Nursyirwani N, Asmara W, Wahyuni AETH et al. Phenotype and genotype of lactic acid bacteria (LAB) isolated from the tiger grouper Epinephelus fuscoguttatus alimentary tract [version 1; peer review: 2 approved, 1 approved with reservations]. F1000Research 2017, 6:1984 (https://doi.org/10.12688/f1000research.12734.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Faculty of Fishery and Marine Science, Universitas Riau, Pekanbaru, Indonesia 2Faculty of Veterinary Medicine, Gadjah Mada University, Yogyakarta, Indonesia 3Department of Fisheries, Faculty of Agriculture, Gadjah Mada University, Yogyakarta, Indonesia 4Faculty of Marine and Fisheries, Syiah Kuala University, Banda Aceh, Indonesia
Nursyirwani Nursyirwani
Roles:
Conceptualization,
Data Curation,
Formal Analysis,
Funding Acquisition,
Methodology,
Project Administration,
Writing – Original Draft Preparation
Lactic acid bacteria (LAB) have been isolated successfully from the tiger grouper Epinephelusfuscoguttatus intestine. However, their genus or species have not been identified. Therefore, the objective of the present study was to characterize the three isolated LAB (KSBU-12C, KSBU-5Da, and KSBU-9) based on their phenotype and genotype. The LAB phenotype was examined by observing morphological features including cell morphology, spore production and motility. The physiological tests were performed in 6.5% NaCl at the temperatures of 10oC and 45oC, and the biochemical tests were evaluated based on the production of enzymes catalase, oxidase and arginine dehydrolase, following the Standard Analytical Profile Index, API 50 CH kit. The genotype was examined based on 16S rDNA gene sequence analysis , and the products were analyzed using the BLAST (Basic Local Alignment Search Tool) NCBI database. The three isolates (KSBU-5Da, KSBU-12C, and KSBU-9) were categorized into the genus Enterococcus. 16S rDNA sequence analysis indicated that the isolates had 99% similarity to E. hirae ATCC 9790, registered in GenBank with accession number NR_075022.1. It was concluded that the three LAB isolates taken from the tiger grouper Epinephelus fuscoguttatus are E. hirae.
Corresponding author:
Nursyirwani Nursyirwani
Competing interests:
No competing interests were disclosed.
Grant information:
This research was partially supported by Universitas Riau.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Lactic acid bacteria (LAB) have been investigated as probiotics in the feed of terrestrial and aquatic farmed animals1–7. This is based on the fact that LAB inhabit the human and animal gastrointestinal tract. This group of bacteria also convert lactose to acetic acid, thus decreasing gastrointestinal pH and naturally preventing colonization of harmful bacteria8. This is because they have the ability to produce the inhibitory materials that inhibit pathogen growth in the digestive tract9. Besides that, the probiotics can stimulate host immunity responses2,9,10.
LAB are a group of gram-positive, non-sporulating bacteria, in cocci or rod form. They can live in aerobic or facultative anaerobic conditions. LAB are able to produce lactic acid through the fermentation process of carbohydrates11,12, and the optimum growth is at pH 5.5 to 6.0, so growth is restricted to neutral and alkaline conditions13. Those naturally occurring bacteria are non-pathogenic to humans and animals, hence being generally recognized as safe (GRAS) organisms14.
According to Parada et al.15 and Carr et al.16 17l generas of LAB have been described worldwide. They are common in food products, for instance milk, meat, fruits, and vegetables, and also in genital and alimentary tracts of humans and animals17, including fish intestines18,19.
Molecular approaches based on DNA such as 16S rRNA sequencing have been used to characterize the intestinal microflora, and this method is proven to be accurate and fast20. For instance, Balcazar et al.21 have identified LAB isolates from salmonid fish using 16S rRNA genes, and they found that there were 6 species of LAB, namely Lactobacillus curvatus, L. sakei, L. plantarum, Lactococcus lactis subsp. cremoris, L. lactis subsp. lactis, Carnobacterium maltaromaticum, and Leuconostoc mesenteroides. Sun et al.22 investigated microbiota from gastrointestinal tracts of the orange-spotted grouper Epinephelus coioides, using a standard isolation procedure in combination with analysis of 16S rRNA sequences, and found several bacterial species, namely Vibrio parahaemolyticus, V. harveyi, V. metschnikovi, V. alginolyticus, Delftia acidovorans, Pseudomonas putida, Acinetobacter baumannii, Burkholderia cepacia, Erwinia carotovora, S. aureus, L. lactis, L. casei, E. faecium, Bacillus pumilus, B. clausii, and Psychrobacter sp.
Nursyirwani et al.23 successfully isolated three LAB from the tiger grouper alimentary tract, and found that LAB have antibacterial activity against Vibrio alginolyticus. However, the taxonomic classification of these bacteria has not been examined. Therefore, the objective of the present study was to characterize the three LAB isolates from tiger grouper based on their phenotype and genotype.
