Keywords
connective tissue, embryology, anatomy and physiology, movement patterns, fascial manipulation.
This article is included in the Fascia research collection.
connective tissue, embryology, anatomy and physiology, movement patterns, fascial manipulation.
Before being ejected from the ovarian follicle, the ovum is covered by a double sac: theca interna and theca externa (Moore & Persaud, 1999). Once released, the ovarian follicle is covered — like every other cell — by a phospholipid bilaminar membrane, which acts as a double sac surrounding the content of the cell. During ovulation, the ovum released from the follicle is surrounded by another membrane, a lining of mucopolysaccharides’ translucent gel called zona pellucida. After fertilization, the ovum divides itself an indefinite number of times within the zona pellucida. This shell of ‘fundamental substance’ covering the zygote constitutes the first meta-membrane of the organism. This is the first connective tissues product to fulfill this function, followed by the reticulum and collagen fibroid components. This ooze is the primordial organismic membrane as well as the original organismic environment. A little amount of cytoplasm leaves the two daughter cells in the process of the first division and it creates a thin layer of fluid which surrounds the two cells and between the cells and the zona pellucida. (Moore & Persaud, 1999). This is the interstitial or lymphatic fluid, the principal means of exchange in the community of cells within the organism. The cells keep dividing and five days later they form a hollow sphere of cells called the blastocyst. During the following two weeks the blastocyst invaginates and undergoes gastrulation, a process in which pseudopods form in each corner of the blastocyst that join with other cells to make a whole, then a crater, and finally a tunnel, differentiating into two layers: an internal and an external one (Moore & Persaud, 1999). Thus, three spaces are created: the inside of the internal sac, the environment of the external sac and a space in between these two. When invagination is complete, it gives rise to two double sacs: the amniotic sac and the yolk sac surrounded by three germinal disc layers consisting of Ectoderm, Mesoderm and Endoderm (Figure 1).

These layers are intertwined between the two sacs in the following way:
The Ectoderm will form the nervous system, neural networks and skin in contact with the amniotic sac and fluid.
The Mesoderm will form the muscles, the connective tissues (so being the precursor of the fibrous net), the blood, lymph, kidneys, part of the genital organs and the adrenal cortex glands. It is located in the middle of the ectoderm and the endoderm.
The endoderm will form the tracts of our blood vessels, the organs of the digestive system and the glands. It is in contact with the yolk sac.
We will now focus on the formation of the fascial network within the mesoderm or the middle layer. During the second week of embryological development, there is hardly any cellular differentiation; almost all cells are exact copies of each other. However, as the embryo develops, growing at an exponential rate, there is also further specialization which means that spatial reorganization becomes essential (Figure 2). At the center of the mesoderm there is a thickening called the notochord, which gives rise to the spinal column, vertebrate bodies and intervertebral discs. To the sides of the paraxial mesoderm there is a special section known as mesenchyme (literally ‘tissue in the middle’) (Snyder, 1975). Mesenchymal stem cells are progenitors of fibroblasts and other connective tissue cells. Mesenchymal stem cells migrate among cells to settle in the three linings and they secrete reticular fibers (a primitive form of collagen) into the interstitial space. These fibers are chemically connected, much like velcro, to form a network that covers all