Keywords
cell-cell interactions, cadherin, adherin junctions, cell-cell adhesion, catenin, intracellular signaling pathways
cell-cell interactions, cadherin, adherin junctions, cell-cell adhesion, catenin, intracellular signaling pathways
The ability of cells to communicate and adhere to each other represents an ultimate prerequisite for the formation and maintenance of a multicellular organism. By sensing their microenvironment, cells can decide whether to continue or stop proliferating, change shape, accept a new identity, move out of the neighborhood, or simply cease to exist. How do the external signals get transmitted inside and prompt the cells to respond accordingly? In the past several years, cadherin-catenin protein complexes emerged as important regulators of morphogenesis and adult tissue homeostasis, linking cell-cell adhesion to multiple major signaling networks. In this short review, we will focus on the most recent studies that address the mechanisms and the functional relevance of the cadherin-mediated intracellular signaling.
Cadherin-catenin complexes comprise the core of a specialized type of adhesion junction named an adherens junction (AJ) (Figure 1). Among the family of classic cadherins, which includes E (epithelial)-, N (neural)-, P (placental)-, VE (vascular-endothelial)-, R (retinal)-, and K (kidney)-cadherins, E-cadherin is the most frequently employed in the formation of AJs in epithelial cells. To initiate the adhesion process, extracellular domains of cadherins engage in the Ca2+-dependent homophilic trans-interaction with identical cadherin molecules on an adjacent cell, while their cytoplasmic tails bind to p120- and β- (or its homolog γ-) catenin proteins. In turn, β-catenin interacts with α-catenin, which contains an actin-binding domain and physically links AJ complexes to the actin cytoskeleton1,2. Interaction between the actomyosin cytoskeleton and the AJs is prominently regulated by the mechanical forces and Rho-family of small GTPases (covered in detail in 3–6). This regulation is necessary for proper tissue morphogenesis and is highly dynamic, facilitating not only the coupling but also the detachment of cadherin-catenin complexes from actomyosin cytoskeleton, allowing cell-cell separation, cell sorting, and cell migration.
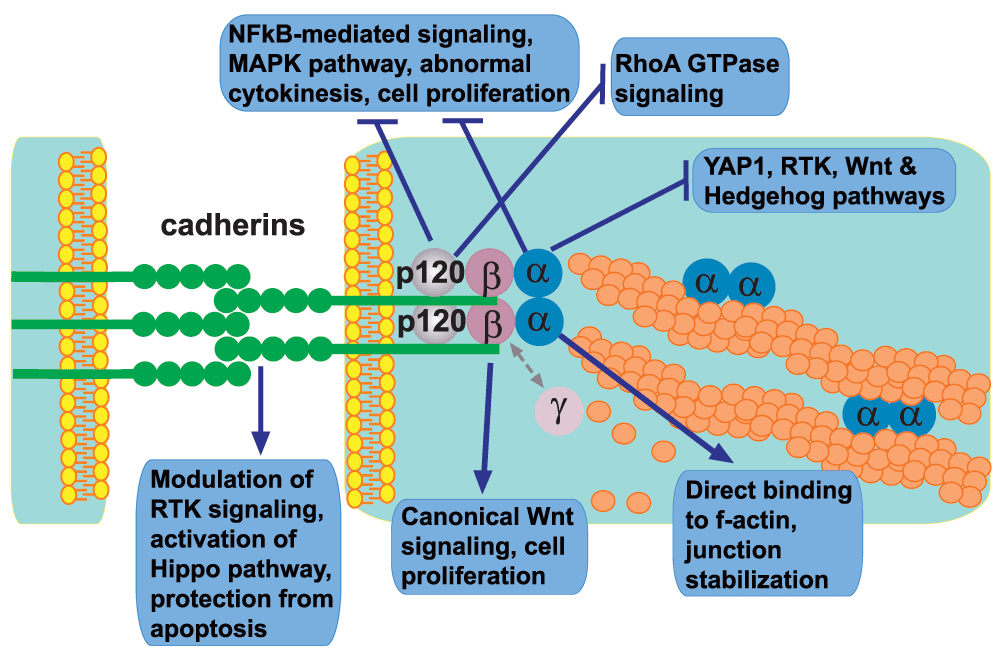
The diagram depicts protein members of the adherens junctions clustered at the plasma membranes of two juxtaposed cells and summarizes their individual roles in the intricate network of intracellular signaling pathways. Note that, despite their unique structural features and separate functions, both cadherins and catenins often work in concert and may also participate in the regulation of the same signaling pathway though via a distinct mechanism. Abbreviations: MAPK, mitogen-activated protein kinase; NFκB, nuclear factor-kappa-B; RTK, receptor tyrosine kinase; YAP1, yes-associated protein 1.
