Millius A, Ode KL and Ueda HR. A period without PER: understanding 24-hour rhythms without classic transcription and translation feedback loops [version 1; peer review: 2 approved]. F1000Research 2019, 8(F1000 Faculty Rev):499 (https://doi.org/10.12688/f1000research.18158.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Laboratory for Synthetic Biology, RIKEN Quantitative Biology Center, 1-3 Yamadaoka, Suita, Osaka, 565-0871, Japan 2Laboratory of Systems Immunology and Laboratory of Host Defense, Immunology Frontier Research Center, Osaka University, Suita, Osaka, 565-0871, Japan 3Department of Systems Pharmacology, Graduate School of Medicine, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo, 113-0033, Japan
Arthur Millius
Roles:
Writing – Original Draft Preparation,
Writing – Review & Editing
Koji L. Ode
Roles:
Writing – Original Draft Preparation
Since Ronald Konopka and Seymour Benzer’s discovery of the gene Period in the 1970s, the circadian rhythm field has diligently investigated regulatory mechanisms and intracellular transcriptional and translation feedback loops involving Period, and these investigations culminated in a 2017 Nobel Prize in Physiology or Medicine for Michael W. Young, Michael Rosbash, and Jeffrey C. Hall. Although research on 24-hour behavior rhythms started with Period, a series of discoveries in the past decade have shown us that post-transcriptional regulation and protein modification, such as phosphorylation and oxidation, are alternatives ways to building a ticking clock.
Corresponding author:
Hiroki R. Ueda
Competing interests:
No competing interests were disclosed.
Grant information:
This work was supported by a grant from the Japan Agency for Medical Research and Development (AMED)-Core Research for Evolution Science and Technology (CREST) (JP17gm0610006; AMED/Ministry of Education, Culture, Sports, Science and Technology [MEXT]; to HRU), CREST (Japan Science and Technology Agency/MEXT; to HRU), Brain Mapping by Integrated Neurotechnologies for Disease Studies (Brain/MINDS) (AMED/MEXT; to HRU), Basic Science and Platform Technology Program for Innovative Biological Medicine (AMED/MEXT; to HRU), a Grant-in-Aid for Scientific Research (Japan Society for the Promotion of Science 18H05270 to HRU), a Grant-in-Aid for Scientific Research on Innovative Areas (23115006 to HRU), the strategic programs for research and development (President’s Discretionary Fund) of RIKEN (to HRU), an intramural Grant-in-Aid from the RIKEN Quantitative Biology Center (to HRU), a Grant-in-Aid for Young Scientists (18K14683 to KLO), the RIKEN Special Postdoctoral Research Program (to AM), and a Grant-in-Aid for Early-Career Scientists (18K14755 to AM).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
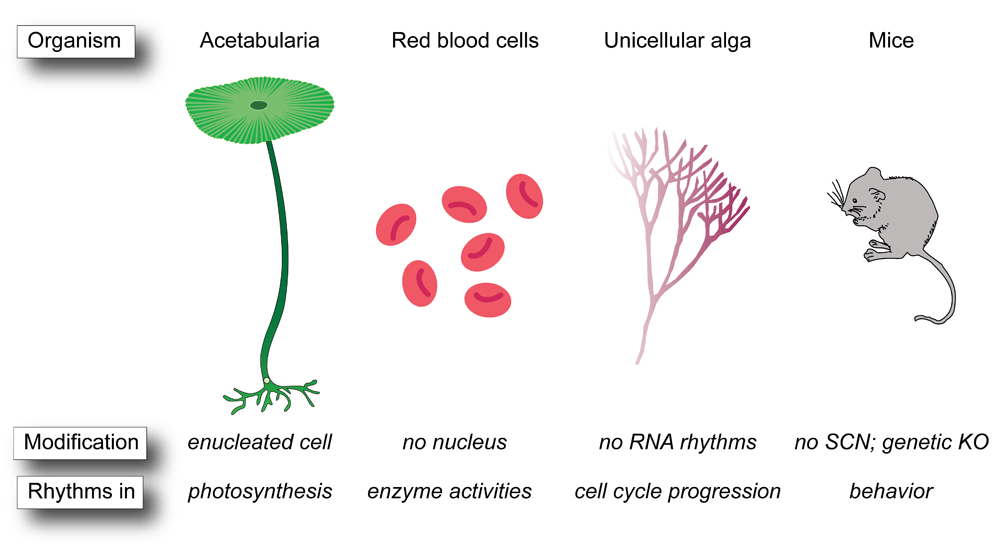
The time-keeping mechanisms of circadian rhythms can be regulated by multiple layers of different cellular networks, including transcription-translation feedback loops (TTFLs) and post-translation oscillators (PTOs)1. Circadian TTFLs generate oscillations in gene expression through delayed negative feedback whereby expression of a transcription factor negatively regulates its own transcription2. The core of this genetic network in mammals is the expression of a heterodimer of BMAL1 (also called ARNTL) with either CLOCK or NPAS2, which binds at promoter cis-elements called E-boxes to drive expression of genes encoding period (PER1-3), cryptochrome (CRY1-2), and nuclear receptor subfamily (NR1D1-2) proteins, which then repress Bmal1 expression by a series of separate and interconnected feedback loops3,4. In contrast to behaviors driven by cyclic differences in gene expression, PTOs generate rhythms independent of transcription and translation through biochemical processes, such as phosphorylation, protein–protein interactions, and other post-translational modifications. These post-translational processes also alter TTFLs as well as post-transcriptional modification of transcripts involved in TTFLs5,6. The most well-known PTO is the cyanobacteria KaiABC system, which consists of only three proteins and ATP7, but novel PTOs may also exist in red blood cells (RBCs)8–14, which lack a nucleus and the molecular machinery to drive TTFL rhythms. In addition, a series of new and old observations of 24-hour rhythms in biological contexts where classic TTFLs are absent or diminished (Figure 1)15–23 continue to puzzle researchers and demonstrate that there are multiple ways to build a clock.
