Keywords
aminoglycoside, antibiotics, Resistance
aminoglycoside, antibiotics, Resistance
The discovery of streptomycin from Streptomyces griseus by Waksman in 1943 as a pharmaceutical legend against tuberculosis inspired massive mining of similar compounds in other bacterial species, leading to the discovery of neomycins, kanamycins, gentamicins, and a variety of other aminoglycoside (AG) antibiotics1–4. Structurally, AGs possess a featured aminocyclitol core, usually as streptamine, streptidine, or 2-deoxystreptamine (2-DOS), which is decorated by one or more amino-sugars at the C4, C5, and C6 positions. These positively charged amino groups enable AGs to interact with negatively charged molecules, especially rRNAs of bacterial 30S and 50S ribosomal subunits, thereby inhibiting protein synthesis and causing cell death5,6. Such a bactericidal property of AGs makes them broad-spectrum antibiotic agents that are effective against both Gram-negative and Gram-positive bacteria7. However, as has been observed for most antibiotics, heavy usage of AGs resulted in the rapid spread of AG-resistant pathogens, which has greatly limited their clinical practice8. The most common mechanism for bacterial resistance to AGs is inactivation of AGs by structural modifications9. Thus far, over 100 AG-modifying enzymes (AMEs), including AG N-acetyltransferases (AACs), AG O-nucleotidyltransferases (ANTs), and AG O-phosphotransferases (APHs), have been found in a broad range of both Gram-positive and -negative organisms10. Also, AG resistance can be achieved by efflux pumps and, as has been revealed recently, by 16S rRNA methyltransferase-catalyzed modifications11. The bacterial resistance issues have been partly addressed by chemical modification of natural AGs to improve their pharmacological properties, as has been exemplified by the so-called second generation of AG drugs (the semisynthetic dibekacin, amikacin, netilmicin, and isepamicin) during the 1970s12.
Whilst chemical engineering is an effective way to generate novel AGs, rational biosynthetic engineering of new antibiotics represents an attractive alternative, and the past decade has witnessed a massive upsurge of studies in AG biosynthesis13,14. Among the largest and best-studied subgroup of AG natural products are the 2-DOS-containing AGs14. It has been well established that 2-DOS is produced from D-glucose-6-phosphate by a set of enzymes, including 2-deoxy-scyllo-inosose synthase, the dual-functional L-glutamine:2-DOI aminotransferase, and 2-deoxy-scyllo-inosamine dehydrogenase (the genes encoding these enzymes are conserved in all of the biosynthetic gene clusters identified so far for 2-DOS-containing AGs and are designated as “C, S, E”-type genes, respectively)7,15,16. Decoration by various sugar moieties at the C4, C5, and C6 positions of 2-DOS proceeds via different patterns, yielding 4,5-disubstituted AGs (e.g. butirosin and neomycin), 4,6-disubstituted AGs (e.g. kanamycin, tobramycin, and gentamicin), or 4- and 5-monosubstituted AGs (e.g. apramycin and hygromycin). Readers are referred to many excellent reviews discussing the biosynthesis and engineering of AG antibiotics13–15,17,18.
A common observation in AG biosynthesis is that one producer strain usually produces many AG congeners simultaneously. Recent studies have shown that the presence of multiple AG congeners is, in many cases, not merely because of inefficient downstream modifications but mainly the result of parallel biosynthetic pathways19–21. This review discusses the recent advances in the biosynthesis of three representative AGs—kanamycin, gentamicin, and apramycin—which all involve parallel pathways. We attempt to highlight nature’s ingenuity in accessing diverse AGs through a limited set of genes and the potential for the generation of novel AG antibiotics by pathway engineering efforts.
The kanamycin class consists of several members, most of which were isolated from Streptomyces kanamyceticus1. Structurally, these congeners contain the same kanosamine moiety (3-amino-3-deoxy-D-glucose) at the C6 position of 2-DOS but possess different amino-sugars at the C4 position of 2-DOS (i.e. 6-amino-6-deoxy-D-glucose in kanamycin A, neosamine in kanamycin B, and D-glucosamine in kanamycin C). Several second-generation AG drugs, such as amikacin, dibekacin, and arbekacin, are produced by semi-synthetic modification of the kanamycin core skeleton9, stimulating interest in deciphering kanamycin biosynthesis and developing novel kanamycin by bioengineering.
