Keywords
retina, photoreceptor, processing, interaction
retina, photoreceptor, processing, interaction
When one of us was a graduate student in the early ’70s, Edwin Land, the inventor of the Polaroid camera, came to Harvard to give a seminar on his “retinex” theory of color vision1,2. Though a successful entrepreneur and businessman, Land retained an immense curiosity about the eye and especially about color. His retinex theory attempted to account for what most scientists would call color constancy—the remarkable ability of the visual system to maintain the perceived color of an object in quite different conditions of illumination. Apples continue to be red or green at dawn and at noon, even though the light reflected from the apples at different times of day can be quite different in wavelength composition.
After the seminar, Gordon told Land about his experiments on interactions between rod and cone signals in the mudpuppy retina3, and Land invited him to the research laboratory of the Polaroid Corporation to meet Land’s associate John McCann, who was working on rod contributions to color vision. Although we are accustomed to think of rods as being responsible for dim-light vision and cones for color vision, McCann had devised a psychophysical demonstration with light stimulating only the rods and long-wavelength cones. He showed with little doubt that rods could contribute to the perception of hue4,5. His results and those of other psychophysicists (see, for example,6,7) have provided clear evidence of a contribution of rods to color.
In addition to these experiments, a large body of psychophysical literature indicates that signals derived from rods and cones can interact at many levels in the retina (see 8). Rod signals can influence cone spatial acuity and temporal sensitivity in sometimes quite surprising ways. Because rod signals are slower than cone signals, the signals from the two kinds of photoreceptors to flickering illumination can arrive at downstream targets out of phase with one another so that the flickering light stimulating both kinds of photoreceptors can seem to be steady9. Even when the rods are not stimulated, they can decrease cone sensitivity to flicker during dark adaptation10, and cones can depress rod sensitivity and move rod saturation to lower light intensities11. These interactions probably occur somewhere in the retina, but nothing is known about the details of their mechanisms.
Renewed interest in interactions between rods and cones has been stimulated by several recent findings. Single-cell RNA expression profiling is being used in many parts of the nervous system to identify and distinguish different cell types from the profiles of the RNAs they express (see, for example,12,13). This method has also been used to good advantage on the retina, where it has now provided a comprehensive classification of all types of mouse bipolar cells14. Careful examination of contacts of these cells with another technique—serial-section electron microscopy followed by computer-guided reconstruction of cell morphology15—has given us a more complete understanding of rod and cone pathways through the mammalian retina and shown that they intermingle more than previously thought16. Physiological studies have also provided new perspectives (for example,17). Joesch and Meister18 have shown, for example, that mice have a specific kind of ganglion cell (called the J-RGC or JAMB; see 19) with a color-opponent receptive field (see also 20). These cells resemble ganglion cells thought to mediate color vision in many vertebrates, but the center OFF response of the J-RGC can come specifically from ultraviolet-sensitive cones, and the ON surround from rods. The center and surround are mutually antagonistic, suggesting that the mouse retina contains specific microcircuits to provide color information resulting from interactions between rod and cone signals.
Another even more surprising finding is the claim of Tikidji-Hamburyan and colleagues21 that rods can continue to function at much higher luminance than previously supposed. These investigators recorded from the retinas (and the central nervous system) of mice genetically engineered to lack cone function and showed that responses could be recorded from photoreceptors and ganglion cells even in bright, bleaching light. These findings challenge earlier psychophysical measurements in humans (22; see 23) and mice24 as well as electrical recordings from single mammalian photoreceptors (for example,25–27) and from other retinal neurons18,28, all of which seem to show that rods saturate and become essentially non-functional in relatively dim background light to allow the cones with their kinetically faster responses to dominate perception.
To put this research into perspective and stimulate new avenues of research, we will endeavor to describe our present understanding of rod and cone interactions in the retina, emphasizing more recent findings from the mouse and primate. There are several demonstrated pathways for rod and cone signals to be communicated through the retina (Figure 1), and each of these pathways provides opportunities for the two signals to influence one another.
