Keywords
Intestinal stem cell, Homeostasis, Regeneration, Plasticity, Epigenetics, Nich
Intestinal stem cell, Homeostasis, Regeneration, Plasticity, Epigenetics, Nich
Intestinal epithelium is one of the fastest renewing tissues in mammals. Owing to the constant exposure of its luminal surface to injurious factors, such as pathogens and toxins, the average turnover rate is every three to five days in mice to replenish damaged cells1. These single-layered intestinal epithelial cells are organized into a crypt-villus structure and are predominantly composed of absorptive enterocytes and four secretory lineages known as enteroendocrine, Paneth, goblet, and tuft cells (Figure 1). The thin intestinal epithelial sheet is responsible for nutrient absorption and stool compaction and also serves as a front-line barrier against microorganisms and infections2. Maintenance and regeneration of this high-turnover tissue upon injury are fueled primarily by intestinal stem cells (ISCs) that reside at the bottom of the crypt, while differentiated absorptive and secretory cells are assembled as clusters or scattered along the crypt-villus axis3,4. More recently, considerable cellular plasticity was noted within intestine, and lineage-restricted progenitors or fully differentiated cells were able to replenish the tissue under certain conditions, such as DNA damage–induced injuries5–11.
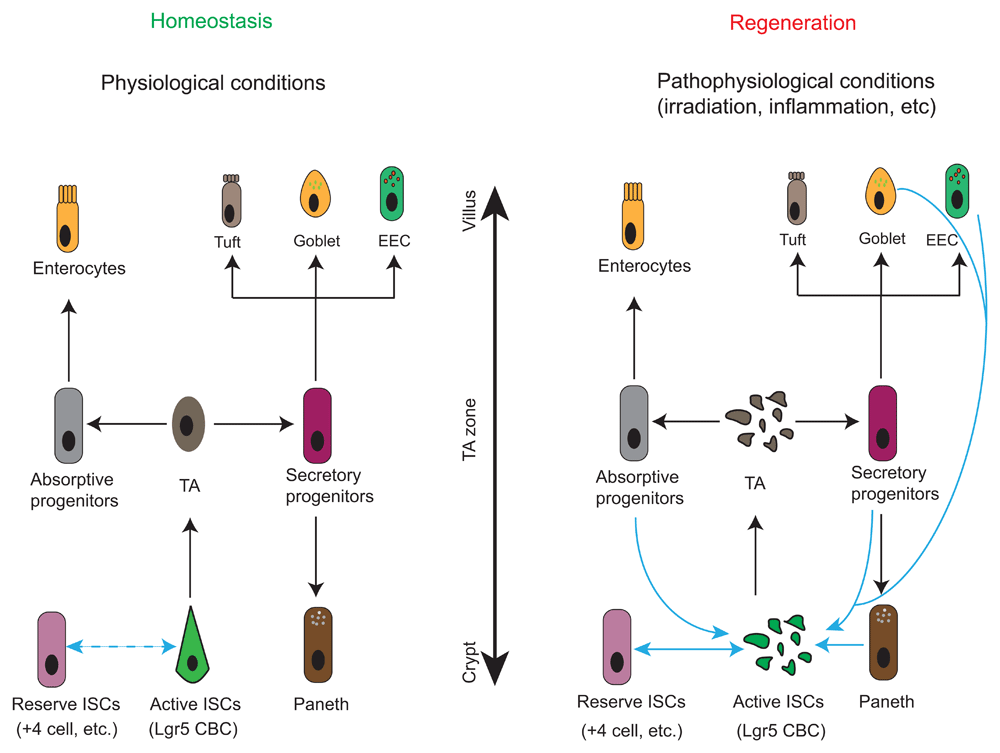
Under homeostatic conditions, active ISCs such as Lgr5 crypt base columnar cells (CBCs) migrate upwards to become transit-amplifying (TA) cells in the TA zone. TA cells divide rapidly and specify into either absorptive or secretory progenitors. Absorptive progenitors further differentiate into large quantities of enterocytes, while secretory progenitors commit to the Paneth, goblet, enteroendocrine (EEC), or goblet cells. Interconversion between reserve ISCs and active ISCs occurs occasionally in this setting (demonstrated by double-headed dash line). In response to radio- or chemo-therapies, the highly proliferative Lgr5 CBCs and TA cells are ablated. Reserve ISCs enter the cell cycle to replenish CBCs for subsequent regenerative process. Plasticity of differentiated progenies, including secretory and absorptive progenitors as well as terminally differentiated EEC and Paneth cells, has been observed when CBCs are damaged. These cells can revert to active ISCs and give rise to all intestinal cell types. However, whether they can bypass CBCs to transdifferentiate directly into other intestinal lineages and their functional importance to intestinal regeneration upon injuries remain to be determined.
