Keywords
microbial activity, inorganic compounds, metal-metabolism interactions
microbial activity, inorganic compounds, metal-metabolism interactions
In the book, The Hitchhiker’s Guide to the Galaxy, the Earth is described as “a computer of such infinite and subtle complexity that organic life itself shall form part of its operational matrix”1. Understanding the workings of the Earth as a deterministic computational entity remains a tantalizing object, and characterizing the relationship between organic life and the rest of the “operational matrix” (that is, inorganic geochemistry) is a central theme in the environmental sciences. Although careful studies have yielded insights into how physical and chemical laws influence microbial fitness and function in response to some environmental parameters, a major challenge lies in scaling laboratory experiments to landscape-wide predictions of gene and microbial fitness (Figure 1).
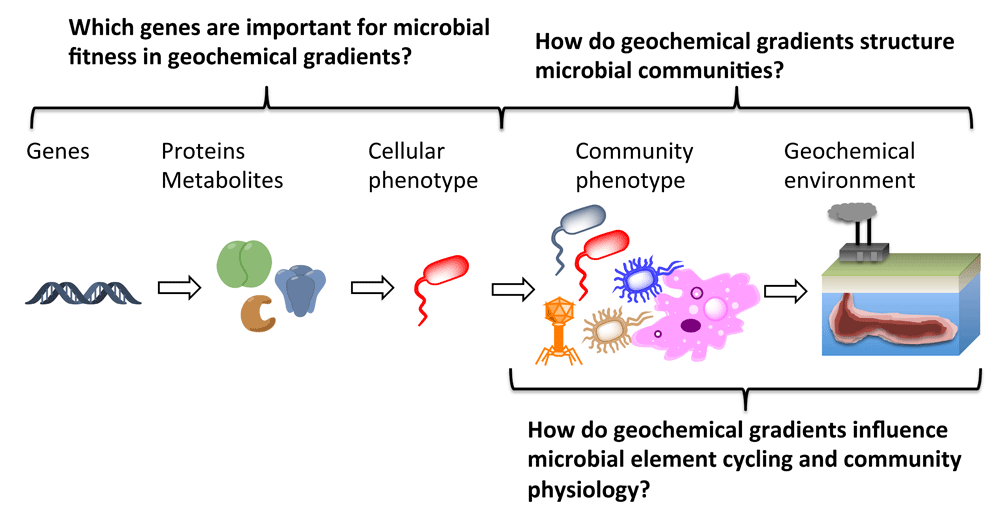
Understanding mechanisms whereby microorganisms survive in geochemical gradients is a central goal of environmental microbiology.
Great strides have been made in our ability to characterize the molecular composition of matter on Earth—from elemental analyses of sediments2,3 and structural characterization of natural organics4 to 'omics measurements of gene and protein content of natural environmental communities5,6. Technological advances in computation, data storage, and analytical tools enable this revolution. Alongside these advances is a similar, though often overlooked, revolution in robotics and laboratory automation. High-throughput cultivation in microtiter plates is possible both aerobically and anaerobically, and plate readers can be used to monitor optical density or metabolites using colorimetric assays7,8. It is also possible to fill microplates with arrays of compounds or serially diluted solutions to simultaneously evaluate the influence of hundreds to tens of thousands of parameters (for example, small-molecule libraries, inorganic ions, carbon sources, or other nutrients) on microbial growth kinetics and metabolism7. Additionally, recent advances in high-throughput genetics can be leveraged (in microbial isolates) to rapidly identify genetic determinants important for fitness in a given growth condition9–11 (Figure 2). Importantly, high-throughput assays can be used to quantitatively measure growth and respiratory activity of microbial cultures to define the fitness of a given microbial respiratory metabolism to defined gradients of compounds. Mass spectrometry–based metabolite analysis can give further insights into important metabolic signatures of this activity, and 16S amplicon sequencing can be used to monitor changes in the microbial community in response to these parameters. Subsequent growth-based assays with isolates from a given microbial enrichment culture can be used to measure isolate fitness and isolate gene fitness in response to the same gradients of compounds in which the enrichment was cultivated. Through measuring metabolic activity, microbial community structure, isolate fitness, and gene fitness in the context of gradients of environmentally relevant parameters, we can build models that link gene-, microbe-, and metabolism-specific fitness to environmental context (Figure 2). Through such workflows, environmental microbiologists now are able to re-array and reconstitute the purified organic and inorganic components of microbial ecosystems at an unprecedented scale and speed.