the body (Schultz & Freitis, 1996). The mesenchymal stem cells are scattered throughout the body in order to differentiate themselves into other cell types where needed. They are a perfect example of the remarkable ability of connective tissue to adapt and respond to the continuous changes of the organism. Mesenchymal cells gradually replace the reticular fibers with the collagen fibers forming an only one fibrous net in the organism. That is the reason why favor the singular “fascia” over the plural “fasciae” (Myers, 2009). Once the three layers and the fascial cohesive network are established, it begins to fold onto and over itself repeatedly. The mesoderm covers the endoderm reaching to the ventral zone, where, from its in-between position, it gives rise to the ribs, the abdominal muscles and the pelvis. At the same time, it lodges and supports the endodermic digestive tract within (Cinnamon, 1999). The mesoderm moves to the dorsal zone where it forms the neural arch of the spinal cord as well as the cranial cavity that covers and protects the Central Nervous System (CNS) (Duprez, 2002). The palate is one of the last folds to connect together (when this process is not completed, a cleft lip results) (Moore & Persaud, 1999). In the musculoskeletal system, the internal sac fixed to bone is referred to as the periosteum, but when it is a fibrous cuff surrounding an articulation it is known as an articular capsule. These connective tissue structures are always connected and they are continuous with one another throughout the body. The external sac forms, deforms, reforms and stabilizes different tensional lines in the intermuscular septum or investing fascia according to the growth and movement to which they are subjected. The tissue’s tensional lines and the direction of the muscular fibers start from this process (Myers, 2009). The pattern of Tcf4 expression will determine how the muscles of the limbs within the mesoderm will be formed. From the beginning, the limbs will be organized according to these characteristics: from proximal to distal, from dorsal to ventral and from anterior to posterior. The signal that Tcf4-expressing cells give at a local level is through the secretion of inductive factors, or also through the deposition of an instructive extracellular matrix to induce myogenic precursors to distinguish into myotubes, which are in specific areas in the limb, serving as the nucleus for the anatomical formation of a future muscle. Following these primary signals, secondary signals join and are integrated to produce an upregulation of Tcf4 in specific areas of the limb mesoderm (Blasi et al., 2015; Kardon et al., 2003). Connective tissue controls the patterns of muscle formation by guiding the migration of myoblasts through the mesoderm (Brand-Seberi & Christ, 1999). Towards the end of the fifth week, future muscular cells are grouped together into two portions: a smaller epimere portion, organized in myotomes, and a larger hypomere portion in the ventral region. These portions subsequently form the epaxial and hypaxial muscles, respectively. Nerves that traverse segmental muscles, are also divided into a primary dorsal ramus for the epimere and a ventral ramus for the hypomere. They accompany the muscles throughout their migratory path (Figure 3). In particular:

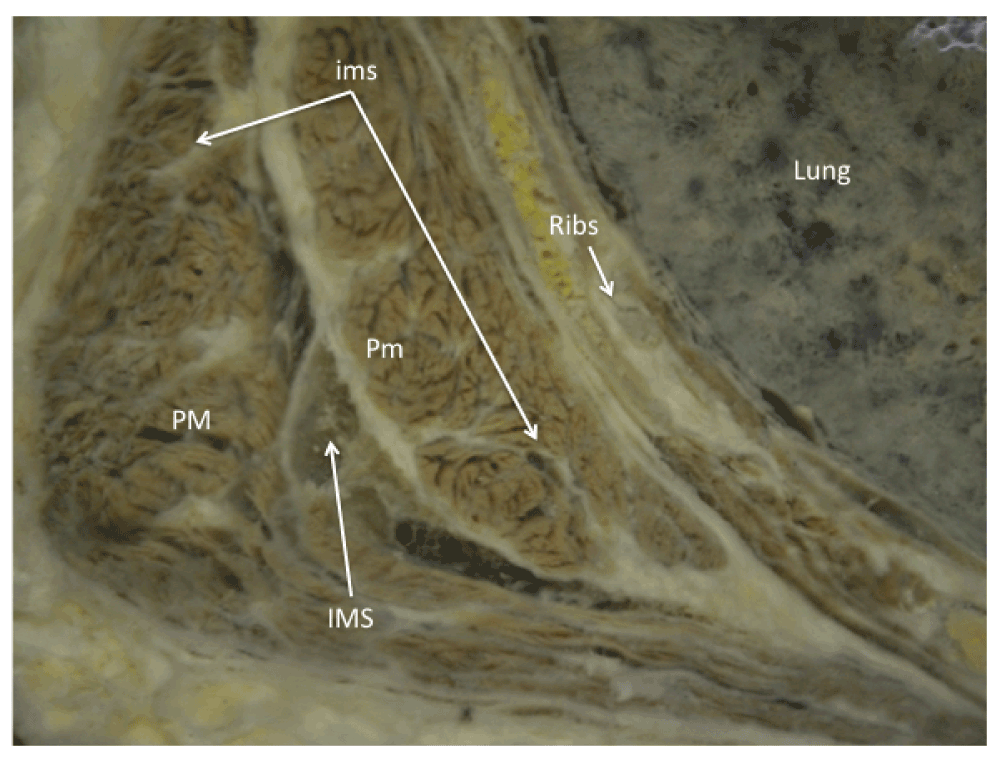
Pectoralis Major PM; pectoralis minor pm; intramuscular septa ims; intermuscular septa IMS. Department of Anatomy at the University of Salvador, School of Medicine, Buenos Aires, Argentina.
Voluntary muscles of the head: from the neural crest cells derive connective tissue elements that guide the patterns of muscle formation in the head; they derive from paraxial mesoderm (somitomeres and somites) and include the muscles of the tongue, the eye (some of them) and are associated with the pharyngeal arches (visceral) (Langman´s Medical Embryology, 2001).