The ability of cadherins to transmit signals from the extracellular microenvironment inside the cell body is likely a direct consequence of their adhesive function, which stimulates clustering of cadherin molecules involved in AJ formation. In cell culture experiments, formation of a confluent cell monolayer results in prominent clustering of cadherin-catenin molecules at the AJs. This clustering not only strengthens cell-cell adhesion but also provides important cues for apical-basal cell polarization and significantly influences the downstream signaling events (for review, see 3,5,7). It was noticed a long time ago that formation of a confluent cell monolayer results in cell cycle withdrawal8. This phenomenon is known as “contact inhibition of cell proliferation”7. Re-expression of E-cadherin in human epithelial cancer cell lines that lack E-cadherin expression or disruption of E-cadherin with neutralizing antibodies in cell lines that maintained endogenous E-cadherin demonstrated that cadherin-mediated cell-cell adhesion plays a pivotal role in execution of contact inhibition of cell proliferation9. Similarly, activation of cadherin-catenin-mediated cell-cell adhesion by re-expression of α-catenin in a carcinoma line that was missing endogenous α-catenin resulted in retardation of cell proliferation10. A negative impact of E-cadherin expression on tumor progression was also revealed in genetic mouse experiments in vivo11. Since restoration of cadherin-catenin-mediated cell-cell adhesion results in prominent changes in cell morphology and re-establishment of apical-basal cell polarity, these early experiments were unable to determine whether cadherin clustering plays a direct or indirect role in negative regulation of cell proliferation. This question was later addressed by elegant experiments in Dr. Gumbiner’s laboratory, which demonstrated that clustering of cellular cadherins by E-cadherin-coated extracellular beads is sufficient to induce proliferation inhibitory signaling, thus directly implicating cadherin clustering in cell signaling events12.
How do cadherins exert their signaling functions? Multiple signaling molecules are located at the cell-cell contact sites in direct proximity to the AJ complexes. Many growth- and proliferation-promoting signaling pathways are initiated at the cell surface by receptor-type tyrosine kinases (RTKs). Cadherins can physically interact with several RTKs and they prominently impact their signaling abilities. For example, E-cadherin associates with epidermal growth factor receptor (EGFR) and negatively regulates its kinase activity12–14. Tumor-suppressor protein neurofibromatosis type 2 (NF2 or Merlin) promotes association between E-cadherin and EGFR, links EGFR to the cortical actin cytoskeleton, and blocks its internalization, which is necessary for EGFR activation and signaling15,16. Loss of Merlin in mouse liver results in prominent activation of EGFR signaling, expansion of progenitors, and development of liver cancer17. In addition to EGFR, E-cadherin can also negatively impact signaling of other RTKs, including ErbB2, insulin-like growth factor receptor (IGFR), and c-Met14. Similar to E-cadherin in epithelial cells, VE-cadherin in endothelial cells interacts with vascular-endothelial growth factor receptor 2 (VEGFR2) and negatively regulates its mitogen-activated protein kinase (MAPK) signaling by preventing the clathrin-dependent internalization of VEGFR2 and promoting the association between VEGFR2 and tyrosine phosphatase PTPRJ, which dephosphorylates and inactivates VEGFR218,19.
It is important to note that in some cases cadherins can promote growth factor receptor signaling. For example, N-cadherin stimulates fibroblast growth factor receptor signaling by preventing ligand-induced receptor internalization20. Both E-cadherin and VE-cadherin can promote PI3-kinase (PI3K) signaling and protect cells from apoptotic cell death21,22. VE-cadherin associates with the transforming growth factor-beta (TGF-β) receptor complex and potentiates cell proliferation inhibitory TGF-β signaling events23.
