Introduction
Finding appropriate therapeutic cells for bone regeneration has been a challenge for decades. Recently, stem cells from the skin, a potentially large cell source with easy access, have caught the attention of clinicians and scientists. More and more evidence indicates that skin stem cells are a potential cell source for bone regeneration. For example, heterozygous inactivating mutations of GNAS (encoding guanine nucleotide-binding G protein alpha subunit) cause diseases, including progressive osseous heteroplasia, Albright hereditary osteodystrophy, pseudohypoparathyroidism, and osteoma cutis1–4. These disorders have the common features of superficial ossification, starting with cutaneous ossification, with some involving subcutaneous and deeper tissues and some restricted to the skin. Multipotent progenitor cells and bone morphogenetic proteins (BMPs) were reported to be responsible for ectopic ossification5,6.
Despite a decade of investigations using skin stem cells for regenerative medicine, most literature concerns their application in skin tissue engineering7 and nerve regeneration8, which was well covered by a recent review article9. However, few review articles are available on skin cell-based osteogenesis. This review first summarizes the latest findings on stem cells or progenitors in skin and their niches and then discusses the strategies of skin cell-based osteogenesis (Figure 1). We hope this article elucidates this topic and generates new ideas for future studies.
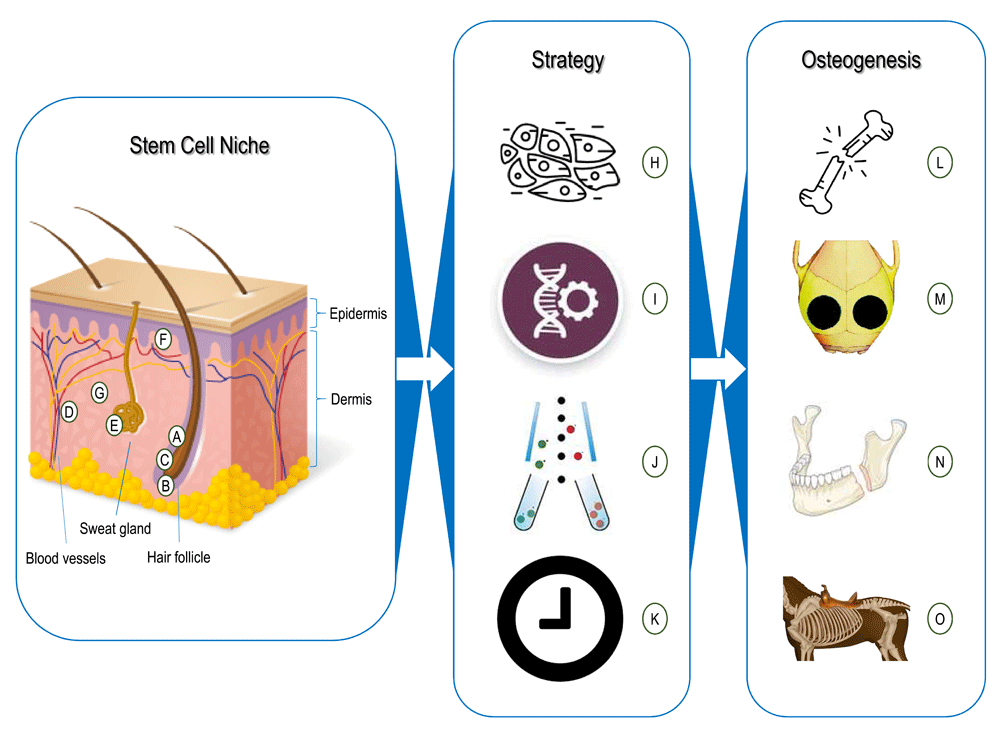
Figure 1. Skin cells for osteogenesis.