Figure 1. Post-translation oscillators without transcription-translation feedback loops.
Examples of post-translation oscillators in enucleated cells such as Acetabularia and red blood cells, in unicellular alga lacking RNA rhythms, and in mice in which the classic transcription-translation feedback loop module is disrupted genetically or anatomically. KO, knockout; SCN, suprachiasmatic nucleus.
One of the first modern uses of the term “circadian” was to describe 24-hour endogenous oscillators that alter Drosophila fly behavior rhythms24, and the persistence of oscillations at various temperatures was viewed as a defining feature of circadian rhythms25,26. The first genetic component of circadian rhythms was discovered in the 1970s when Ronald Konopka in Seymour Benzer’s lab used chemical mutagenesis of Drosophila to discover three alleles of the Period gene27. In the 1980s, rhythmicity of Period mutants was shown to be restored by gene transfer28,29, confirming that Period both is necessary and can restore rhythmic behaviors, such as eclosion and locomotor activity, in flies. In 1990, Hardin et al. proposed that PER protein altered the levels of Period mRNA in a negative feedback loop30, but at the time it was unclear whether PER directly suppressed Period transcription or whether the negative feedback occurred through an indirect route. A few years later, researchers discovered that this negative feedback was direct in the bread mold Neurospora crassa model of circadian rhythms because the frequency (FRQ) directly repressed its own transcription31. In addition to Neurospora32 and Drosophila33, TTFL models of circadian rhythms from plants34 to mammals35 have been elucidated and reviewed extensively.
Post-translational oscillators and post-translational modifications: breaking the transcription-translation feedback loop mold
The modern idea that TTFLs were necessary for 24-hour rhythms was shattered in 2005 when Nakajima et al. reconstituted rhythmic 24-hour oscillations in protein phosphorylation with just a small number of cyanobacterial proteins36. This seminal moment in the circadian rhythm field spurred investigators to examine other non-canonical rhythm-generating mechanisms and to unearth forgotten studies of PTOs. For example, in the 1960s, it was shown that the unicellular alga Acetabularia undergoes diurnal rhythms of photosynthesis, which persist even after the nucleus has been artificially removed21.
There are a few more recent examples of organisms that have circadian rhythms in the absence of TTFLs. In the unicellular red alga Cyanidioschyzon merolae, circadian rhythms control cell cycle progression in the absence of RNA translation22, and the unicellular dinoflagellate Lingulodinium has daily rhythms in bioluminescence and photosynthesis without a detectable change in RNA transcript abundance and in the presence of transcription inhibitors23. These studies suggest that protein activities and post-translational modifications can serve as 24-hour oscillators. Research has centered on phosphorylation as the period-determining post-translational modification37–40, but other post-translational modifications, including methylation, acetylation, sumoylation, and ubiquitination, also alter clock function41–44.
Importantly, circadian rhythms are insensitive to temperature and this property of temperature compensation was identified in biological time-keeping systems, such as those of bees, flies, and marine organisms, as early as the 1950s and 1960s25,26,45,46. Transcription and translation are temperature-dependent reactions47–50, which suggests that post-translational activities are important for temperature compensation. For example, Isojima et al. revealed that phosphorylation by casein kinase I (CKI) is a temperature-insensitive period-determining process, and the degradation rate of PER2, which is regulated by CKI phosphorylation, was found to be insensitive to temperature38. Importantly, the phosphorylation of PER2-derived peptide by CKI is insensitive to temperature in vitro. In 2015, the degradation of PER2 was found to occur in a more complex mode composed of three distinct stages, and the duration of the second stage depended on circadian time, which led to the identification of temperature-sensitive and -insensitive PER2 phosphorylation sites51. Thus, differences in the temperature sensitivity of phosphorylation sites on the repressor, which alter degradation rates at different temperatures, are responsible for temperature compensation, PER2 stability, and ultimately the length of the circadian period. In 2017, Shinohara et al. identified a short sequence region around residue K224 in CKI, which was responsible for temperature compensation and converted a temperature-sensitive kinase into a temperature-insensitive one in vitro52. Mutation of K224 shortens circadian behavioral rhythms and alters the temperature dependency of the circadian clock in the sub-hypothalamic region of the brain52, called the suprachiasmatic nucleus (SCN), which controls circadian response to light. It is though noteworthy that K224 is part of the consensus KRQK monopartite nuclear localization signal in CKI, which makes it difficult to disentangle the effects of temperature dependence from that of localization in vivo. These studies provide evidence for how post-translational activities modify TTFL rhythms, but a series of new and old studies have revealed that PTOs can drive rhythms even in the absence of TTFL clocks.
