Keywords
ciliogenesis, centriole assembly, Deuterosome, deuterosome-dependent,
ciliogenesis, centriole assembly, Deuterosome, deuterosome-dependent,
The centriole is a cylinder-shaped organelle that serves as the core of the centrosome or the basal body of the cilium1–5. Nascent centriole formation usually depends on pre-existing mother centrioles. Normally in one cell cycle each mother centriole produces only one daughter centriole, that is directly adjacent (Figure 1). Such tight control ensures proper mitosis, since only two centrosomes are required to function as the spindle poles. It also guarantees that the centriole number remains constant after cell division (Figure 1).
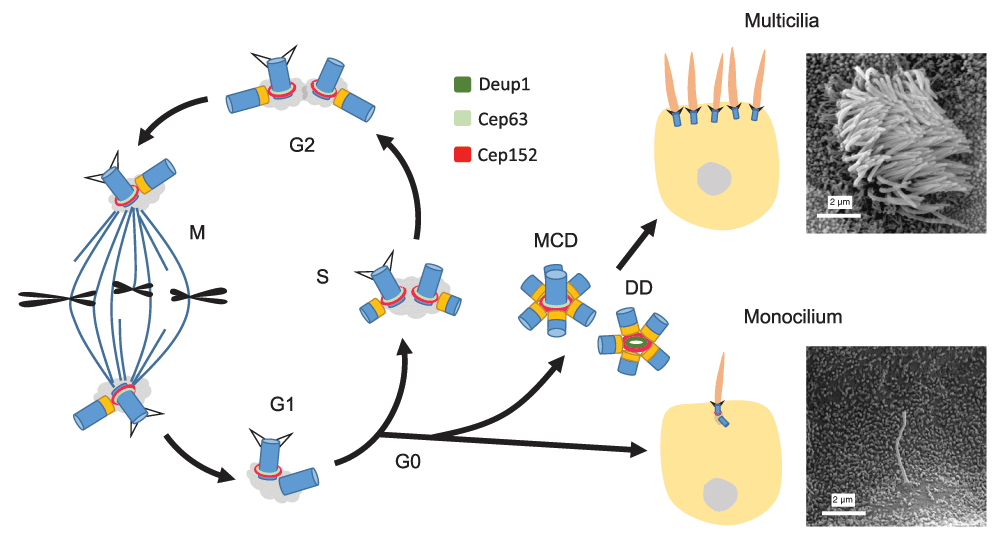
The centrosome in a G1 cell contains a pair of mother-daughter centrioles. Upon entering the S phase, each centriole starts to duplicate one daughter centriole so that the centriole number remains constant after mitosis (a). When the cell enters G0, the mother centriole can be transformed into the basal body to support monocilium formation (b). Alternatively, both the mother centriole-dependent (MCD) and deuterosome-dependent (DD) pathways can be activated to generate an abundance of centrioles for dense multicilia formation (c). The scanning electron microscopy images show a primary cilium in the collecting duct of mouse kidney and multicilia of a multiciliated cell in mouse tracheal epithelium, respectively. Centrioles are drawn in blue and their cartwheels in orange.
Ciliogenesis occurs at the G0 or G1 stage of the cell cycle (Figure 1)1,3,6. In vertebrates, most cells can possess a primary cilium, which functions as a sensory organ for diverse environmental signals. Mammalian epithelial tissues such as those lining the inner surface of the trachea, the oviduct, and the brain ventricles, however, have abundant multiciliated cells (MCCs) with hundreds of cilia (Figure 1). These multicilia are motile and their beating is critical for mucus clearance, ovum transport, or cerebrospinal fluid circulation7. How then do such cells generate sufficient numbers of basal bodies?
The mystery was initially uncovered by electron microscopy (EM) on a variety of MCC-containing tissues in the 1960’s and 1970’s. The mother centriole was observed to be surrounded by multiple daughter centrioles in MCCs. Moreover, many granular or ring-shaped EM structures termed deuterosomes (this name will be used in this review), procentriole precursor bodies, dense granules, and generative complexes were also able to initiate procentriole assembly8–12. Importantly, the deuterosomes were estimated to produce most of the basal bodies required. Nevertheless, it is only recently that we have begun to understand the molecular mechanisms involved, which will be the major focus of this review.