Voluntary muscles of extremities: the mesenchyme is derived from dorsolateral cells of the somites, which move from each limb bud to form the muscles; this occurs near the seventh week of development and can be seen as a condensation (Chevallier et al., 1977; Cristo et al., 1977; Duprez, 2002; Zhi et al., 1996). From somatic mesoderm derives the connective tissue that makes the muscle pattern formation of the limb bones (Langman´s Medical Embryology, 2001; Sadler, 2001).
Spinal nerves play an important role in the differentiation and motor innervation of the limb musculature, and they, as well, provide sensory innervation to dermatomes (Langman´s Medical Embryology, 2001).
Sobolevskii explains in his article that "Comparative anatomy of the spinal cord of the terrestrial, aquatic and semi-aquatic mammals of animals" to the nerve fascicle takes its origin from the spinal cord and elongation of nerve bands, going from the spinal cord to spinal ganglia.
Classically, more attention is given to the connections between the muscles and the skeleton via the tendons of origin and insertion. The recruitment of muscle fibers generates mechanical tension that, via these tendon connections, produces either musculoskeletal movement or maintenance of a static position, thus stabilizing the body (Turrina et al., 2012). Different authors (Hujing, 2003; Huijing & Baan, 2001a; Huijing & Baan, 2001b; Stecco, 2009a; Stecco, 2009b) describe myotendinous expansions that extend from the deep fascia, or extramuscular connective tissue, of muscles to attach to the deep fascia in adjacent segments, periarticular soft tissues, intermuscular septa, interosseous membranes and neurovascular sheaths (Platzer, 1978; Standring et al., 2005; Tidball & col., 1991). Due to these connections, muscles acquire additional surfaces for leverage and the generation of movement (Huijing & Jaspers, 2005; Stecco, et al., 2010a; Stecco, et al., 2010b; Yucesoy et al., 2010). Via these myotendinous expansions, which are a type of extension of the tendons, muscles can directly stretch deep fascia in a number of body segments in a longitudinal sense (Huijing, 2009; Stecco et al., 2007a). They can also stretch it transversally through intramuscular connective tissue (endomysium, perimysium and epimysium) (Huijing & Jaspers, 2005; Monti et al., 1999; Purslow, 2010), as well as through other parts of the musculoskeletal connective tissue (such as the intermuscular septum and neurovascular bundles).
It is also well established that, during a muscular contraction, not all motor units within recruited muscles are activated simultaneously (Finni et al., 2003; Pappas et al., 2002). The presence of intramuscular connective tissue plays a fundamental role in the synchronization of the many variables involved in force production. (Rowe, 1981), as does the retinacula of deep fascia (Sanchis-Alfonso & Rosello-Sastre, 2000).
Intramuscular connective tissue, from the macroscopic to the microscopic aspect, can be defined as:
Epimysium: thicker than the other elements of the intramuscular tissue, it is formed by large diameter collagen fibers (Sakamoto, 1996). In the extreme of the muscle the epimysium thickens before it merges with the tendons of origin and insertion (Benjamin, 2009) and converges in the paratenon, while the muscle belly, which it covers, forms a thin layer that determines the volume of the muscle. Wherever epimysium of two muscles connects, it forms a connective tissue sheath that envelops vessels and nerves destined to reach these muscles. Between the collagen fibers, there is the ground substance, which is rich in hyaluronic acid and other glycosaminoglycans (McCombe et al., 2001). In particular, the hyaluronic acid component allows collagen fibers to slide freely, allowing relative mobility of intrafascial layers (Stecco et al., 2011).
Perimysium: surrounding the muscle and the tendon of origin and insertion of the same muscle, this perimysium does not present a solution of continuity with the epimysium. The amount of perimysium within each muscle and each region of the body varies. The perimysium divides the muscle belly into fascicles of different sizes. Histologically, perimysium has a smaller percentage of elastic fibers and especially collagen fibers type I, III, IV, V, VI and XII, immersed in a matrix of proteoglycans (Kurose et al., 2006; Petibois et al., 2006). The collagen type I fibers provide the perimysium with great resistance to traction. It is probable that, in the transmission of force generated by the muscle towards bone levers, intramuscular connective tissue play an important role (Turrina et al., 2012). In addition, in between two adjacent muscle bundles is placed the same perimysium that contacts with both fascicles by its opposite surface.