The parallel pathways of AG biosynthesis were firstly revealed for kanamycin21. For a long time, it had been postulated that kanamycin is assembled via a linear pathway7. However, the recent study by Yoon, Sohng, and co-workers indicated that kanamycin A and B are actually produced by two parallel pathways (Figure 1)21. KanF (also known as KanM1), the glycosyltransferase in the kan gene cluster, was shown to be able to transfer both NDP-glucose and NDP-N-acetylglucosamine to 2-DOS to generate 2'-hydroxyparomamine and paromamine, respectively, thereby directing the metabolic flux of 2-DOS into two branches. The two resulting pseudodisaccharides are then aminated at the C6' position by the sequential action of a dehydrogenase KanQ (also known as KanI) and an aminotransferase KanB (also known as KacL), yielding 2'-hydroxyneamine and neamine, respectively. KanE (also known as KanM2), the second glycosyltransferase in the kan gene cluster, exhibits a very relaxed substrate promiscuity, accepting four different pseudodisaccharides (i.e. paromamine, neamine, 2'-hydroxyneamine, and 2'-hydroxyparomamine) as glycosyl acceptors21,22, resulting in the production of kanamycin A and B via the two independent pathways (Figure 1). The complex network of kanamycin biosynthesis is completed by the action of the α-ketoglutarate-dependent dioxygenase KanJ and the NADH-dependent reductase KanK, which together convert kanamycin B to kanamycin A23. The remarkable substrate promiscuity of kanamycin biosynthetic enzymes suggests the great potential in AG combinatorial biosynthesis. Indeed, integration of the S-4-amino-2-hydroxybutyric acid (AHBA) biosynthesis genes into the kanamycin pathway successfully led to the production of a semisynthetic AG, amikacin, and a new AG analogue, 1-N-AHBA-kanamycin X, both of which show improved antibacterial activity compared with kanamycin A21.
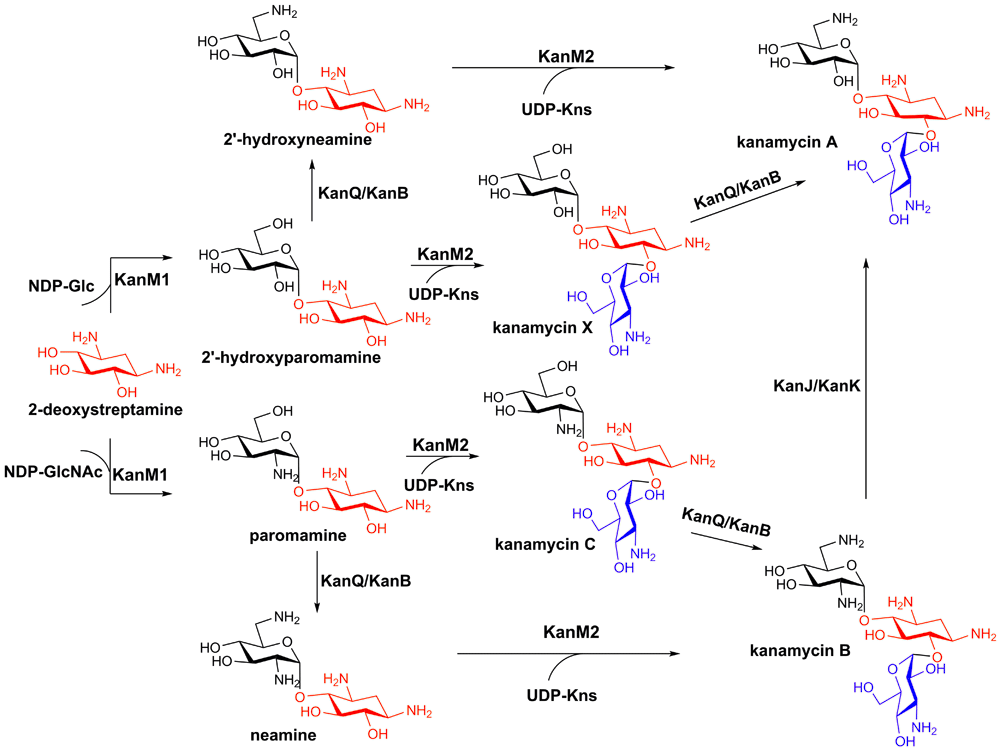
The 2-deoxystreptamine (2-DOS) moiety is highlighted in red, and the amino-sugar kanosamine (Kns) is highlighted in blue. GlcNAc, N-Acetylglucosamine.