Tremendous progress has been made toward understanding how a daughter centriole is born in cycling cells. A group of proteins, including Cep152 and Cep63, are specifically located around the proximal side of the mother centriole. In the G1 phase, the polo-like kinase PLK4 binds to Cep152 to form the site of centriole assembly13–17. In the S phase, a cartwheel structure is formed at the PLK4 site, followed by the assembly of the nine sets of microtubule triplets and other components of the daughter centriole. Centriole assembly is completed by the G2 phase and, following mitosis, each daughter cell inherits a mother-daughter pair of centrioles (Figure 1)1–5.
Interestingly, mother centrioles in cycling cells are capable of generating more than one daughter centriole. For instance, overexpression of PLK4 results in multiple PLK4 foci around the mother centriole and overproduction of daughter centrioles18,19. Overexpression of Cep152 or the cartwheel proteins SAS-6 or STIL also has a similar effect20–23. These observations not only indicate that cycling cells execute the one-daughter-centriole-per-mother rule by restricting the levels of several critical proteins but also suggest that MCCs may break this rule by simply upregulating the protein levels. Indeed, when mouse tracheal epithelial cells (MTECs) are induced to form multicilia, they express high levels of these proteins19,24,25. The importance of PLK4 and Cep152 in mother centriole-dependent (MCD) centriole overduplication of MTECs is also verified19.
The discovery of an essential deuterosome component, Deup1 (also called Ccdc67), has promoted the understanding of deuterosome-dependent (DD) centriole biogenesis. Strikingly, Deup1 is a paralog of Cep6319. Cep152 binds to both Cep63 and Deup1 to stabilize them and be recruited, respectively, to the mother centriole and the deuterosome19,26. Therefore, if we consider the Cep63-Cep152-containing proximal ring of the mother centriole as a platform, or ‘cradle’, that supports nascent centriole assembly, deuterosomes are analogous cradles, independent of mother centrioles (Figure 2A). In MTECs, deuterosomes appear initially as foci with zero to two associated procentrioles (Figure 2A-B, stage II). Their sizes then enlarge, accompanied by an increase in procentriole numbers (Figure 2A-B, stage III). They are disassembled upon completion of centriole assembly (Figure 2A)19,27. Usually 50–100 deuterosomes can be found in a MTEC, sufficient for the production of hundreds of centrioles (Figure 2B)19. Mouse ependymal cells (MEPCs) displayed a similar centriole amplification process, but their deuterosomes are usually much larger in size and smaller in number (Figure 2C).
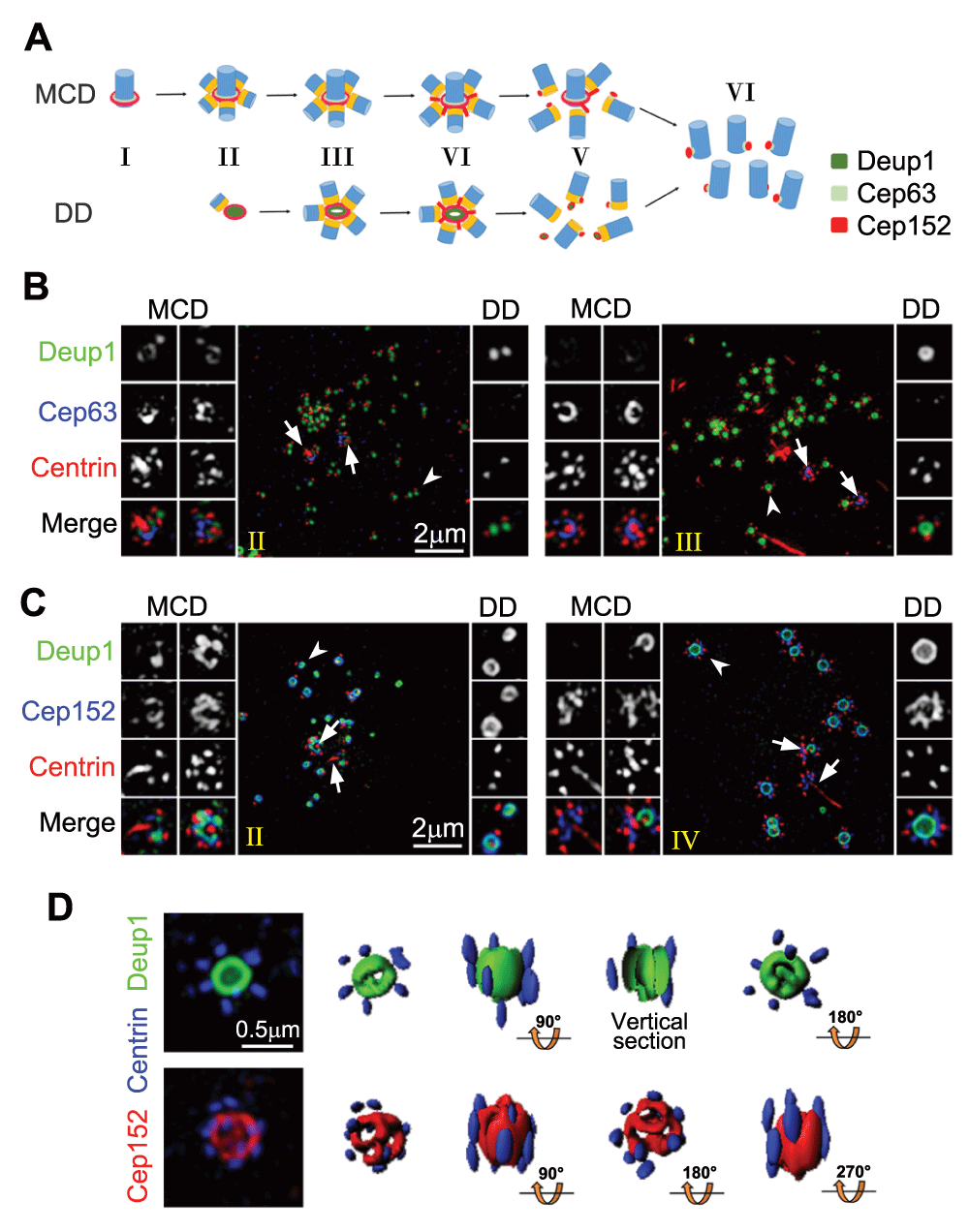
