Keywords
heritability, behavioral genetics, genes, development, evolution, environment
heritability, behavioral genetics, genes, development, evolution, environment
Behavioral genetics shares with many other scientific disciplines the goal of learning about the various factors that shape human beings and about what such knowledge implies for the possibility of positive change. However, the particular way in which behavioral genetics has traditionally raised and answered those questions is the subject of continuing controversy (Barnes et al., 2014; Burt & Simons, 2014; Burt & Simons, 2015; Moffitt & Beckely, 2015; Wright et al., 2017).
The problems noted by critics are diverse: they range from statistical assumptions made in estimating heritability, the meaning of those estimates and their relationship to the causes of human traits, to the conceptual distinction between nature (genes) and nurture (environment) underlying behavioral genetics’ core methodology. On a more abstract level, these issues translate into doubts about whether the questions asked by behavioral geneticists make sense, and, if so, whether the methods used to answer them are valid.
This paper has two aims: to diagnose the situation and to offer a way out. The diagnosis will involve showing why the concept of heritability in behavioral genetics lacks utility. The way out leads from recognizing the relationship of heritability in the behavioral genetic sense to its common meaning in biology and then to generalizing this biological notion into a principled concept of heritability based on its causal role in evolution. In the process, a modern view of heritability, development and evolution, united by causal principles, will emerge, and will complete the diagnosis of why, in our view, the traditional behavioral genetic approach has become obsolete.
This paper will focus exclusively on the classical methods of behavioral genetics - variance partitioning into a genetic and one or two environmental components - and will ignore more modern approaches involving the actual measurement of specific genes or environmental factors (i.e. it focusses on G vs E, and ACE models). The reason for this is twofold: first, even though some behavioral geneticists may now disavow the classical methods, these have produced the main body of literature in the field, and understanding the problems with this body is a prerequisite to understanding whether or not modern extensions of behavioral genetics have successfully overcome the problems. Second, even if "only" the classic results were shown to be invalid, it would force the abandonment of a great many core findings about heritability which still inform modern thinking about the genetic nature of traits such as intelligence and diseases such as schizophrenia.
In what follows, we assume the falsity of genetic determinism. This means we reject the idea that genes have more power than other developmental causes to determine the phenotype. We thus exclude a spectrum of views, from strong genetic determinism, the claim that genes fully determine a phenotype no matter what other developmental causes do, to the so-called "information metaphor", the claim that genes, and only genes, carry information in the sense of instructions or a program to control development. The information metaphor is the most widespread form of genetic determinism today, but it has been shown to lack any solid theoretical or empirical basis (Godfrey-Smith, 1999; Godfrey-Smith, 2008; Griffiths, 2001; Griffiths & Gray, 1994; Johnston, 1987; Keller, 1995; Neumann-Held, 2006; Nijhout, 1990; Moss, 2004; Oyama, 1985; Sarkar, 1996; Šustar, 2007).
Charles Darwin shattered the complacent view of his predecessors that God was taking care of guaranteeing the similarity of parents and offspring. After Darwin, the fact of parent-offspring resemblance found itself in urgent need of a non-theistic explanation. Theories of biological inheritance sprang up, and the role of environment and upbringing called for clarification. Darwin’s cousin Francis Galton was perhaps the first to explicitly bring biology (the innate qualities of a person, which he called “nature”) into opposition with the environmental influences that impinge on a person after birth (calling the acquired qualities “nurture”). As is well known, the biologically inherited part later became identified with genes. The view that emerged saw human development as the battleground of two fundamental forces: genes and environment. The question became: which of the two was stronger, and how could their relative strengths be measured?
Galton had already suggested that observing identical twins would be useful in addressing these questions, as their virtual indistinguishability from birth indicated that their biological inheritances (genes) were the same. The degree to which they turned out different should then be a measure of the influence of environment.
By the 1930s, the statistical tools necessary to decompose the effects of nature and nurture had become available in the form of the analysis of variance. It allowed quantification of how much of the variance in an organismic trait was “due to” the variance in any number of explanatory variables. For example, the height to which a plant would grow was expected to depend both on its genotype and on the environment, such as the amount of water in the soil. In a simple experimental design, an agriculturist could plant seeds of different genotypes and expose each genotype to different amounts of water. After the plants had grown to their full height, she would measure how much height varied among the different genotypes (averaged across all water conditions), obtaining the amount of genetic variance. Similarly, she would measure the variation in height across the different levels of exposure to water (averaged across all genotypes) and obtain the environmental variance. If the effects of genotype and water were independent of each other, summing the two variances would yield the total variance in height across all the plants. The ratio of the genetic variance to the total height variance (a number between 0 and 1) could be taken as a measure of the relative strength of the effect of differences in genes on differences in plant height. This number became known as the “heritability” of a trait (Lush, 1937). The ratio of environmental variance to height variance was the complement of heritability and was taken to indicate the strength of environmental differences on trait differences. Comparing these two ratios would supply the answer to Galton’s question as to which was stronger, nature or nurture?