Methods
Collection of LAB isolates
The experiment was carried out in compliance with the ethical guidelines provided by the Research Institution of Riau University (SOP/02/PL/LPPM/2016). Lactic acid bacteria (LAB) had been isolated from the tiger grouper (E. fuscoguttatus) alimentary tract following the procedure of Bucio et al24. The tiger grouper Epinephelus fuscoguttatus was taken from the aquaculture pond in Situbondo of East Java, Indonesia. The alimentary tract was removed from the fish, opened, and the gut content was removed. Then, the internal wall of the tract was scraped gently, and the mucus was collected in a sterile test tube containing 9 ml of phosphate buffer saline (PBS) solution at pH 7.2. The suspension was serial in PBS solution from 10-1 to 10-6. A volume of 0.1 ml of each serial dilution was inoculated in petri-dish containing deMan-Rogasa-Sharpe (MRS, Oxoid) agar medium. All inoculated dishes were incubated at 37°C for 24–48 hours. The bacterial colonies were reinoculated in new fresh MRS agar medium. Then, the grown colonies were inoculated on medium agar of glucose-yeast extract-pepton, supplemented with CaCO3 (GYP+CaCO3) agar and incubated at 37°C for 24 hours. The presence of LAB was indicated by the growth of white colonies surrounded by clear zones. The isolates were then examined for gram staining and catalase tests. Gram-positive and negative catalase isolates were selected and reinoculated in new fresh MRS agar and incubated at 37°C for 24 hours. The isolates were stored in a refrigerator at a temperature of -10°C before being used for the next tests.
Bacterial isolates
The three LAB isolates (KSBU-12C, KSBU-5Da, and KSBU-9) previously isolated from the intestine of the tiger grouper were reinoculated in MRS broth (MRS, Merck). Phenotype characterization was based on cell morphology, physiology, and biochemical tests. Cell morphology involves observing the shape of the cells, spore production, and motility. Physiological tests were performed in 6.5% NaCl at 10°C and 45°C. Biochemical tests were based on the production of enzymes catalase, oxidase, and arginine dehydrolase following the Standard Analytical Profile Index, API 50 CH kit (BioMerieux SA, Marcy I’Etoil, France). Molecular characterization of 16S rDNA gene sequences allowed identification of the genotype of the LAB isolates.
DNA extraction
DNA of isolates KSBU-12C, KSBU-5Da, and KSBU-9 was extracted following the procedures of Ausubel et al25. Each of the isolates were reinoculated in 5ml of MRS broth and then incubated at 30°C for 48 hours.
The 1.5 ml culture was centrifuged at 13000 rpm for 2 minutes, the supernatant was discarded, the pellet was resuspended in 467 µL of TE buffer, and 30 µL of lysozyme was added and then incubated at 37°C for 30 minutes. The suspension was added to 30 µL of 10% SDS and mixed with 3 µL of K proteinase and incubated at 37°C for an hour. The mixture was added to 100 µL 5 M NaCl, mixed well, added to 80 µL of CTAB/NaCl solution, mixed thoroughly, and incubated for 10 minutes at 65°C. The mixture was further incubated at 65°C for 10 minutes, mixed by stirring up and down 100 times, and the same volume (0.7–0.8 ml) of phenol/chloroform/isoamyl-alcohol (PCIAA) was added, mixed well, and centrifuged for 5 minutes. The supernatant beneath the interphase was transferred to a new microtube, mixed with an equal volume of PCIAA, and centrifuged for 5 minutes.
The supernatant was placed into a new the microtube, then the isopropanol (60% of the volume) was added to the tube and centrifuged for two minutes. The supernatant was discarded, the DNA pellet was washed with 70% ethanol (±50 µL), centrifuged for 5 minutes, the ethanol was discarded, and the pellet was dried for 1 hour. Finally, the pellet was added to 100 µL TE buffer, vortexed, and stored at -20°C until it was used for further experiments.
Amplification of 16S rDNA
The genome of bacterial 16S rDNA was amplified by PCR using a thermal cycler (Eppendorf, Mastercycler Personal). The PCR reaction consisted of 25 µL of final solution of 12.5 µL PCR mix (Promega), 1.0 µL of universal primer (1st base) 24F (5-‘AGAG TTT GAT CCT GGC T-3’) and 1.0 µL primer 1540R (5-‘AAG G AGGT GAT CC AG CC GCA-3’), 0.5 µL of DNA template, and 10.0 µL of dH2O.
The thermocycler program was run with the following conditions: pre-denaturation at 94°C for 4 minutes; 29 cycles of denaturation at 94°C for 3 minutes; annealing at 55°C for 1 minute; extension at 72°C for 1 minute and 30 seconds; and the final extension at 72°C for 10 minutes. The PCR product was detected by running electrophoresis in 1% of agarose gel, stained with ethidium bromide (1 µL/10 ml), and visualized under a UV lamp for 1500 base pairs of the target product.