(A–G) Stem cells and niches found in skin. (A) Hair follicle bulge-derived stem cells11,12,15. (B) Hair follicle papilla-derived stem cells18,22–24. (C) Hair sheath-derived stem cells16,22. (D) Pericytes10,51. (E) Sweat gland-derived stem cells25,26. (F) Interfollicle epidermis-derived stem cells13,14. (G) Stem cells from dermal niches that are not fully characterized27–34,50,52,53. (H–K) Strategies for using skin cells. (H) Total skin fibroblasts35,36. (I) Genetic modification38–48. (J) Cell sorting33,50–53. (K) Cell reprogramming56–58,65. (L–O) Skin cells’ osteogenesis. (L) Limb bone defect regeneration35,41,42. (M) Cranial bone defect regeneration38,43,44,53. (N) Mandibular bone defect regeneration40,48. (O) Rib bone defect regeneration45.
Characteristics of skin stem cells and niches
Besides the primary structure of the epidermis, dermis, and subcutaneous tissue, there are hair follicles, vessels, capillaries, neurons, sweat glands, sebaceous glands, lymphatic capillaries, and erector pili muscles in skin, implying that there could be numerous niches for stem cells and progenitors in this tissue (Table 1). Evidence also indicates that stem cells in skin, so-called pericytes, might be of perivascular origin10.
Table 1. Characterization of skin stem cells and niches.
| Location | Niche | Culture | Name | Markers | Differentiation potential | References |
|---|
| Epidermis | Interfollicle
epidermis | Adherence | Epidermal
stem cells | α6 integrin, β1 integrins,
CD133, CD90, and keratin 15 | Keratinocytes | 13,14 |
Hair
follicle and
appendages | Hair follicle
bulge | Adherence | Keratinocyte
stem cells/
epidermal
neural crest
stem cells | Keratin 15, keratin 19,
β1 integrins, CD200,
PHLDA1, follistatin, frizzled
homolog 1, CD24lo, CD34lo,
CD71lo, and CD146lo | Keratinocytes, all major
neural crest lineages,
including neurons, Schwann
cells, myofibroblasts,
melanocytes, and bone/
cartilage cells | 11,12,14,15 |
Hair follicle
sheath | Floating
spheres | Dermal
sheath cells | Nestin, fibronectin, CD34,
and keratin 15(−) | Adipogenic and osteogenic
lineages | 16,22 |
Hair follicle
papillae | Floating
spheres | Skin-derived
precursor
cells | βIII-tubulin, p75NTR, NF-M;
CNPase, GFAP, and S100β | Adipogenic, osteogenic,
chondrogenic, and myogenic
lineages, neurons, glia, and
Schwann cells | 18–23 |
| Sweat gland | Adherence | Sweat gland
stroma-
derived
stem cells | α6 integrin and nestin | Adipogenic, chondrogenic,
and osteogenic lineages | 25,26 |
| Dermis | Perivascular | Adherence | Pericytes | CD146, NG2, CD31(−),
CD34(−), CD144(−), and
VWF(−) | Adipogenic, chondrogenic,
myogenic, and osteogenic
lineages | 10,51 |
Undefined
niches of
dermis | Adherence | Dermal stem
cells/dermis-
derived
stromal cells | CD13, CD29, CD44, CD49d,
CD71, CD73, CD90, CD105,
CD166, SSEA4, vimentin,
CD14(−), CD31(−), CD34(−),
CD45(−), CD106(−),
CD133(−), SSEA3(−), and
nestin(−) | Adipogenic, chondrogenic,
myogenic, and osteogenic
lineages | 27–34,50,52,53 |
Epidermis
Epidermal stem cells are found in both hair follicle bulge11,12 and interfollicular epidermis13,14. They are also viewed as keratinocyte stem cells because they generate cells that produce keratin11,14. Recent reports indicate that human epidermal stem cells are able to create all major neural crest derivatives containing neurons, Schwann cells, myofibroblasts, melanocytes, and bone/cartilage cells15,16. Despite the investigation of many stem cell markers, such as α6 integrin 5-bromo-2-deoxyuridine, β1 integrins, CD133, CD200, CD90, keratin 15, delta 1, and p6317, the molecular signature of epidermal stem cells remains undetermined.
