1Cologne Biocenter, Botanical Institute II and Cluster of Excellence on Plant Sciences (CEPLAS), University of Cologne, Cologne, Germany 2Department of Biology and Cluster of Excellence on Plant Sciences (CEPLAS), Heinrich-Heine University, Düsseldorf, Germany 3Plant Molecular Biology, Department of Biology I, Ludwig-Maximilians-University, Munich, Germany
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
Photosynthesis is central to all life on earth, providing not only oxygen but also organic compounds that are synthesized from atmospheric CO2 and water using light energy as the driving force. The still-increasing world population poses a serious challenge to further enhance biomass production of crop plants. Crop yield is determined by various parameters, inter alia by the light energy conversion efficiency of the photosynthetic machinery. Photosynthesis can be looked at from different perspectives: (i) light reactions and carbon assimilation, (ii) leaves and canopy structure, and (ii) source-sink relationships. In this review, we discuss opportunities and prospects to increase photosynthetic performance at the different layers, taking into account the recent progress made in the respective fields.
Corresponding author:
Ulf-Ingo Flügge
Competing interests:
The authors declare that they have no competing interests.
Grant information:
Work in the authors´ laboratories cited in this review was supported by the German Research Foundation (DFG), the Federal Ministry of Education and Research (BMBF), and the European Union (project 3to4).
Photosynthesis is a process that all life on earth depends on. Photosynthetic organisms convert more than 109 metric tons of atmospheric CO2 into biomass per year. With the global human population rising from ~7 billion now to 9–10 billion by 2050, the worldwide trend towards a more meat-rich human diet, the loss of harvest and grazing land, and the negative effects of global warming on crop production have put forward the question of whether this incredibly high amount of biomass production can be further increased.
Crop yield is determined by the available solar irradiation energy across the growing season (0.487 St), the genetically encoded properties of how the radiation is intercepted (εi), the efficiency by which the light energy is converted into biomass (εc), and what fraction of the total biomass is partitioned into the harvestable part of the plant (harvest index, εp). This results in the Monteith equation: yield = 0.487 • St • εi• εc• εp1–3.
The Green Revolution raised the yield potential of the major grain crops mainly by increasing the harvest index, which is now about 0.6. Breeders were also able to improve the light interception efficiency, which in modern cultivars is up to about 0.8–0.9. All available evidence suggests that additional grain yields by further increasing the harvest index or optimizing light interception are rather unlikely; they appear to be close to their biological limits already. In contrast, the best light conversion efficiency (εc) observed in field experiments (0.24 in C3- and 0.37 in C4-crops) is far below the theoretical maxima (0.46 in C3- and 0.6 in C4-plants) and thus not yet close to its biological limit. Enhancing, and in the long term re-designing, photosynthesis with respect to light energy conversion efficiency is therefore a prime target when aiming to increase crop yield3–6.
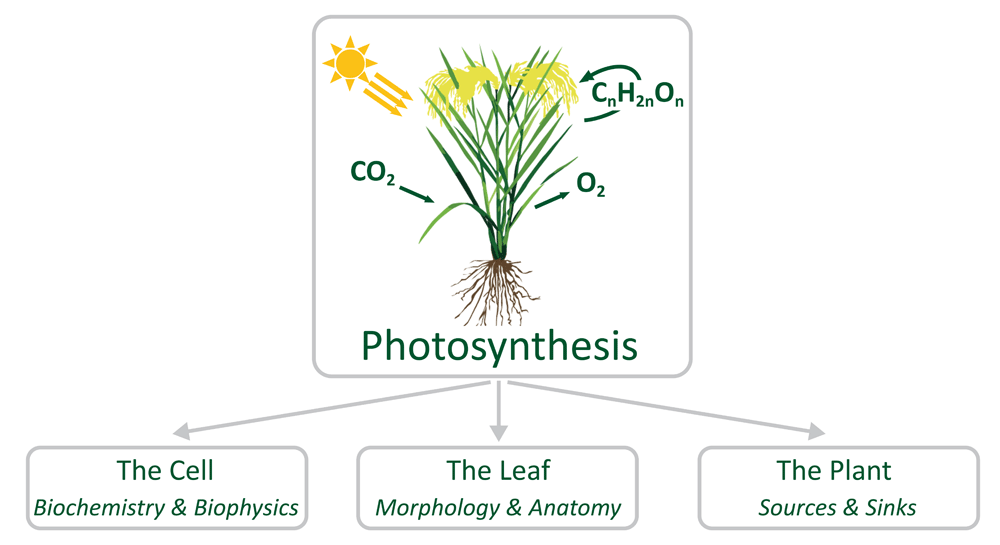
In general, photosynthesis can be described as a cellular trait that uses light energy to convert atmospheric CO2 into carbohydrates. At a higher level, photosynthesis is determined by the activity of leaves and by the canopy structure. And, finally, photosynthesis is related to the capacities of source and sink tissues, i.e. mature leaves and heterotrophic organs, respectively (Figure 1).