The way to obtain these numbers in humans is, however, importantly different from that in the example above. The facts that monozygotic (“identical”) twins share all of their genes, and that dizygotic (“fraternal”) twins share half of their genes can be used as a shortcut to estimating genetic and environmental variances (how this is done mathematically will not concern us here). The simplest partitioning only comprises a term for genes (G) and one for everything else, called environment (E). The total variance in some phenotypic trait (P) is then simply the sum of the genetic and environmental variance:
P = G + E
A slightly more complex approach is to split the environmental variance in two: one term for environmental influences that make twins more similar to each other (called "shared environmental effects") and one for environmental influences that do not make twins more similar (called "non-shared environmental effects"). This is called the ACE model, where A stands for (additive) genetic effects, C for shared and E for non-shared environmental effects1.
What is important to understand is that apart from the trait of interest (e.g. intelligence, antisocial behavior, political preference), nothing is measured in a classic behavioral genetic study. The subjects’ genotype is not quantified or identified in absolute terms. All that enters the calculations is the assumption that all the genetic variance of monozygotic (MZ) twins contributes to their phenotypic similarity (since they share all of their genes), while only half of the genetic variance of dizygotic (DZ) twins contributes to their phenotypic similarity (since they share only half of their genes). Environmental influences are not measured at all. Instead, the term for the environment (E) is simply the remainder after the phenotypic (P) and genetic (G) variances have been calculated (E = P - G).
This has an important corollary: since the environment is treated as a leftover category, any environmental differences that produce the same pattern of effects as do genetic differences will be counted as genetic effects, not environmental ones. This is true even if the environmental differences actually cause the differences in phenotype.
Perhaps the most striking example of this is the fact that the phenotypic effects of the common egg cell that MZ twins develop from is entirely neglected in twin study designs. MZ twins are created when one single fertilized egg splits in two and the two resulting daughter cells each grow to become a separate individual, while DZ twins stem from two different egg cells fertilized by two different sperm cells. Twin studies make use of the fact that in MZ twins, the two daughter cells will have the same genome - copies of the parent cell’s genome - but ignore that the whole non-genetic part of the two cells (called the “cytoplasm”) also stems from the common parent cell and will therefore also be the same or very similar, whereas the two cells from which DZ twins develop are likely to have less similar cytoplasms. The implication is that in addition to (and in conjunction with) their common DNA, their common cytoplasm will also make MZ twins more similar to each other than DZ twins. To the extent that this effect operates in the 2:1 fashion specified by the mathematical formalism of the ACE model, it is subsumed under the genetic component A, while in reality it is a shared environmental effect belonging to component C. The result is high values for A and low values for C, as is repeatedly found in behavioral genetics studies.
One context in which this matters is the continued debate about the causes of individual or racial IQ differences (see, e.g., recent debate in Vox [Haider, 2017a; Haider, 2017b; Turkheimer et al., 2017a; Turkheimer et al., 2017b]). If twin-based methods of identifying sources of variation confound genetic with cytoplasmic effects, the importance of genes for IQ will have to be re-evaluated. The same is true in another important context: the debate about the relative importance of parents vs. peers, where behavioral genetic findings have given rise to dramatic claims about the global inefficacy of parenting (Boutwell, 2015a; Boutwell, 2015b; Boutwell, 2017a; Boutwell, 2017b; Boutwell & Khan, 2016; Harris, 1998; Rowe, 1994). In fact, what makes cytoplasmic confounding particularly nasty is that there is no known way of disentangling genetic from cytoplasmic effects. The problem affects the entire body of research on twins and heritability. Everything, from antisocial personality to political preferences, that has been claimed to be heritable could potentially be an effect of cytoplasmic similarity, or, more realistically, a joint effect of genetic and cytoplasmic similarity.
A more fundamental charge is sometimes leveled at variance partitioning in the context of developmental causes, a charge that has been considered by a number of critics to be the fatal flaw of behavioral genetics (Burt & Simons, 2014; Burt & Simons, 2015; Moore, 2003). This “interactionist challenge” states that the development of organismic traits is a process of interacting causes that are inextricably entangled, such that their individual causal contributions cannot be separated, let alone quantified. Therefore, attempting to attach numbers to the relative importance of genes and environment in a trait is doomed to fail. Genes and environmental causes are both necessary (and none by itself sufficient) to create a phenotype, it is said, and if that is the case, it is impossible to quantify their relative contributions. Variance partitioning with regard to the causes of development is fatally flawed.
For example, in their recent challenge to behavioral genetics, Burt and Simons (Burt & Simons, 2015) put the case as follows:
“[G]enes and environments are involved in an interpenetrating and interdependent dynamic relationship that renders the attempt to demarcate separate influences— the goal of heritability studies—illogical at both the individual and population levels” (p. 105).