Hair follicle and appendages
Hair follicles have long been considered an important niche for stem cells because of the versatility in regeneration of hair and epidermis and wound repair. For example, skin-derived precursors (SKPs) from both murine and human origins residing in the papillae of hair follicles18 can differentiate into neuron, glia, smooth muscle, and adipose cells19,20. As non-adherent cells, the SKPs are cultured as floating spheres with a neural crest origin21. Although lineage differentiation crosses both ectoderm and mesoderm18,20, their potential for osteogenesis has seldom been tested, although a cell subpopulation characterized from hair follicle dermal papilla and dermal sheath of both rats and humans has the capacity for adipogenesis, myogenesis, chondrogenesis, and osteogenesis22–24. In addition, since keratinocytes can be generated from the hair follicle bulge, the hair follicle is an important niche for epidermal stem cells11,12. These findings indicate that the hair follicle is one of the most important niches in skin with stem cells and progenitors generating mesenchymal lineages. Recent studies indicate that sweat glands, a skin appendage, are also characterized as a niche for stem cells which can be isolated and induced into three mesodermal lineages25,26.
Dermis
Dermis constitutes the majority of skin in both thickness and cell number. Dermal fibroblasts, the principal cells in dermis, have long been considered terminally differentiated cells and served as a negative control of mesenchymal stem cells (MSCs). When preserved in saline at 4°C for 6 days before digesting, non-hair follicle human dermis has been successfully proven to be an MSC source, indicative of a potential niche for stem cells27. This finding is supported by another report, in which clonal analysis of a single dermal fibroblast isolated from human foreskin exhibited tripotent, bipotent, and unipotent ability28, indicating multiple differentiation potential in dermal fibroblasts. Increasing evidence also demonstrates that these cells are positive for surface markers CD29, CD44, CD73, CD90, CD105, and CD166, indicating their MSC nature, and negative for CD14, CD31, CD34, CD45, and CD133, indicating non-hematopoietic lineage29–34.
Strategies for using skin cells for osteogenesis
Fibroblasts from rabbit skin were osteoinduced followed by seeding on porous titanium pylon; this construct exhibited enhanced osseointegrative properties compared with unseeded pylon in both in vitro and in vivo studies35. This study and others36 suggest the possibility of using skin fibroblasts for osteogenesis, although an early report showed the inhibition of rat skin fibroblasts on mineralization of bone marrow MSCs37. Unfortunately, owing to the low osteogenic potential of total skin fibroblasts with mixed cell populations, this kind of trial is far from successful. Therefore, it is critical to isolate skin cells with a preference for differentiation toward osteogenesis.
Genetic modification
Using modification of genes to increase the expression of specific osteogenesis-related genes, skin fibroblasts, acting as “protein secretors” without differentiating by themselves or having the paracrine/exosomal effects that are found in MSCs, were promoted for bone tissue engineering and regeneration38–41. These genes of interest include BMP-241–45, BMP-442, BMP-738,42, Runx2 (runt-related transcription factor 2)39,43,46,47, and LMP-3 (lim mineralization protein-3)40,48. In in vivo studies using skin fibroblasts, both ectopic osteogenesis and orthotopic bone regeneration are achieved through gene therapy42,44 from small animals like mice44, rats38,42,48, and rabbits41 to large animals like equines45. A study comparing different genes of interest for modification efficiency of skin fibroblasts determined that BMP-2 is more powerful than Runx243 and that the mineralization ability of Runx2-modified skin fibroblasts is scaffold-dependent39. Gene therapy is a promising method with a prominent effect; however, the safety of viral genetic modification needs further characterization49.
Cell sorting
Mixed populations isolated from total skin make cell therapy strategies for osteogenesis unsuccessful. Consequently, there are increasing efforts in sorting cells from skin to get target subpopulations. For example, type IV collagen-coated dishes have been used to attract CD29(+) human dermal stem cells via adherence, which exhibited higher osteogenic, adipogenic, and chondrogenic capacity compared with unsorted cells33. CD271(+) and CD146(+) cells isolated from human skin and CD73(−)CD105(+) cells isolated from mouse skin by immunosorting also showed elevated multi-differentiation potential50–52. Interestingly, subpopulations sorted by other markers from human skin, such as CD73, stage-specific embryonic antigen-4 (SSEA-4), and BmprIB, show relatively restricted differentiation potential. For instance, BmprIB(+) cells can generate only an osteogenic lineage50,53, indicating that these subpopulations can be applied as therapeutic cells for osteogenesis because of their established lineage preference. However, concern due to low harvest rate resulting from cell sorting still exists50,51,53.
