Keywords
GWAS, VIGS, T-DNA, CRISPR, Heat stress, Functional genomics
GWAS, VIGS, T-DNA, CRISPR, Heat stress, Functional genomics
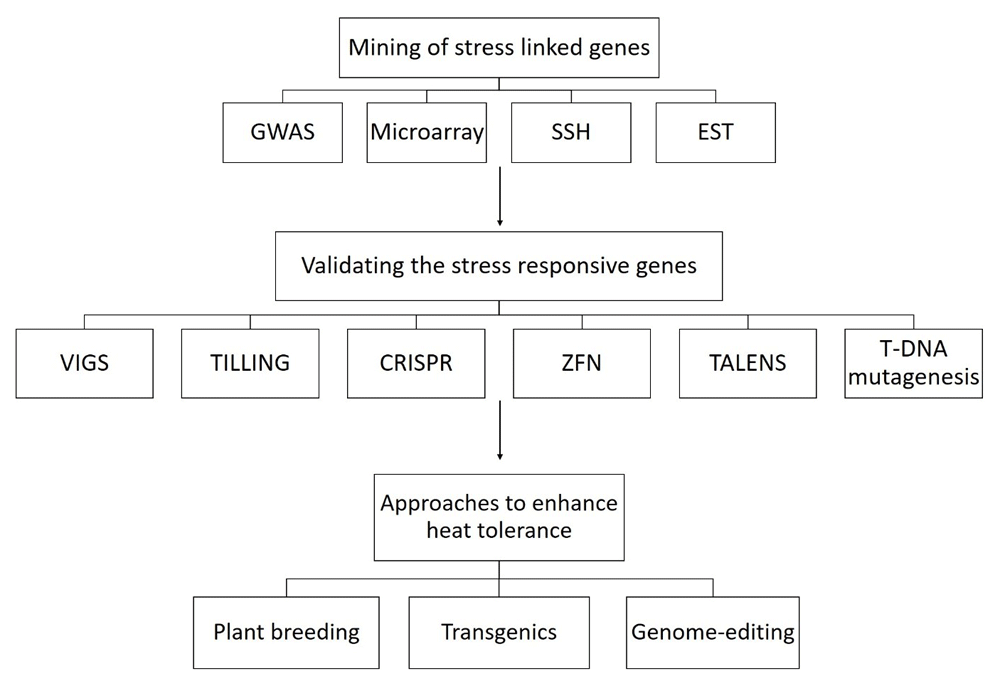
Abiotic stresses have numerous adverse effects on crop plants, which further lead to yield and quality losses (Figure 1). To feed the whole world in the scenario of the changing climate, new and better heat tolerant varieties of various crops is needed1. The understanding of various physiological, molecular and biochemical pathways can facilitate the development of superior heat tolerant varieties2. However, previous efforts, aimed at improving plant heat stress tolerance, have had limited success3,4 because of the poor understanding of the genetics of heat tolerance. Fortunately, nowadays reference genomes of major food crops and model plant species are available publicly, which provide a solid platform for crop improvement. Moreover, wild species and various landraces of various crops have unknown heat tolerant genes that should be identified and incorporated to high yielding modern cultivars5. The functional genomic approaches such as genome wide association studies (GWAS) and gene expression profiling using microarrays can catalyse the discovery of novel genes associated to heat stress6–8. In addition, suppression subtractive hybridization (SSH) is another effective and productive technique used for the screening and cloning of the genes/ESTs that express differentially under heat stress9,10. Reverse genetic techniques can improve the understanding of their expression patterns under heat stress. The plant breeding strategies and new biotechnological tools including genome editing techniques can use these validated genes to enhance heat stress tolerance in crop plants (Figure 2).