By acting at the plasma membrane, cadherins are ideally positioned to attract and retain their cytoplasmic partners, thus modulating their activation, stability, or nuclear accumulation or a combination of these.
This is important because some of these intracellular proteins are pivotal signaling molecules in their own right. For example, β-catenin is a very potent transcriptional co-activator and a key member of the canonical Wnt signaling pathway (for review, see 24–26). The levels of cytoplasmic β-catenin available for signaling are tightly controlled by the activity of the β-catenin-destruction protein complex, which is inhibited by activation of Wnt signaling24,25. Sequestration of β-catenin at the cell junctions can attenuate its ability to enter the cell nucleus and participate in transcriptional regulation. Indeed, multiple studies demonstrated that the loss of cadherin-mediated cell adhesion can promote β-catenin release and signaling26. The exact relationship between cadherin-mediated adhesion and β-catenin signaling is highly complex and context-dependent. In some cases, not only do cadherins not inhibit but they actually potentiate the β-catenin signaling pathway (for review, see 27).
Similarly to β-catenin, cadherins can sequester at the plasma membrane and prevent cytoplasmic accumulation of another member of AJs, p120-catenin (for review, see 28). p120-catenin binds to the transcriptional repressor KAISO and inhibits its function29–31. In addition, p120-catenin is a potent regulator of Rho-family GTPases and the nuclear factor-kappa-B (NFκB) signaling pathway28,32. p120-catenin is critical for stabilization of cadherin-catenin complexes and formation of AJs, and this function is likely to be responsible for its tumor-suppressor function in squamous cell carcinoma (SCC), which was revealed by genetic loss-of-function experiments in mice33.
α-catenin is crucial for AJ formation because it is necessary for the direct linkage of cadherin-catenin complexes at the membrane with the actin cytoskeleton34. Although there are three α-catenin genes in mammalian genomes (alpha E-catenin CTNNA1, alpha N-catenin CTNNA2, and alpha T (testis)-catenin CTNNA3), most epithelial cells express only one α-catenin (CTNNA1), and the knockout of this gene is usually sufficient for the complete loss of AJ function and loss of cell polarity35,36. This is different from inactivation of E-cadherin or β-catenin, which may often have redundant functions in the AJs because of the expression of other cadherins and γ-catenin. Notably, this is not the case in the adult heart, where inactivation of all expressed alpha-catenins (Ctnna1 and Ctnna3) does not cause a severe cell adhesion defect comparatively to N-cadherin knockout mice37,38.
Similar to p120 catenin (Ctnnd1), genetic loss-of-function experiments in mice revealed prominent tumor-suppressor activity of epithelial α-catenin (Ctnna1), as epidermal stem cell-specific deletion of α-catenin in mice results in the development of SCC tumors35,39,40. Like p120-catenin, α-catenin has been linked to NFκB signaling pathway in skin39 and in E-cadherin-negative basal-like breast cancer cells41, where it interacts with and stabilizes IκBα by preventing its ubiquitylation and association with proteasomes41. In addition to its critical role in cell-cell adhesion, via direct interaction with the dynactin protein complex, α-catenin can regulate dynactin-dynein-mediated traffic and integrate the microtubule and actin cytoskeletons during intracellular trafficking events42.
Loss-of-function experiments in vivo and in vitro revealed an important role of α-catenin in regulation of several major signaling networks, including Ras-MAPK35, canonical Wnt27,43, and Hedgehog44 pathways. Since α-catenin acts as a tumor suppressor in skin epidermis, our laboratory performed a small interfering RNA (siRNA) screen for genes necessary for this function in keratinocytes, which revealed a connection between α-catenin and yes-associated protein 1 (YAP1), a pivotal target of the Hippo signal transduction pathway40. The connection between cadherin-catenin proteins and the Hippo pathway components has been demonstrated by multiple studies and we will discuss these findings in more detail45–47 (see below).