DNA sequence analysis
The PCR product with clear bands was sent to 1st Base Laboratories in Singapore for sequencing. The sequencing program was performed by ABI PRISM 3730×L GENETIC ANALYZER (Applied Biosystems, USA). The sequenced products were blasted (NCBI Basic Local Alignment Search Tool)26, and the results were presented as homology (%) of bacterial DNA sequences to the database sequences27.
Results
The three LAB isolates were categorized as cocci, gram-positive, non-motile, and non-spore forming. The LAB did not produce catalase, oxidase, and arginine dehydrolase, and were facultative anaerobes, with growth at 45°C at concentrations of 6.5% NaCl. However, differences were observed in the ability to produce acid from carbohydrates provided in the API 50 CH kit. Acid was not produced from substrates L-arabinose, D-raffinose, and D-xylose by the KSBU-12C isolate, meanwhile the KSBU-5Da isolate did not use L-arabinose, gluconate, glycerol, D-mannose, sorbitol, D-tagatose, and D-xylose; and the KSBU-9 isolate used L-arabinose, and D-xylose. The cell morphology, and the physiological and biochemical characteristics of the isolates are presented in detail in Table 1.
Table 1. Phenotype of KSBU-12C, KSBU-5Da, and KSBU-9 isolates in comparison to Enterococcus sp.28.
Phenotype characters
KSBU-12C
KSBU-5Da
KSBU-9
Group and species of Enterococcus
E. faecium
E. durans
E. hirae
Cell morphology
Shape
Coccus
Coccus
Coccus
Coccus
Coccus
Coccus
Gram staining
+
+
+
+
+
+
Spore formation
-
-
-
-
-
-
Motility
-
-
-
-
-
-
Use of pyruvate
n.a
n.a
n.a
-
-
-
Production of:
Catalase
-
-
-
-
-
-
Oxidase
-
-
-
-
-
-
Alkaline phosphatase
n.a
n.a
n.a
-
-
-
Arginine dehydrolase
-
-
-
+
+
+
Pyrrolidonyl arylamidase
n.a
n.a
n.a
+
+
+
O/F
O/F
O/F
O/F
n.a
n.a
n.a
Growth at:
10°C
n.a
n.a
n.a
+
+
+
45°C
+
+
+
+
+
+
NaCl 6.5%
+
+
+
+
+
+
Oxygen requirement
Anaerobic facultative
Anaerobic facultative
Anaerobic facultative
Anaerobic facultative
Anaerobic facultative
Anaerobic facultative
Production of :
Acetoin
n.a
n.a
n.a
+
+
+
Antigen group D
n.a
n.a
n.a
+
+
+
Leucine arylamidase
n.a
n.a
n.a
+
+
+
α-Galactosidase
n.a
n.a
n.a
-
-
+
β-Galactosidase
n.a
n.a
n.a
+
D
+
β-Glucuronidase
n.a
n.a
n.a
-
-
-
Hydrolysis of:
Esculin
+
+
+
+
+
+
Hippurate
n.a
n.a
n.a
d
D
D
Starch
n.a
n.a
n.a
-
D
Acid from:
Adonitol
-
-
-
-
-
-
D-Arabitol
-
-
-
-
-
-
Inulin
-
-
-
-
-
-
Melezitose
+
+
+
-
-
D
Ribose
+
+
+
+
+
+
L-Sorbose
-
-
-
-
-
-
N-Acetylglucosamine
+
+
+
+
+
+
Amygdalin
+
+
+
+
+
+
D-Arabinose
-
-
-
-
-
-
L-Arabinose
-
-
+
+
-
-
L-Arabitol
-
-
-
-
-
-
Arbutin
+
+
+
+
+
+
Cellobiose
+
+
+
+
+
+
Dulcitol
-
-
-
-
-
-
Erythritol
-
-
-
-
-
-
D-Fructose
+
+
+
+
+
+
D-Fucose
-
-
-
-
-
-
L-Fucose
-
-
-
-
-
-
Galactose
+
+
+
+
+
+
β-Gentiobiose
+
+
+
+
+
+
Gluconate
+
-
+
d
-
-
D-Glucose
+
+
+
+
+
+
Glycerol
+
-
+
d
-
d
Glycogen
-
-
-
-
-
-
Inositol
-
-
-
-
-
-
2-Keto-gluconate
-
-
-
-
-
-
5-Keto-gluconate
-
-
-
-
-
-
Lactose
+
+
+
+
+
+
D-Lyxose
-
-
-
-
-
-
Maltose
+
+
+
+
+
+
Mannitol
+
+
+
d
-
-
D-Mannose
+
-
+
+
+
+
Melibiose
+
+
+
d
D
d
α-Methyl-D-glucoside
-
-
-
-
-
-
α-Methyl-D-mannose
-
-
-
d
-
-
D-Raffinose
-
+
+
d
D
d
Rhamnose
-
-
-
d
-
-
Sorbitol
+
-
+
d
-
-
Starch (Amidon)
-
-
-
-
-
d
Sucrose
+
+
+
d
D
d
D-Tagatose
+
-
+
d
-
d
Trehalose
+
+
+
+
+
+
D-Turanose
-
-
-
-
-
d
Xylitol
-
-
-
-
-
-
D-Xylose
-
-
+
d
-
-
L-Xylose
-
-
-
-
-
-
Note: +, 90% or most strains are positive; -, 90% or most data are negative; O/F, oxidative or fermentative; nd, data not available; d, variable.