Figure 1. The process of photosynthesis viewed from different perspectives.
Here, we will discuss the process of photosynthesis from these various perspectives and address putative targets for the improvement of photosynthetic performance.
Photosynthesis as a cellular trait: (1) light-dependent reactions
Novel insights into the dynamic interactions of photosynthetic complexes
The light-dependent reactions involve five major multi-protein complexes: photosystem I (PSI), photosystem II (PSII), cytochrome b6f, ATP synthase, and NADPH dehydrogenase (NDH). In particular, PSI can (transiently) form larger super-complexes with other complexes like cytochrome b6f7, NDH8, and PROTON GRADIENT 5 (PGR5)-PGR5-LIKE 1 (PGRL1)9,10. More recently, interactions of PSI with PSII11 and light-harvesting complex II (LHCII)12 have been characterized. Traditionally, PSI and PSII are thought to be spatially dispersed. However, in cyanobacteria, a megacomplex consisting of PSII, PSI, and phycobilisomes was found13. Aro and co-workers reported that PSI and PSII complexes coexist with LHCs in Arabidopsis14. More recently, Tanaka and colleagues suggested that about half of PSIIs are physically connected to PSI complexes11. The obtained results suggest that when PSII becomes excessively excited, it can divert excitation energy directly to PSI to avoid photo-damage. Conversely, photoinhibition of PSI can downregulate PSII via a mechanism that involves increased reduction of the intersystem electron carrier system due to damage to the FeS clusters of PSI. This is associated with the activation of thylakoid phosphorylation-based mechanisms that increase energy flow towards PSI15.
Bassi and colleagues have recently generated plants with a PSI that lacks its natural LHCI antenna system12. Despite the absence of LHCI, LHCII can still attach to PSI, and in fact each LHCI-less PSI complex binds one LHCII trimer, thus fully replacing the four Lhca proteins from LHCI by the three Lhcb proteins of LHCII. This demonstrates that LHCI is not necessary for excitation energy transfer from LHCII to PSI, as was previously suggested16. Intriguingly, the transfer of energy from LHCII to PSI appears to be more efficient than from LHCI, raising the question of why PSI uses LHCI instead of LHCII. A plausible explanation is that LHCI absorbs photons with lower energy than LHCII, avoiding competition between PSI and PSII for the same photons. Therefore, in low-intensity and far-red enriched light (like the one available under canopy cover), PSI is favored over PSII, avoiding over-reduction of plastoquinone and thus preventing photo-damage.
With respect to photosynthetic improvements, the structural flexibility of the photosynthetic complexes might allow the construction of novel combinations of photosystem cores and LHC antenna complexes. Indeed, it appears to be possible to design an entire set of such combinations that could be tested under different light regimes for their efficiency and use under controlled (such as in greenhouses) and natural (such as in the field) light conditions. In a complementary approach, conditional regulation of various light-harvesting genes might allow for tuned optimization in an ever-changing and challenging environment. Therefore, biotic interactions also need to be considered, taking into account, for instance, trade-offs between photosynthesis and herbivore resistance17.
Novel auxiliary components of photosynthetic light reactions
During the last few decades, the structural components of the light reactions, including the multi-protein complexes PSI and PSII, cytochrome b6f, ATP synthase, and NDH, have been identified and extensively characterized at the gene, protein, and mutant level. In contrast, the inventory of the auxiliary components of the photosynthetic light reactions is far from being complete. Novel proteins involved in the biogenesis, repair, regulation, and protection of the photosynthetic machinery are continuously identified owing to genetic screens and functional genomics approaches and employing mainly the model species Arabidopsis thaliana, Chlamydomonas reinhardtii, and Synechocystis sp. PCC 6803. In Table 1, recently identified auxiliary proteins of the photosynthetic light reactions are listed. Adding their numbers to the ones of the factors already identified before (reviewed in 18), it becomes clear that the number of different proteins controlling the biogenesis and repair of the multi-protein complexes exceeds the one of structural subunits. While for PSI and NDH, and particularly PSII, substantial sets of assembly factors have been known for many years already, until recently only one auxiliary protein (ALB419) involved in the biogenesis of the chloroplast ATP synthase (cpATPase) has been isolated. Three additional cpATPase assembly factors have now been identified: CGL16020,21, PAB22, and CGLD11/BFA323,24. CGL160 represents a protein of prokaryotic origin, whereas CGLD11/BFA3 and PAB are specific to photosynthetic eukaryotes. Interestingly, CGLD11/BFA3 is also targeted to mitochondria but not (yet) essential for the assembly of the mitochondrial ATP synthase23. Similar pronounced evolutionary dynamics are displayed by PAM68L, which evolved from a PSII assembly factor to mediate the assembly of the chloroplast NDH complex in Arabidopsis25.