Blood: a novel source of post-translational oscillator rhythms
Mammals have a natural supply of enucleated cells in RBCs, and researchers have plumbed this cell type for non-TTFL rhythms. In the 1970s, circadian rhythms in ATPase activity and periodic rhythms in enzymes—such as acetylcholinesterase, glyceraldehyde-3-phosphate dehydrogenase, and glucose-6-phosphate dehydrogenase—in RBCs were found (Table 1), but it was unclear whether the rhythms were robust or persistent beyond 24 hours10.
Table 1. Oscillatory phenomena observed in human red blood cells.
Molecule
Year
Period
Impact
Reference
Glucose-6-phosphate dehydrogenase
1975
~12 hours
Observed two peaks in enzyme activity over a 24-hour period in three different individuals
Observed two peaks in activity over a 24-hour period with one pattern peaking at 4 p.m. and midnight and the other peaking at midnight and 8 p.m. in six and five individuals, respectively
Observed two peaks in activity over a 24-hour period with one pattern peaking at noon and midnight and the other peaking at 4 a.m. and 4 p.m. in four and seven individuals, respectively
Observed variations in acetylcholinesterase activity over a 24-hour period in four individuals, but variations had lower amplitude than reference13 and were not circadian
In 2011, an anti-oxidant enzyme called peroxiredoxin (PRX) in cultured human RBCs was found to have temperature-independent circadian cycles of hyperoxidation for up to 76 hours11. Because RBCs lack a nucleus and the rhythms persisted in the presence of transcription and translation inhibitors, a novel non-transcriptional-based circadian oscillator in mammals was proposed. Analysis of the PRX rhythms relied solely on PRX1, PRX2, and PRX-SO2/3 (hyperoxidized PRX form) antibodies. In particular, the PRX-SO2/3 antibody recognizes multiple hyperoxidized forms of PRX53, results in up to eight different bands on non-reducing sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE)11, and produces multiple non-specific bands that can confound interpretation of the hyperoxidized signal54,55, which make determination of the correct PRX isoform technically difficult and in-gel controls essential. Nevertheless, the same researchers discovered that hyperoxidized PRX-SO2/3 rhythms were conserved in a wide range of species55,56.
In mice, blocking hemoglobin oxygen transport by incubation with carbon monoxide eliminates PRX2 hyperoxidized rhythms57. Hemoglobin auto-oxidation in RBCs generates superoxide, which is converted to H2O2 by superoxide dismutase 1 (SOD1)58,59, and H2O2 is subsequently reduced by catalase, glutathione peroxidase, and PRXs54,58,60, which results in the oxidation of these proteins61. Oxidation of PRX2 is reversed by sulfiredoxin (SRX)62–64, but rhythms in PRX2 oxidation in mice are not mediated by the rhythmic reduction of hyperoxidized PRX2 by SRX but rather through rhythmic degradation by 20S proteasomes, and only about 1% of the total PRX pool is modified in a circadian manner57. Mitochondria-specific PRX (PRX3) is also reversibly inactivated by hyperoxidation, reduced, and reactivated by SRX, and hyperoxidized PRX3 and SRX undergo anti-phasic circadian oscillations in the mitochondria in various mice tissues, which links mitochondria function to circadian rhythms65. Another group revealed about three peaks in hyperoxidized PRX-SO2/3 rhythms in mice over a 48-hour period (instead of two as would be expected for a circadian rhythm) and showed that rhythms were impaired in SOD1-mutant mice66. There is still uncertainty regarding the origin of PRX hyperoxidation rhythms, but the data suggest that PRX-SO2/3 oscillations are more an output of rhythm-generating machinery involving the 20S proteasome rather than a daily oxidation-reduction cycle.
Deconstructing PRX rhythms biochemically and using non-antibody methods, such as mass spectrometry, to directly detect the hyperoxidized cysteine residue or redox-sensitive fluorescent proteins may bolster understanding of this novel PTO. However, biochemical reconstruction is difficult because RBC lysis causes gradual loss of PRX-SO2/3 signal over a 48-hour period9. However, these types of approaches have revealed that potassium-containing media enhances PRX2-SO2/3 rhythms9, and chemical perturbation with Conoidin A, a PRX2 inhibitor67, shortens PER2:LUCIFERASE rhythms in immortalized mouse fibroblasts68.
The exact mechanism of rhythmic PRX oxidation is still unclear, but researchers have begun to examine other general rhythmic behaviors in RBCs. Although no one has followed up on the circadian, ultradian, and irregular rhythms of various enzyme activities in the 1970s, in 2011 researchers reported circadian changes in NADH and NADPH levels11, and in 2017 researchers reproduced circadian changes in RBC membrane potential9 observed in an article published in 197612. Paradoxically, the researchers reported circadian changes in potassium concentration in 2017, whereas no rhythms in potassium were observed in 1976; instead, a gradual and steady increase in potassium efflux occurred over the 48-hour observation period. Whether these differences arise from measuring slightly different potassium populations (intracellular versus extracellular), a small sample size, technical differences in methods and individuals, or an actual biological phenomenon remains to be determined.