Present crop varieties have limited heat tolerance because earlier domestication, green revolution and conventional breeding were focused to increase yield and qualitative traits11. However, the knowledge of genes/markers/QTL regions associated to heat tolerance is now required to improve thermo tolerance. Previous studies suggested that vast genetic diversity still exists in the germplasms of various crops12–14. GWAS emerged as a powerful tool to identify the genetic basis behind complex phenotypic traits15,16, and it provides high mapping resolution compared with conventional genetic mapping17,18. So far this approach has been applied to major food crops, including wheat7,19,20, rice21, maize22, sorghum23and Brassica napus L.24, to identify the natural variation associated with heat stress and to understand this genetic basis. Another way to identify and understand the key molecular mechanisms in response to heat stress is a transcriptomic study25,26; plants respond to heat stress by inducing various heat responsive genes, thus transcriptomic studies provide an effective screening of heat responsive candidate genes6. For example, microarray studies allow the screening of genes on the basis of their expression patterns under stressed conditions at a particular plant developmental stage6,26,27. Singh et al. (2015)6 investigated the heat responsive genes for potato tuberization and Ginzberg et al.28 identified the candidate heat responsive genes for potato periderm formation using microarrays. In addition, SSH is an easy and efficient approach for the identification of genes/ESTs with differential expression under heat stress. This technique is preferred when the genome sequence information is not available9. It can identify the tissue specific differentially expressed transcripts. To identify heat responsive ESTs cDNA libraries can be generated from plants grown under heat stressed conditions29. For example, SSH library of potato skin present 108 candidate genes for suberin and periderm formation30. To investigate the genes/ESTs involved in heat tolerance at the stage of grain filling in wheat, SSH library was constructed by using the leaf RNA samples from heat stressed plants9,29. The results of these studies provided many heat responsive genes/ESTs, which can be used to develop thermo tolerant wheat varieties.
The above approaches can identify potential candidate genes linked to heat stress tolerance. However, the functions of the candidate genes must be validated before incorporating them into present cultivars. Both forward and reverse genetic approaches can be employed for functional validation of genes (see examples in Table 1). Forward genetics detect variations in the nucleic acid sequence responsible for a given phenotype31, while reverse genetics detect the gene’s functionality by observing the change in the phenotype due to alterations in known genetic sequence32.
| Plant/crop | Gene | Technique used | Reference |
|---|---|---|---|
| Arabidopsis thaliana | HSF1 and HSF3 | Transcription control (genetic engineering using protein fusion) | 33,34 |
| DREB2A CA | Microarray | 35 | |
| Hsp70 | antisense gene approach | 36 | |
| ATHSF1 (HSF) | Recombinant DNA technology | 33 | |
| FAD7 | T-DNA | 37 | |
| HSP101 | Transformation | 38 | |
| Rice (Oryzasativa) | spl7 | Transcription control | 39 |
| Athsp101 | Agrobacterium mediated transformation | 40 | |
| Wheat (Triticumaestivum) | TamiR159 | MiR159 (miRNA) | 41 |
| TaGASR1 | Agrobacterium mediated transformation | 42 | |
| Carrot (Daucuscarota) | Hsp17.7 | Hsps and molecular Chaperones | 43 |
| Chilli pepper (Capsicum annuum) | CabZIP63 | Virus induced gene silencing | 44 |
| CaWRKY40 | Virus induced gene silencing | 45 | |
| Tomato (Solanumlycopersicum) | Hsa32 | Subtracted cDNA libraries | 46 |
| MT-sHSP | freezing transformation method | 47 | |
| ATG5, ATG7, NBR1, WRKY33 | Virus induced gene silencing | 48 | |
| 2-CP1, 2-CP2, 2-CP1/2, ATG5, ATG7 | Virus induced gene silencing | 49 | |
| RBOH1, MPK1, MPK2 | Virus induced gene silencing | 50 | |
| Hsc70.1 | Virus induced gene silencing | 51 | |
| SlLrgB | RNA interference | 51 | |
| Barley (Hordeumvulgare) | APX 1 | Agrobacterium mediated transformation | 52 |
Virus-induced gene silencing (VIGS) is a rapid, efficient and cost effective post-transcriptional gene silencing (PTGS) technique used to study target gene(s) functionality53. It can be used as both the forward and reverse genetic approach48,54. Plants can sense and then respond to heat stress by activating various transcriptional cascades55. Being a PTGS technique, VIGS can be used to knockdown the expression of target genes after transcription. VIGS takes an advantage of a plant’s innate defence mechanism against virus infection. In this technique, a fragment of the target gene is first inserted into a suitable viral vector and then that vector is transformed into the plant, where the viral genome harbouring the fragment of target gene start replicating and produce dsRNA. Then an enzyme DICER cut this dsRNA into multiple siRNA of about 21 nucleotide long. Later these siRNA unwind into two single stranded RNAs, one out of which is degraded and the other one binds to RNA induced silencing complex (RISC), which later degrade the targeted endogenous gene and the effect on gene knockdown can be observed on by phenotypic analysis56–58. Many candidate genes and transcriptional factors associated with heat stress response/tolerance have been validated successfully through VIGS58,59. For example, TRV-VIGS based silencing of CabZIP63 gene lowered the tolerance to heat stress in pepper plants, suggesting that CabZIP63 is a positive regulator for thermos tolerance44. The functionality of ATG5, ATG7,FAD7 and NtEDS1genes in response to heat stress have been successfully studied in tomato using tobacco rattle virus (TRV) based VIGS technique48,60. Recently, VIGS has also been employed to investigate the involvement of small heat shock proteins (CaHSP16.4 and CaHsp25. 9) in heat stress tolerance61,62.