Cell reprogramming
Characterized by unlimited proliferation and differentiation potential like embryonic stem cells54,55, induced pluripotent stem cells (iPSCs) can be used in numerous stem cell therapies. As skin fibroblasts are the most abundant and easily accessed cells, they are commonly chosen as the parent cells of iPSCs. It has been well characterized that iPSC-derived osteoblasts can form osteoid both in vitro and in vivo56–58. A recent study revealed that bone defect repair is also achieved by human iPSCs in a radial defect model of immune-deficient mice59. Furthermore, the involvement and mechanism of microRNAs in the regulation of mouse iPSCs during osteogenic differentiation have been preliminarily investigated60.
Conclusions and perspectives
In past decades, investigations using skin cells for osteogenesis have achieved significant progress. Many niches for stem cells in skin have been revealed and preliminarily characterized. Also, skin cells, enriched or not enriched, modified or not modified, are used for osteogenesis in vitro and in vivo and have achieved success in limb, cranial, mandibular, and rib bone defect regeneration (Figure 1). However, some key problems remain unsolved. For example, since the niche for stem cells in dermis is not completely characterized, the efficiency of enriching stem cells or progenitors from skin is still restricted. For cell modification strategies, like gene therapy and cell reprogramming, the efficacy might be readily apparent, but the safety needs more in-depth research.
Recent developments in epigenetic conversion may shed some light on cell reprogramming. Unlike in iPSCs, epigenetic conversion does not completely reverse cells to the pluripotent stem cell stage61–64. This approach may avoid undesired side effects such as teratoma, which often occurs in the application of iPSCs and embryonic stem cells. Epigenetic conversion has achieved progress in directing fibroblasts from human skin and mouse embryos into cardiomyocytes, neuronal cells, and insulin-secreting cells with a mature phenotype61,63,64. Although not much is known about converting skin fibroblasts into osteoblasts, there is a report of converting non-osteogenic cells into osteoblasts by epigenetic stimulation of BMP-2 expression65. By transient use of platelet-derived growth factor-AB and 5-azacytidine, mature bone and fat cells can also be converted into multipotent stem cells62. Thus, although there are no studies characterizing the cells converted for bone regeneration, the most common candidate for epigenetic conversion, skin cells, may play a significant role in this strategy.
Taken together, two of these strategies are promising. One strategy is the enrichment of stem cells and progenitors from different skin niches. By improving the current low-efficiency cell isolation, a mass of therapeutic cells can be gathered from skin for better bone tissue engineering and regeneration. The other strategy is based on the easy access and abundant amount of skin fibroblasts. Via modification of the cell, either through iPSCs or the recent concept of epigenetic conversion, a differentiation-specific cell population can be manipulated and gathered. In that case, therapeutic cells for osteogenesis can be harvested on a large scale, making both the autologous and allogeneic approaches possible.
Competing interests
The authors declare that they have no competing interests.
Grant information
This work was supported by research grants from the Musculoskeletal Transplant Foundation and the National Institutes of Health (1R03AR062763-01A1 and 1R01AR067747-01A1).
Acknowledgments
We thank Suzanne Danley for editing the manuscript.
Faculty Opinions recommendedReferences
- 1.
Kaplan FS, Craver R, MacEwen GD, et al.:
Progressive osseous heteroplasia: a distinct developmental disorder of heterotopic ossification. Two new case reports and follow-up of three previously reported cases.
J Bone Joint Surg Am.
1994; 76(3): 425–36. PubMed Abstract
| Publisher Full Text
- 2.