The argument is often illustrated by analogy. One such comes from Richard Lewontin (Lewontin, 1974), who has us imagine two men building a brick wall, where one is laying the bricks and the other is applying the mortar. The two men are analogous to two different developmental causes, and the brick wall represents a phenotypic trait. After their work is done, does it make sense to ask which of the two men has contributed more to the finished wall? The answer is no, because both their contributions are necessary, but not by themselves sufficient, to build the wall.
Another popular analogy is the area of a rectangle (analogous to a trait) and its two causes, length and width. Does it make sense to ask which of them, its length or its width, contributes more to the rectangle’s area? Obviously not. Both length and width are necessary for the area to even exist, so neither can be considered more important. Trying to quantify their relative contributions makes no sense.
How can behavioral geneticists respond to this challenge? First, there is some common ground: the defender of behavioral genetics agrees with the critic’s characterization of how causes operate in development (Barnes et al., 2014; Rutter, 2006)2. Second, however, she correctly notes that this characterization pertains to how individual traits arise in an individual organism, i.e. to the etiology of traits. It says nothing about what causes differences in traits between different individuals. Third, she points out that the latter is what behavioral geneticists are interested in and what they use variance partitioning for. Fourth, she claims that variance partitioning is still possible, even in the case of inextricably interacting developmental causes.
This last claim is correct. In their behavioral genetics textbook, Plomin et al. (2008, p. 85) use the example of the rectangle to show that, although both its length and width are necessary for any individual rectangle’s area to exist, a population of rectangles may not only show variation in length and width, but these two variations may well contribute unequally to the variation in area. In other words, in a particular population of rectangles, the variation in length might be much greater than that in width, and therefore contribute much more to the variation in surface area. In the context of the causes of differences (variation) in a population, length and width are not inseparable; either one of them can exhibit larger variation than the other one and therefore contribute more to differences in the trait of interest, in this case surface area.
Having established the feasibility of variance partitioning for interacting causes, the crucial question becomes: can it actually tell us anything useful about such causes? Critics of behavioral genetics may think not; they may think that variance partitioning is irrelevant, or even illogical when it comes to questions of trait etiology (Burt & Simons, 2014; Burt & Simons, 2015; Moore, 2003). However, far from being irrelevant, variance partitioning is actually one of the major scientific tools used to identify causes in general, including the causes of organismic traits. It is the core of two of the most widespread statistical methods in all of the natural sciences: ANOVA and linear regression, both employed nearly universally in experimental and observational studies to provide clues as to the causes of human and animal traits. Crucially, variance partitioning provides these clues not only when causes are operating independently, but also in the case of interacting developmental causes. This is true simply because if something is a cause of some outcome, varying its level or its presence will change the outcome. Co-variation with an outcome is therefore an indicator of causation. If, for example, some genes are causes of a trait, changing or removing those genes will typically change the trait. (The 'typically' qualifier is necessary because there might be other causal mechanisms preventing the co-variation.) It does not matter whether a cause interacts with or is independent of other causes. One can begin to see this by setting up a simple statistical simulation that embodies the constraints of two interacting causal variables and running a regression analysis, as has been done by Gamma & Liebrenz (2017). Results show positive relationships between the two variables and the outcome, correctly identifying them as possible causes.
The upshot is that the interactionist charge that variance partitioning is useless in the case of interacting developmental causes is invalid. Analysis of variance on the population level can, in principle, identify the causes of traits in individuals, even if these causes are "non-separable" in the sense of being jointly necessary, but singly insufficient, to cause a trait. There is nothing illogical or categorically wrong about this endeavor. Employing variance partitioning per se is not what makes heritability estimates useless. They are useless for quite another reason.
It is not uncommon for technical terms used in science to have a pre-existing meaning. “Heritability” is an example. Its meaning in behavioral genetics is very different from that in common language, including its common meaning in biology. Behavioral genetic heritability (which we will henceforth call “heritabilityBG“) is a single number estimated from twin studies that denotes the proportion of total variance in an outcome variable that is genetic variance. In other words, it is simply the ratio of genetic to total phenotypic variance. As such, its value lies between 0 (no heritabilityBG) and 1 (complete heritabilityBG). In the context of variance partitioning as discussed above, heritabilityBG is the relative size of the variance component due to genes, where “genes” are one of two (or three) causes being considered (the other being “environment”).
Before discussing the implications of the different meanings of “heritability”, we need to assess the utility or lack thereof of heritabilityBG for identifying the developmental causes of traits. We have shown that variance partitioning per se can be useful for identifying such causes, but that is not at all the same as showing that estimating heritabilityBG is useful for this. In fact, the very reason why heritabilityBG estimates lack utility is because they are a particularly unhelpful way of partitioning variance.
The main problem is that the two (or three) sources of variance in behavioral genetic studies are such broad, unspecific and unsystematic categories as to make it virtually impossible to draw any useful causal inferences from them. The ACE model seems to make a potentially useful distinction between two kinds of environmental effects (shared vs. non-shared), but these effects are identified in terms of whether they make twins more similar to each other or not. They do not identify any actual environmental conditions3. Even assuming that heritabilityBG precisely estimates the proportion of genetic variance, all a non-zero value tells us is that genes make a difference to a trait in a population. It leaves us entirely in the dark as to which specific genes are involved in the formation of that trait (Sober, 2001).