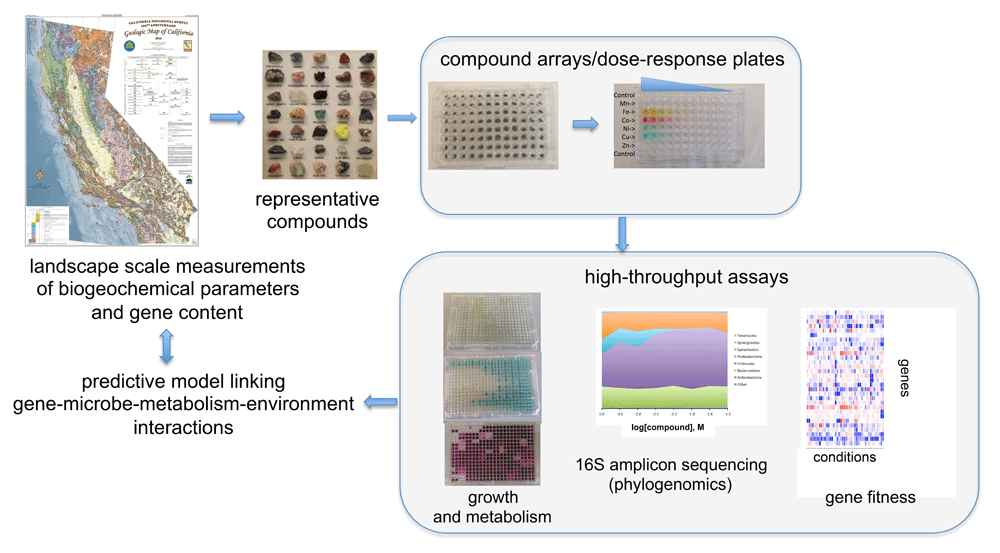
Measurements of metal content across rock, soil, and water samples can be obtained, and landscape-scale elemental maps can be constructed. Mineral samples and metal ions can be arrayed in microplates, and tagged-transposon pool assays, 16S amplicon sequencing, and metabolism-specific colorimetric assays can be employed to quantify the influence of concentrations of various metals on gene-microbe-metabolism fitness. Linking landscape-scale measurements of geochemistry to high-throughput laboratory measurements of microbial activity in response to geochemistry will enable higher-resolution biogeochemical models.
Microbes rely on both organic and inorganic cofactors and nutrients and live in complex multidimensional gradients of beneficial, neutral, and toxic compounds12,13. Microbial niche space is often viewed as an n-dimensional matrix in which antimetabolites, carbon sources, and essential nutrients influence the ability of a microorganism or microbial community to grow and survive13,14. Thus, only by altering the concentrations of multiple inorganic or organic compounds in a massively parallel, high-throughput cultivation platform can we gain a quantitative bottom-up picture of gene-microbe-metabolism-environment interactions. Several high-throughput approaches developed in the biomedical sciences can be applied to problems in environmental microbiology, including high-throughput screens to identify inhibitory compounds, dose-response microplate assays to quantify the inhibitory potency of compounds, checkerboard synergy assays to evaluate non-linear interactions between compounds, and leave-one-out assays to evaluate formulation potencies8,15. However, most of the compounds used in the biomedical industry (for example, drugs) are not environmentally relevant. To address this shortfall, we have begun to array metals and other inorganic compounds such as in an “80 metals plate” (Table 1) to create compound collections that more accurately capture the microbial stressors present in the environment. This arrayed compound collection can be serially diluted and added to microbial cultures to determine inhibitory concentrations such as minimal inhibitory concentration (MIC) or the concentration required to inhibit 50% of control growth (IC50). By varying the inoculum, respiratory substrates, and other parameters, experimentalists can gain insights into how other dimensions of the environment influence the inhibitory potency of these inorganic ions on gene-microbe-metabolism fitness.