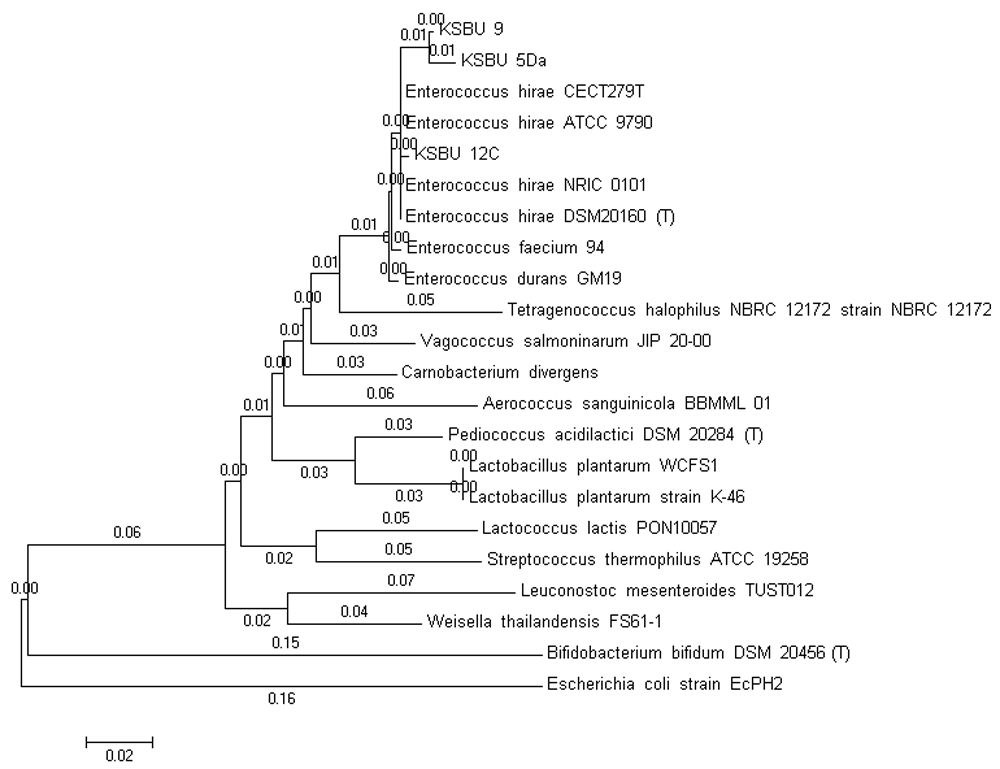
The 16S rRNA derived from the LAB isolates were amplified by PCR with two universal primers 24F (5-‘AGAG TTT GAT CCT GGC T-3’) and 1540R (5-‘AAG G AGGT GAT CC AG CC GCA-3’). The LAB isolates have a highly similar 16S rRNA sequence (99% similarity as shown in Table 2; those were 1510-bp for KSBU-12C and 1526-bp for KSBU-5Da and KSBU-9) (Figure 1). A phylogenetic tree was constructed for the LAB isolates sequences using Clustal W, followed by the Mega 5 neighbor-joining program as shown in Figure 2.
Table 2. BLAST results of LAB isolates KSBU-12C, KSBU-5Da, and KSBU-9.
Figure 1. Purification product of 16S rDNA LAB genome of (1) KSBU-5Da; (2) KSBU-9; (3) KSBU-12C isolates; (4) marker DNA ladder (bp).
Figure 2. Phylogenetic tree constructed by the neighbor-joining method based on partial sequence of 16S rRNA gene for KSBU-12C, KSBU-5Da, and KSBU-9 isolated from tiger grouper.
Scale 0.02 indicates sequence difference percentage. Escherichia coli strain EcPH2 was used as the outer group.
Discussion
Based on morphological, physiological, and biochemical characteristics, the LAB isolates (KSBU-12C, KSBU-5Da, and KSBU-9) show similarity to three bacterial species of the Enterococcus genus found by Ludwig et al.28. The analysis of the 16S rDNA sequence indicated that KSBU-5Da, KSBU-12C, and KSBU-9 isolates were of the Enterococcus genus, and with high similarity to E. hirae ATCC-9790 registered in GeneBank with accession number NR_075022.1 (Table 2). Each of the LAB isolates indicated 99% homology with E. hirae ATCC 9790. Strains of bacteria with the same or more than 97% 16S rRNA gene sequence belonged to one species29.