Adegbite NS, Xu M, Kaplan FS, et al.:
Diagnostic and mutational spectrum of progressive osseous heteroplasia (POH) and other forms of GNAS-based heterotopic ossification.
Am J Med Genet A.
2008; 146A(14): 1788–96. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 3.
Shore EM, Kaplan FS:
Inherited human diseases of heterotopic bone formation.
Nat Rev Rheumatol.
2010; 6(9): 518–27. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 4.
Myllylä RM, Haapasaari KM, Palatsi R, et al.:
Multiple miliary osteoma cutis is a distinct disease entity: four case reports and review of the literature.
Br J Dermatol.
2011; 164(3): 544–52. PubMed Abstract
| Publisher Full Text
- 5.
Wosczyna MN, Biswas AA, Cogswell CA, et al.:
Multipotent progenitors resident in the skeletal muscle interstitium exhibit robust BMP-dependent osteogenic activity and mediate heterotopic ossification.
J Bone Miner Res.
2012; 27(5): 1004–17. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 6.
Myllylä RM, Haapasaari K, Lehenkari P, et al.:
Bone morphogenetic proteins 4 and 2/7 induce osteogenic differentiation of mouse skin derived fibroblast and dermal papilla cells.
Cell Tissue Res.
2014; 355(2): 463–70. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 7.
Kamel RA, Ong JF, Eriksson E, et al.:
Tissue engineering of skin.
J Am Coll Surg.
2013; 217(3): 533–55. PubMed Abstract
| Publisher Full Text
- 8.
Chen Z, Pradhan S, Liu C, et al.:
Skin-derived precursors as a source of progenitors for cutaneous nerve regeneration.
Stem Cells.
2012; 30(10): 2261–70. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 9.
Vapniarsky N, Arzi B, Hu JC, et al.:
Concise Review: Human Dermis as an Autologous Source of Stem Cells for Tissue Engineering and Regenerative Medicine.
Stem Cells Transl Med.
2015; 4(10): 1187–98. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 10.
Crisan M, Yap S, Casteilla L, et al.:
A perivascular origin for mesenchymal stem cells in multiple human organs.
Cell Stem Cell.
2008; 3(3): 301–13. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 11.
Lyle S, Christofidou-Solomidou M, Liu Y, et al.:
The C8/144B monoclonal antibody recognizes cytokeratin 15 and defines the location of human hair follicle stem cells.
J Cell Sci.
1998; 111(Pt 21): 3179–88. PubMed Abstract
- 12.
Ohyama M, Terunuma A, Tock CL, et al.:
Characterization and isolation of stem cell-enriched human hair follicle bulge cells.
J Clin Invest.
2006; 116(1): 249–60. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 13.
Ghazizadeh S, Taichman LB:
Multiple classes of stem cells in cutaneous epithelium: a lineage analysis of adult mouse skin.
EMBO J.
2001; 20(6): 1215–22. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 14.
Ito M, Liu Y, Yang Z, et al.:
Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis.
Nat Med.
2005; 11(12): 1351–4. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 15.
Clewes O, Narytnyk A, Gillinder KR, et al.:
Human epidermal neural crest stem cells (hEPI-NCSC)--characterization and directed differentiation into osteocytes and melanocytes.
Stem Cell Rev.
2011; 7(4): 799–814. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 16.
Kanno H, Kubo A, Yoshizumi T, et al.:
Isolation of multipotent nestin-expressing stem cells derived from the epidermis of elderly humans and TAT-VHL peptide-mediated neuronal differentiation of these cells.
Int J Mol Sci.
2013; 14(5): 9604–17. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 17.
Ghadially R:
25 years of epidermal stem cell research.
J Invest Dermatol.
2012; 132(3 Pt 2): 797–810. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 18.
Fernandes KJ, McKenzie IA, Mill P, et al.:
A dermal niche for multipotent adult skin-derived precursor cells.
Nat Cell Biol.
2004; 6(11): 1082–93. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 19.
Toma JG, Akhavan M, Fernandes KJ, et al.:
Isolation of multipotent adult stem cells from the dermis of mammalian skin.
Nat Cell Biol.