First identified in Drosophila, the Hippo signaling pathway is evolutionarily conserved and functions as a key regulator of organ size and tumorigenesis by inhibiting cell proliferation and promoting (and, in some cases, inhibiting) apoptotic cell death (for review, see 48,49). In vertebrates, the core of the canonical Hippo pathway consists of two sequentially acting sets of kinases, MST1/2 and LATS1/2 (Hippo and Warts in Drosophila), and several associated co-activators and scaffold proteins. The MST1/2 kinases phosphorylate and activate LATS1/2, which in turn phosphorylates the growth-promoting transcriptional co-activator YAP1 (Yorkie in Drosophila) and its homolog TAZ (also known as WWTR1), leading to their cytoplasmic retention. When the Hippo pathway is inhibited, YAP1 translocates to the nucleus, where it binds multiple transcriptional factors and promotes their transcriptional activity48,49. It is important to note that, in addition to the canonical Hippo signaling pathway, YAP1/TAZ nuclear localization and activity can be regulated independently from MST1/2 and LATS1/245,50,51. In both Drosophila and mammalian model systems, the Hippo signaling is exquisitely sensitive to changes in the actin cytoskeleton or cellular tension which functions as a pivotal regulator that integrates and transmits upstream signals to the Hippo signal transduction pathway (for review, see 49,52). Increase in F-actin and actomyosin contractility blocks Hippo signaling and prominently activates Yorkie/YAP1/TAZ51,53.
For a long time, it remained largely unknown whether extracellular cues play any role in activating the Hippo pathway in mammals. The identity of the upstream transmembrane receptors responsible for transmitting the external signals inside the cell was undetermined. Elegant experiments in Dr. Guan’s laboratory identified G-protein-coupled receptors as important upstream regulators of Hippo signaling in mammalian cells54. The evidence that the nuclear localization and activity of YAP1 are inversely correlated with cell density55 pointed in the direction of the cell-cell junctions as potential upstream regulators of the Hippo signaling pathway. Indeed, it was recently demonstrated that E-cadherin homophilic binding at the cell surface in mammalian MDA-MB-231 cells is sufficient to control the subcellular localization of YAP1 independently of other cell interactions46. In addition, two recent studies using primary mouse keratinocytes revealed that α-catenin can bind to YAP1 and sequester it in the cytoplasm, thus modulating the level of YAP1 phosphorylation and its activity40,45 (for review, see 56,57). Importantly, there was an inverse correlation between α-catenin levels and nuclear YAP1 localization in both cultured keratinocytes and human SCC tumors, indicating that α-catenin may act as an inhibitor of YAP1 both in vitro and in vivo40. Of interest, although Ca2+ depletion, which abolishes cadherin homophilic interactions, triggered translocation of YAP1 into the nucleus, the depletion of E/P-cadherin or β-catenin in cultured keratinocytes did not affect the cellular localization of YAP145, pointing at the possibility that the expression of other cadherins and catenins might be sufficient to maintain AJs in E/P-cadherin or β-catenin knockdown keratinocytes.
In addition to α-catenin, β-catenin interacts with YAP1 and these proteins prominently impact each other’s nuclear localization and activity47,58,59. Constitutive activation of β-catenin in human cancer cells results in the formation of a β-catenin-YAP1-TBX5 transcriptional complex, which is essential for cancer cell survival60.
In Drosophila, the Hippo pathway can be regulated by multiple upstream transmembrane modules, which include atypical cadherins Dachsous and Fat (for review, see 61). Recently, another AJ protein, Echinoid, was shown to activate Hippo signaling via its physical interaction with and stabilization of the Hippo-binding partner Salvador62. This interaction is triggered by cell-cell contacts and requires the dimerization of Echinoid cytoplasmic domain. It is of interest to mention that, although there is no known Echinoid homolog in mammals, this protein is able to interact with Drosophila E-cadherin, thus contributing to the formation and maintenance of AJs63. Overall, although there are a lot of similarities between Drosophila and mammalian Hippo signaling pathways, at least some of the upstream regulators may be quite different64. Drosophila Yorkie is missing the C-terminal PDZ-binding motif, which is necessary for the connection between YAP1/TAZ and tight junction (TJ) proteins in mammalian cells. Although α-catenin is a potent negative regulator of YAP1 in mammalian cells38,40,45,46,65, it is a positive regulator of Yorkie in Drosophila66,67. While E-cadherin is a cell autonomous-positive regulator of Hippo pathway in mammalian cells46, it is a cell autonomous-negative regulator of Hippo in Drosophila67. Fat4, the mammalian ortholog of Drosophila fat gene, does not regulate the Hippo pathway in mouse liver, the organ highly sensitive to changes in the canonical Hippo signaling pathway64. However, mammalian FAT4 and Dachsous cadherins appear to negatively regulate YAP1 in neural progenitor cells68,69, indicating that at least some of the important connections in Hippo signaling may be tissue- and species-specific.