E. hirae ATCC 9790 is a gram-positive LAB used as a model organism in basic research for four decades30. E. hirae was first isolated in the intestines of pigs and chickens31. E. hirae strain NRIC 0101 was collected by the Japan Nodai Culture Collection Center, NCCC32. In addition, E.hirae C17456 was a species isolated from chickens33. In summary, Enterococcus hirae has been found in humans, poultry, foods for instant dosa batter, and in the environment34–37. Together with E. faecium, E. faecalis, E.casseliflavus, and E. mundtii they are enterococci which were predominant in coastal waters and sediments of South of California38. E. hirae was one of the homofermentative strains obtained from traditional fermentation products such as pla-som and pla-chom (fermented fish) in Thailand39. It has been shown that E. hirae K34, L. plantarum K39, and L. plantarum K50 isolated from Kung-Som (fermentative shrimp) display antimicrobial activity against pathogenic bacterial strains of Bacillus cereus, E. coli, Staphylococcus aureus, Salmonella typhimurium, Vibrio cholerae, and Listeria monocytogenes40.
Information on the use of E. hirae as a probiotic in aquaculture is still limited. Mazurkiewicz et al.41 reported that E. hirae isolated from the intestine of common carp (Cyprinus carpio L.) and applied into feed did not have a significant effect on the growth performance of the common carp. In contrary, another report by Adnan et al.42 and Carlos et al.43 showed that E. hirae from freshwater fish Catla catla and rainbow trout Oncorhynchus mykiss had a significant effect on inhibiting the growth of pathogens such as Escherichia coli, Staphylococcus aureus, Salmonella typhi and Pseudomonas spp.
In the present study, the role of E. hirae isolated from tiger grouper fish the growth performance of fish has not been evaluated yet. Therefore, further study will be needed to examine the effect of these bacteria on the feeding of cultured fish.
Conclusions
This research has successfully characterized three of the LAB isolates (KSBU-12C, KSBU-5Da, and KSBU-9) based on their phenotype and genotype. All isolates were determined to be E. hirae.
Data availability
Sequenced DNA of LAB isolates can be found in NCBI GenBank, with accession numbers MF977716 to MF977718.
Competing interests
No competing interests were disclosed.
Grant information
This research was partially supported by Universitas Riau.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgements
The authors would like to thank the staff at the Microbiology Laboratory of the Fishery Department, Gadjah Mada University, Indonesia.
Faculty Opinions recommended
References
1.
Arisa II, Widanarni W, Yuhana M, et al.:
The application of probiotics, prebiotics and synbiotics to enhance the immune responses of vannamei shrimp (Litopenaeus vannamei) to Vibrio harveyi infection.
AACL Bioflux.
2015; 8(5): 772–778. Reference Source
2.
Yulvizar C, Dewiyanti I, Defira CN, et al.:
Pathogenicity assay of probiotic potential bacteria from the common carp Cyprinus carpio.AACL Bioflux.
2015; 8(5): 694–698. Reference Source
3.
Muchlisin ZA, Murda T, Yulvizar C, et al.:
Growth performance and feed utilization of keureling fish Tor tambra (Cyprinidae) fed formulated diet supplemented with enhanced probiotic. [version 1; referees: 2 approved].
F1000Res.
2017; 6: 137. PubMed Abstract
| Publisher Full Text
| Free Full Text
4.
Amenu D:
Probiotic properties of Lactic Acid Bacteria from human milk.
J Med MicrobDiagn.
2014; S3: 005. Publisher Full Text
5.
Serrano-Nino JC, Solis-Pacheco JR, Gutlerrez-Padilla JA, et al.:
Isolation and identification of lactic acid bacteria from human milk with potential probiotic role.
J Food Nutr Res.
2016; 4(3): 170–177. Reference Source
6.
Wang D, Liu W, Ren Y, et al.:
Isolation and Identification of Lactic Acid Bacteria from Traditional Dairy Products in Baotou and Bayannur of Midwestern Inner Mongolia and q-PCR Analysis of Predominant Species.
Korean J Food Sci Anim Resour.
2016; 36(4): 499–507. PubMed Abstract
| Publisher Full Text
| Free Full Text
7.
Chowdhury T, Islam S:
Isolation, identification and determination of probiotic potential of lactic acid bacteria from local curd.
Int J Sci Eng Res.
2016; 7(4): 263–267. Reference Source
8.
Watson AK, Kaspar H, Lategan MJ, et al.:
Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes.
Aquaculture.
2008; 274(1): 1–14. Publisher Full Text
9.
Sugimura Y, Hagi T, Hoshina T:
Correlation between in vitro mucus adhesion and the in vivo colonization ability of lactic acid bacteria: screening of new candidate carp probiotics.
Biosci Biotechnol Biochem.