Post-translation control of circadian period in transcription-translation feedback loop model organisms
There are shared design principles between the period-determination processes of PTO-based and TTFL-based oscillators. A PTO generates rhythmic changes in protein states without changing the amount of protein itself. On the other hand, rhythmic protein synthesis and degradation are essential for TTFL-based oscillations, and mechanisms that control protein abundance are critical for controlling the circadian period. This idea is widely accepted for circadian TTFL oscillators because there is a significant correlation between the half-life of transcription repressor mutants, such as in Drosophila PER and Neurospora FRQ, and circadian period69,70. The correlation suggests that faster degradation of circadian repressors accelerates clock speed. In mammalian circadian clocks, F-box proteins recruit E3 ubiquitin ligase complexes that license PER and CRY degradation, which modulates period length71. Although the circadian TTFL-based oscillators involve post-translational regulation as period-determination mechanisms, modification of transcription repressors regulates period length by changing repressor stability. For example, a mutation in CKIε that destabilizes mammalian PER results in period shortening72,73, and mutation of a phosphorylation site on PER that destabilizes PER also results in period shortening74,75. Other kinases, such as AMPK and DNA-PK, control period length by altering CRY stability through phosphorylation76,77. In addition, stabilization of CRY by small molecules lengthens the period78, and destabilization of CRY by degron tagging of CRY shortens the period79, strongly suggesting the causal relationship between CRY stability and period length.
However, a recent study of the Neurospora circadian clock challenged this protein stability–period length paradigm of period determination in a TTFL-based oscillator80. Researchers used an FWD-1–deleted strain, which is an F-box protein that causes proteolysis of phosphorylated FRQ. The Δfrd-1 strain results in a markedly increased FRQ half-life, and new FRQ is produced even in the presence of hyperphosphorylated FRQ. Nonetheless, circadian oscillation of FRQ-promoter activities persists with modest change in period length, and several short-period mutations of FRQ still have a short period in a Δfrd-1 background in which the stability of FRQ is significantly increased. Because mutation of phosphorylation sites in FRQ still alters the period and because generic inhibition of kinase activity lengthens the period even in the absence of FWD-1, these data suggest that a protein-state not a protein-abundance attribute, namely phosphorylation, controls period length.
A similar uncoupling of protein stability and circadian period may occur even in the TTFL clock in mammals. A recent study of CRY1 mutations in phosphorylation sites by Ode et al. revealed that multiple phosphorylation sites near the co-factor binding pocket of CRY1 markedly changes period length while having only a modest effect on CRY1 half-life79. Mutagenesis of CRY1 and CRY2 revealed mutations in a secondary co-factor binding pocket which shorten the period without reducing CRY1 stability81. Furthermore, an exon-skipping mutation in CRY1 found from a human family with delayed sleep phase syndrome lengthens the period without affecting CRY1 stability82. Therefore, mammalian CRY may also control the circadian period independently of its abundance.
If protein abundance control does not explain all aspects of period determination, what is the nature of state control of TTFL-based oscillator proteins such as multisite phosphorylation of FRQ, PER, and CRY? One of the shared properties of period-determining repressor proteins is structural flexibility. Most FRQ and PER regions modified by multisite phosphorylation are intrinsically disordered, highly flexible, and variable83,84. The multisite phosphorylation region of CRY1 critical for period control also occurs on a flexible loop region. These flexible regions may undergo a relatively large conformation change that may underlie slow dynamics (that is, 24 hours) of protein activity change. The intrinsically disordered C-terminal domain of BMAL1 controls the period through a slow conformation change with a high energy barrier85. Conformation changes may lead to a slow and coherent re-organization of the macromolecular repressor complex86, which is consistent with the dynamics of the cyanobacteria PTO87,88 in which the slow dynamics of the intrinsic conformational change of KaiC89 couple to the re-organization of the KaiABC complex90. An atomic-scale understanding of the repressor complex in a TTFL-based oscillator may reveal subtle differences in molecular mechanisms of 24-hour period determination between PTO- and TTFL-based oscillators.
Oscillations without classic transcription-translation feedback loop oscillators
Several classic models of circadian rhythms have persistent 24-hour rhythms even when the circadian TTFL machinery is absent or disrupted. In S2 cells, which are generally regarded as non-rhythmic, a multi-omics approach recently revealed hundreds of genes, proteins, and metabolites with 24-hour rhythms20. Although this approach seems to suggest the presence of a novel non-canonical oscillator with 24-hour periodicity, it does not preclude possible cell cycle effects from the roughly 24-hour doubling time of S2 cells or the possibility of classic circadian clock components operating below the experimental limits of detection. For example, large-scale proteomics studies of circadian variation frequently fail to detect circadian proteins91,92 because there may be only a few hundred to a thousand protein copies per cell93. Thus, genetic knockout (KO) of canonical clock genes is needed to definitively determine whether rhythms derive from a novel oscillator.
In mammals, genetic and anatomical ablation of the circadian machinery normally disrupts 24-hour behavioral rhythms, but rhythms persist under specialized situations. For example, SCN-lesioned rats administered methamphetamine in the drinking water retain circadian behaviors of activity in constant light conditions17. This so-called methamphetamine-sensitive oscillator also does not depend on classic circadian genes, such as Per1-2, Cry1-2, Bmal1, Npas2, and Clock18,94. Recent data suggest that the methamphetamine-sensitive oscillator is a long-period manifestation of a tunable dopamine ultradian oscillator95,96. KO of a dopamine transporter in SCN-lesioned or Bmal1 KO mice, which prevents dopamine reuptake in dopaminergic neurons, increases the period of the ultradian rhythms. Similarly, administration of methamphetamine, which increases extracellular dopamine concentrations, lengthens ultradian rhythms in a dose-dependent manner from 4 hours to an astonishing 48 hours. In contrast, the anti-psychotic drug haloperidol, which selectively blocks the dopamine D2 receptor, shortens long-period rhythms induced by methamphetamine in wild-type and Bmal1 KO mice95. These data suggest that dopamine neurons are a second independent rhythm-generating mechanism in the brain, and future studies using chemical and genetic approaches to perturb dopamine pathways coupled with recently developed brain-clearing techniques97–100 may enable a more complete understanding of the neural architecture of this dopamine ultradian oscillator.