It may be objected that the simple statement that genes contribute to a trait (that a trait “is genetic”) already constitutes significant knowledge. Thus, the behavioral geneticist Michael Rutter (Rutter, 2006) writes
"…the overall pattern demands acceptance of the importance of genetic influences. That is, the findings are incompatible with a zero genetic effect". (p. 60)
Frankly, however, that genes are involved universally in the formation of traits is one of the fundamental facts of developmental biology. There was never a reason to assume that a ubiquitous cellular component whose contributions to most aspects of cell life have been extensively documented would somehow magically fail to affect some classes of traits. Another behavioral geneticist, Eric Turkheimer, calls this a “pointless null hypothesis” (Turkheimer, 2011).
Looking at variance partitioning in any field other than behavioral genetics will show how differently it is used. For example, a study of obesity may measure intake of fat, carbohydrates and proteins and find a statistical effect of carbs. This result gives a useful pointer to possible causes, and a follow-up study might then look at different types of sugary foods to narrow down the causes of the effect. Studies all over the natural sciences use experimental manipulations or observational variables specific enough to give potentially useful hints for further investigation. In contrast, knowing that some variance in a trait is genetic, even knowing the amount of that variance, provides little useful knowledge and little idea how to proceed further.
As far as the identification of causes is concerned, heritabilityBG tells us little that is useful. Nevertheless, the brute fact of genetic involvement as indicated by a positive heritabilityBG estimate might be regarded as carrying further significance: it might tell us something about the malleability of a trait.
According to a still-common view, genetic involvement means that a trait, being inherited and therefore “inborn”, is more hard-wired, more fixed, and less amenable to change than a trait that is environmentally caused. Therefore, the fact of genetic involvement is taken to be much more consequential than, for example, the involvement of diet, in causing a trait.
This is not the case, however. Whether genes are or are not involved in the causation of a trait says nothing about how difficult it is to change that trait. In other words, genetically influenced traits are not a priori more difficult to change. A given trait’s malleability is always an empirical question, and the answer may not only be different for different traits, but even for different individuals carrying the same trait.
Perhaps, if a genetically influenced trait could only be altered by changing the relevant genes, an argument could be made that due to the relative inaccessibility as well as the structural and functional complexity of the genome, changing such a trait would always be difficult compared to changing other traits whose etiology does not involve genes. But changing a genetically influenced trait does not necessarily require changing the relevant genes. Traits are always multi-causal, and each of the causes involved, whether genetic or environmental, is, in principle, an access point for causal manipulation.
A famous example is that of the disease phenylketonuria (PKU). It involves a faulty gene that fails to produce an enzyme required to break down a certain amino acid. If left untreated, the amino acid accumulates in the body, leading to early mental retardation. There is a treatment that essentially cures the disease, but it has nothing to do with genes. It consists of a special diet that is free of the offending amino acid, so that a toxic build-up is prevented. A causally simple environmental intervention prevents all symptoms of a genetic disease. Many other such examples could be found, all illustrating the same underlying principle: if multiple causes co-determine an outcome, manipulating any of them can potentially change the outcome. This is true regardless of whether any one of the causes is genetic or not.
It is important to realize that heritabilityBG estimates are not informative about contexts such as this, in which a treatment for a disease trait, or an intervention to improve a beneficial trait (e.g. intelligence) is sought. In particular, the question of whether there could be an environmental intervention to change the trait in a desired direction is not addressed by heritabilityBG, since such an as-yet-unknown intervention cannot be part of the environment of the population from which the heritabilityBG estimate was derived. For example, before a dietary treatment became available, the heritabilityBG of PKU was high, as environmental variation in the disease phenotype was low. This meant nothing, however, about the prospects of finding a successful environmental intervention.
HeritabilityBG does not indicate a trait’s malleability. Genetic involvement per se does not determine malleability, and even the fact that a trait such as PKU is “genetic”, i.e. originates from a gene defect, does not allow any a priori conclusions about how difficult it is to change, and where best to intervene to change it.
In biology, heritability is most commonly understood as a trait's potential to recur across generations due to genetic inheritance. We will henceforth call this notion “heritabilityBio“. To say that a trait is heritableBio means that if it exists in parents, it can recur in offspring because they receive from their parents the genes that give rise to the trait. HeritabilityBio therefore incorporates not only the aspect of "passing on" something to someone, but also a particular causal mechanism for doing so. That mechanism is genetic inheritance, because in the standard view of biology, genes are seen as the only causal substrate for trait inheritance. In "The Selfish Gene", Richard Dawkins puts this bluntly by stating that "genetic factors replicate themselves, blemishes and all, but non-genetic factors do not" (Dawkins, 1976, pp. 98/99).
What is the relationship between heritabilityBG and heritabilityBio? Why do behavioral geneticists call a statistical measure of the proportion of genetic variance in a trait "heritability"?