2011; 75(3): 511–515. PubMed Abstract
| Publisher Full Text
10.
Oke AO, Olaoye OJ, Nnali KE:
Recent advances in fish diseases treatment: Probiotics as alternative therapy to antibiotics in aquaculture “A Review”.
Advances in Agriculture Sciences and Engineering Research.
2013; 3(2): 668–676. Reference Source
11.
Axelsson L:
Lactic acid bacteria: classification and physiology.
Food Science and Technology.
2004; 139: 1–66. Reference Source
12.
Hayek SA, Ibrahim SA:
Current limitations and challenges with lactic acid bacteria: A Review.
Food Nutr Sci.
2013; 4: 73–87. Publisher Full Text
13.
Khalil MI, Anwar MN:
Isolation, identification and characterization of lactic acid bacteria from milk and yoghurts. Research and Review.
Journal of Food and Dairy Technology.
2016; 4(3): 17–26. Reference Source
14.
Patil MM, Pal A, Anand T, et al.:
Isolation and characterization of lactic acid bacteria from curd and cucumber.
Indian J Biotechnol.
2010; 9: 166–172. Reference Source
15.
Parada JL, Sambucetti ME, Zuleta A, et al.:
Lactic acid fermented products as vehicles for probiotics. In:
New Horizons in Biotechnology.
Kluwer Academic Publishers, Boston, 2003; 335–351. Publisher Full Text
16.
Carr FJ, Hill D, Maida N:
The lactic acid bacteria: a literature survey.
Crit Rev Microbiol.
2002; 28(4): 281–370. PubMed Abstract
| Publisher Full Text
17.
Françoise L:
Occurrence and role of lactic acid bacteria in seafood products.
Food Microbiol.
2010; 27(6): 698–709. PubMed Abstract
| Publisher Full Text
18.
Yang GM, Bao BL, Peatman E, et al.:
Analysis of the composition of the bacterial community in puffer fish Takifugu obscurus.Aquaculture.
2007; 262(2–4): 183–191. Publisher Full Text
19.
Ringo E:
The ability of carnobacteria isolated from fish intestine to inhibit growth of fish pathogenic bacteria: a screening study.
Aquaculture Research.
2008; 39(2): 171–180. Publisher Full Text
20.
O’Sullivan DJ:
Methods for analysis of the intestinal microflora.
Curr Issues Intest Microbiol.
2000; 1(2): 39–50. PubMed Abstract
21.
Balcázar JL, De-Blas I, Ruiz-Zarzuela I, et al.:
Sequencing of variable regions of the 16S rRNA gene for identification of lactic acid bacteria isolated from the intestinal microbiota of healthy salmonids.
Comp Immunol Microbiol Infect Dis.
2007; 30(2): 111–118. PubMed Abstract
| Publisher Full Text
22.
Sun Y, Yang H, Ling Z, et al.:
Gut microbiota of fast and slow growing grouper Epinepheluscoioides.
Afr J Microbiol Res.
2009; 3(11): 713–720. Reference Source
23.
Nursyirwani, Asmara W, Wahyuni AETH, et al.:
Isolasi bakteri asam laktat dari usus ikan kerapu macan (Epinephelus fuscoguttatus) dan potensinya sebagai antivibrio.
Ilmu Kelautan Indonesian Journal of Marine Sciences.
2011; 16(2): 70–77. Reference Source
24.
Bucio A, Hartemink R, Schrama JW, et al.:
Presence of Lactobacilli in the intestinal content of freshwater fish from a river and from a farm with a recirculation system.
Food Microbiol.
2006; 23(5): 476–482. PubMed Abstract
| Publisher Full Text
25.
Ausubel FM, Brent R, Kingston RE, et al.:
Current Protocols in Molecular Biology. John Wiley, and Sons, Inc. Cambridge, Massachusetts. 2003. Reference Source
26.
Muchlisin ZA, Batubara AS, Fadli N, et al.:
Assessing the species composition of tropical eels (Anguillidae) in Aceh Waters, Indonesia, with DNA barcoding gene cox1. [version 1; referees: 1 approved, 2 approved with reservations].
F1000Res.
2017; 6: 258. PubMed Abstract
| Publisher Full Text
| Free Full Text
27.
Diop MB, Dubois-Dauphin R, Tine E, et al.:
Bacteriocin producers from traditional food products.
Biotechnol Agron Soc Environ.
2007; 11(4): 275–281. Reference Source
28.
Ludwig W, Schleifer KH, Whitman WB:
Family IV. Enterococcaceae fam. nov. In: De Vos P, GM Garrity, D Jones, NR Krieg, W Ludwig, FA Rainey, K-H Schleifer and WB Whitman (Eds) Bergey’s Manual of Systematic Bacteriology. Second Edition. The Firmicutes. Springer Dordrecht Heidelberg, London New York, 2009; 3: 594–623. Reference Source
29.