Conclusions
From blood to brain, these studies suggest that non-canonical PTOs have an impact on circadian rhythms beyond the classic PER negative feedback loop. However, recent studies of PER itself, including temperature-sensitive phosphorylation sites51, three prime untranslated region (3′-UTR) regulation101, and the separation of Period2 rhythms from Bmal1 rhythms in the SCN102, indicate that even a gene as well studied as Period can still teach us new tricks about the period-determining mechanisms of circadian rhythms.
Abbreviations
AMPK, AMP-activated protein kinase; BMAL1, brain and muscle Arnt-like protein 1; CKI, casein kinase I; CLOCK, circadian locomotor output cycles kaput; CRY1-2, cryptochrome1-2; FRQ, frequency; FWD-1, F-box/WD-40-repeat-containing protein 1; KO, knockout; NPAS2, neuronal PAS domain-containing protein 2; NR1D1-2; nuclear receptor subfamily 1 group D member 1-2; PER; period; PRX, peroxiredoxin; PTO, post-translational oscillator; RBC, red blood cell; S2, Schneider 2; SCN, suprachiasmatic nucleus; SOD1, superoxide dismutase 1; SRX, sulfiredoxin; TTFL, transcription-translation feedback loop.
Grant information
This work was supported by a grant from the Japan Agency for Medical Research and Development (AMED)-Core Research for Evolution Science and Technology (CREST) (JP17gm0610006; AMED/Ministry of Education, Culture, Sports, Science and Technology [MEXT]; to HRU), CREST (Japan Science and Technology Agency/MEXT; to HRU), Brain Mapping by Integrated Neurotechnologies for Disease Studies (Brain/MINDS) (AMED/MEXT; to HRU), Basic Science and Platform Technology Program for Innovative Biological Medicine (AMED/MEXT; to HRU), a Grant-in-Aid for Scientific Research (Japan Society for the Promotion of Science 18H05270 to HRU), a Grant-in-Aid for Scientific Research on Innovative Areas (23115006 to HRU), the strategic programs for research and development (President’s Discretionary Fund) of RIKEN (to HRU), an intramural Grant-in-Aid from the RIKEN Quantitative Biology Center (to HRU), a Grant-in-Aid for Young Scientists (18K14683 to KLO), the RIKEN Special Postdoctoral Research Program (to AM), and a Grant-in-Aid for Early-Career Scientists (18K14755 to AM).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Faculty Opinions recommended
References
1.
Hurley JM, Loros JJ, Dunlap JC:
Circadian Oscillators: Around the Transcription-Translation Feedback Loop and on to Output.
Trends Biochem Sci.
2016; 41(10): 834–46. PubMed Abstract
| Publisher Full Text
| Free Full Text
2.
Partch CL, Green CB, Takahashi JS:
Molecular architecture of the mammalian circadian clock.
Trends Cell Biol.
2014; 24(2): 90–9. PubMed Abstract
| Free Full Text
3.
Brown SA, Kowalska E, Dallmann R:
(Re)inventing the circadian feedback loop.
Dev Cell.
2012; 22(3): 477–87. PubMed Abstract
| Publisher Full Text
4.
Ueda HR, Hayashi S, Chen W, et al.:
System-level identification of transcriptional circuits underlying mammalian circadian clocks.
Nat Genet.
2005; 37(2): 187–92. PubMed Abstract
| Publisher Full Text
8.
Cornelius G, Rensing L:
Daily rhythmic changes in Mg2+-dependent ATPase activity in human red blood cell membranes in vitro.
Biochem Biophys Res Commun.
1976; 71(4): 1269–72. PubMed Abstract
| Publisher Full Text
10.
Mabood SF, Newman PF, Nimmo IA:
Circadian rhythms in the activity of acetylcholinesterase of human erythrocytes incubated in vitro [proceedings].
Biochem Soc Trans.
1978; 6(1): 305–8. PubMed Abstract
| Publisher Full Text
12.
Hartman H, Ashkenazi I:
Circadian changes in membrane properties of human red blood cells in vitro, as measured by a membrane probe.
FEBS Lett.
1976; 67(2): 161–3. PubMed Abstract
| Publisher Full Text
13.
Ashkenazi IE, Hartman H, Strulovitz B, et al.:
Activity rhythms of enzymes in human red blood cell suspensions.
J Interdiscipl Cycle Res.
1975; 6(4): 291–301. Publisher Full Text
14.
Brok-Simoni F, Ashkenazi YE, Ramot B, et al.:
The diurnal rhythm of enzymes in human red cells.
Br J Haematol.
1976; 32(4): 601–8. PubMed Abstract
| Publisher Full Text
15.
Clarke JD, Coleman GJ:
Persistent meal-associated rhythms in SCN-lesioned rats.