The answer is that, given the view that heritabilityBio is based exclusively on the transmission of genes, measuring the influence of genetic differences on trait differences in a population would simultaneously capture the extent to which such trait differences are biologically inherited. While superficially plausible, it is clear that the procedure by which heritabilityBG is estimated does not directly probe any aspect of the process of genetic transmission from parents to offspring. It is not surprising, then, that the behavioral genetic measure of heritability sometimes diverges sharply from the biological concept.
For example, consider traits such as bipedality (having two legs) or pentadactyly (having five fingers) in humans. These are paradigmatic features of the "design" of Homo sapiens and universally considered to be highly heritable. However, almost all variation in these traits is environmental, stemming from accidents, and therefore heritabilityBG is basically zero.
The inverse case is possible, too: consider the trait of wearing skirts, which, as most would agree, is primarily culturally determined and not heritableBio. Wearing skirts, however, is almost perfectly correlated with sex, which itself is usually considered genetically determined. Giving precedence to genetic over environmental variation, even when the two correlate, the heritabilityBG of wearing skirts would turn out to be very high. Analogous considerations would show that racism and sexism are also highly heritable, even if in reality they were entirely sociocultural phenomena.
How do these discrepancies occur? In the case of bipedality, what goes wrong is that there is genetic causation, but no genetic variation. And without genetic variation, heritabilityBG is zero. In the other cases, there is environmental causation and therefore correlation, but it is congruent with a genetic correlation, so the effect is ascribed to genes and heritabilityBG is high.
These examples illustrate that behavioral genetic heritability does not track biological heritability as commonly understood.
To summarize the previous three sections, heritabilityBG is of little use in determining the specific causes of a trait, in indicating the malleability of a trait, and in tracking heritability as commonly understood in biology.
Truth is not decided by authority, but it should at least be known that authorities in (the philosophy of) biology share this verdict (Bateson, 2001; Feldman & Lewontin, 1975; Keller, 2010; Lewontin, 2006; Sober, 2001), even if they are perhaps more diplomatic about it than we are here. We specifically mention these scholars because it is philosophers of biology who have done most of the work in developing the conceptual basis of biology, particularly addressing issues such as the role of genes in evolution and development (see for example the collection of essays in Neumann-Held & Rehmann-Sutter, 2006). But prominent behavioral geneticists also acknowledge that estimating heritabilityBG in itself is not very useful. According to Michael Rutter (Rutter, 2006), "It is not that, on its own, it matters very much whether the heritability is 20 percent or 80 percent” (p. 89), while Eric Turkheimer finds variance partitioning as practiced by behavioral genetics to be without “scientific content” entirely (Turkheimer, 2011, p. 598).
Unfortunately, it turns out that the common biological concept of heritability (heritabilityBio) is itself seriously wanting. By fixing it, however, we can arrive at an elegant, unified view of evolution based on a principled definition of heritability. This definition will abstract from contingent, particular mechanisms of inheritance and instead focus on the causal role of heritability in Darwinian evolution. It will be called heritabilityX (the “X” stands for “extended”).
Why was Darwin able to formulate his theory of evolution by natural selection without knowing anything about the actual mechanisms of biological inheritance? Because the actual mechanisms do not matter for the theory, only the fact of heritability itself, understood as the fact that parental traits tend to recur in the offspring generation or, simply put, that offspring tend to resemble their parents.
This basic observation is one part of the “holy trinity” of Darwinism, the three criteria that enable evolution by natural selection: phenotypic variation, differential fitness, and heritability (Sterelny & Griffiths, 1999, p. 32). Together they specify a causal recipe for evolution by natural selection. Importantly, this recipe is abstract enough to leave its physical implementation open. It is defined, rather, by the causal roles it specifies. As long as these roles are implemented in any way in any system, the system will exhibit Darwinian evolution.
In the world of organisms, the recipe specifies that there must be variation in a trait among a population of individuals. Some of the variants cause their carriers to have higher fitness, i.e. to produce more offspring than carriers of other variants. Finally, the trait in question must be heritable, i.e. offspring of carriers of the “successful” trait must be more likely to have the trait themselves. As a result, the trait increases in frequency in the offspring generation.
This three-step process is one iteration of the cycle of Darwinian evolution, and it will repeat as long as there is a fitness differential and no countervailing forces. The successful trait will continue to increase in the population across many generations, which is another way of saying that it evolves by natural selection.
Nothing in this account depends on the particularities of the mechanism by which offspring come to be similar to parents. Rather, the heritability requirement is defined by a particular causal role. This role consists of causing parental traits to recur in offspring. This can happen in potentially many ways, but, following Oyama (1985), we suggest that these ways can be subsumed under three broad categories, using one simple background assumption.
The assumption is that, since traits are multiply caused, the presence of the same or similar causes (in different organisms of the same population) will tend to result in the same or similar traits. Put somewhat informally, the idea is simply “same input - same output”. This is certainly plausible.