Madigan MT, Martinko JM, Stahl DA, et al.:
Brock Biology of Microorganisms. Thirteenth Edition. Benjamin Cummings, San Francisco. 2012. Reference Source
30.
Gaechter T, Wunderlin C, Schmidheini T, et al.:
Genome Sequence of Enterococcus hirae (Streptococcus faecalis) ATCC 9790, a model organism for the study of ion transport, bioenergetics, and copper homeostasis.
J Bacteriol.
2012; 194(18): 5126–5127. PubMed Abstract
| Publisher Full Text
| Free Full Text
31.
Farrow JAE, Collins MD:
Enterococcus hirae, a new species that includes amino acid assay strain NCDO 1258 and strains causing growth depression in young chickens.
Int J Syst Bacteriol.
1985; 35(1): 73–75. Publisher Full Text
32.
TanakaN, Nakano M, Okada S:
16S rRNA gene sequences of NRIC Lactic Acid Bacteria strains.
Published Only in Database.
2007.
33.
Chadfield MS, Christensen JP, Juhl-Hansen J, et al.:
Characterization of Enterococcus hirae outbreaks in broiler flocks demonstrating increased mortality because of septicemia and endocarditis and/or altered production parameters.
Avian Dis.
2005; 49(1): 16–23. PubMed Abstract
| Publisher Full Text
34.
Iweriebor BC, Gaqavu S, Obi LC, et al.:
Antibiotic susceptibilities of Enterococcus species isolated from hospital and domestic wastewater effluents in Alice, Eastern Cape Province of South Africa.
Int J Environ Res Public Health.
2015; 12(4): 4231–4246. PubMed Abstract
| Publisher Full Text
| Free Full Text
35.
Arokiyaraj S, Hairul Islam VI, Bharanidharan R, et al.:
Antibacterial, anti-inflammatory and probioticpotential of Enterococcus hirae isolated from the rumen of Bos primigenius.World J Mirobiol Biotechnol.
2014; 30(7): 2111–2118. PubMed Abstract
| Publisher Full Text
36.
Gupta A, Tiwari SK:
Probiotic potential of bacteriocin-producing Enterococcus hirae strain LD3 isolated from dosa batter.
Ann Microbiol.
2015; 65(4): 2333–2342. Publisher Full Text
37.
Fisher K, Phillips C:
The ecology, epidemiology and virulence of Enterococcus.
Microbiology.
2009; 155(Pt 6): 1749–1757. PubMed Abstract
| Publisher Full Text
38.
Ferguson DM, Moore DF, Getrich MA, et al.:
Enumeration and speciation of enterococci found in marine and intertidal sediments and coastal water in southern California.
J Appl Microbiol.
2005; 99(3): 598–608. PubMed Abstract
| Publisher Full Text
40.
Dangkhaw N, Maneerat S, Sumpavapol P:
Characterization of Lactic Acid Bacteria Isolated From Kung-Som, A Traditional Fermented Shrimp, in Respect of Their Probiotic Properties.
International Conference on Nutrition and Food Sciences IPCBEE.
IACSIT Press, Singapore. 2012; 39: 121–125. Reference Source
41.
Mazurkiewickz J, Przybyl A, Sip A, et al.:
Effect of Carnobacterium divergens and Enterococcus hirae as probiotic bacteria in feed for common carp, Cyprinuscarpio L.
Archives of Polish Fisheries.
2007; 15(2): 93–102. Reference Source
42.
Adnan M, Patel M, Hadi S:
Functional and health promoting inherent attributes of Enterococcus hirae F2 as a novel probiotic isolated from the digestive tract of the freshwater fish Catla catla.PeerJ.
2017; 5: e3085. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Araújo C, Muñoz-Atienza E, Hernández PE, et al.:
Evaluation of Enterococcus spp. from rainbow trout (Oncorhynchus mykiss, Walbaum), feed and rearing environment against fish pathogens.
Foodborne Pathog Dis.
2015; 12(4): 311–322. PubMed Abstract
| Publisher Full Text
1
Faculty of Fishery and Marine Science, Universitas Riau, Pekanbaru, Indonesia 2
Faculty of Veterinary Medicine, Gadjah Mada University, Yogyakarta, Indonesia 3
Department of Fisheries, Faculty of Agriculture, Gadjah Mada University, Yogyakarta, Indonesia 4
Faculty of Marine and Fisheries, Syiah Kuala University, Banda Aceh, Indonesia
Nursyirwani Nursyirwani
Roles:
Conceptualization,
Data Curation,
Formal Analysis,
Funding Acquisition,
Methodology,
Project Administration,
Writing – Original Draft Preparation
This research was partially supported by Universitas Riau.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Nursyirwani N, Asmara W, Wahyuni AETH et al. Phenotype and genotype of lactic acid bacteria (LAB) isolated from the tiger grouper Epinephelus fuscoguttatus alimentary tract [version 1; peer review: 2 approved, 1 approved with reservations]. F1000Research 2017, 6:1984 (https://doi.org/10.12688/f1000research.12734.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Thank you, here are some comments about the contents of the article.