Gentamicin was isolated from Micromonospora echinospora as a mixture of five C-series components consisting of C1, C2, C1a, C2a, C2b4. These compounds have received much attention in recent years as a broad-spectrum antibiotic for the treatment of various infections as well as a promising drug lead for the treatment of several inherited diseases and a sensitizing agent for lung cancer cells24–26. Gentamicin contains the core 2-DOS moiety with the amino-sugars purpurosamine and garosamine attached at positions C4 and C6, respectively. The biosynthetic pathway for gentamicin was firstly proposed several decades ago, which is mostly corrected as shown by recent studies19. Heterologous expression of a subset of the gentamicin gene cluster in Streptomyces venezuelae indicated that gentamicin A2 is the first pseudotrisaccharide precursor of the gentamicin C complex27. Gentamicin A2 is then converted to gentamicin X2 by a series of enzymes, including the dehydrogenase GenD2 and the pyridoxal phosphate (PLP)-dependent aminotransferase GenS2, the S-adenosylmethionine (SAM)-dependent N-methyltransferase GenN, and the radical SAM methyltransferase GenD1. GenD2 and GenS are responsible for the amination of the C3'' position of gentamicin A2, and the resulting amino group is methylated by GenN to afford a C3'' methylamino group28. GenD1 belongs to the class B radical SAM methyltransferases29,30, which utilizes a cobalamin cofactor and installs a methyl group on the C4'' position31,32. Gentamicin X2 is then methylated at the C6' position by GenK, which, like GenD1, also belongs to the class B radical SAM methyltransferase, leading to the production of gentamicin G41833.
Gentamicin G418 and X2 then serve as the starting substrates of the two parallel pathways and are first aminated at C6' by the sequential action of the dehydrogenase GenQ and the aminotransferase GenB1, resulting in gentamicin JI-20A and JI-20B, respectively19. Gentamicin JI-20A is then dideoxygenated by a set of uncharacterized enzymes (likely including a phosphotransferase, GenP) to produce gentamicin C1a, whose C6' amino group is then methylated by an as-yet-unknown methyltransferase to produce gentamicin C2b. Gentamicin JI-20B is also dideoxygenated and methylated to afford gentamicin C1. However, because of the presence of several gentamicin congeners, including gentamicin JI-20Ba, gentamicin C2a, and gentamicin C2, and the involvement of the epimerase GenB2 that catalyzes the stereochemical interconversion of the C6' amino group, the exact reaction order and timing in gentamicin C1 production is currently unclear. A possible scenario is shown in Figure 2, in which GenB2 is responsible for the third parallel pathway and is not essential for gentamicin C biosynthesis; this hypothesis awaits testing in future studies. Unlike the parallel pathways in kanamycin biosynthesis, in which the final product of one pathway (kanamycin B) can be converted to the final product of the second one (kanamycin A), gentamicin C2b is the ultimate product and cannot be methylated by GenK to produce gentamicin C1. Clearly, many aspects in gentamicin biosynthesis remain elusive and warrant investigations.