(A) Illustration for centriole amplification stages in MTECs19. Centrioles are drawn in blue and their cartwheels in orange. (B) Three-dimensional structured illumination microscopy (3D-SIM) images for MTECs at early stages (II and III) of centriole amplification. MTECs cultured as described previously19 were immunostained for Deup1, Cep63, and Centrin and imaged using a DeltaVision OMX V3 microscopic system (GE Healthcare). The mother centrioles (arrows) and representative deuterosomes (arrowheads) are magnified 2× to show details. (C) 3D-SIM images showing centriole amplification in MEPCs. MEPCs were isolated from neonatal C57BL/6J mice and cultured as described32. The cells were fixed at day three after serum starvation and immunostained for Deup1, Cep152, and Centrin. The stages (II and IV) are defined as in the MTECs. Note that MEPC deuterosomes (C) are usually much larger than those in MTECs (B). (D) SIM images of two large MEPC deuterosomes immunostained for Deup1 and Centrin (top row) or Cep152 and Centrin (bottom row). Their 3D profiles are also shown. Abbreviations: DD, deuterosome dependent; MCD, mother centriole dependent.
The beauty of such a DD pathway is obvious: cycling cells only need to turn off the DD pathway by shutting down Deup1 expression to avoid the production of extra centrioles. On the other hand, as MCCs are terminally differentiated and no longer able to enter the cell cycle, turning on the DD pathway and upregulating other genes critical for basal body assembly can safely fulfill their demand on large numbers of basal bodies. For instance, the Multicilin-E2F4/5 complex is known to activate the transcription of Deup1, Plk4, Cep152, and many other centriolar protein genes in MCCs28–30. Other proteins such as cyclin O appear to fine-tune the transcription program31.
Deuterosome size varies remarkably in different tissues and species: for instance, from 100–200 nm (diameter) in rat or mouse MTECs8,19 to more than 500 nm in the mouse oviduct10. Larger deuterosomes are capable of supporting more procentrioles. As deuterosomes look mostly ring shaped in transmission EM, they were proposed to be roughly sphere shaped, capable of assembling centrioles in all directions8,10. Serial ultra-thin sections of MEPCs support this notion32.
Three-dimensional profiling of subdiffraction images from both MTECs and MEPCs, however, suggests that Deup1 and Cep152 are arranged in a ring-shaped configuration in the deuterosome, with the Cep152 signals enwrapping those of Deup1 from outside (Figure 2C)19. Such a configuration is topologically analogous to the mother centriole cradle. Only the ends of the deuterosome appear relatively amorphous. For instance, in large deuterosomes such as those of MEPCs, the Cep152 signals may exhibit several ‘holes’ at each end (Figure 2D). Procentrioles tend to be assembled on the outer wall of the deuterosome but can be found at both ends as well (Figure 2D)19.