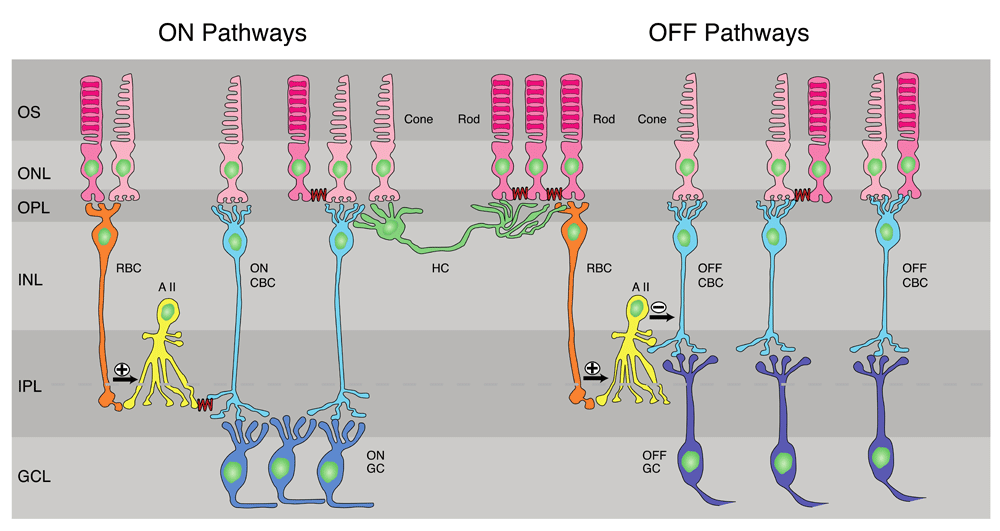
(Left) ON pathways. Rod bipolar cell (RBC) receives input mostly from rods but also from cones and makes excitatory glutamatergic synapses onto AII amacrine cells, which in turn make gap junctions (squiggly lines) onto ON cone bipolar cells (ON CBCs). The ON CBCs then synapse onto ON-type ganglion cells (ON GCs). Rods also make gap junctions onto other rods and onto cones (squiggly lines), and the cones then carry rod signals to ON CBCs. (Center) Mouse horizontal cell (HC), whose dendrites contact exclusively cones and whose axon terminal contacts exclusively rods. Both cell body and axon can receive both rod and cone signals indirectly by passive spread through gap junctions between receptors. (Right) OFF pathways. RBCs make excitatory synapses onto AII amacrine cells, which make inhibitory glycinergic synapses onto OFF cone bipolar cells (OFF CBCs). These in turn synapse onto OFF ganglion cells (OFF GCs). Rods make gap-junctional contacts onto cones, which carry rod signals through OFF CBCs to OFF GCs. Finally, some OFF bipolar cells receive input from both rods and cones (far right). Dashed line through inner plexiform layer (IPL) indicates sublaminae; upper layer (sublamina a) contains terminations of OFF bipolar cells and dendrites of OFF GCs, and lower (sublamina b) contains ON bipolar cell terminals and ON GC dendrites42. GCL, ganglion-cell layer; INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segments of photoreceptors. After 43,44.
A first stage of interaction between rods and cones occurs within the photoreceptor layer itself. Rods are electrically coupled to cones in many vertebrate species29, including teleost fish30, amphibians31, mice32,33, and primates34,35. Although the coupling is known to require connexin 3636,37, and connexin 36 immunolabeling can be readily identified at gap junctions between rods and cones on the cone side of the junction33,38,39, the identity of the rod connexin has long been uncertain. We think it quite likely, however, that the rod protein in the mouse is also some form of connexin 36. The evidence is, first, that connexin 36 expression has been shown to be widespread within the photoreceptor layer, and there is so much labeling that it is unlikely to be confined only within the cones36. In addition, Jin and colleagues40 have recently shown that coupling is essentially abolished when connexin 36 gene expression is deleted specifically within the rods. However, it remains possible that two different isoforms of connexin 36 are expressed in the two kinds of photoreceptors.
The preferred direction of current flow will be from rods to cones because of the much greater number of rods in most mammalian retinas41. Therefore, the extent of coupling between rods and cones will set the magnitude of rod signals seen within the cone pedicle. Rod–cone coupling has some interesting features. Coupling of rods and cones in goldfish retina is under circadian control so that it is strong in darkness at night but much weaker in brighter illumination during the day30. The extent of coupling is regulated by dopamine and D2 receptors, such that increases in dopamine and D2-receptor stimulation during the day decrease coupling. In primates, on the other hand, neither background light nor dopamine has been reported to influence the extent of coupling35.
Asteriti and colleagues32 have recorded from photoreceptors with perforated patch in mouse retina and observed that the amplitude of the rod voltage signal recorded in cones gradually increases over many minutes during the duration of the recording. The mechanism of this potential modulation is unclear. Our graduate student Norianne Ingram has recorded rod input in mouse cones with whole-cell patch recording45 and has seen no evidence for time dependence of the rod input. Clearly, much more remains to be learned about the physiology of rod–cone coupling and its control by light intensity and circadian rhythm.