The role of candidate genes in heat stress tolerance can also be verified through the generation of transfer (T)-DNA mutants63. Like VIGS, insertion of T-DNA in the target gene’s sequence disrupt its functionality, which results in the change of phenotype. This approach is widely accepted because of the genome wide distribution of transposable elements with superior insertions in the gene sequences, resulting in the direct gene knockout64. In addition, T-DNA can also be used as a gain of function approach to study the target gene’s functionality, called as activation tagging65. For example, T-DNA having a tetramer of cauliflower mosaic virus 35S promoter can cause gene activation mutations65. Since plant responses to environmental stresses are polygenic and complex traits, model plants, such as Arabidopsis, are used to first study adaptive responses66. T-DNA mutant lines of many heat tolerant ecotypes of Arabidopsis have been discovered and are available at Nottingham Arabidopsis Stock Centre (NASC) or The Arabidopsis Information Resource (TAIR).
Targeting induced local lesions in genome (TILLING) is another non-transgenic approach that allows the PCR based identification of directed mutations in the target gene sequence and the function of target gene can be analysed from the modified phenotype due to that mutation67,68. It is a fast and cost efficient technique for the screening of point mutations and for the functional validation gene of interest69. It take advantage of conventional insertional mutagenesis and availability of genomic sequences70. These point mutations can be generated with the help of chemical mutagens such as ethyl methane sulfonate (EMS)71. Nowadays, the genome sequences of many crop plant species are available, which make this technique more effective. The plants under heat stress exhibit different phenotypes associated with allelic variations in their genomic sequence. The TILLING approach used to study these natural variations or SNP mutations in individuals is called as EcoTILLING. The next generation sequencing techniques allow inexpensive TILLING by sequencing method to screen SNP variations72. Recently, the functionality of heat shock binding protein 1 (HSBP1) was examined with TILLING. The chemically induced mutations disrupted the functionality of HSBP1 partially and the mutant plants exhibited increased heat stress tolerance. These findings confirmed that HSBP1 is a negative regulator of heat stress response in tomato73.
In addition, the genome-editing techniques such as transcription activator-like effector nucleases (TALENs), zinc finger nucleases (ZFNs) and clustered regularly interspace short palindromic repeat (CRISPR) can also be used as reverse genetic approaches to study the target gene function. TALENs using sequence specific nucleases (SSNs) became a powerful genome editing technique, which can also be applied as a reverse genetic approach to understand the function of a target gene. It consists of one customizable DNA-binding domain and a nuclease, which generate double stranded DNA breaks (DSBs) at the target gene sequence74. Similar to CRISPR, these DSB are repaired either via NHEJ pathway or via homologous recombination. Both these recovery pathways allow insertion, deletion and intentional replacements in the target gene sequence. These modifications in the target gene’s sequence may cause a variation in the phenotype, which suggests the function of that gene74,75. The ZFNs are the synthetic proteins having a DNA binding domain that consists two finger modules and a DNA cleaving domain. ZFNs causes DSBs in the targeted DNA sequence and facilitate site-specific mutagenesis, or base substation, which alter or may knockout the gene expression76. The ZFNs have revealed the function of various genes in model plants as well as in crop species77.