2001; 3(9): 778–84. PubMed Abstract
| Publisher Full Text
- 20.
Toma JG, McKenzie IA, Bagli D, et al.:
Isolation and characterization of multipotent skin-derived precursors from human skin.
Stem Cells.
2005; 23(6): 727–37. PubMed Abstract
| Publisher Full Text
- 21.
Biernaskie JA, McKenzie IA, Toma JG, et al.:
Isolation of skin-derived precursors (SKPs) and differentiation and enrichment of their Schwann cell progeny.
Nat Protoc.
2006; 1(6): 2803–12. PubMed Abstract
| Publisher Full Text
- 22.
Jahoda CA, Whitehouse J, Reynolds AJ, et al.:
Hair follicle dermal cells differentiate into adipogenic and osteogenic lineages.
Exp Dermatol.
2003; 12(6): 849–59. PubMed Abstract
| Publisher Full Text
- 23.
Hoogduijn MJ, Gorjup E, Genever PG:
Comparative characterization of hair follicle dermal stem cells and bone marrow mesenchymal stem cells.
Stem Cells Dev.
2006; 15(1): 49–60. PubMed Abstract
| Publisher Full Text
- 24.
Bajpai VK, Mistriotis P, Andreadis ST:
Clonal multipotency and effect of long-term in vitro expansion on differentiation potential of human hair follicle derived mesenchymal stem cells.
Stem Cell Res.
2012; 8(1): 74–84. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 25.
Lu CP, Polak L, Rocha AS, et al.:
Identification of stem cell populations in sweat glands and ducts reveals roles in homeostasis and wound repair.
Cell.
2012; 150(1): 136–50. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 26.
Nagel S, Rohr F, Weber C, et al.:
Multipotent nestin-positive stem cells reside in the stroma of human eccrine and apocrine sweat glands and can be propagated robustly in vitro.
PLoS One.
2013; 8(10): e78365. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 27.
Bartsch G, Yoo JJ, De Coppi P, et al.:
Propagation, expansion, and multilineage differentiation of human somatic stem cells from dermal progenitors.
Stem Cells Dev.
2005; 14(3): 337–48. PubMed Abstract
| Publisher Full Text
- 28.
Chen FG, Zhang WJ, Bi D, et al.:
Clonal analysis of nestin- vimentin+ multipotent fibroblasts isolated from human dermis.
J Cell Sci.
2007; 120(Pt 16): 2875–83. PubMed Abstract
| Publisher Full Text
- 29.
Jääger K, Islam S, Zajac P, et al.:
RNA-seq analysis reveals different dynamics of differentiation of human dermis- and adipose-derived stromal stem cells.
PLoS One.
2012; 7(6): e38833. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 30.
Crigler L, Kazhanie A, Yoon TJ, et al.:
Isolation of a mesenchymal cell population from murine dermis that contains progenitors of multiple cell lineages.
FASEB J.
2007; 21(9): 2050–63. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 31.
Lorenz K, Sicker M, Schmelzer E, et al.:
Multilineage differentiation potential of human dermal skin-derived fibroblasts.
Exp Dermatol.
2008; 17(11): 925–32. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 32.
Huang HI, Chen SK, Ling QD, et al.:
Multilineage differentiation potential of fibroblast-like stromal cells derived from human skin.
Tissue Eng Part A.
2010; 16(5): 1491–501. PubMed Abstract
| Publisher Full Text
- 33.
Shim JH, Kang HH, Lee TR, et al.:
Enrichment and characterization of human dermal stem/progenitor cells using collagen type IV.
J Dermatol Sci.
2012; 67(3): 202–5. PubMed Abstract
| Publisher Full Text
- 34.
Hill RP, Gledhill K, Gardner A, et al.:
Generation and characterization of multipotent stem cells from established dermal cultures.
PLoS One.
2012; 7(11): e50742. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 35.
Shevtsov MA, Galibin OV, Yudintceva NM, et al.:
Two-stage implantation of the skin- and bone-integrated pylon seeded with autologous fibroblasts induced into osteoblast differentiation for direct skeletal attachment of limb prostheses.