ISCs possess both self-renewing capacity and multipotency to give rise to all types of intestinal epithelial cells. They divide and migrate upward to the middle region of the crypt to convert into transit-amplifying (TA) cells that divide rapidly for massive expansion before specializing into absorptive or secretory lineages4. Owing to the tremendous regenerative capacity and the simple anatomic structure, intestine and particularly its ISCs have become an elegant model system for studying homeostasis, regeneration, and oncogenic transformation of mammalian adult tissues12,13. Self-renewal and multipotency of ISCs and plasticity of intestinal epithelial cells are largely controlled by external signals emanated from neighboring niche cells and intrinsic molecular processes, including epigenetic regulation14,15. In support of a central role of extracellular niche factors, ISCs embedded in the crypt bottom are sustained by signals emanated from both epithelial and mesenchymal niches. In the in vitro 3D culture system, ISCs are able to self-organize into crypt-villus–like structures referred to as “organoids” (or precisely enteroids or colonoids if derived from small intestine or colon, respectively) in the presence of a defined set of growth factors16. These organoids comprise self-renewing ISCs intermingled with Paneth cells at the base of budding crypt and various differentiated lineages at blunt villus-like compartments and can be grown and maintained for many passages without losing normal karyotype over time17.
In this review, we summarize the latest advances in our understanding of ISC identity, cellular plasticity, the basis for intestinal homeostasis and regeneration as well as how ISC self-renewal and multipotency are regulated, with a particular focus on extrinsic niche-derived signaling and intrinsically epigenetic regulation. Considering such progress in the mechanistic understanding of intestinal homeostasis and regeneration as well as the development of new models and techniques to faithfully mimic intestinal pathophysiology, we envision a variety of potent and effective therapeutic approaches for the treatment of intestinal diseases.
For decades, crypts have been known as compartments comprising cellular sources for continuous intestinal homeostasis and robust post-injury regeneration18. However, the cellular basis and nature of ISCs that fuel the rapid renewal of intestine have been among the mysteries in the field of adult stem cell biology. It has long been assumed that mammalian tissue-resident adult stem cells, including ISCs, predominantly reside out of the cell cycle in a relatively quiescent G0 state so that genomic integrity can be sustained in response to genotoxic insults2,19. However, this prevailing notion has been amended by the identification of long-lived yet rapidly dividing intestinal crypt base columnar cells (CBCs) with relatively specific expression of Lgr520. They self-renew and are capable of differentiating into all types of intestinal epithelial cells in in vivo, as evidenced by lineage-tracing studies of in vivo and in vitro cultured organoids16,20,21. Owing to their mitotically active feature, Lgr5 CBCs were termed active ISCs and thought to sustain physiological homeostasis of the rapid renewing intestine3. Intriguingly, a subset of epithelial cells residing specifically at +4 position relative to the base of crypts was observed to share some properties of tissue-resident adult stem cells, such as the ability of long-term DNA label retention and a strong resistance to stress, including chemotherapy and irradiation19,22,23, and thus had been postulated to represent ISCs long before Lgr5 CBCs were identified.
