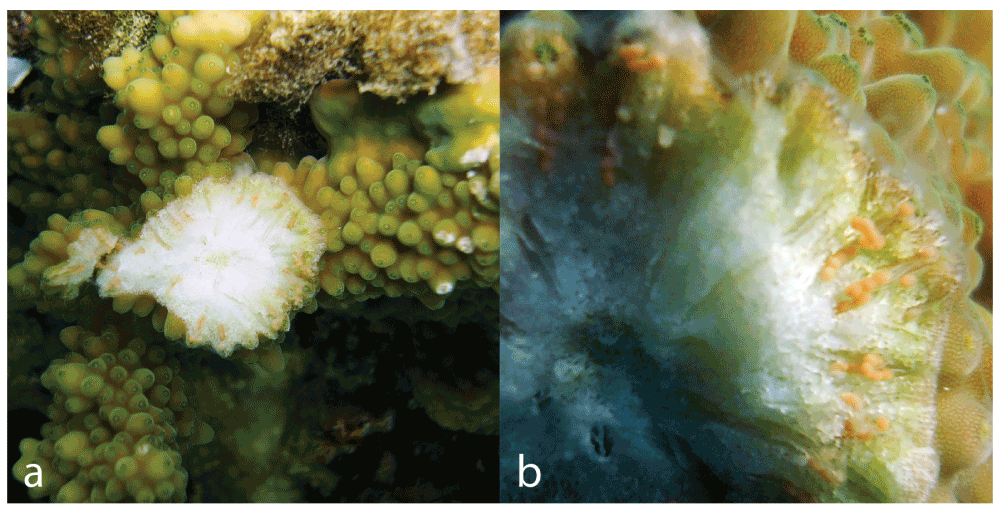
Figure 2.
a Exposed oocytes in a mature colony of Acropora variolosa b close-up of pigmented oocytes.
Seventy-one percent of Acropora colonies had mature oocytes and an additional three percent had immature oocytes (Table 1). No oocytes were observed in the remaining colonies. Fourteen out of 15 species had at least one colony with mature eggs, and in seven of those species, 100% of the sampled colonies had mature eggs (Table 1).
The reproductive condition in the Acropora assemblage at Dahab in June is very similar to that estimated in Acropora assemblages on the Egyptian coast of the northern Red Sea, where 85% of colonies from 12 species had mature oocytes in Marsa Alam in April 2008 and 99% of colonies from 17 species had mature oocytes in Hurghada in April 2009 (Hanafy et al., 2010). Subsequent sampling in both years revealed the absence of oocytes in all but one of these species, indicating that spawning had occurred sometime in the previous couple of weeks, most likely around the full moon of April (Hanafy et al., 2010). Nighttime observations in 2012 in Hurghada revealed spawning of 12 Acropora species over two consecutive nights around the full moon of May (Kotb, 2012). Similarly, 13 Acropora species in Thuwal, central Red Sea (Figure 1a), were observed to spawn together on the same night, both in April 2011 and in April 2012, following initial reproductive surveys which revealed 65% of mature Acropora colonies from 9 species in 2011 and 39% of mature Acropora colonies from 16 species in 2012 (Bouwmeester et al., 2014). The high percentage of species and colonies with mature oocytes in Dahab one week before the June full moon strongly suggests they will spawn synchronously as observed in Thuwal in the central Red Sea (Bouwmeester et al., 2011; Bouwmeester et al., 2014) and in Hurghada in the northern Red Sea (Hanafy et al., 2010; Kotb, 2012). Broadcast spawning of corals in most locations of the Indo-Pacific occurs as sea surface temperatures are increasing or when temperatures are close to their annual maxima (Baird et al., 2009a). In Dahab, waters start warming in the months of March-April, rising from 21–22°C to a maximum of 26–27°C in the month of August (Cornils et al., 2007; Plähn et al., 2002). Spawning in June most likely occurs when temperatures are ~24–25°C, possibly an optimum temperature for spawning and early larval development in the Gulf of Aqaba.
The month of spawning of Acropora species in the Gulf of Aqaba is two months later than in the northern and central Red Sea, where most Acropora spawn in April (Bouwmeester et al., 2014; Hanafy et al., 2010). This one or two-month offset is not surprising due to the difference in local temperature regimes and is similar to the latitudinal pattern observed along the east coast of Australia and from the Philippines to Japan (Baird et al., 2009b). Spawning in Dahab does not seem to occur before the waters reach 24–25°C, suggesting that a minimal temperature threshold is required during the warming of surface waters for spawning. In the central Red Sea, those temperatures are reached in March-April, and indeed multi-species spawning of Acropora has been recorded in April at 25–27°C (Bouwmeester et al., 2014).
Our data from Dahab match the data from Eilat (Shlesinger & Loya, 1985) for the timing of Acropora spawning in the Gulf of Aqaba, however, the larger number of Acropora species examined in the present study allows us to understand reproductive synchrony within this genus much more effectively. Indeed, we predict that with a comparable sampling protocol, similar levels of Acropora reproductive synchrony would be detected at Eilat, only 125 km north of Dahab, and would support the hypothesis that high levels of spawning synchrony are a feature of all speciose coral assemblages (Guest et al., 2005). The length of the scleractinian reproductive season can be established by sampling distantly related species from the coral assemblage, which in Eilat lasts four months for broadcast spawning species (Guest et al., 2005; Shlesinger & Loya 1985; Shlesinger et al., 1998), but sampling closely related species such as Acropora species will determine whether overlap in spawning occurs and will allow estimation of the level of synchrony in the assemblage.
Comments on this article Comments (0)