J Biomed Mater Res A.
2014; 102(9): 3033–48. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 36.
Hee CK, Jonikas MA, Nicoll SB:
Influence of three-dimensional scaffold on the expression of osteogenic differentiation markers by human dermal fibroblasts.
Biomaterials.
2006; 27(6): 875–84. PubMed Abstract
| Publisher Full Text
- 37.
Ogiso B, Hughes FJ, Melcher AH, et al.:
Fibroblasts inhibit mineralised bone nodule formation by rat bone marrow stromal cells in vitro.
J Cell Physiol.
1991; 146(3): 442–50. PubMed Abstract
| Publisher Full Text
- 38.
Krebsbach PH, Gu K, Franceschi RT, et al.:
Gene therapy-directed osteogenesis: BMP-7-transduced human fibroblasts form bone in vivo.
Hum Gene Ther.
2000; 11(8): 1201–10. PubMed Abstract
| Publisher Full Text
- 39.
Phillips JE, Hutmacher DW, Guldberg RE, et al.:
Mineralization capacity of Runx2/Cbfa1-genetically engineered fibroblasts is scaffold dependent.
Biomaterials.
2006; 27(32): 5535–45. PubMed Abstract
| Publisher Full Text
- 40.
Lattanzi W, Parrilla C, Fetoni A, et al.:
Ex vivo-transduced autologous skin fibroblasts expressing human Lim mineralization protein-3 efficiently form new bone in animal models.
Gene Ther.
2008; 15(19): 1330–43. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 41.
Ishihara A, Weisbrode SE, Bertone AL:
Autologous implantation of BMP2-expressing dermal fibroblasts to improve bone mineral density and architecture in rabbit long bones.
J Orthop Res.
2015; 33(10): 1455–65. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 42.
Rutherford RB, Moalli M, Franceschi RT, et al.:
Bone morphogenetic protein-transduced human fibroblasts convert to osteoblasts and form bone in vivo.
Tissue Eng.
2002; 8(3): 441–52. PubMed Abstract
| Publisher Full Text
- 43.
Hirata K, Tsukazaki T, Kadowaki A, et al.:
Transplantation of skin fibroblasts expressing BMP-2 promotes bone repair more effectively than those expressing Runx2.
Bone.
2003; 32(5): 502–12. PubMed Abstract
| Publisher Full Text
- 44.
Wang R, Zou Y, Yuan Z, et al.:
Autografts and xenografts of skin fibroblasts delivering BMP-2 effectively promote orthotopic and ectopic osteogenesis.
Anat Rec (Hoboken).
2009; 292(6): 777–86. PubMed Abstract
| Publisher Full Text
- 45.
Ishihara A, Zekas LJ, Weisbrode SE, et al.:
Comparative efficacy of dermal fibroblast-mediated and direct adenoviral bone morphogenetic protein-2 gene therapy for bone regeneration in an equine rib model.
Gene Ther.
2010; 17(6): 733–44. PubMed Abstract
| Publisher Full Text
- 46.
Phillips JE, Guldberg RE, Garcia AJ:
Dermal fibroblasts genetically modified to express Runx2/Cbfa1 as a mineralizing cell source for bone tissue engineering.
Tissue Eng.
2007; 13(8): 2029–40. PubMed Abstract
| Publisher Full Text
- 47.
Phillips JE, García AJ:
Retroviral-mediated gene therapy for the differentiation of primary cells into a mineralizing osteoblastic phenotype.
Methods Mol Biol.
2008; 433: 333–54. PubMed Abstract
| Publisher Full Text
- 48.
Parrilla C, Lattanzi W, Rita Fetoni A, et al.:
Ex vivo gene therapy using autologous dermal fibroblasts expressing hLMP3 for rat mandibular bone regeneration.
Head Neck.
2010; 32(3): 310–8. PubMed Abstract
| Publisher Full Text
- 49.