Lgr5 CBCs are mitotically active and can regenerate whole intestinal epithelium under homeostatic conditions20. However, owing to their exquisite sensitivity to genotoxic stresses, Lgr5 CBCs are rapidly lost upon radio-/chemo-induced damage and thus could not account for the robust regenerative potential of post-injury intestine24. Moreover, studies with genetic ablation of Lgr5 CBCs by diphtheria toxin (DT) treatment of mice harboring Lgr5-driven DT receptor (DTR) allele revealed that these cells are dispensable for normal intestinal homeostasis, implying the existence of other epithelial cells with both stem cell activity and DNA damage–resistant capacity to replace Lgr5 CBC loss for intestinal regeneration25. Multiple populations of rare crypt cells marked by Bmi126, Hopx26, mTert27, Krt1928, Lrig129, Sox930, Mex3a31, or Prox16 have been found to reside at approximately +4 position by short-term CreER-activated cell fate mapping assay. In sharp contrast to Lgr5 CBCs, most cells labeled by these reporter alleles are slowly cycling and injury-resistant and can give rise to clonal lineage-tracing events albeit at much lower frequency than Lgr5 CBCs5. In light of the above features, these reporter-marked, predominantly +4 resident cells were defined as reserve ISCs in the literature3.
In contrast to their unique spatial localization noted in genetic-marked reporter assays, transcriptomic analyses revealed that endogenous Bmi1, mTert, and Hopx are broadly expressed throughout crypt cells, even in the active Lgr5 CBCs, reflecting a certain inconsistency between reporter activity and actual mRNA expression of the endogenous alleles32–34. Multiple reasons could underlie this discrepancy, such as (1) difference in the 3′ untranslated region (UTR) sequence between CreER reporter and endogenous alleles. A direct comparison between the mRNA level of CreER reporter and endogenous alleles among distinct populations of crypt cells could determine whether CreER reporter can faithfully recapitulate expression of its endogenous counterpart at transcriptional and post-transcriptional levels. (2) As activation of genetic reporters in lineage-tracing studies requires reaching a certain threshold of CreER activity, cells marked by genetic reporters following short-term tamoxifen administration may point to a stronger CreER activity in these cells than in other populations of crypt cells, which could result from differential levels of CreER protein or tamoxifen permeability in distinct types of crypt cells. Ideally, development of immunohistological grade antibodies that can specially recognize the endogenous protein of reported markers for ISCs will resolve these discrepancies and help determine their actual distribution pattern throughout crypts.
As reserve ISCs marked by Bmi1+, Hopx+, mTert, Lrig1, and so on can give rise to all types of epithelial lineages, including the active Lgr5 CBCs in lineage-tracing studies, they have initially been posited to sit at the apex of the cellular hierarchy in intestine25,26. However, single- or bulk-cell transcriptomic profiling analyses have invariably detected the expression of a few enteroendocrine markers within reserve populations of ISCs, indicating some common features between reserve ISCs and committed enteroendocrine cells or hinting at a potential developmental plasticity of lineage-restricted secretory progenitors in intestine35. It should be mentioned that definition of reserve ISCs hinges solely on the functional criteria, including injury resistance, multipotency, cell cycle entry from G0 to G1 upon damage, and long-term maintenance, all of which can be illustrated by Cre recombinase–induced lineage-tracing assays3,5. Cells marked with Bmi1+, Hopx+, mTert, and so on fulfill these functionally defining criteria and could act as bona fide reserve ISCs for tissue regeneration24,26,27,36. The fact that stem cells can express multiple-lineage genes in a fluctuating way, assumed as priming, has been reported in adult stem cells37 as well as in pluripotent embryonic stem cells, which express a significant level of representative genes for primordial germ cells in an undifferentiated pluripotent state38–40. Therefore, caution should be exercised when defining ISC identity simply on the basis of transcriptomic analysis. Nevertheless, more intense studies to further investigate the heterogeneity of +4 epithelial cells with single-cell high-throughput strategies are required to deepen our understanding of the identity of reserve stem cells in intestine.