Organic life exists and evolves in a matrix of both organic and inorganic compounds. One indelible mark of this evolutionary history consists of the diverse metallocofactors incorporated into enzymes that enable chemistry impossible for catalysts composed solely of C, H, N, O, P, and S16. High concentrations of metals are toxic to cells, and many metals also serve no catalytic role. Therefore, resistance mechanisms to metals have evolved. Metals are toxic to microorganisms because of their redox activity and because antimetabolic metals can compete with cofactor metals for binding to biological ligands and proteins17. Not surprisingly, microorganisms have evolved mechanisms for coping with metal stress, and these mechanisms vary by microorganism, metabolic state, or metal and are different depending on the metal concentration16,18. As an example, iron and its interactions with other transition metals and microbial cells are fairly well studied. Iron is an essential metal for a variety of metalloproteins. Under limiting concentrations of iron, other transition metals can interfere with high-affinity iron uptake systems and metalloregulatory proteins19, but at higher concentrations, some transition metals are toxic because of their ability to catalyze the production of reactive oxygen species20. Thus, the mechanism of toxicity and the mechanisms of resistance will be different depending on the concentrations of the metals. Very few studies systematically evaluate metal toxicity under both excess and limiting concentrations of essential metals, but by quantifying the toxicity of larger panels of metals under these conditions, we can obtain “structure-activity” information for inorganic compounds and their toxicity against, for example, uptake and efflux systems (Figure 3). A microplate array involving compounds such as the “80 metals plate” described in Table 1 could be serially diluted and used to evaluate the toxicity of many metals simultaneously against microbial isolates, enrichments, and pooled transposon mutants. Metal cations and oxyanions with varying ionic radii, charge, and electron affinity will vary in their interaction with different cellular systems. Only by quantifying the inhibitory potency of these metals under the various conditions under which these different cellular systems are important can we gain insights into how these systems have or have not evolved resistance to various metals. Ultimately, the data obtained through such studies will help geomicrobiologists to infer which metals may have been present in the environment in which a microbe evolved and to quantify the geochemical and genetic parameters that limit the growth of a microbial isolate or community in the environment.

The influence of a toxic metal will vary depending on the metabolism. For example, metabolism 1 and metabolism 2 could be aerobic respiration, nitrate reduction, sulfate reduction, and photosynthesis. Similarly, other metals (I) can serve as antimetabolic inhibitors of respiratory enzymes, competing with substrate (Sred) for binding and turnover to product (Sox). Depending on the inhibitory potency of the toxic metal (Mtox.), the requirements of the essential metal (Mnut.), and the inhibitory potency of a respiratory inhibitor (I), different metabolisms will have different environmental ranges in response to metal gradients.
Metal requirements and toxicity are influenced by the metabolic state of a microorganism. Microorganisms can grow with a range of electron donors, carbon sources, and electron acceptors. All of these metabolisms have unique metal requirements and sensitivities to inorganic antimetabolites and toxins. As such, metals can be selective inhibitors or promoters of different metabolisms. For example, zinc can be more inhibitory of bacteria growing under glucose catabolic conditions versus other carbon sources because zinc inhibits key enzymes in glycolysis21. Against respiratory sulfate reduction, monofluorophosphate, molybdate, and perchlorate are all selective inhibitors with varying selectivities, potencies, and modes of inhibition against the central enzymes in the sulfate reduction pathway22. Some redox-active metals are more inhibitory of aerobically growing cells than anaerobic cells because they can reduce oxygen to superoxide and catalyze Fenton chemistry. By quantifying the inhibitory or stimulatory potencies of large panels of inorganic compounds against microbial isolates and enrichments carrying out various metabolic activities selective compounds can be identified and the degree of their selectivity quantified. Quantification of these tipping points will improve biogeochemical reactive transport models that incorporate predictions of microbial metabolic activities.
Multidimensional microbiology is poised to become the norm in the 21st century. Alongside rapidly improving computational and analytical tools, high-throughput microbial physiology will enable massively parallel measurements of microbial fitness in complex gradients of environmentally relevant conditions. Rarefaction curves from genome sequencing datasets imply that the genetic diversity of life is not infinite23, nor is the elemental composition of biosphere. From this perspective, the “infinite and subtle complexity”1 of the natural world has more to do with the fractal complexity of natural gradients, heterogeneous mixtures in soil, complex water currents, and the corresponding conglomerate of microbial activity in this geochemical milieu. Thus, although we may not reach a comprehensive and flawless model of biogeochemical processes on Earth from bottom-up measurements of microbial fitness and physiology, we are likely to greatly increase the resolution of our models through careful, high-throughput experimentation.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Thank you for this beautiful piece of Science.
Recently, nickel-resistant bacteria have been isolated from the first time in human gut. https://doi.org/10.1016/j.nmni.2017.06.001
While the role of ... Continue reading To the Authors,
Thank you for this beautiful piece of Science.
Recently, nickel-resistant bacteria have been isolated from the first time in human gut. https://doi.org/10.1016/j.nmni.2017.06.001
While the role of heavy resistant bacteria in the environment has been extensively studied, the impact of these bacteria in human health needs further investigations.
Elena Angela Lusi
Thank you for this beautiful piece of Science.
Recently, nickel-resistant bacteria have been isolated from the first time in human gut. https://doi.org/10.1016/j.nmni.2017.06.001
While the role of heavy resistant bacteria in the environment has been extensively studied, the impact of these bacteria in human health needs further investigations.
Elena Angela Lusi