Zwaka TP:
Use of genetically modified stem cells in experimental gene therapies. In: Templeton NS (Eds.). Gene and Cell Therapy: Therapeutic Mechanisms and Strategies, third edition. Taylor & Francis/CRC Press, Boca Raton, FL. 2009; 731–6. Publisher Full Text
- 50.
Vaculik C, Schuster C, Bauer W, et al.:
Human dermis harbors distinct mesenchymal stromal cell subsets.
J Invest Dermatol.
2012; 132(3 Pt 2): 563–74. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 51.
Ruetze M, Knauer T, Gallinat S, et al.:
A novel niche for skin derived precursors in non-follicular skin.
J Dermatol Sci.
2013; 69(2): 132–9. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 52.
Lee SB, Shim S, Kim MJ, et al.:
Identification of a distinct subpopulation of fibroblasts from murine dermis: CD73- CD105+ as potential marker of dermal fibroblasts subset with multipotency.
Cell Biol Int.
2016; 40(9): 1008–16. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 53.
Wang T, He J, Zhang Y, et al.:
A Selective Cell Population From Dermis Strengthens Bone Regeneration.
Stem Cells Transl Med.
2016; pii: sctm.2015-0426. PubMed Abstract
| Publisher Full Text
- 54.
Takahashi K, Yamanaka S:
Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors.
Cell.
2006; 126(4): 663–76. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 55.
Takahashi K, Tanabe K, Ohnuki M, et al.:
Induction of pluripotent stem cells from adult human fibroblasts by defined factors.
Cell.
2007; 131(5): 861–72. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 56.
Bilousova G, Jun du H, King KB, et al.:
Osteoblasts derived from induced pluripotent stem cells form calcified structures in scaffolds both in vitro and in vivo.
Stem Cells.
2011; 29(2): 206–16. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 57.
Jin GZ, Kim TH, Kim JH, et al.:
Bone tissue engineering of induced pluripotent stem cells cultured with macrochanneled polymer scaffold.
J Biomed Mater Res A.
2013; 101(5): 1283–91. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 58.
Lou X:
Induced Pluripotent Stem Cells as a new Strategy for Osteogenesis and Bone Regeneration.
Stem Cell Rev.
2015; 11(4): 645–51. PubMed Abstract
| Publisher Full Text
- 59.
Sheyn D, Ben-David S, Shapiro G, et al.:
Human Induced Pluripotent Stem Cells Differentiate Into Functional Mesenchymal Stem Cells and Repair Bone Defects.
Stem Cells Transl Med.
2016; 5(11): 1447–60. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 60.
Okamoto H, Matsumi Y, Hoshikawa Y, et al.:
Involvement of microRNAs in regulation of osteoblastic differentiation in mouse induced pluripotent stem cells.
PLoS One.
2012; 7(8): e43800. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 61.
Brevini TA, Pennarossa G, Maffei S, et al.:
Epigenetic Conversion as a Safe and Simple Method to Obtain Insulin-secreting Cells from Adult Skin Fibroblasts.
J Vis Exp.
2016; (109): e53880. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 62.
Chandrakanthan V, Yeola A, Kwan JC, et al.:
PDGF-AB and 5-Azacytidine induce conversion of somatic cells into tissue-regenerative multipotent stem cells.
Proc Natl Acad Sci U S A.
2016; 113(16). E2306–15. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 63.
Cao N, Huang Y, Zheng J, et al.:
Conversion of human fibroblasts into functional cardiomyocytes by small molecules.
Science.
2016; 352(6290): 1216–20. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
- 64.
Black JB, Adler AF, Wang HG, et al.:
Targeted Epigenetic Remodeling of Endogenous Loci by CRISPR/Cas9-Based Transcriptional Activators Directly Converts Fibroblasts to Neuronal Cells.
Cell Stem Cell.
2016; 19(3): 406–14. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
- 65.
Cho YD, Yoon WJ, Kim WJ, et al.:
Epigenetic modifications and canonical wingless/int-1 class (WNT) signaling enable trans-differentiation of nonosteogenic cells into osteoblasts.
J Biol Chem.
2014; 289(29): 20120–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
Comments on this article Comments (0)