A parental trait will therefore recur in offspring to the extent that the same causes that gave rise to the trait in parents are also present in offspring. For a trait to be inherited, therefore, the same or very similar “trait-forming” causes need to be present in both parents and offspring. Oyama (1985) identifies three broad ways in which this can happen: first, the trait-forming causes simply persist across generations; second, the trait-forming causes are physically copied and transported from parents to offspring; and, third, the trait-forming causes are reconstructed in each generation anew. These categories should cover most, if not all, of the possibilities of trait inheritance.
To make the idea more concrete, we will give examples of each category. Persistent trait-forming causes include fixed features of the environment such as sunlight, oxygen, gravity, and humidity. These are always there, and will exert their effect in every generation. Copied trait-forming causes include the familiar case of DNA, which is physically replicated before being passed on to offspring. DNA methylation patterns, if reliably copied across generations, are another example. Reconstructed trait-forming causes are probably the major category. They include all trait-forming causes that recur in every generation without having been directly physically copied or having been always there. Examples are diverse and include the womb (present in every generation, but reconstructed anew from “parts” in every generation), nutrition, social interaction with parents and conspecifics, behavioral habits, cultural norms and practices, and language.
As these examples show, heritability in the evolutionarily relevant sense is more accurately described as “repeated presence across generations”, because this removes the narrow focus on something physical having to be passed on from parents to offspring. Here, however, we will stick to the shorter term “heritabilityX”.
At this point, it is natural to ask what the empirical evidence for non-genetic inheritance in humans is. The answer is that it is all around. This is less a case of having to discover novel non-genetic inheritance mechanisms (although that is part of it, as for example with epigenetic inheritance), but one of many basic and well-known observations that fall into place as soon as they are put in a new conceptual framework. That oxygen and sunlight are reliably present in every generation of our species is a simple fact. That every child starts out from a cell containing its mother’s cytoplasm and begins growing in the rich environment of a womb is another one. That diets can remain stable for long periods of time in local populations is also well-known. That offspring in each generation experience very similar maternal behavior, soak up local cultural norms and practices and learn the local language is no secret, either.
Vastly more than just DNA is stable across generations (Griffiths & Gray, 2001; Griffiths & Gray, 1994; Jablonka & Lamb, 2005; Jablonka & Lamb, 2007; Jablonka & Raz, 2009; Oyama, 1985). This should be uncontroversial. All that is needed then is to recognize that these other trans-generationally stable trait-forming causes meet the heritabilityX requirement for Darwinian evolution just as genes do. They all fulfill the causal role of being reliably present in repeated generations, poised to contribute to the building of the phenotype. They are thus heritable in the one sense that matters, the sense that enables evolution by natural selection.
In human beings, culture will be the major source of non-genetic inheritance (Acerbi & Mesoudi, 2015; Richerson & Boyd, 2008). Factors such as how we raise our children, how we school them, what norms and values we teach, what habits and lifestyles we have, how and where we live, how and where we work, what societies we build, what political and legal systems we have, what technologies we use, what we eat, whether and how we exercise, all this and more is part of a large package of culture children in every generation are shaped by. These packages will be different across different parts of the globe, and different parts of the package will have different “time constants”, i.e. will change faster or more slowly than others. But all will to some extent contribute to the stability or similarity of the phenotypes related across generations by descent.
Non-genetic inheritance is also found in animals with little or no culture. First, persistent trait-forming causes belonging to the physical, geological or biological environment affect most animals just as they do humans. Second, so do reconstructed trait-forming causes like wombs and eggs, a population- or species-specific diet, social interaction with parents and conspecifics, specific forms of housing, etc. Third, there are other, more particular non-genetic inheritance systems in animals that are described in detail in the work of geneticists Jablonka & Lamb (2005); Jablonka & Lamb (2007). The subject matter, particularly in humans, far transcends the current fashionability of “epigenetics” (here, narrowly understood as chemical and structural modifications of DNA).
An extended view of heritability also resolves a contradiction inherent in popular notions that deny genetic determinism, while also finding nothing wrong with the standard biological concept of heritability based exclusively on genetic inheritance. The problem becomes evident in a phrase we used earlier to characterize heritabilityBio: “To say that a trait is heritableBio means that if it exists in parents, it can recur in offspring because they receive from their parents the genes that give rise to the trait.”
This mechanism of inheritance can only work as described if having the right genes more often than not causes the trait in question, regardless of the configuration of other, non-genetic trait-forming causes. In other words, it can only work with a liberal measure of genetic determinism. When genetic determinism is rejected, however, a “paradox” arises: how could trans-generationally similar genes “give rise to” trans-generationally similar traits without other trait-forming causes also being trans-generationally similar?
If genes do not have special deterministic or instructive powers, how could the mere presence of certain genes guarantee the formation of a certain trait, irrespective of the status of other, non-genetic co-determinants of the trait?
The answer is that it could not. Development is paradigmatically a matter of multiple causes interacting to create a phenotype. If only one type of cause (genes) were stable across generations, and all other causes were free to vary arbitrarily, there would be no way of making sure that offspring develop the same traits as their parents4. It is here that the interactionist challenge really bites!