Table 1. Overview of novel auxiliary components of photosynthetic light reactions identified in the last few years.
The network of protein kinases and phosphatases involved in reversible thylakoid phosphorylation (reviewed in 26) and the two ferredoxin-plastoquinone reductase complexes involved in cyclic electron flow (reviewed in 27) are now relatively well characterized, and also several factors regulating the dynamics of thylakoid ultrastructure and the acclimation and protection of the photosynthetic machinery have been identified (reviewed in 28–30). Three recent examples are highlighted here: (i) the levels and phosphorylation states of CURT1 proteins were shown to control the formation of the appressed regions of thylakoids in the so-called grana stacks31; it remains to be elucidated how the activity of CURT1 is regulated under physiological conditions. (ii) The calcium sensor CAS was localized to thylakoids32, appears to regulate acclimation and cyclic electron flow in C. reinhardtii33,34, and was more recently associated with retrograde signaling in A. thaliana35; here, future research needs to clarify the role of chloroplasts in cellular calcium signaling and how CAS is involved in this process. (iii) Very recently, the chloroplast-splicing factor HPE1 was shown to be involved in the regulation of photosynthetic efficiency; in fact, plants without HPE1 re-adjust their light-harvesting pigments, thereby reducing antenna size, improving light capture, decreasing energy loss, mitigating photodamage, and enhancing photosynthetic quantum yield during photosynthesis36.
As suggested before, the most promising targets for genetic engineering of the light reactions of photosynthesis––in terms of manipulating one or a few genes––are modifying light harvesting and regulators of photosynthetic electron flow37. The resulting plants have the potential to exhibit more efficient photosynthesis under controlled conditions, e.g. in greenhouses, or in regions that cannot otherwise be extensively used for agriculture because of their short growing seasons. In fact, plants without the LHCII protein phosphatase TAP38 and concomitantly with hyper-phosphorylated thylakoid proteins38 or without HPE136 (see above) appear to be superior to normal plants under certain conditions, implying that the modification of single regulatory proteins might positively impact photosynthesis. Moreover, since evolutionary-based adaptation of plants might not cope with the relatively rapid progression of human-made climate change that we experience today, transgenic plants with altered acclimation capacity due to altered activity of accessory photosynthetic proteins might contribute to the generation of improved crop varieties. Proof of this concept was provided very recently. The parallel overexpression of three proteins involved in photoprotection (subunit S of PSII [PsbS], violaxanthin de-epoxidase [VDE], and zeaxanthin epoxidase [ZEP]) accelerated the photoprotective response to natural shading events in tobacco, resulting in increased plant dry matter productivity in fluctuating light and under field conditions39.
Photosynthesis as a cellular trait: (2) assimilation reactions
During photosynthesis, plants capture energy from sunlight and turn it into biochemical energy in the form of ATP and reducing equivalents, NADPH. In C3-plants, the first product of the CO2-fixing ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), the C3-compound 3-phosphoglycerate, is transformed to a C3-sugar (triose phosphate) as part of the Calvin-Benson cycle using both ATP and NADPH as products of the light reaction. In most plants, triose phosphates are subsequently converted into sucrose as the main photoassimilate exported from photosynthetically active leaves (the source tissue). The long-distance transport of photosynthates to heterotrophic organs (the sink tissue) serves to supply these organs with carbon and energy. In harvestable organs, carbon can be stored as, e.g., sucrose, starch, oil, or, in combination with nitrogen, protein.
In C3-plants, the category to which most of our crop plants belong to, RuBisCO is CO2 limited and works, at best, at half of its maximal reaction velocity40. The oxygenase activity of RuBisCO initiates the wasteful process of photorespiration that finally results in the release of previously photoassimilated CO2 and ammonia41. Multiple efforts have been or are still being undertaken to improve RuBisCO activity either directly42, albeit with very limited success so far6, or by targeting auxiliary proteins, for instance RuBisCO activase43. Alternatively, by introducing CO2 pumping devices into C3-species, the limitations of RuBisCO are expected to be reduced. These CO2 pumps are to operate via either a C4 cycle44 or inorganic carbon-concentrating devices of cyanobacterial origin45. There were also attempts to reduce photorespiratory losses by introducing alternative salvage pathways46,47. In addition, promising approaches have also been initiated to overcome bottlenecks in the Calvin-Benson cycle48,49.