Endomysium: is the thinner portion of intramuscular connective tissue. It is directly in contact with the sarcolemma and therefore with every single muscle fiber. Covering the muscular fiber in its entire surface, it takes on a structural role, similar to the parenchyma within organs (Moore, 1983). Endomysium creates a network that unites adjacent fibers. It is the only intramuscular element that, within each muscular fascicle, has contact with the individual muscular fibers innervated by the same motor unit. Other muscular fibers may be interposed but they may remain inactive during the activation of a given motor unit. Thanks to the endomysium, the muscle fibers that are not recruited represent a ‘true tendon’ for the transmission of force laterally, without having to change length (Trotter, 1990). To analyze the properties and structure of a muscle must take into account the arrangement of fibers in relation to the aponeurosis and tendons. (Finni et al., 2003; Turrina et al., 2012) Figure 4.

Department of Anatomy at the University of Salvador, School of Medicine, Buenos Aires, Argentina.
It is evident that right from embryological development, the fascia determines the structural and functional characteristics of the muscle.
During evolution: the independence of the various segments meant that the myomeres and myosepta were required to align themselves according to new lines of force, thus abandoning metamerism altogether. Previously the entire musculature of a body intervened every time it was moved in any direction, or plane, but at this stage each part of the body required its own musculature (Chiarugi, 1975; Stecco, 2004). In this way, the method of Fascial Manipulation organized into six myofascial units, formed by mono and biarticular fibres as well as muscle spindles, were created for each segment.
The evolutionary process proceeded in the following manner:
- at first metameres lengthened according to the lines of tension;
- then the myosepta, or metameric septa, in part joined with the unidirectional muscle fibres to form the muscle spindles and, in part, surrounded the entire muscular mass to form the epimysium (Chiarugi, 1975; Stecco, 2004).
From the observation of this process in bony fishes we find that lateral flexion of the trunk stimulated the myomeres to elongate in a cephalo-caudal sense which in turn induced the myosepta-fasciae to align parallel to the traction. Hence, these fibres lengthened and they connected to a number of segments (Figure 5).

Department of Anatomy at the University of Salvador, School of Medicine, Buenos Aires, Argentina.
As a consequence, the myosepta-fasciae, which are no longer metameric, lengthened between the muscle fibres to form the perimysium. The deep muscles maintained parallel fibres between one vertebra and the next, somewhat similar to the first metameric stage. The more superficial muscles however formed links extend over a number of metameres. The multisegmentary muscular fibres have the muscle spindles, which are placed in parallel to them, as their point of reference. The muscle spindle is the connection between the unidirectional muscular fibres and the relative portion of the primordial fascia (Stecco, 2004). Only the presence of the muscle spindles, plus the activation of the muscular fibres in succession, allows for the arrest of movement at any angle of the entire joint range. Due to the muscle spindles and the Golgi tendon organs, the unidirectional fibres intervene in succession during the movement of a body part in one plane (Stecco, 2004). The evolution of peripheral receptors clearly demonstrates how the brain depends upon the fascia in order to be able to formulate the concept of space and time. To enable it to organise movement, in general the brain requires feedback concerning whatever is taking place in the periphery. Normally the free nerve endings are involved in deep somaesthetic activity or the perception of the body´s position and movement in space. This feedback transmits information to the CNS regarding the exact position of any given body segment. The fascia and, in particular, its directional sequences, have a predefined length and therefore can act as a type of measuring device in the periphery. Furthermore, because of its elasticity, it is also capable of applying stretch to neuroreceptors (Kent, 1997); (Stecco, 2004). According to the angle of the articulation and therefore, the fascial stretch, either a sequence, a diagonal or a spiral is activated with each step or movement. In other words the mind initiates a motor scheme and the fascia assists in the process of its realisation (Stecco, 2004). The functional meaning of this relationship between the activity of the muscle, the mobility of intra and extramuscular connective tissue and, function of intermuscular septa to achieve the formation of sequences, diagonals, and the spirals of movement, should be studied by also taking into account the presence of numerous embedded receptors that may play a role in peripheral coordination of movement. (Stecco, 2004; Stecco, 2007a; Stiwell, 1957; Turrina et al., 2012; Yahia et al., 1992).
In the 1940s, when Doctor Herman Kabat began to develop the concept of Proprioceptive Neuromuscular Facilitation (PNF) (Voss et al., 2001), it was sustained that normal neuromuscular mechanisms were capable of developing a wide range of motor activities within the limits of the anatomical structure, the level of development, and innate and previously learnt neuromuscular responses. In other words, the countless combinations of movement that an individual employs to satisfy everyday needs are acquired by means of a well-established process of development and an abundance of learning situations. Neuromuscular mechanisms are assimilated and become efficient without paying attention to individual muscular activity, reflex activity, or the enormous amount of neurophysiologic reactions involved. In the introduction to the PNF method (Voss et al., 2001) it is stated that ‘movement patterns are mass movement patterns. Mass movement is a characteristic of normal motor activity and it is in consonance with Beevor’s Axiom: the brain knows nothing of individual muscle action, but knows only of movement. In normal motor activity, the varied combinations of movement, or mass movement, demand shortening and lengthening reactions from many muscles and in different degrees. Mass movement, which has the purpose of setting out a specific demand, has to consist of a special combination of movements, ideal to the specific sequence of muscles responsible for that movement and has to allow those muscles to contribute with their action components in a harmonic way’.