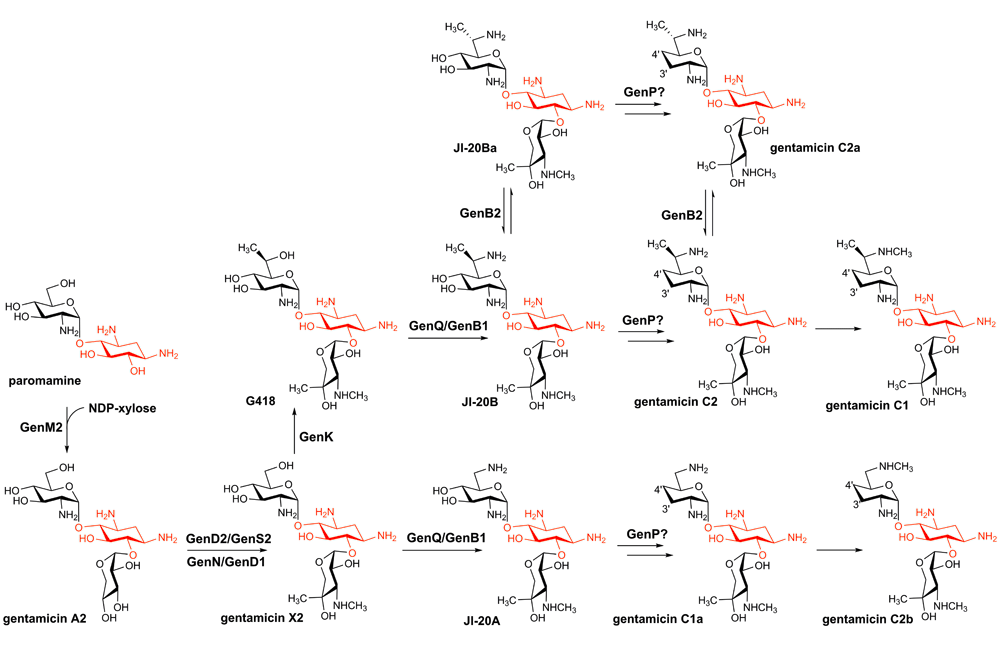
The 2-deoxystreptamine (2-DOS) moiety is highlighted in red.
Apramycin is a highly modified AG containing a unique bicyclic octose moiety, which is deoxygenated at the C3 position. Although currently only licensed for veterinary practice use, apramycin represents the only known AG that is active against carbapenem-resistant Enterobacteriaceae isolates harboring the 16S rRNA MTases gene34. In addition, apramycin is able to mimic the features of an incompatible plasmid, thereby re-sensitizing bacteria to conventional antibiotic treatments by causing plasmid loss in vivo35. More importantly, in contrast to common AGs that possess substantial ototoxicity and could cause irreversible hearing loss36, recent studies have shown that apramycin has less ototoxicity, further promising its pharmaceutical applications37.
The biosynthetic gene cluster of apramycin was reported more than a decade ago38, but the biosynthetic timing of apramycin, particularly with regard to the C3 deoxygenation step, has remained obscure until very recently. In contrast to the previous proposal in which the C3 deoxygenation occurs as the penultimate step, biochemical studies have shown that the C3 deoxygenation is catalyzed by the radical SAM diol dehydratase AprD4 and its NADPH-dependent reductase partner AprD320,39,40. This C3 deoxygenation reaction does not occur on oxyapramycin but on the pseudodisaccharide substrate paromamine, demonstrating the two parallel biosynthetic pathways, which affords oxyapramycin and apramycin, respectively (Figure 3)20. Consistent with this observation, biochemical analysis has shown that AprQ, a FAD-dependent dehydrogenase, catalyzes the C6' dehydrogenation of both paromamine and lividamine, with the latter as the preferred substrate20. Remarkably, AprQ is also able to act on the tripseudodisaccharide substrate gentamicin X2 and G418 and, in the latter case, produce a novel gentamicin analogue with a C6' carboxylate20. A similar observation was also made on AprD4 and AprD3, which are able to act on tripseudodisaccharide substrates, such as kanamycin C and kanamycin B40. The remarkable substrate promiscuity of apramycin biosynthetic enzymes demonstrates the great potential to produce novel AGs by pathway engineering efforts. It is also noteworthy that although the radical-mediated dehydration catalyzed by cobalamin-dependent diol dehydratase has been well studied, AprD4 represents the only known dehydratase in the radical SAM superfamily, and its catalytic mechanism remains to be determined.