Rods and cones make synapses in the outer plexiform layer with two kinds of cells (Figure 1): bipolar cells, whose axons transmit information to the next layer of retina called the inner plexiform layer, and horizontal cells, whose lateral processes interconnect photoreceptors and bipolar cells. Vertebrates have two kinds of bipolar cells (see 46): ON-type depolarizing to light in the center and hyperpolarizing to surrounding illumination and OFF-type hyperpolarizing to central illumination and depolarizing to the surround47,48. In teleost fish49 and other lower vertebrates (for example,3,50), rods and cones synapse together onto the same ON and OFF bipolar cells, although some bipolar cells receive much more rod input than others.
In mammals, rod signals are conveyed down to the inner plexiform layer by only a few specific kinds of bipolar cells (see Figure 1 and 43). There is a single so-called “rod” bipolar cell, which is exclusively ON and which uses metabotropic mGluR6 synaptic receptors51. Although these cells receive almost entirely rod input, we now know that they can also make a small number of contacts with cones16,17. These rod bipolar cells form the primary pathway of rod transit through the retina in dim light52–54 but can also continue to signal contrast modulation at ambient intensities above the cone threshold55. It is currently unknown whether they can also respond in even brighter, bleaching light, when rods would be hyperpolarized well below their dark resting-membrane potential.
In addition to the rod bipolar, there are a further 13 or 14 kinds of bipolar cell in mouse14, which can be either ON-type or OFF-type and use either mGluR6 glutamate receptors or ionotropic kainate/AMPA glutamate receptors. Most of these bipolar cells receive direct input exclusively from cones, but at least three types of OFF bipolar cell are also contacted by rods16,56–58, in mouse as apparently also in primates59. At present, there is little evidence for direct synaptic input of rods to ON bipolar cells other than the “rod” bipolar (but see 17). Each of the different bipolar cell types has a distinguishing response profile and different spatial and temporal characteristics60. These differences are probably the combined result of differences in their populations of synaptic receptors, voltage-gated ion channels, receptor modulation, and inhibition from horizontal cells as well as from amacrine cells in the inner plexiform layer (see 46,60,61).
Horizontal cells in lower vertebrates receive input from cones or from both rods and cones and can have light responses of two kinds. Some cells (called C-type) have mutually antagonistic input from two spectral classes of cones or from rods and cones, but most cells are L-type and receive exclusively hyperpolarizing input (see, for example,3,62,63). Mammals seem not to have C-type horizontal cells but can have two morphologically different kinds of L-type horizontal cells: an A-type, which lacks an axon and contacts only cones, and a B-type, whose dendritic terminals contact only cones and whose often large axon terminal contacts only rods64,65. The mouse retina has only the B-type (Figure 1; see 66).
Early recordings from mammalian horizontal cells nearly 50 years ago revealed that the same cell could receive both rod and cone input67. Similar recordings from mice have also demonstrated that both the cell body and axon terminal of the B-type cell can receive input from rods and cones68. Because the axon is slender and does not conduct action potentials, it is usually assumed that signals from the two kinds of photoreceptors cannot be conducted by the axon between the cell body and axon terminal69 but must travel via gap junctions between the rod and cone photoreceptors before the photoreceptors synapse onto the horizontal cells. However, Trümpler and colleagues68 have shown that cone signals can be recorded from horizontal-cell axon terminals in mice lacking connexin 36, suggesting that some signal can make it down the axon at least from the cell body to the axon terminal. They were unable to demonstrate that rod signals could be recorded from the cell bodies after elimination of the gap junctions. This result is surprising: if signal spreads from the cell body to the axon terminal, we should expect it to spread in the opposite direction as well, given the passive properties of signal conduction down the axon and the large size of the axon terminal relative to the cell body.
Horizontal cells provide inhibitory surrounds by feeding back onto both rods and cones (see, for example,70,71). Because both the soma and the axon terminals of horizontal cells receive signals from both kinds of photoreceptors, the rod signal in the horizontal-cell somata could feed back onto cones and the cone signal in the horizontal-cell axons could feed back onto rods. An effect of this kind was recently demonstrated by Szikra and colleagues28, who showed that rod photoreceptors can be modestly depolarized by cone signals coming from horizontal cells. Because horizontal cells receive input from both kinds of photoreceptors, they are potentially capable of mediating antagonistic rod–cone interactions in either direction.
Bipolar cells synapse onto both amacrine and ganglion cells in the next layer of processing, the inner plexiform layer (Figure 1). There are at least 40 different types of amacrine cell, each with a distinguishable morphology and pattern of synaptic contact15. Amacrine cells receive signals from rods or both rods and cones, either directly from bipolar cells or indirectly via the gap junctions between the two kinds of receptors. The A17 amacrine cells, for example, receive synapses almost exclusively from rod bipolar cells72 and make reciprocal GABAergic inhibitory synapses back onto these same bipolar cells73,74. In this way, they provide local feedback inhibition at rod bipolar cell terminals75, which can shape the temporal characteristics of the rod signal conveyed to the rest of the retina and visual system76.