In the background it has not been explained specifically why the characterization of lactic acid bacteria isolated from the grouper intestine. The question is
... Continue reading
Thank you, here are some comments about the contents of the article.
In the background it has not been explained specifically why the characterization of lactic acid bacteria isolated from the grouper intestine. The question is that the bacteria will be mixed with fish feed in order to increase the digestibility of the food or the prevention of pathogens that attack groupers.
Procedure of taking bacterial inoculum in the grouper's bowel. Is the fish fasted first? This is to know the bacteria do live in the intestines of grouper fish instead of bacterial contaminants derived from food given.
In the discussion section has not been submitted phenotype characteristics of three types of bacterial isolates LAB. These traits include color, colony shape, surface roughness of the colony and will be better when displayed images with given information.
The RNA sequence results also need to be displayed to make the right conclusions in the characterization process. It would be great if supported with comparative literature.
It would be great if the discussion section uses more literature to discuss especially the latest journals.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
I cannot comment. A qualified statistician is required.
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Yes
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: My research in the field of ecotoxicology. My focus is primarily about the expression of protein or certain enzymes as biomarkers due to pollution of the aquatic environment.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
This article has excellent relevance value to both the development of marine science and is suitable for publication. This article should be indexed when minor improvements have been made.
The abstract shows that this research is interesting.
... Continue reading
This article has excellent relevance value to both the development of marine science and is suitable for publication. This article should be indexed when minor improvements have been made.
The abstract shows that this research is interesting.
How many isolation works? Of all isolation, three isolates were selected. The method used in the study should be listed on the abstract. Show keywords, keywords generally refer from general to specific, and not mentioned in title or abstract.
Introduction: It needs to be rewritten more purposefully, including backgrounds on topic selection and importance. Writing in English should pay attention to diction and grammar.
Method: It needs to include the reference method used. The exposure method has not been written in a systematic or accurate way, so it can be repeated by others.
Results and Discussion: There are still repeated discussions. References used are not current, need to use the latest references.
An annotated version of the Word document can be viewed here.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Yes
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Yes
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Marine natural product
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
The paper describes the identification of three LAB isolates from the alimentary tract of a tiger grouper. The identification was done using various phenotypic and genotypic methods.
However, there are some remarks or suggestions that can be made:
... Continue reading
The paper describes the identification of three LAB isolates from the alimentary tract of a tiger grouper. The identification was done using various phenotypic and genotypic methods.
However, there are some remarks or suggestions that can be made:
- First of all, the authors mention that LAB strains were previously isolated; why they included then in the Methods section and how they were isolated?; on the other hand, they do not mention from how many individuals they isolated these strains from? Were they isolated from one fish? Then, in fact, it is only one strain, since all three were identified in the end as Ent. hirae.
- Introduction:
"According to Pareda et al. ... 17l genera..." - that "l" doesn't belong there.
I think the authors should use "microbiota" instead of "microflora".
- Methods:
- Collection of LAB isolates
"The suspension was serial in PBS..." - I think "diluted" is missing.
"Gram positive and negative catalase isolates..." should be "Gram positive and catalase negative isolates..."
It is not clear how they kept the isolates, at -10 degrees, but on agar media?
The subtitle "Bacterial isolates" should be "Identification of the bacterial isolates" since they are talking about identification.
"Physiological tests were performed in 6.5% NaCl..." should be maybe "... in the presence of 6.5% NaCl..."
"... production of enzymes such as..."
- Amplification of 16S rDNA
"The genome of bacterial 16S rDNA..." should be " The 16S rDNA gene from the bacterial genome..."
- Results:
In Table 1 there is a discrepancy between the legend and the table. For instance, in the Table they have "n.a"., while in the legend is "n.d".; in the table they have both "d" and "D", and in the legend only "d". Moreover, there are many rows with data only for the Enterococcus species, but for the strains isolated by them, there are no data available. What is the point to show these things?
Some information from the Methods is repeated in the Results, such as the primers used.
The paragraph with the similarities among the sequences and 16S rDNA amplicons, with Fig. 1 and Table 2, is not very clear. Perhaps they should mention first that the amplicons obtained with the universal primers were very similar (and give their size and the reference to Fig. 1) and then they should mention about the high similarity among the sequences (99%) with the sequence of Ent. hirae (reference to Table 2).
- Discussion:
This section can be improved, maybe with information about the source of this strain in the alimentary tract, the importance for the host, why was the only strain found in their samples, etc.
There is a paragraph that I would suggest to be changed in: " Strains... with the same or more than 97% homology of the 16S rRNA gene sequence may be considered to belong to the same species".
"Ent. hirae was first isolated..." - I think the word "from" is missing.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Partly
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Not applicable
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Partly
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles

Comments on this article Comments (0)