Cellular plasticity has been observed in several mammalian tissues and could act as an additional mechanism for tissue regeneration41. Direct evidence showing developmental plasticity of enteroendocrine progenitors comes from lineage tracing of cells marked with Prox1, which is a transcription factor expressed predominantly in mature enteroendocrine cells and is essential for its commitment from ISCs6,42–45. Prox1 enteroendocrine cells can function as reserve ISCs as they assume both homeostatic and injury-inducible stem activity in lineage-tracing reporter assays6. Besides mature enteroendocrine cells, considerable plasticity and clonal lineage-tracing events have been noted in other progenies of Lgr5 CBCs, such as enteroendocrine progenitors marked by Bmi17, Alpi+ absorptive progenitors8, secretory precursors expressing Atoh110 or Dll111, CD69+ CD274+ goblet precursor7, fully differentiated Paneth cells46,47, and a small population of Dclk1+ tuft cells48 in certain situations, such as inflammation, radio-/chemo-induced intestinal injuries, or upon loss of Lgr5 CBCs by DT administration (Figure 1). However, the functional robustness and contribution of these lineage-committed cells to intestinal homeostasis and restoration upon tissue damage remain unclear and will be worth future investigation. Furthermore, whether these differentiated epithelia can bypass Lgr5 CBCs and transdifferentiate directly into other epithelial lineages is also instrumental for understanding the mechanisms of regenerative process and awaits clarification in forthcoming studies. Another concern to be considered is that haploinsufficiency in the knocked-in allele used in current genetic tracing assays was recently reported to result in a misleading phenotype as seen in pancreatic systems49.
The activity of ISCs is stringently controlled to ensure proper proliferation and differentiation. Tight regulation of ISCs is achieved primarily through extrinsic signaling molecules emanated from their surrounding cells that altogether constitute a unique niche microenvironment15,50. Recent intensive studies have led to gradual identification of vital niche components that include both epithelial progenies of ISCs and mesenchymal cells, such as Paneth cells, fibroblasts, immune cells, enteric neurons, and endothelial cells51,52. When perturbed by injurious factors, these intestinal niche cells can be rewired for coordinated production of cytokines and growth factors to activate ISCs for rapid regeneration. To date, various niche factor–derived signaling pathways have been identified to be essential for ISC activity, intestinal homeostasis, and regeneration. In this section, we will briefly summarize the central roles of Wnt, bone morphogenetic protein (BMP), Notch, epidermal growth factor (EGF), and Hippo signaling in ISC regulation.
In the Wnt pathway, binding of Wnt ligands to their Frizzled receptor on targeted cells induces collapse of cytoplasmic APC destruction complex and subsequent nuclear translocation of β-catenin to activate Wnt target genes through association with T-cell factor (TCF) transcription factors53. Wnt signaling is progressively reduced from crypt to villus axis and is indispensable for ISC maintenance and intestinal regeneration32,54,55. Its abrogation via inactivating Tcf1/2 mutation56, Tcf4 deletion57, or exogenous expression of Wnt inhibitor Dkk158,59 invariably leads to Lgr5 CBC loss and decreased crypt cell proliferation. Augmentation of activity of this pathway by Apc inactivating mutation, constitutive activating mutation in β-catenin, or simultaneous deletion of two E3 ligases (Rnf43 and Znrf3) targeting Wnt ligand receptors for degradation unanimously result in crypt expansion and rapid appearance of intestinal adenomas in mice60–64. Although the functional importance of Wnt signaling and its contribution to intestinal regulation have been well appreciated55, the cellular source for Wnt ligands remains incompletely understood. Various types of niche cells have been found to express a significant level of Wnt ligands, such as small intestinal Paneth cells51, colonic Reg4+65 or cKit+66 secretory cells, and numerous subsets of stromal cells67,68. Although epithelial Wnts can promote expansion of Lgr5 CBCs during in vitro organoid culture, their ablation does not demonstrate a notable impact on crypt cell proliferation and intestinal homeostasis in vivo, suggesting a significant redundancy of cellular origin of Wnt ligands in sustaining intestinal integrity67–70. In line with this assumption, recent studies found that Foxl1+71,72, Pdgfrα+73, Gli1+55,74, and CD34+ Gp38+ αSMA–75 mesenchymal cells serve as major sources of Wnt activity and play pivotal roles in sustaining intestinal homeostasis in vivo. Abolition of Wnt secretion in Foxl1+, Gli1+, or Pdgfrα+ cells by genetic excision of porcupine (Porcn) impairs proliferation of Lgr5 CBCs and leads to corruption of intestinal integrity71,73,74. Whether these mesenchymal cells with distinct markers overlap with each other to some extent or even represent the same subsets of non-epithelial cells is currently unclear. Moreover, how Wnt activity in these mesenchymal cells is regulated in response to intestinal injuries, such as inflammation and chemo-/radio-toxicity, remains unanswered. Future investigations combining genetic approaches with single-cell RNA-sequencing technology will address these critical questions.