Photosynthesis: leaves as target for photosynthesis improvement
Leaves are the plant’s organs that are dedicated to photosynthesis. Under high-light conditions, e.g. at midday, plants receive excess energy that they are not able to cope with. It was proposed earlier that photosynthetic efficiency could be maximized by improving the plants’ canopy light distribution in a way that minimizes light saturation of the upper leaves and light starvation of the lower leaves6. This could be achieved by (i) varying the angle of the leaves in the canopy, (ii) altering the size of LHCs per photosystem, i.e. fewer LHCs in the upper leaves and larger LHCs in lower leaves, and/or (iii) extending the light absorption spectrum of photosynthetic pigments into the near-infrared region in the lower leaves in order to use this light quality more efficiently6,50,51.
The leaves of the angiosperms vary greatly in size and shape, from single and entire to highly complex compound leaves52. However, the role and importance of leaf anatomy in contributing to the photosynthetic output of leaves is largely unexplored. It was suggested earlier that both leaf shape and leaf anatomy contribute to the photosynthetic output of leaves53 and are therefore promising targets for improving photosynthesis54. Genetic studies with rice support this notion by demonstrating that in rice the anatomy of leaves is closely associated with leaf photosynthesis55,56 and, moreover, with crop yield57,58. Unfortunately, global genetic analyses, preferably with easily accessible genetic model plants, which aim to identify genes affecting the inner anatomy of leaves and concomitantly their photosynthetic output, are still missing.
One approach to tackle this issue would be to compare leaf form and anatomy in established model systems, e.g. A. thaliana and Cardamine hirsuta, which both belong to the Brassicaceae. In contrast to A. thaliana, C. hirsuta has complex leaves subdivided into leaflets. Major steps in understanding the genetic basis for variation in leaf shape both within and between species have already been made59–63. Both species have been documented to be suitable for an easy identification of genes and their functions. In both species, genes could be identified that affect primarily leaf form and/or the structure and organization of palisade and spongy parenchyma tissues of the leaf. A second step would be to investigate how and to which degree these genes can be used to optimize the photosynthetic output of leaves. The forward genetic approaches could rely on either mutagenesis or the available natural genetic diversity. Both photosynthesis and leaf differentiation are rather conserved in evolution, at least among the angiosperms. The transfer of knowledge from these model systems to the major crop species of this plant family, namely the Brassicas, should therefore be relatively straightforward.
Photosynthesis and the interaction with source-sink metabolism
Photosynthesis is part of a superordinate system, namely the whole plant. From the system’s perspective, the production of goods and their utilization have to be coupled, i.e. previous to the storage of products derived from photosynthetic assimilation processes in sink tissues (i.e. in harvestable organs), and carbon and nitrogen have to be assimilated in the source tissue (i.e. in mature leaves). To increase biomass production, strengthening the capacities of both source and sink tissues have been aimed at, although with varying success64–71. Molecular targets were mainly related to genes involved in sugar and starch metabolism. A promising approach was recently suggested comprising the simultaneous boosting of both source and sink capacities using the crop plant potato as an example72. Here, the moderate repression of the leaf ADP-glucose pyrophosphorylase, a key enzyme of transitory starch formation or, alternatively, the mesophyll-specific expression of a bacterial pyrophosphate to stimulate sucrose synthesis and to prevent sucrose breakdown were combined with plants that had an increased sink strength. An increase in sink strength was accomplished by tuber-specific overexpression of two plastidic transporters responsible for the import of carbon (glucose 6-phosphate73) and energy (ATP74), respectively, into starch-storing amyloplasts. Both procedures resulted in an enhanced allocation of sucrose from source to sink tissues at the expense of leaf starch accumulation. The combined push-pull approach resulted in doubling the tuber starch yield72. This way of proceeding is currently being transferred to cassava, an important staple food in developing countries (Bill & Melinda Gates Foundation, CASS, OPP1113365).
It remains to be elucidated whether or not the push-pull approach can be combined with recent knowledge on novel photosynthetic components, e.g. by novel combinations of photosystem cores and LHC antenna complexes or by modifying light harvesting and regulators of photosynthetic electron flow to improve photosynthetic efficiency (Table 1). In addition, the push-pull strategy could potentially be teamed up with approaches to alter leaf form and/or anatomy to further improve εc, i.e. photosynthetic performance and biomass production.
Competing interests
The authors declare that they have no competing interests.
Grant information
Work in the authors´ laboratories cited in this review was supported by the German Research Foundation (DFG), the Federal Ministry of Education and Research (BMBF), and the European Union (project 3to4).
Faculty Opinions recommended
References
1.
Monteith JL, Moss CJ:
Climate and the efficiency of crop production in Britain.
Philos Trans R Soc Lond B Biol Sci.
1977; 281(980): 277–294. Publisher Full Text
2.
Long SP, Zhu X, Naidu SL, et al.:
Can improvement in photosynthesis increase crop yields?
Plant Cell Environ.
2006; 29(3): 315–30. PubMed Abstract
| Publisher Full Text
3.
Zhu X, Long SP, Ort DR:
Improving photosynthetic efficiency for greater yield.