Among all genome-editing techniques, CRISPR-Cas9 has emerged as a powerful tool for precise genome editing to study the molecular pathways linked to heat stress and to enhance thermo tolerance in crop plants78,79. It is comparatively simpler, more accurate and faster than other genome editing techniques. In brief, CRISPR involves designing of a guide RNA of ~20 nucleotides complementary to the gene of interest and a Cas9 nuclease enzyme that cut 3–4 bases next to the protospacer adjacent motif, which is later repaired either by homology directed repair pathway or via error prone non-homologous end joining80,81. Therefore, this technique can be used to generate gene knockout mutant lines to study the function of targeted gene(s). For example, annexin gene OsAnn3 knockout mutant lines developed via CRISPR-Cas9 technique revealed the role of OsAnn3 gene in cold stress tolerance in rice82.
Plants have inherent mechanisms to survive under heat stressed conditions but the heat tolerance capacity of plants varies species-to-species and even within the species. If heat tolerant genes are present in sexually compatible species, then marker-assisted selection (MAS), new generation molecular breeding, precision breeding and genome editing techniques can be used.
Thermo tolerance is a complex multigenic trait, which is influenced by genotype X environment interactions83. Development of heat tolerant crop varieties through traditional breeding is very labours and time consuming. However, precision breeding with the help of MAS can accelerate the plant breeding programs with high efficiency84. SNPs and simple sequence repeats (SSR) are being used widely in plant breeding experiments aimed to enhance abiotic stress tolerance. Presently the use of SNPs become more common in plant breeding than SSR markers85,86. Garg et al.87 found one SNP in the sequence of heat shock protein (HSP16.9) between a heat tolerant and heat susceptible wheat genotypes. This SNP contribute 29.89% phenotypic variation for grain weight per spike. Recently, many SNPs associated to heat stress tolerance have been identified in major food crops7,88–90. However, heat stress tolerance is a polygenic trait and a single molecular marker contribute little to improve it. Therefore, it is important to incorporate several SNPs associated to various QTLs that are controlling the heat stress tolerance mechanisms89,91. However, the accessibility of genome editing techniques opened various new windows to introduce targeted editing of plant genomes to understand the molecular aspects involved in heat stress tolerance92,93. For example, ethylene response factors (ERFs) are the stress induced transcriptional factors that take part in abiotic stress tolerance. The CRISPR-Cas9 based genome editing of one such ethylene response factor from AP2/ERF superfamily enhanced abiotic stress tolerance in crop plants94.
In cases when heat tolerant genes do not exist in sexually compatible species, these methods cannot be applied. Advanced biotechnological tools can increase the limited heat stress tolerance in crop plants. The transfer of heat tolerant genes through recombinant DNA technology can generate heat tolerant transgenic lines in a short amount of time. This method also allows utilisation of potential genes from other species to enhance thermo tolerance in target crops, e.g. AmDREB2C, from Ammopiptanthus mongolicus has been used to increase heat stress tolerance in transgenic Arabidopsis plants95. In addition, many genes responsible for heat stress tolerance have been identified and validated in model plants and also in major food crops that can be introduced to heat susceptible cultivars or their expression levels can be increased by generating their stable overexpression lines. For instance, the overexpression TaPEPKR2 gene enhanced heat stress tolerance in wheat and Arabidopsis plants96.
Heat stress affects crop production significantly. Plants respond to heat stress by activating complex molecular networks, such as signal transduction, metabolite production and expressions of heat stress-associated genes. With the developments in plant functional genomics techniques, many novel genes related to heat stress tolerance have been identified and are being used to improve stress tolerance with the help of advanced biotechnological approaches. Next generation sequencing and genome-editing techniques will play crucial roles in crop improvement. In the near future, the scientists will have a better understanding of plant heat tolerant mechanisms and farmers will be able to grow better high yielding heat tolerant crop varieties in the fields.
No data is associated with this article.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)