BMP signaling assumes an increasing gradient along the intestinal crypt-villus axis and serves as a critical inducer for ISC differentiation, thereby playing a vital role in balancing the effect of Wnt signaling on intestinal homeostasis76,77. BMPs belong to the transforming growth factor-beta superfamily of ligands and can induce phosphorylation of cytoplasmic R-Smads (Smad1/5/8) through binding to the membrane-embedded serine/threonine kinase type I and II receptors. Phosphorylated R-Smads form a complex with Co-Smad (Smad4) and subsequently enter the nucleus, where they regulate expression of targeted genes through association with a variety of co-factors78,79. The initial experimental evidence implicating an inhibitory role of BMP signaling in ISC self-renewal comes from conditional deletion of BMP receptor Bmpr1a and transgenic expression of BMP antagonist Noggin in mice, both of which unanimously lead to development of multiple polyposis in small intestine80–82. Similarly, aberrant expression of BMP antagonist Gremlin1 was noted in patients with hereditary mixed polyposis syndrome and its transgenic expression in mice leads to the appearance of ectopic crypts and subsequent oncogenic transformation83,84. Gene expression and in situ hybridization analyses demonstrated that BMP ligands and BMP antagonists are produced primarily by mesenchymes residing at different regions of the crypt-villus unit. BMP2 and BMP4 are secreted by intravillus and intercrypt mesenchymal cells, whereas Noggin, Gremlin1, Gremlin2, and chordin-like 1 antagonists are expressed by cryptal myofibroblasts and smooth muscle cells beneath the crypt bottom80,85. To date, little is known about the role and cellular source of other BMP ligands in intestine and whether ablation of these niche cells has an impact on intestinal integrity and tumorigenesis. Future studies are needed to address questions in this regard.
Notch receptors are single-pass type I transmembrane heterodimer proteins that comprise functional extracellular truncation, transmembrane, and intracellular (NICD) domain. Activation of Notch receptors begins with their binding to Notch ligands presented on an adjacent cell, which leads to cytoplasmic proteolytic cleavage of Notch receptors by γ-secretase complex and subsequent nuclear translocation of intracellular NICD to regulate transcription of target genes86. Notch signaling has been shown to contribute to Lgr5 CBC maintenance and proliferation87–89 and its inhibition skews intestinal differentiation toward goblet lineage90–92. Paneth cells express and present Notch ligands Dll1 and Dll4 to adjacent Lgr5 CBCs to facilitate their self-renewal51. Moreover, activation of Notch signaling in Paneth cells by forced expression of NICD can result in acquisition of a stem-like property in these cells46. Notch signaling is also involved in controlling secretory and absorptive lineage determination when CBCs start to differentiate. Its activation stimulates Hes1 expression for transcriptional silencing of Atoh1, a master transcription factor for secretory lineage determination, thereby blocking secretory differentiation but promoting enterocyte specification of Notch receptor activated cells93. Expression of Notch ligand Dll1 on secretory cells prevents a secretory fate of neighboring cells but drives these cells toward absorptive lineage through Notch signaling activation, which is a biological event referred to as lateral inhibition88. Although Notch signaling contributes to CBC proliferation and intestinal homeostasis, the role of niche cells expressing Notch ligands remains unclear.