Whether there are additional proteins to construct the outer wall, fill the center, or cap the ends of the deuterosome is presently unknown. Ccdc78, a coiled coil domain-containing protein, is reported as a deuterosome-specific protein required for centriole amplification in the Xenopus embryonic epidermis33. Nonetheless, mouse Ccdc78, expressed either endogenously or exogenously, was not detected on Deup1-positive deuterosomes in our hands, raising the possibility that Ccdc78 may be either an amphibian-specific deuterosome component or even not a bona fide one.
How deuterosome components are packed together to form the supramolecular structure is also an important issue. Fibrous granules (also called fibrogranular material or proliferative elements), clouds of material abundant in 40 to 80 nm granules that coincide with deuterosome formation in MCCs, were proposed to be precursors of the deuterosome8–10. PCM-1, a component of fibrous granules, however, failed to show deuterosome localization34. Its depletion by RNA interference also didn’t impair centriole amplification25. Likewise, neither Deup1 nor Cep152 exhibited obvious fibrous granule-like distributions (Figure 2B-C)19. Since small deuterosomes tend to emerge in bulk and then grow in synchrony and ectopic expression of Deup1 in cycling cells is sufficient to induce the formation of functional deuterosomes (Figure 2B)19, we propose that deuterosomes can be assembled spontaneously (Figure 2A).
Interestingly, a recent publication argues for a totally different mechanism32. Based mainly on studies in MEPCs, a model is proposed in which an unknown mechanism recruits Deup1, Ccdc78, and other cradle proteins to a site in the cradle of the young mother centriole to initiate the assembly of both the deuterosome and the daughter centrioles. The deuterosome-procentriole halo is then released so that the site can begin the next assembly cycle. After the release of the last halo, procentrioles on all the deuterosomes start to elongate and mature. Thus, both the deuterosome formation and the massive centriole biogenesis are MCD processes. Deuterosomes function merely as shuttles to carry the daughter centrioles away from their mother centriole into the cytoplasm32,35.
This model, despite its uniqueness, still needs further verification. Firstly, it remains to be shown whether this is the sole and universal way of deuterosome generation. Deup1 is capable of mother centriole localization (Figure 2B-C)19. It is thus understandable that some of the protein there may serve as seeds to initiate deuterosome assembly. Since live imaging in the MEPCs suggests that the generation of one halo requires about two hours32, such an efficiency would demand several days to generate the 50–100 deuterosomes in MTECs, while the entire centriole amplification process takes roughly only one day19,32. Thus, both the spontaneous and MCD pathways may contribute. Furthermore, there might be multiple deuterosome nucleation sites on both the young and the old mother centrioles (Figure 2B-C). Secondly, the model is apparently incompatible with the observation that the numbers of deuterosome-associated procentrioles increase over time (Figure 2B-C)19. Even if one or two daughter centrioles could be carried away from the mother centriole by each nascent deuterosome, their subsequent increase in numbers still argues for the existence of de novo DD centriole biogenesis. Finally, what defines the deuterosome nucleation site on the mother centriole and how the cytoplasmic halos can wait until the last one is released are also issues for future clarification.
Phylogenetic analysis suggests that Deup1 is originated from a common fish ancestor of the lobe-finned fish and tetrapods in the vertebrate evolution to boost cilia density in MCCs19,36. Accordingly, in contrast to the lobe-finned fish (such as lungfish), MCCs of the ray-finned fish (such as zebrafish), which have no Deup1, contain only sparse cilia37,38. Many invertebrates, however, possess MCCs with dense multicilia39–42. Deuterosome-like ultrastructures have also been reported in some invertebrate species43,44. A comprehensive knowledge of strategies for centriole amplification throughout metazoan evolution will thus require an understanding of the mechanisms for multiciliogenesis in the invertebrate.
The mechanism of centriole amplification is both exciting and challenging. Because the sizes of centrioles and deuterosomes are below or close to the optical diffraction limit, technical limitations of imaging are a current major bottleneck restraining studies of centriole amplification in MCCs. Although 3D structured illumination microscopy (SIM)45 has proven its power in the past19,32, the development and introduction of super-resolution techniques with higher spatial (especially the z-axis) and temporal resolutions46–49 are expected to greatly facilitate studies in the field. Furthermore, other cutting-edge techniques such as cryo-electron tomography, omics analysis, and computational biology may help to solve issues on the structure, formation, growth, disassembly, and function of the deuterosome as well as the entire mechanism that controls appropriate on-and-off switching of the centriole amplification program.
DD, deuterosome dependent; EM, electron microscopy; MCC, multiciliated cell; MCD, mother centriole dependent; MEPC, mouse ependymal cell; MTEC, mouse tracheal epithelial cell; SIM, structured illumination microscopy.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)