Annu Rev Plant Biol.
2010; 61: 235–61. PubMed Abstract
| Publisher Full Text
4.
Zelitch I:
The close relationship between net photosynthesis and crop yield.
BioScience.
1982; 32(10): 796–802. Publisher Full Text
8.
Peng L, Shimizu H, Shikanai T:
The chloroplast NAD(P)H dehydrogenase complex interacts with photosystem I in Arabidopsis.
J Biol Chem.
2008; 283(50): 34873–9. PubMed Abstract
| Publisher Full Text
| Free Full Text
9.
DalCorso G, Pesaresi P, Masiero S, et al.:
A complex containing PGRL1 and PGR5 is involved in the switch between linear and cyclic electron flow in Arabidopsis.
Cell.
2008; 132(2): 273–85. PubMed Abstract
| Publisher Full Text
11.
Yokono M, Takabayashi A, Akimoto S, et al.:
A megacomplex composed of both photosystem reaction centres in higher plants.
Nat Commun.
2015; 6: 6675. PubMed Abstract
| Publisher Full Text
13.
Liu H, Zhang H, Niedzwiedzki DM, et al.:
Phycobilisomes supply excitations to both photosystems in a megacomplex in cyanobacteria.
Science.
2013; 342(6162): 1104–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
14.
Järvi S, Suorsa M, Paakkarinen V, et al.:
Optimized native gel systems for separation of thylakoid protein complexes: novel super- and mega-complexes.
Biochem J.
2011; 439(2): 207–14. PubMed Abstract
| Publisher Full Text
16.
Benson SL, Maheswaran P, Ware MA, et al.:
An intact light harvesting complex I antenna system is required for complete state transitions in Arabidopsis.
Nat Plants.
2015; 1: 15176. PubMed Abstract
| Publisher Full Text
19.
Benz M, Bals T, Gügel IL, et al.:
Alb4 of Arabidopsis promotes assembly and stabilization of a non chlorophyll-binding photosynthetic complex, the CF1CF0-ATP synthase.
Mol Plant.
2009; 2(6): 1410–24. PubMed Abstract
| Publisher Full Text
20.
Fristedt R, Martins NF, Strenkert D, et al.:
The thylakoid membrane protein CGL160 supports CF1CF0 ATP synthase accumulation in Arabidopsis thaliana.
PLoS One.
2015; 10(4): e0121658. PubMed Abstract
| Publisher Full Text
| Free Full Text
21.
Rühle T, Razeghi JA, Vamvaka E, et al.:
The Arabidopsis protein CONSERVED ONLY IN THE GREEN LINEAGE160 promotes the assembly of the membranous part of the chloroplast ATP synthase.
Plant Physiol.
2014; 165(1): 207–26. PubMed Abstract
| Publisher Full Text
| Free Full Text
22.
Mao J, Chi W, Ouyang M, et al.:
PAB is an assembly chaperone that functions downstream of chaperonin 60 in the assembly of chloroplast ATP synthase coupling factor 1.
Proc Natl Acad Sci U S A.
2015; 112(13): 4152–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
23.
Grahl S, Reiter B, Gügel IL, et al.:
The Arabidopsis Protein CGLD11 Is Required for Chloroplast ATP Synthase Accumulation.
Mol Plant.
2016; 9(6): 885–99. PubMed Abstract
| Publisher Full Text
24.
Zhang L, Duan Z, Zhang J, et al.:
BIOGENESIS FACTOR REQUIRED FOR ATP SYNTHASE 3 Facilitates Assembly of the Chloroplast ATP Synthase Complex.
Plant Physiol.
2016; 171(2): 1291–306. PubMed Abstract
| Publisher Full Text
| Free Full Text
25.
Armbruster U, Rühle T, Kreller R, et al.:
The photosynthesis affected mutant68-like protein evolved from a PSII assembly factor to mediate assembly of the chloroplast NAD(P)H dehydrogenase complex in Arabidopsis.
Plant Cell.
2013; 25(10): 3926–43. PubMed Abstract
| Publisher Full Text
| Free Full Text
26.
Rochaix JD, Lemeille S, Shapiguzov A, et al.:
Protein kinases and phosphatases involved in the acclimation of the photosynthetic apparatus to a changing light environment.
Philos Trans R Soc Lond B Biol Sci.
2012; 367(1608): 3466–74. PubMed Abstract
| Publisher Full Text
| Free Full Text
27.
Yamori W, Shikanai T:
Physiological Functions of Cyclic Electron Transport Around Photosystem I in Sustaining Photosynthesis and Plant Growth.
Annu Rev Plant Biol.
2016; 67: 81–106. PubMed Abstract
| Publisher Full Text
30.
Pribil M, Labs M, Leister D:
Structure and dynamics of thylakoids in land plants.
J Exp Bot.