The Hippo pathway functions as a vital regulator of tissue homeostasis and organ size and its deregulation is implicated in the development of multiple types of cancer in humans. In the presence of extracellular stimuli, transcriptional activator YAP/TAZ complex is phosphorylated by upstream serine–threonine kinase MST1/MST2/Sav1-LATS1/2/MOB1A/B cascade, leading to either cytoplasmic sequestration or proteasomal degradation of YAP/TAZ. Without Hippo activation, non-phosphorylated YAP/TAZ enters the nucleus and acts as a co-activator for the TEAD transcription factor to regulate expression of genes associated with cell growth and proliferation94,95. The Hippo pathway is constitutively active in intestine under homeostatic conditions and its genetic inactivation via transgenic expression of YAP/TAZ or conditional deletion of Sav1, an upstream negative regulator of YAP, increases ISC proliferation and leads to crypt hyperplasia96,97. Hippo signaling is also indispensable for intestinal regeneration in mouse models of dextran sodium sulfate (DSS)-induced colitis and radiation-induced injury97,98. Transgenic expression of YAP induces an EGF signaling–dependent regenerative program to facilitate intestinal regeneration98. Appropriate activation of Wnt signaling is essential for intestinal recovery following injury. However, its aberrant activation could lead to oncogenic transformation or reduce ISC survival through increasing radio-sensitivity of ISCs to DNA damage during intestinal regeneration99. In addition to the transcriptional regulatory activity of nuclear YAP/TAP complex, unphosphorylated cytoplasmic complexes are integral components of β-catenin destruction complex and thus play a critical role in restricting Wnt signaling activity100. Paradoxically, it has been noted that conditional deletion of YAP in intestinal epithelia augments Wnt activity and causes crypt hyperplasia and overgrowth throughout small intestine and colon after radiation-induced injury101. These findings suggest that the Hippo pathway has dual roles in regulating both Wnt and Egf signaling and its impact on intestinal regeneration is context-dependent and could be determined by net activity of these two pathways. More work is needed to deepen our understanding of the molecular mechanisms that control the balanced activation of Wnt and Egf pathways by Hippo signaling during intestinal regeneration.
EGF communicates with target cells through the EGF receptor, which is a member of the ErbB family of tyrosine receptor kinases102. Paneth cells express EGF and sustain proliferation of Lgr5 CBCs through its receptor ErbB103. EGF supplementation of culture medium dramatically increases the efficiency of organoid formation in vitro17. Lrig1 is a negative feedback regulator of ErbB receptor and its ablation causes crypt expansion in mice29,104. Although these findings support a crucial role of EGF signaling in CBC proliferation, the additional cellular source of EGF growth factor except for Paneth cells in intestine remains ambiguous.
Intestinal homeostasis and regeneration are accompanied by drastic transcriptional alterations that are achieved through the cooperation between extrinsic niche signaling–controlled transcription factors/co-activators and intrinsic epigenetic regulators. Epigenetics refers to the inherited alteration in gene expression and phenotype that occur without changes in DNA sequence. It consists primarily of DNA methylation, histone modification, and chromatin remodeling105,106. In contrast to the extensively studied extrinsic niche factors, the role of intrinsically epigenetic mechanisms in intestinal homeostasis and regeneration is poorly understood. In this section, we will briefly discuss the currently limited knowledge of epigenetic mechanisms in intestinal regulation.