Rod and cone inputs have been particularly well studied for another amacrine cell, called the AII amacrine (or sometimes A2; see, for example,15). In mammals, this cell serves an essential function in dim-light vision because rod bipolar cells do not make direct synaptic connection with ganglion cells. Instead, the rod bipolars make glutamatergic excitatory synapses onto AII amacrine cells, which then transfer the rod signal to cone ON bipolars via gap junctions (see Figure 1 and 46). The AII amacrine also makes glycinergic inhibitory synapses onto OFF cone bipolars (see also 54). As a result, the depolarizing signal of ON rod bipolars produces a depolarizing AII amacrine response, which then depolarizes ON cone bipolars and hyperpolarizes OFF cone bipolars. The AII amacrines also make gap junctions with one another, and, like the rod–cone junctions, these gap junctions are more conductive at night than during the day and are modulated by presynaptic activity77 and by dopamine—but by D1 receptors instead of D2 receptors78,79.
This classic picture of rod signal flow is subject to several qualifications. First, AII amacrines also receive a relatively large direct synaptic input from OFF cone bipolar cells80–82. The functional consequence of this input remains largely unexplored. Second, the gap-junctional input from AII amacrines to ON cone bipolars can proceed in either direction83; as a consequence, depolarization of ON cone bipolars can depolarize AII amacrines and hyperpolarize OFF cone bipolars, some of which receive rod input. Finally, AII amacrine cells are known to synapse directly onto ganglion cells80–82, specifically onto certain classes of OFF ganglion cells84. All of these pathways provide opportunities for rod and cone signals to interact.
Recordings from ganglion cells could have special significance for our understanding of rod and cone interactions because ganglion cell responses reflect the aggregate of integration within the whole retina and specify the nature of the signal sent by the retina to the central nervous system. Some ganglion cells receive only cone signals, most notably the midget ganglion cells in the primate fovea, but the large majority appear to receive both rod and cone input from amacrine cells and cone bipolar cells (see, for example,53,85), allowing their activity to span a large, dynamic range of light intensities.
Although it is possible to stimulate rods and cones selectively with careful selection of light intensity and spectral composition, attempts to investigate rod and cone interactions in ganglion cells have been few and far between (see 8). One of the earliest was the paper of Gouras and Link86, who made extracellular recordings from large (presumably parasol) ganglion cells in the primate perifovea. When brief flashes were given at short intervals to stimulate rods and then cones, or cones and then rods, the first response suppressed the second. Gouras and Link hypothesized that the first response—whether rod or cone—produced a transient inhibition that depressed the second response.
This phenomenon was studied in greater detail and with more powerful techniques nearly 50 years later by Grimes and colleagues87. They also made extracellular recordings from ON parasol cells from primate retina and presented two flashes at short intervals selectively stimulating the rods or cones. They were able to demonstrate clear suppression when stimulation of rods preceded stimulation of cones, but they found little or no effect when the cone signal came first. They then made patch-clamp recordings of voltage responses from horizontal cells, ON cone bipolar cells, and AII amacrine cells, and in each of these cells the rod and cone signals summed linearly. However, they noticed a clear difference in the waveform of rod and cone responses in ON cone bipolar cells: rod responses to brief flashes consisted of a depolarization followed by a pronounced hyperpolarizing undershoot, whereas cone responses were monotonic depolarizations lacking an undershoot. They then constructed a simple model in which rod and cone responses arriving independently at an ON cone bipolar cell were summed linearly and then passed through a common non-linear filter in transit to ganglion cells. This model successfully explained the rod suppression of cone responses, ultimately attributable to the inhibitory undershoot of the rod signal in the cone ON bipolar. Grimes and colleagues showed that this inhibition could be blocked by a cocktail of GABA and glycine receptor inhibitors and may reflect, at least in part, feedback inhibition coming from amacrine cells.
Although there is a large psychophysical literature describing rod and cone interactions in visual behavior as well as detailed information about the anatomy and physiology of sites within the retina where rod and cone interactions can occur, we still know very little about which sites produce which behaviors. The tools currently available for investigating retinal processing continue to evolve and are vastly more powerful now than even 10 years ago. As these tools improve, we may be able to explain more clearly how the rod and cone systems in the eye function separately and together to produce a coherent perception of the world around us.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)