More recently, (Myers, 2009) has suggested that perhaps there are not in fact 600 muscles but a single connective tissues system with "pockets, which gives muscles and muscles fibers to settle their spaces in different directions to perform its function.
In neurophysiology, we find that movement is programmed by the mind and carried out by the contraction of the muscle fibers. Nerve impulses generated from the brain determine the displacement of a certain segment and it is subject to a vast number of variables in the periphery (Stecco & Stecco, 2010).
Given that each muscle has fibers with different functions and innervations (Burke et al., 1974), according to the Fascial Manipulation® concept (Stecco, 2004), the musculoskeletal system could be divided into myofascial units (MFUs). Each MFU is said to be composed of motor units responsible for moving a segment in a specific direction and, along with the fascia surrounding these fibers, these forces generate movement vectors. A MFU necessarily includes the nerve components innervating these elements. Furthermore, the main vectors of each MFU generate other smaller, more distant vectors (Stecco, 2013).
The fascial context in which muscle fibers are inserted could potentially coordinate movement in the periphery. The MFU is the element that could synchronize the action of motor units innervated by the same axon. While the brain sends impulses to muscle fibers scattered within the muscle via the axon of a single motor neuron, diminishing the activity of the more distal fibers and increasing it in the most proximal ones in relation to changing positions of the limb or the required force presents neurological challenges. An elastic structure, sensitive to stretch, such as the fascia, could assist modulation of recruitment or inhibition via the activation of mechanoreceptors embedded in this tissue (Stecco & Stecco, 2004) The potential role of these mechanoreceptors in the coordination and synchronization of movement via their proprioceptive function is emphasized. As connective tissue continues in the tendons and the muscle insertions (Hujing, 2003; Huijing & Baan, 2001a; Huijing & Baan, 2001b; Stecco, 2009a; Stecco, 2009b), it also distributes and directs the muscular force to the bone in an appropriate way (Stecco & Stecco, 2010).
1) If the brain is not aware of individual muscle activity, but knows and plans movement in terms of patterns (Beever’s axiom), should we continue thinking in terms of individual muscles bringing about movement?
2) Is there a central command that sends an order through nerve impulses, all or nothing, and are there peripheral systems that regulate coordination and synchronization according to the requirements of movement patterns? Is the brain really in charge of everything?
3) If the brain coordinates, regulates and synchronizes everything, what is the purpose of the peripheral nervous system, the fascia and its embedded mechanoreceptors?
From this review, a new comprehension of the connective tissue in general, and more specifically, of the musculoskeletal system emerges:
- The connective tissue drives musculoskeletal development from conception.
- In the 1940s, prompted by his rehabilitative work with patients affected by poliomyelitis, Kabat initiated discussion concerning how the brain interprets movement in terms of finalized gestures or patterns of movement.
- In the 70s, Stecco developed a comprehensive method (Fascial Manipulation®) based on similar reasoning, introducing the potential role of muscular fascia in musculoskeletal activity.
- More recent studies concerning the anatomy and physiology of intramuscular and extramuscular connective tissue and its innervation suggest that the myo–fascial-skeletal system presents peripheral response mechanisms that are capable of responding to movement patterns.
- These mechanisms include functions such as musculoskeletal coordination, synchronization, regulation of movement, and the transmission of contractile force from the muscle fibers and fascicles within muscles and as well as along body segments via myotendinous expansions and intramuscular septa. Lastly, the neurovascular tracts, wrapped in fascia, also transmit contractile force (Karassolik et al., 2009; Lindsay, 2008). As suggested by Langevin (2006), the fascial system represents a ‘body-wide proprioceptive/communicating organ’.
Connective tissue guides our developmental phases during intrauterine life and continues to evolve through movement patterns. In addition, the brain apparently knows very little about individual muscle action, but recognizes movement patterns catalogued by learning. Muscle fibers and fascicles are oriented in a direction dictated by their connective tissue in a pre-established pattern, reinforced through movement patterns and modified to adjust to the individual, making it effectively redundant to name every muscle involved in the process. With ongoing research concerning the anatomy and physiology of the myo-fascial-skeletal system is it not time for a paradigm shift with regards to this system?
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)