With the tremendous advancements in fluorescence-activated cell sorting, identification of lineage-specific markers, and next-generation sequencing technology, epigenetic characterization of distinct types of intestinal epithelial cells has recently become feasible. Initial studies with DNase I mapping for accessible chromatin regions and H3K4me2 and H3K27ac ChIP-seq (chromatin immunoprecipitation sequencing) for active enhancers showed that ISCs and enterocyte and secretory progenitors assume remarkable similarity in distribution pattern of open chromatin elements and active enhancers throughout the genome107. However, these observations are not in line with the subsequent studies that employ assay for transposase-accessible chromatin using sequencing (ATAC-seq) to profile open chromatin regions among ISCs and enterocyte and secretory progenitors. The latter studies identified thousands of unique open chromatin regions among these populations and found that accessible chromatin states in secretory progenitors shift to resemble ISCs during dedifferentiation upon injuries (Figure 2)7. Although the discrepancy between the two studies can be explained by cellular heterogeneity and the different sensitivity of technologies employed, the functional importance of those unique open chromatin elements in secretory lineage specification, dedifferentiation, and regeneration has not been determined experimentally. In addition, comprehensive profiling of distinct histone modifications in purer populations of intestinal lineages will determine whether the epigenetic landscape varies during intestinal homeostasis and regeneration.
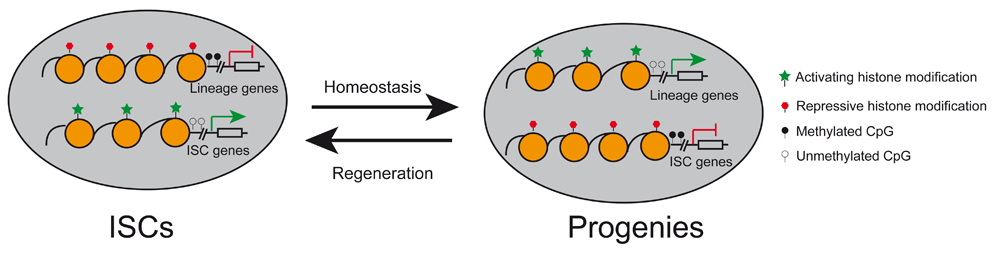
During homeostasis, intestinal stem cells (ISCs) adopt a unique epigenetic signature and chromatin accessibility that collaboratively lead to expression of ISC-related genes and shutdown of lineage-specific factors. Lineage specification of ISCs is accompanied by epigenetic remodeling and chromatin accessibility changes that turn off ISCs genes while activating lineage-restricted genes. During the regenerative process, epigenetic landscape and chromatin accessibility are reconfigured to resemble ISCs during dedifferentiation of ISC progenies.
Initial bisulfite-sequencing of ISCs and their differentiated progenies in villus revealed minimal dynamics of DNA methylome during intestinal differentiation108. This study used a cutoff of a minimal differential methylation of 40% to identify differentially methylated regions (DMRs) between ISCs and villous cells. As the distal regulatory regions are low-methylated at an overall methylation level of 30% in intestinal cells and display average change of about 15% in DNA methylation during intestinal differentiation, the cutoff criteria of analysis used in the study eliminates the identification of DMRs during differentiation of ISCs108,109. Subsequent bisulfite-sequencing studies with distinct analytic standards identified many DMRs at enhancers and disclosed a tight correlation between the DMRs and transcriptional alterations between two populations of cells, implying that the DNA methylation may have a vital role in intestinal renewal110. This viewpoint has been proven by genetic ablation of Dnmt1 and Tet1 in intestine, which are maintenance methyltransferase and hydroxylase for DNA methylation, respectively106. Dnmt1 is essential for intestinal development in newborn mice as inducible deletion of Dnmt1 at perinatal stage causes genomic instability, premature differentiation, apoptosis, loss of villi, and decreased proliferation of crypt cells111,112. The essential role of DNA methylation for intestinal development is also reflected in Tet1-deleted mice. Tet1-null mice exhibit retarded growth, shorter intestines, weaker capability to form in vitro organoids, and reduced postnatal viability113. Acute ablation of Dnmt1 in adult intestinal epithelia causes a slight expansion of the proliferative crypt zone. However the crypt morphology recovers and DNA methylation restores to normal level several days after Dnmt1 deficiency. A subsequent study revealed that de novo methyltransferase Dnmt3b can compensate for Dnmt1 loss to maintain intestinal integrity as ablation of both enzymes leads to genome demethylation, genomic instability, increased apoptosis, and decreased survival114. Taken together, these pieces of genetic evidence strongly support that dynamic regulation of DNA methylation acts as an essential epigenetic mechanism underlying intestinal development and homeostasis.