2014; 65(8): 1955–72. PubMed Abstract
| Publisher Full Text
32.
Vainonen JP, Sakuragi Y, Stael S, et al.:
Light regulation of CaS, a novel phosphoprotein in the thylakoid membrane of Arabidopsis thaliana.
FEBS J.
2008; 275(8): 1767–77. PubMed Abstract
| Publisher Full Text
33.
Petroutsos D, Busch A, Janssen I, et al.:
The chloroplast calcium sensor CAS is required for photoacclimation in Chlamydomonas reinhardtii.
Plant Cell.
2011; 23(8): 2950–63. PubMed Abstract
| Publisher Full Text
| Free Full Text
38.
Pribil M, Pesaresi P, Hertle A, et al.:
Role of plastid protein phosphatase TAP38 in LHCII dephosphorylation and thylakoid electron flow.
PLoS Biol.
2010; 8(1): e1000288. PubMed Abstract
| Publisher Full Text
| Free Full Text
40.
Spreitzer RJ, Salvucci ME:
Rubisco: structure, regulatory interactions, and possibilities for a better enzyme.
Annu Rev Plant Biol.
2002; 53: 449–75. PubMed Abstract
| Publisher Full Text
41.
Bauwe H, Hagemann M, Fernie AR:
Photorespiration: players, partners and origin.
Trends Plant Sci.
2010; 15(6): 330–6. PubMed Abstract
| Publisher Full Text
43.
Parry MA, Andralojc PJ, Scales JC, et al.:
Rubisco activity and regulation as targets for crop improvement.
J Exp Bot.
2013; 64(3): 717–30. PubMed Abstract
| Publisher Full Text
44.
von Caemmerer S, Quick WP, Furbank RT:
The development of C4rice: current progress and future challenges.
Science.
2012; 336(6089): 1671–2. PubMed Abstract
| Publisher Full Text
45.
Price GD, Pengelly JJ, Forster B, et al.:
The cyanobacterial CCM as a source of genes for improving photosynthetic CO2 fixation in crop species.
J Exp Bot.
2013; 64(3): 753–68. PubMed Abstract
| Publisher Full Text
47.
Maier A, Fahnenstich H, von Caemmerer S, et al.:
Transgenic Introduction of a Glycolate Oxidative Cycle into A. thaliana Chloroplasts Leads to Growth Improvement.
Front Plant Sci.
2012; 3: 38. PubMed Abstract
| Publisher Full Text
| Free Full Text
48.
Raines CA:
Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: current and future strategies.
Plant Physiol.
2011; 155(1): 36–42. PubMed Abstract
| Publisher Full Text
| Free Full Text
50.
Blankenship RE, Chen M:
Spectral expansion and antenna reduction can enhance photosynthesis for energy production.
Curr Opin Chem Biol.
2013; 17(3): 457–61. PubMed Abstract
| Publisher Full Text
52.
Nicotra AB, Leigh A, Boyce CK, et al.:
The evolution and functional significance of leaf shape in the angiosperms.
Funct Plant Biol.
2011; 38(7): 535–552. Publisher Full Text
53.
Nicotra AB, Cosgrove MJ, Cowling A, et al.:
Leaf shape linked to photosynthetic rates and temperature optima in South African Pelargonium species.
Oecologia.
2008; 154(4): 625–35. PubMed Abstract
| Publisher Full Text
54.
Tholen D, Boom C, Zhu XG:
Opinion: prospects for improving photosynthesis by altering leaf anatomy.
Plant Sci.
2012; 197: 92–101. PubMed Abstract
| Publisher Full Text
56.
Adachi S, Nakae T, Uchida M, et al.:
The mesophyll anatomy enhancing CO2 diffusion is a key trait for improving rice photosynthesis.
J Exp Bot.
2013; 64(4): 1061–72. PubMed Abstract
| Publisher Full Text
57.
Fujita D, Trijatmiko KR, Tagle AG, et al.:
NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars.
Proc Natl Acad Sci U S A.
2013; 110(51): 20431–6. PubMed Abstract
| Publisher Full Text
| Free Full Text
58.
Zhang G, Li S, Wang L, et al.:
LSCHL4 from Japonica Cultivar, which is allelic to NAL1, increases yield of indica super rice 93-11.
Mol Plant.
2014; 7(8): 1350–64. PubMed Abstract
| Publisher Full Text
| Free Full Text
59.
Barkoulas M, Hay A, Kougioumoutzi E, et al.:
A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta.
Nat Genet.
2008; 40(9): 1136–41. PubMed Abstract
| Publisher Full Text
60.
Piazza P, Bailey CD, Cartolano M, et al.:
Arabidopsis thaliana leaf form evolved via loss of KNOX expression in leaves in association with a selective sweep.
Curr Biol.
2010; 20(24): 2223–8. PubMed Abstract
| Publisher Full Text
63.