As soon as this is accepted, the pieces fall into place: trait heritability in the evolutionarily relevant sense can only be achieved if all or most or many trait-forming causes are stable across generations - certainly not if only one kind of cause is stable. The rest is re-description: we call all trans-generationally stable trait-forming causes inherited, because that is what they are in the sense relevant to Darwinian evolution (Griffiths, 2001; Griffiths & Gray, 1994; Griffiths & Gray, 1997; Griffiths & Gray, 2001)5. And we call all trans-generationally stable traits heritable, because that is what they are in the sense that is relevant to Darwinian evolution. What we end up with is a view in which many or most traits are heritable, and many or most causes of traits are inherited, at least to some extent. In her book-length treatment of the nature-nurture question, philosopher of science Evelyn Fox Keller (2010) summarizes this new view of heritability:
It can now be seen that both the behavioral genetic and the standard biological concept of heritability are based on the false view that only genes are inherited, or at least that it is only genetic inheritance that matters. That view does not reflect the reality of non-genetic, in particular cultural, inheritance. A deeper understanding of heritability, development, and evolution can only follow from an extended notion of heritability which has a principled causal basis in evolutionary theory.
To elaborate: first, the concept of heritabilityX deepens the understanding of heritability qua parent-offspring resemblance. Far from being a phenomenon with a narrow causal basis limited to genetic inheritance, it is actually supported by a broad range of causal mechanisms. Everything that contributes to the stability of traits across generations is part of an inheritance mechanism. If, for example, offspring reliably get their dietary habits from their parents, then the underlying process of social learning is part of an inheritance mechanism, and diet becomes an inherited (and heritable) trait in the full, evolutionarily licensed, sense of the word.
Broadening the notion of heritability makes sense of fundamental observations about the world, in particular the human world: that a vast array of conditions and stimuli that impinge on us are present in every generation, from the obvious, such as sunlight and oxygen, to the more subtle, such as cultural norms of behavior. Although it is certainly impressive that the DNA molecule is faithfully copied from parents to offspring, it is not only genes that provide stable inputs to the developmental process, and DNA-grade fidelity of replication is required neither for trait heritability nor for biological or cultural evolution (Henrich et al., 2008; Lewis & Laland, 2012; Richerson & Boyd, 2008, p. 83 ff). Rather, there are gradients of similarity of inherited trait-forming causes that lead to gradients of similarity in inherited traits.
Second, if non-genetic inheritance leads to stable changes in the phenotype of a population, there is no reason to deny this process the status of evolution. One might object that the time frame of many non-genetic inheritance phenomena is much shorter than the millions of years traditionally associated with genetic evolution, and that this should militate against them being counted as truly evolutionary. However, we do not see this as a principled objection. For one thing, research in recent decades has shown many examples of genetically driven evolution occurring at time scales as small as months (Gibbs & Grant, 1987; Herrel et al., 2008; Losos et al., 2006; Reznick et al., 1997). Further, it would be unprincipled just to draw a line somewhere and sort all instances of phenotypic change into evolutionary and non-evolutionary based on an arbitrary length threshold.
To take an example: adult body height has increased in Western populations for over a century. On average, current Westerners are over 10 centimeters taller than their ancestors 150 years ago (Danubio & Sanna, 2008). The main reasons for this secular trend are thought to be improvements in diet and hygiene, while genetic effects are deemed unlikely (Silventoinen, 2003). If this is correct, then, since diet and standards of hygiene are certainly (at least partly) culturally inherited, there is no reason not to view this increase in height as true evolutionary change6.
Third, an extended notion of heritability connects with the nature of development in various ways. The most important connection has already been mentioned: it is the fact that rejecting genetic determinism actually implies the notion of heritabilityX. If genes alone cannot and do not determine traits, how could it ever be sufficient for a parental trait to be "rebuilt" in offspring to just inherit the "right" genes? It could not, and therefore non-genetic trait-forming causes must also be inherited from parents7.
Traits are always the product of multiple, interacting causes, none of which is a priori more determinative of the outcome than any other. Heritable traits do not remain stable across generations because there is one special trait-forming cause that instructs the developmental process, but because a sufficient number of different trait-forming causes are inherited so that the process results in a stable phenotype. The principle “same input - same output” applies.
Finally, the view of evolution that emerges is one in which an interacting network of internal and external trait-forming causes envelops and permeates the developing organism, spanning the generations and leading to the reliable recurrence - the heritability - of traits, or to trait variation, depending on the stability or variability of the network components involved (see Figure 1).