Post-translational modification of histones, such as methylation and ubiquitination, constitutes an additional component of epigenetic mechanism in transcriptional regulation, embryonic development, and adult tissue homeostasis115. Monoubiquitination of lysine 119 on H2A (H2A119 mUb) and methylation of lysine 27 on H3 (H3K27me) correlate with transcriptional repression and are implemented by polycomb repressive complex 1 (PRC1) and polycomb repressive complex 2 (PRC2), respectively116. The activity of PRC1 and PRC2 stems from Ring1a/1b E3 ligase and Ezh1/2 methyltransferases and has been shown to be essential for normal intestinal homeostasis or regeneration following injuries. Loss of function of the total PRC1 activity via Ring1a/b double deletion in intestinal epithelia compromises ISC self-renewal and intestinal integrity and results in morbidity through de-repression of a number of transcription factors that negatively regulate Wnt signaling117.
Loss of PRC2 activity via AhCre or VillinCreER-induced epithelial deletion of the Eed, a scaffold protein of the complex, leads to a clear defect in cell proliferation in crypts, a marked increase in the number of goblet cells, mislocalized Paneth cells, and compromised regenerative capacity118–120. The homeostatic and regenerative defect of intestine in the absence of PRC2 results at least in part from aberrant upregulation of Cdkn2a and some master regulators for secretory lineages that are bivalently marked, normally targeted, and repressed by this complex118. Conditional ablation of catalytic subunit of PRC2 complex Ezh2 does not show any abnormalities in the intestinal homeostasis, indicating that the other H3K27 methyltransferase Ezh1 could compensate for Ezh2 loss to maintain intestinal integrity120. Further studies are needed to know the role and target genes of individual non-canonic PRC1 and PRC2 complexes in intestinal renewal and regeneration.
Over the past decade, remarkable progress has been made in our understanding of intestinal biology. It has become evident that active Lgr5 CBCs and reserve ISCs work in a coordinated manner to maintain intestinal homeostasis and replenish the tissue upon injuries. In addition, differentiated epithelial cells possess considerable plasticity and can dedifferentiate to ISCs for intestinal regeneration in mouse models when active CBCs are damaged or artificially removed. Many niche-derived factors and a few epigenetic regulators have been identified and characterized for their roles in intestinal morphogenesis, homeostasis, and regeneration. Despite these considerable advances in recent years, several fundamental questions remain to be addressed about the precise molecular mechanisms controlling self-renewal, lineage commitment, and plasticity of ISCs. First, the cellular source of distinct environmental factors crucial for ISC function and the role of immune cells, peripheral nerve cells, and other ISC-proximal cells remain poorly understood; thus, extensive studies of these niche components will offer more insight into the extrinsic regulatory mechanisms for ISC function. Second, comprehensive characterization of the epigenetic landscape of various intestinal cells coupled with conditional ablation of key chromatin regulators in murine models will help determine the molecular basis underlying the remarkable plasticity of intestine in response to injuries. Third, the physiological function and contribution of differentiated crypt cells to radio-/chemo-induced intestinal regeneration remain to be experimentally evaluated. Fourth, the molecular mechanisms that reserve ISCs and differentiated crypt cells use to sense and replenish lost CBCs are largely unexplored; future investigation in this regard will help design effective therapeutic approaches to facilitate intestinal recovery after damage. Lastly, does aging-associated loss of regenerative potential of ISCs result from alterations in niche factors or epigenetic regulation or both121–123? The more we understand the above fundamental questions, the better we can employ ISCs for regenerative medicine.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)