Cartolano M, Pieper B, Lempe J, et al.:
Heterochrony underpins natural variation in Cardamine hirsuta leaf form.
Proc Natl Acad Sci U S A.
2015; 112(33): 10539–44. PubMed Abstract
| Publisher Full Text
| Free Full Text
64.
Leggewie G, Kolbe A, Lemoine R, et al.:
Overexpression of the sucrose transporter SoSUT1 in potato results in alterations in leaf carbon partitioning and in tuber metabolism but has little impact on tuber morphology.
Planta.
2003; 217(1): 158–67. PubMed Abstract
65.
Leidreiter K, Heineke D, Heldt HW, et al.:
Leaf-specific antisense inhibition of starch biosynthesis in transgenic potato plants leads to an increase in photoassimilate export from source leaves during the light period.
Plant Cell Physiol.
1995; 36(4): 615–624. Reference Source
66.
Sweetlove LJ, Kossmann J, Riesmeier JW, et al.:
The control of source to sink carbon flux during tuber development in potato.
Plant J.
1998; 15(5): 697–706. Publisher Full Text
67.
Zhang L, Häusler RE, Greiten C, et al.:
Overriding the co-limiting import of carbon and energy into tuber amyloplasts increases the starch content and yield of transgenic potato plants.
Plant Biotechnol J.
2008; 6(5): 453–64. PubMed Abstract
| Publisher Full Text
68.
Baroja-Fernández E, Muňoz FJ, Montero M, et al.:
Enhancing sucrose synthase activity in transgenic potato (Solanum tuberosum L.) tubers results in increased levels of starch, ADPglucose and UDPglucose and total yield.
Plant Cell Physiol.
2009; 50(9): 1651–62. PubMed Abstract
| Publisher Full Text
69.
Ferreira SJ, Sonnewald U:
The mode of sucrose degradation in potato tubers determines the fate of assimilate utilization.
Front Plant Sci.
2012; 3: 23. PubMed Abstract
| Publisher Full Text
| Free Full Text
70.
Li J, Baroja-Fernández E, Bahaji A, et al.:
Enhancing sucrose synthase activity results in increased levels of starch and ADP-glucose in maize (Zea mays L.) seed endosperms.
Plant Cell Physiol.
2013; 54(2): 282–94. PubMed Abstract
| Publisher Full Text
72.
Kammerer B, Fischer K, Hilpert B, et al.:
Molecular characterization of a carbon transporter in plastids from heterotrophic tissues: the glucose 6-phosphate/phosphate antiporter.
Plant Cell.
1998; 10(1): 105–17. PubMed Abstract
| Publisher Full Text
| Free Full Text
73.
Kampfenkel K, Möhlmann T, Batz O, et al.:
Molecular characterization of an Arabidopsis thaliana cDNA encoding a novel putative adenylate translocator of higher plants.
FEBS Lett.
1995; 374(3): 351–5. PubMed Abstract
| Publisher Full Text
74.
Che Y, Fu A, Hou X, et al.:
C-terminal processing of reaction center protein D1 is essential for the function and assembly of photosystem II in Arabidopsis.
Proc Natl Acad Sci U S A.
2013; 110(40): 16247–52. PubMed Abstract
| Publisher Full Text
| Free Full Text
75.
Wang P, Liu J, Liu B, et al.:
Evidence for a role of chloroplastic m-type thioredoxins in the biogenesis of photosystem II in Arabidopsis.
Plant Physiol.
2013; 163(4): 1710–28. PubMed Abstract
| Publisher Full Text
| Free Full Text
76.
Calderon RH, Garcia-Cerdán JG, Malnoë A, et al.:
A conserved rubredoxin is necessary for photosystem II accumulation in diverse oxygenic photoautotrophs.
J Biol Chem.
2013; 288(37): 26688–96. PubMed Abstract
| Publisher Full Text
| Free Full Text
77.
Huang W, Chen Q, Zhu Y, et al.:
Arabidopsis thylakoid formation 1 is a critical regulator for dynamics of PSII-LHCII complexes in leaf senescence and excess light.
Mol Plant.
2013; 6(5): 1673–91. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
78.
Wang Q, Sullivan RW, Kight A, et al.:
Deletion of the chloroplast-localized Thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves.
Plant Physiol.
2004; 136(3): 3594–604. PubMed Abstract
| Publisher Full Text
| Free Full Text
79.
Torabi S, Umate P, Manavski N, et al.:
PsbN is required for assembly of the photosystem II reaction center in Nicotiana tabacum.
Plant Cell.
2014; 26(3): 1183–99. PubMed Abstract
| Publisher Full Text
| Free Full Text
80.
Schneider A, Steinberger I, Strissel H, et al.:
The Arabidopsis Tellurite resistance C protein together with ALB3 is involved in photosystem II protein synthesis.