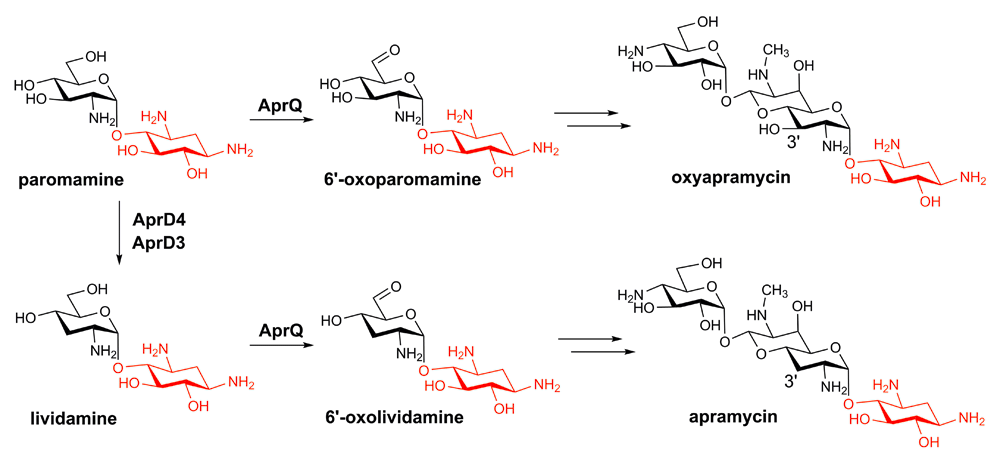
The 2-deoxystreptamine (2-DOS) moiety is highlighted in red.
In 2015, Drs Youyou Tu, William Campbell, and Satoshi Ōmura jointly received the Nobel Prize in Physiology or Medicine for their seminal discoveries of the natural products avermectin and artemisinin, partly renewing interest in natural drug discovery and development. Despite their long history of clinical and veterinary usage, AGs still represent an important class of antibiotic for the treatment of severe infections, particularly for Gram-negative bacterial infections. Because of the recent global spread of AG-resistant pathogens and the inherent drawbacks of AGs such as ototoxicity and nephrotoxicity, the discovery and development of novel AGs is in urgent need41,42. Mining of novel AGs from bacterial genomes is a promising way because of the rapid development of sequencing technology and the expansion of sequenced genomes that are publically available (https://gold.jgi.doe.gov/)43. Pathway engineering serves as another attractive strategy to generate novel and more robust AGs18. The parallel biosynthetic pathways of AGs discussed above reflects nature’s wisdom in structural diversity-oriented biosynthesis, highlighting the potential to access novel and more robust AGs by combinatorial biosynthesis while taking advantage of the substrate promiscuity of AG biosynthetic enzymes. Plazomicin is a new semisynthetic AG designed to evade all clinically relevant AMEs and is currently under phase III clinical trials9,44. It contains a gentamicin-related skeleton with a hydroxy-aminobutyric acid substituent and a hydroxyethyl substituent at its N-1' and N-6' positions, respectively45. The promise of plazomicin to be approved as a third-generation AG drug and its structural similarity with gentamicin will undoubtedly spark increased interest in engineering AG pathways46. Given that many aspects of AG biosynthesis remain unclear, further functional and mechanistic investigation of AG biosynthetic enzymes could be rewarding, which very likely entails novel and interesting biochemistries. With the expansion of our knowledge in AG biosynthesis and the availability of more and more synthetic biological tools, the resurgence of this old class of antibiotics is possible in the foreseeable future.
AG, aminoglycoside; AHBA, S-4-amino-2-hydroxybutyric acid; AME, aminoglycoside-modifying enzyme; 2-DOS, 2-deoxystreptamine; SAM, S-adenosylmethionine.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)