As discussed above, one of the ways for cadherins to regulate contact inhibition of cell proliferation is by antagonizing the activity of a variety of RTKs, including the EGFR. Interestingly, changes in RTK activity may indirectly impact Hippo signaling. For example, it was recently demonstrated that, in immortalized mammary cells, EGF treatment triggers the nuclear accumulation of YAP1 through activation of PI3K and phosphoinositide-dependent kinase (PDPK1) and this is largely independent of AKT signaling70. Interestingly, in Drosophila, EGF signaling also inhibits the Hippo pathway but through a different mechanism, which uses MAPK and the inhibitor of Warts, Jub71. Taken together, those findings point at the important connection between AJs, mitogenic factor pathways, and growth-inhibitory Hippo signaling. Of note, the Drosophila Jub was also shown to associate with α-catenin in a cytoskeleton tension-dependent manner, thus linking the actomyosin cytoskeleton, regulation of Hippo pathway activity, and AJs66.
In addition to the AJs, cadherin-mediated adhesion plays an important role in the formation of TJs and the apical-basal cell polarity domains. In turn, the polarity complex proteins can interact with structural components of both AJs and TJs, thus potentially centralizing the regulation of several signaling pathways (for review, see 72), although it is possible that the AJs and cell polarity regulate the Hippo signaling via multiple, genetically separable mechanisms67. The TJ-associated proteins angiomotin and angiomotin-like 1 and 2 directly interact with YAP1/TAZ, localize them to the cytoplasm and TJs, and negatively regulate their transcriptional activity73–76. Remarkably, at least in some cases, angiomotin proteins promote YAP1 activity by antagonizing YAP1-LATS2 interaction and increasing YAP1 dephosphorylation and translocation to the nucleus77. Interestingly, via its interaction with Merlin, angiomotin can localize to the AJs and facilitate AJ-specific recruitment and activation of LATS78. In both Drosophila and mammals, Merlin promotes Hippo signaling by targeting LATS to the cell membrane79. However, since angiomotin proteins are missing in the Drosophila genome, the angiomotin-mediated localization and activation of LATS at the AJs are likely to be species-specific, and this may potentially explain the differences in AJ-mediated regulation of YAP1 between Drosophila and mammalian model systems.
The unique aspect of cadherin-mediated signaling is that the clustering of cadherin molecules is mediated by the direct cell-cell contacts. This enables cells to identify and map the positions of their immediate neighbors, helping to integrate individual cells into the tissues not only at physical but also at biochemical levels. Although we are continually learning about novel aspects of cadherin-mediated signaling, it is clear that the unifying picture is still not within reach. Knowledge remains highly fragmented with distinct and frequently seemingly opposite findings generated in different model organisms, tissues, or cell culture conditions. Future studies are clearly necessary to accumulate more data in the hope that the sheer quantity of information will inevitably result in a qualitative change in our understanding of how individual cells use their cell-cell adhesion structures to coordinate their behavior in building and homeostatic maintenance of multicellular organisms.
AJ, adherens junction; E, epithelial; EGF, epidermal growth factor; EGFR, epidermal growth factor receptor; MAPK, mitogen-activated protein kinase; N, neural; NFκB, nuclear factor-kappa-B; P, placental; PI3K, PI3-kinase; RTK, receptor tyrosine kinase; SCC, squamous cell carcinoma; TGF-β, transforming growth factor-beta; TJ, tight junction; VE, vascular-endothelial; VEGFR2, vascular endothelial growth factor receptor 2; YAP1, yes-associated protein 1.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)