Physiol Behav.
1986; 36(1): 105–13. PubMed Abstract
| Publisher Full Text
16.
Dragovic Z, Tan Y, Görl M, et al.:
Light reception and circadian behavior in 'blind' and 'clock-less' mutants of Neurospora crassa.
EMBO J.
2002; 21(14): 3643–51. PubMed Abstract
| Publisher Full Text
| Free Full Text
17.
Honma K, Honma S, Hiroshige T:
Activity rhythms in the circadian domain appear in suprachiasmatic nuclei lesioned rats given methamphetamine.
Physiol Behav.
1987; 40(6): 767–74. PubMed Abstract
| Publisher Full Text
19.
de Paula RM, Lewis ZA, Greene AV, et al.:
Two circadian timing circuits in Neurospora crassa cells share components and regulate distinct rhythmic processes.
J Biol Rhythms.
2006; 21(3): 159–68. PubMed Abstract
| Publisher Full Text
21.
Sweeney BM, Haxo FT:
Persistence of a Photosynthetic Rhythm in Enucleated Acetabularia.
Science.
1961; 134(3487): 1361–3. PubMed Abstract
| Publisher Full Text
22.
Miyagishima Sy, Fujiwara T, Sumiya N, et al.:
Translation-independent circadian control of the cell cycle in a unicellular photosynthetic eukaryote.
Nat Commun.
2014; 5: 3807. PubMed Abstract
| Publisher Full Text
23.
Roy S, Beauchemin M, Dagenais-Bellefeuille S, et al.:
The Lingulodinium circadian system lacks rhythmic changes in transcript abundance.
BMC Biol.
2014; 12: 107. PubMed Abstract
| Publisher Full Text
| Free Full Text
24.
Pittendrigh CS:
Circadian rhythms and the circadian organization of living systems.
Cold Spring Harb Symp Quant Biol.
1960; 25: 159–84. PubMed Abstract
| Publisher Full Text
26.
Pittendrigh CS:
On Temperature Independence in the Clock System Controlling Emergence Time in Drosophila.
Proc Natl Acad Sci U S A.
1954; 40(10): 1018–29. PubMed Abstract
| Publisher Full Text
| Free Full Text
28.
Bargiello TA, Jackson FR, Young MW:
Restoration of circadian behavioural rhythms by gene transfer in Drosophila.
Nature.
1984; 312(5996): 752–4. PubMed Abstract
| Publisher Full Text
29.
Zehring WA, Wheeler DA, Reddy P, et al.:
P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster.
Cell.
1984; 39(2 Pt 1): 369–76. PubMed Abstract
| Publisher Full Text
30.
Hardin PE, Hall JC, Rosbash M:
Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels.
Nature.
1990; 343(6258): 536–40. PubMed Abstract
| Publisher Full Text
31.
Aronson BD, Johnson KA, Loros JJ, et al.:
Negative feedback defining a circadian clock: autoregulation of the clock gene frequency.
Science.
1994; 263(5153): 1578–84. PubMed Abstract
| Publisher Full Text
38.
Isojima Y, Nakajima M, Ukai H, et al.:
CKIepsilon/delta-dependent phosphorylation is a temperature-insensitive, period-determining process in the mammalian circadian clock.
Proc Natl Acad Sci U S A.
2009; 106(37): 15744–9. PubMed Abstract
| Publisher Full Text
| Free Full Text
39.
Chiu JC, Ko HW, Edery I:
NEMO/NLK phosphorylates PERIOD to initiate a time-delay phosphorylation circuit that sets circadian clock speed.
Cell.
2011; 145(3): 357–70. PubMed Abstract
| Publisher Full Text
| Free Full Text
40.
Lee HM, Chen R, Kim H, et al.:
The period of the circadian oscillator is primarily determined by the balance between casein kinase 1 and protein phosphatase 1.
Proc Natl Acad Sci U S A.
2011; 108(39): 16451–6. PubMed Abstract
| Publisher Full Text
| Free Full Text
41.
Brown SA, Ripperger J, Kadener S, et al.:
PERIOD1-associated proteins modulate the negative limb of the mammalian circadian oscillator.
Science.
2005; 308(5722): 693–6. PubMed Abstract
| Publisher Full Text
44.
Naidoo N, Song W, Hunter-Ensor M, et al.:
A role for the proteasome in the light response of the timeless clock protein.
Science.
1999; 285(5434): 1737–41. PubMed Abstract
| Publisher Full Text
46.
Enright JT:
Temperature compensation in short-duration time-measurement by an intertidal amphipod.
Science.
1967; 156(3781): 1510–2. PubMed Abstract
| Publisher Full Text
47.
Oliveira SM, Häkkinen A, Lloyd-Price J, et al.:
Temperature-Dependent Model of Multi-step Transcription Initiation in Escherichia coli Based on Live Single-Cell Measurements.
PLoS Comput Biol.
2016; 12(10): e1005174. PubMed Abstract
| Publisher Full Text
| Free Full Text
49.
Farewell A, Neidhardt FC:
Effect of temperature on in vivo protein synthetic capacity in Escherichia coli.
J Bacteriol.
1998; 180(17): 4704–10. PubMed Abstract
| Free Full Text
50.
Craig N:
Effect of reduced temperatures on protein synthesis in mouse L cells.
Cell.