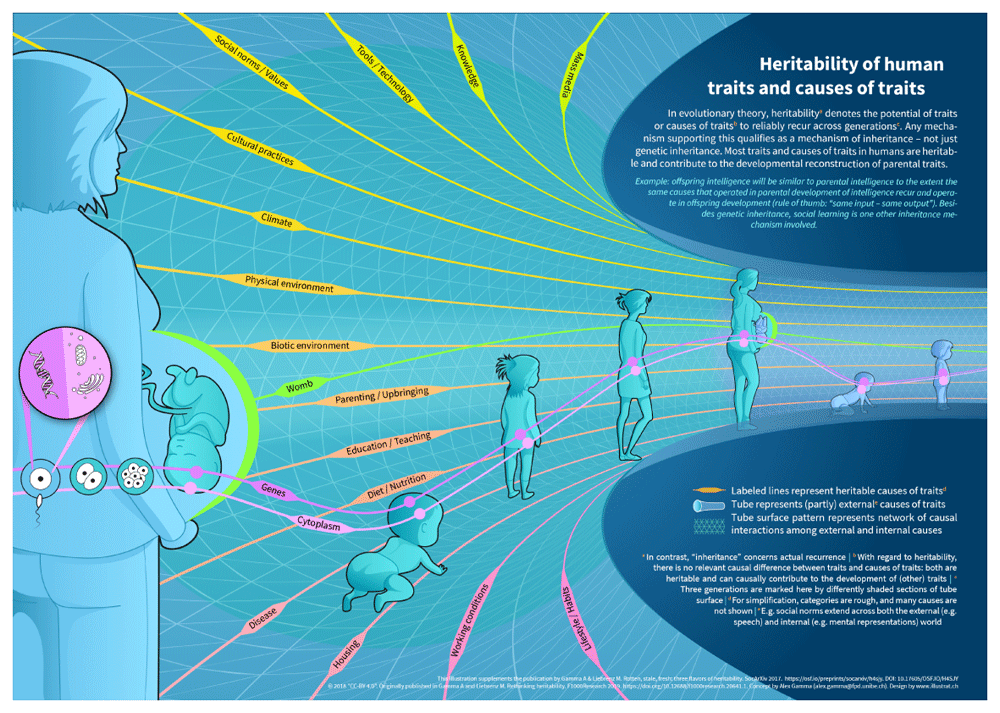
The behavioral genetic concept of heritability - what we have called “heritabilityBG“ - has three major flaws: first, it does not track common expectations about biological heritability; second, it lacks utility for human research since it does not identify specific causes of traits, is silent on the potential success of new therapeutic interventions, and has no bearing on the malleability of traits. Third, what heritabilityBG shares with the common understanding of biological heritability (“heritabilityBio”) is the popular misconception that heritability is based exclusively on genetic inheritance. In other words, both versions confuse “heritable” with “genetic”. This is both theoretically and empirically false; considering the origin and causal role of heritability in Darwin’s scheme of evolution by natural selection shows that, conceptually, any mechanism that guarantees that traits reliably recur across generations will fulfill the heritability requirement of evolution. In human beings, cultural inheritance is the most varied and important such mechanism, but others are possible, and many have in fact been described in various species of animal (Jablonka & Lamb, 2005). A number of fundamental observations about the stability of cultural and physical environments can be re-interpreted as phenomena of inheritance in the Darwinian sense. This leads to an extended causally defined concept of heritability, which we call “heritabilityX“.
HeritabilityX incorporates the insight that most trait-forming causes - genetic and non-genetic - recur in every generation and are therefore heritable in the relevant sense, which is as trans-generational enablers of Darwinian evolution. Standard neo-Darwinism takes inheritance to be exclusively genetic and therefore implicitly requires a form of genetic determinism, typically the so called "information metaphor", the idea that genes embody instructions or even a program to direct the formation of traits. However, there was never any solid theoretical or empirical reason to believe in the information metaphor, as close scrutiny by philosophers of science has demonstrated (Godfrey-Smith, 1999; Godfrey-Smith, 2008; Griffiths, 2001; Griffiths & Gray, 1994; Johnston, 1987; Keller, 1995; Moss, 2004; Neumann-Held, 2006; Nijhout, 1990; Oyama, 1985; Sarkar, 1996; Šustar, 2007).
In a new, updated view of life, traits from the parental generation are successfully re-created in offspring because a sufficient number of parental trait-forming causes - genetic and non-genetic - are inherited and serve as “inputs” to offspring development. Same or similar inputs tend to produce same or similar outputs.
This perspective does justice to the most fundamental observation about development: that it is paradigmatically a process of different kinds of causes interacting constantly to produce traits or phenotypes, and that, in general, these causes are all jointly necessary, but not by themselves sufficient, to generate a certain outcome. There is a “causal parity” (Griffiths & Knight, 1998) of different developmental causes, with none of them playing an a priori more significant or more determinative role than any other.
The causal parity of genes and other developmental factors also implies that genes cannot constitute sufficient causal routes to traits, let alone provide complete explanations of traits. Full-blown explanations will integrate various kinds of causes across different levels of organizational hierarchy, and across the divide between the internal and the external. The impossibly broad categories of nature vs. nurture that captured the imagination of our intellectual ancestors a century ago are no longer fit for the science of today.
No data are associated with this study.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)