Plant J.
2014; 78(2): 344–56. PubMed Abstract
| Publisher Full Text
81.
Knoppová J, Yu J, Konik P, et al.:
CyanoP is Involved in the Early Steps of Photosystem II Assembly in the Cyanobacterium Synechocystis sp. PCC 6803.
Plant Cell Physiol.
2016; 57(9): 1921–31. PubMed Abstract
| Publisher Full Text
82.
Sakata S, Mizusawa N, Kubota-Kawai H, et al.:
Psb28 is involved in recovery of photosystem II at high temperature in Synechocystis sp. PCC 6803.
Biochim Biophys Acta.
2013; 1827(1): 50–9. PubMed Abstract
| Publisher Full Text
83.
Dobáková M, Sobotka R, Tichý M, et al.:
Psb28 protein is involved in the biogenesis of the photosystem II inner antenna CP47 (PsbB) in the cyanobacterium Synechocystis sp. PCC 6803.
Plant Physiol.
2009; 149(2): 1076–86. PubMed Abstract
| Publisher Full Text
| Free Full Text
84.
Yang H, Liao L, Bo T, et al.:
Slr0151 in Synechocystis sp. PCC 6803 is required for efficient repair of photosystem II under high-light condition.
J Integr Plant Biol.
2014; 56(12): 1136–50. PubMed Abstract
| Publisher Full Text
85.
Rast A, Rengstl B, Heinz S, et al.:
The Role of Slr0151, a Tetratricopeptide Repeat Protein from Synechocystis sp. PCC 6803, during Photosystem II Assembly and Repair.
Front Plant Sci.
2016; 7: 605. PubMed Abstract
| Publisher Full Text
| Free Full Text
88.
Fristedt R, Williams-Carrier R, Merchant SS, et al.:
A thylakoid membrane protein harboring a DnaJ-type zinc finger domain is required for photosystem I accumulation in plants.
J Biol Chem.
2014; 289(44): 30657–67. PubMed Abstract
| Publisher Full Text
| Free Full Text
89.
Järvi S, Suorsa M, Tadini L, et al.:
Thylakoid-Bound FtsH Proteins Facilitate Proper Biosynthesis of Photosystem I.
Plant Physiol.
2016; 171(12): 1333–43. PubMed Abstract
| Publisher Full Text
| Free Full Text
90.
Wulfhorst H, Franken LE, Wessinghage T, et al.:
The 5 kDa protein NdhP is essential for stable NDH-1L assembly in Thermosynechococcus elongatus.
PLoS One.
2014; 9(8): e103584. PubMed Abstract
| Publisher Full Text
| Free Full Text
91.
Yamamoto H, Fan X, Sugimoto K, et al.:
CHLORORESPIRATORY REDUCTION 9 is a Novel Factor Required for Formation of Subcomplex A of the Chloroplast NADH Dehydrogenase-Like Complex.
Plant Cell Physiol.
2016; 57(10): 2122–2132. PubMed Abstract
| Publisher Full Text
93.
Jin H, Liu B, Luo L, et al.:
HYPERSENSITIVE TO HIGH LIGHT1 interacts with LOW QUANTUM YIELD OF PHOTOSYSTEM II1 and functions in protection of photosystem II from photodamage in Arabidopsis.
Plant Cell.
2014; 26(3): 1213–29. PubMed Abstract
| Publisher Full Text
| Free Full Text
94.
Liu J, Last RL:
A land plant-specific thylakoid membrane protein contributes to photosystem II maintenance in Arabidopsis thaliana.
Plant J.
2015; 82(5): 731–43. PubMed Abstract
| Publisher Full Text
95.
Seok MS, You YN, Park HJ, et al.:
AtFKBP16-1, a chloroplast lumenal immunophilin, mediates response to photosynthetic stress by regulating PsaL stability.
Physiol Plant.
2014; 150(4): 620–31. PubMed Abstract
| Publisher Full Text
| Free Full Text
1
Cologne Biocenter, Botanical Institute II and Cluster of Excellence on Plant Sciences (CEPLAS), University of Cologne, Cologne, Germany 2
Department of Biology and Cluster of Excellence on Plant Sciences (CEPLAS), Heinrich-Heine University, Düsseldorf, Germany 3
Plant Molecular Biology, Department of Biology I, Ludwig-Maximilians-University, Munich, Germany
Work in the authors´ laboratories cited in this review was supported by the German Research Foundation (DFG), the Federal Ministry of Education and Research (BMBF), and the European Union (project 3to4).
Flügge UI, Westhoff P and Leister D. Recent advances in understanding photosynthesis [version 1; peer review: 3 approved]. F1000Research 2016, 5(F1000 Faculty Rev):2890 (https://doi.org/10.12688/f1000research.9744.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)