1975; 4(4): 329–35. PubMed Abstract
| Publisher Full Text
52.
Shinohara Y, Koyama YM, Ukai-Tadenuma M, et al.:
Temperature-Sensitive Substrate and Product Binding Underlie Temperature-Compensated Phosphorylation in the Clock.
Mol Cell.
2017; 67(5): 783–798.e20. PubMed Abstract
| Publisher Full Text
53.
Cox AG, Pearson AG, Pullar JM, et al.:
Mitochondrial peroxiredoxin 3 is more resilient to hyperoxidation than cytoplasmic peroxiredoxins.
Biochem J.
2009; 421(1): 51–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
54.
Low FM, Hampton MB, Peskin AV, et al.:
Peroxiredoxin 2 functions as a noncatalytic scavenger of low-level hydrogen peroxide in the erythrocyte.
Blood.
2007; 109(6): 2611–7. PubMed Abstract
| Publisher Full Text
57.
Cho CS, Yoon HJ, Kim JY, et al.:
Circadian rhythm of hyperoxidized peroxiredoxin II is determined by hemoglobin autoxidation and the 20S proteasome in red blood cells.
Proc Natl Acad Sci U S A.
2014; 111(33): 12043–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
58.
Johnson RM, Goyette G Jr, Ravindranath Y, et al.:
Hemoglobin autoxidation and regulation of endogenous H2O2 levels in erythrocytes.
Free Radic Biol Med.
2005; 39(11): 1407–17. PubMed Abstract
| Publisher Full Text
60.
Cho CS, Lee S, Lee GT, et al.:
Irreversible inactivation of glutathione peroxidase 1 and reversible inactivation of peroxiredoxin II by H2O2 in red blood cells.
Antioxid Redox Signal.
2010; 12(11): 1235–46. PubMed Abstract
| Publisher Full Text
| Free Full Text
61.
Rhee SG, Chae HZ, Kim K:
Peroxiredoxins: a historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling.
Free Radic Biol Med.
2005; 38(12): 1543–52. PubMed Abstract
| Publisher Full Text
62.
Jeong W, Park SJ, Chang TS, et al.:
Molecular mechanism of the reduction of cysteine sulfinic acid of peroxiredoxin to cysteine by mammalian sulfiredoxin.
J Biol Chem.
2006; 281(20): 14400–7. PubMed Abstract
| Publisher Full Text
64.
Woo HA, Chae HZ, Hwang SC, et al.:
Reversing the inactivation of peroxiredoxins caused by cysteine sulfinic acid formation.
Science.
2003; 300(5619): 653–6. PubMed Abstract
| Publisher Full Text
65.
Rhee SG, Kil IS:
Mitochondrial H2O2 signaling is controlled by the concerted action of peroxiredoxin III and sulfiredoxin: Linking mitochondrial function to circadian rhythm.
Free Radic Biol Med.
2016; 100: 73–80. PubMed Abstract
| Publisher Full Text
66.
Homma T, Okano S, Lee J, et al.:
SOD1 deficiency induces the systemic hyperoxidation of peroxiredoxin in the mouse.
Biochem Biophys Res Commun.
2015; 463(4): 1040–6. PubMed Abstract
| Publisher Full Text
67.
Haraldsen JD, Liu G, Botting CH, et al.:
Identification of Conoidin a as a Covalent Inhibitor of Peroxiredoxin II.
Org Biomol Chem.
2009; 7: 3040–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
68.
Causton HC, Feeney KA, Ziegler CA, et al.:
Metabolic Cycles in Yeast Share Features Conserved among Circadian Rhythms.
Curr Biol.
2015; 25(8): 1056–62. PubMed Abstract
| Publisher Full Text
| Free Full Text
69.
Syed S, Saez L, Young MW:
Kinetics of doubletime kinase-dependent degradation of the Drosophila period protein.
J Biol Chem.
2011; 286(31): 27654–62. PubMed Abstract
| Publisher Full Text
| Free Full Text
70.
Ruoff P, Loros JJ, Dunlap JC:
The relationship between FRQ-protein stability and temperature compensation in the Neurospora circadian clock.
Proc Natl Acad Sci U S A.
2005; 102(49): 17681–6. PubMed Abstract
| Publisher Full Text
| Free Full Text
71.
Hirano A, Fu YH, Ptáček LJ:
The intricate dance of post-translational modifications in the rhythm of life.
Nat Struct Mol Biol.
2016; 23(12): 1053–60. PubMed Abstract
| Publisher Full Text
72.
Meng QJ, Maywood ES, Bechtold DA, et al.:
Entrainment of disrupted circadian behavior through inhibition of casein kinase 1 (CK1) enzymes.
Proc Natl Acad Sci U S A.
2010; 107(34): 15240–5. PubMed Abstract
| Publisher Full Text
| Free Full Text
74.
Shanware NP, Hutchinson JA, Kim SH, et al.:
Casein kinase 1-dependent phosphorylation of familial advanced sleep phase syndrome-associated residues controls PERIOD 2 stability.
J Biol Chem.
2011; 286(14): 12766–74. PubMed Abstract
| Publisher Full Text
| Free Full Text
75.
Vanselow K, Vanselow JT, Westermark PO, et al.:
Differential effects of PER2 phosphorylation: molecular basis for the human familial advanced sleep phase syndrome (FASPS).
Genes Dev.
2006; 20(19): 2660–72. PubMed Abstract
| Publisher Full Text
| Free Full Text
76.
Lamia KA, Sachdeva UM, DiTacchio L, et al.:
AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation.
Science.
2009; 326(5951): 437–40. PubMed Abstract
| Publisher Full Text
| Free Full Text
77.
Gao P, Yoo SH, Lee KJ, et al.:
Phosphorylation of the cryptochrome 1 C-terminal tail regulates circadian period length.
J Biol Chem.
2013; 288(49): 35277–86. PubMed Abstract
| Publisher Full Text
| Free Full Text
79.
Ode KL, Ukai H, Susaki EA, et al.:
Knockout-Rescue Embryonic Stem Cell-Derived Mouse Reveals Circadian-Period Control by Quality and Quantity of CRY1.
Mol Cell.
2017; 65(1): 176–90. PubMed Abstract
| Publisher Full Text
81.
Rosensweig C, Reynolds KA, Gao P, et al.:
An evolutionary hotspot defines functional differences between CRYPTOCHROMES.
Nat Commun.
2018; 9(1): 1138. PubMed Abstract
| Publisher Full Text
| Free Full Text
83.
Querfurth C, Diernfellner AC, Gin E, et al.:
Circadian conformational change of the Neurospora clock protein FREQUENCY triggered by clustered hyperphosphorylation of a basic domain.
Mol Cell.
2011; 43(5): 713–22. PubMed Abstract
| Publisher Full Text
89.
Abe J, Hiyama TB, Mukaiyama A, et al.:
Circadian rhythms. Atomic-scale origins of slowness in the cyanobacterial circadian clock.
Science.
2015; 349(6245): 312–6. PubMed Abstract
| Publisher Full Text
91.
Mauvoisin D, Wang J, Jouffe C, et al.:
Circadian clock-dependent and -independent rhythmic proteomes implement distinct diurnal functions in mouse liver.
Proc Natl Acad Sci U S A.
2014; 111(1): 167–72. PubMed Abstract
| Publisher Full Text
| Free Full Text
92.
Robles MS, Cox J, Mann M, et al.:
In-vivo quantitative proteomics reveals a key contribution of post-transcriptional mechanisms to the circadian regulation of liver metabolism.
PLoS Genet.
2014; 10(1): e1004047. PubMed Abstract
| Publisher Full Text
| Free Full Text
93.
Narumi R, Shimizu Y, Ukai-Tadenuma M, et al.:
Mass spectrometry-based absolute quantification reveals rhythmic variation of mouse circadian clock proteins.
Proc Natl Acad Sci U S A.
2016; 113(24): E3461–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
94.
Honma S, Yasuda T, Yasui A, et al.:
Circadian behavioral rhythms in Cry1/Cry2 double-deficient mice induced by methamphetamine.
J Biol Rhythms.
2008; 23(1): 91–4. PubMed Abstract
| Publisher Full Text
97.
Susaki EA, Tainaka K, Perrin D, et al.:
Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis.
Cell.
2014; 157(3): 726–39. PubMed Abstract
| Publisher Full Text
98.
Ertürk A, Becker K, Jährling N, et al.:
Three-dimensional imaging of solvent-cleared organs using 3DISCO.
Nat Protoc.
2012; 7(11): 1983–95. PubMed Abstract
| Publisher Full Text
1
Laboratory for Synthetic Biology, RIKEN Quantitative Biology Center, 1-3 Yamadaoka, Suita, Osaka, 565-0871, Japan 2
Laboratory of Systems Immunology and Laboratory of Host Defense, Immunology Frontier Research Center, Osaka University, Suita, Osaka, 565-0871, Japan 3
Department of Systems Pharmacology, Graduate School of Medicine, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo, 113-0033, Japan
Arthur Millius
Roles:
Writing – Original Draft Preparation,
Writing – Review & Editing
Koji L. Ode
Roles:
Writing – Original Draft Preparation
This work was supported by a grant from the Japan Agency for Medical Research and Development (AMED)-Core Research for Evolution Science and Technology (CREST) (JP17gm0610006; AMED/Ministry of Education, Culture, Sports, Science and Technology [MEXT]; to HRU), CREST (Japan Science and Technology Agency/MEXT; to HRU), Brain Mapping by Integrated Neurotechnologies for Disease Studies (Brain/MINDS) (AMED/MEXT; to HRU), Basic Science and Platform Technology Program for Innovative Biological Medicine (AMED/MEXT; to HRU), a Grant-in-Aid for Scientific Research (Japan Society for the Promotion of Science 18H05270 to HRU), a Grant-in-Aid for Scientific Research on Innovative Areas (23115006 to HRU), the strategic programs for research and development (President’s Discretionary Fund) of RIKEN (to HRU), an intramural Grant-in-Aid from the RIKEN Quantitative Biology Center (to HRU), a Grant-in-Aid for Young Scientists (18K14683 to KLO), the RIKEN Special Postdoctoral Research Program (to AM), and a Grant-in-Aid for Early-Career Scientists (18K14755 to AM).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Millius A, Ode KL and Ueda HR. A period without PER: understanding 24-hour rhythms without classic transcription and translation feedback loops [version 1; peer review: 2 approved]. F1000Research 2019, 8(F1000 Faculty Rev):499 (https://doi.org/10.12688/f1000research.18158.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)