Serpico AF and Grieco D. Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.12688/f1000research.21185.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1CEINGE, Naples, 80145, Italy 2DMMBM, University of Naples Federico II, Naples, 80131, Italy 3Department of Pharmacy, University of Naples Federico II, Naples, 80131, Italy
The goal of mitosis is to form two daughter cells each containing one copy of each mother cell chromosome, replicated in the previous S phase. To achieve this, sister chromatids held together back-to-back at their primary constriction, the centromere, have to interact with microtubules of the mitotic spindle so that each chromatid takes connections with microtubules emanating from opposite spindle poles (we will refer to this condition as bipolar attachment). Only once all replicated chromosomes have reached bipolar attachments can sister chromatids lose cohesion with each other, at the onset of anaphase, and move toward opposite spindle poles, being segregated into what will soon become the daughter cell nucleus. Prevention of errors in chromosome segregation is granted by a safeguard mechanism called Spindle Assembly Checkpoint (SAC). Until all chromosomes are bipolarly oriented at the equator of the mitotic spindle, the SAC prevents loss of sister chromatid cohesion, thus anaphase onset, and maintains the mitotic state by inhibiting inactivation of the major M phase promoting kinase, the cyclin B-cdk1 complex (Cdk1). Here, we review recent mechanistic insights about the circuitry that links Cdk1 to the SAC to ensure correct achievement of the goal of mitosis.
Corresponding author:
Domenico Grieco
Competing interests:
No competing interests were disclosed.
Grant information:
This work was supported by a grant from AIRC (IG grant 2017; Id. 19851 to DG).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Maintenance of genome stability through cell generations is a crucial feature that grants health to cells, organs and organisms. In humans, genome instability is causally linked to pathological outcomes such as cancer, degenerative disorders and physical and mental retardation1–3. Cells have developed several mechanisms to surveil that each step required for cell division is healthy and thoroughly completed before passing to the next one. This is achieved through mechanisms called cell cycle checkpoints4–8. If cells experience DNA damage or sense that DNA replication or assembly of the mitotic spindle is incomplete, checkpoint mechanisms halt cell cycle progression to repair damage or complete previous cell cycle stages before moving forward in their division process. If repair or completion is frustrated, then healthy checkpoints promote cell death9–13. This short review will be focused on recent advancements in the mechanistic understanding of the Spindle Assembly Checkpoint (SAC), the checkpoint that prevents formation of cells with an abnormal chromosome number by delaying mitosis exit until bipolar attachment of all replicated chromosomes14.
Progression through mitosis: a cycle of Cdk1 activation/inactivation
Progression through mitosis is granted by a wave of cyclin B-cdk1 complex (Cdk1) activity15,16. Cdk1 is activated at the onset of mitosis by reversal of inhibitory phosphorylations of the cdk1 moiety at threonine 14 and tyrosine 15. These phosphorylations, operated by the Myt1 and Wee1 kinases, allow accumulation of enough inactive Cdk1, during S phase and G2, to rapidly induce mitosis upon their reversal17,18. Dephosphorylation and activation of Cdk1 are granted by the dual-specificity phosphatase Cdc2519. Upon initial activation, Cdk1 phosphorylates and inhibits Myt1 and Wee1 while it phosphorylates and further activates Cdc25; this way, Cdk1 promotes positive feedback loops for its own activation20–22. For mitosis onset, Cdk1 activity also represses major phosphatase activities (like that of PP1 and PP2A) that otherwise would antagonize Cdk1 action. The catalytic activity of PP1 is directly inhibited by Cdk1-dependent phosphorylation, while the activity of PP2A in which B55 is the holoenzyme regulatory subunit, PP2A-B55, is kept inhibited in mitosis by the aid of Greatwall kinase (Gwl). Gwl is stimulated by Cdk1 and phosphorylates Ensa/Arpp19, two small molecules, transforming them into potent PP2A-B55 inhibitors22.
Inactivation of Cdk1 at the end of mitosis instead depends on the ubiquitin-dependent degradation of cyclin B14,23–25. This is initiated by the ubiquitin ligase Anaphase Promoting Complex/Cyclosome (APC/C) in association with its coactivator Cdc20. APC/CCdc20 also promotes the degradation of securin, an inhibitor of separase, the protease that cleaves the protein bridge that holds sister chromatid centromeres together14,26–28. This way, the onset of anaphase and Cdk1 inactivation are tightly coupled by this irreversible degradative mechanism. Initial evidence indicated that APC/CCdc20 activity required Cdk1-dependent phosphorylation; recently, the APC/C members that are directly phosphorylated by Cdk1 were identified29–33. Thus, Cdk1 is also promoting a negative feedback for its own inactivation. Nevertheless, final APC/CCdc20 activation is under the control of the SAC, which inhibits APC/CCdc20 until bipolar attachment of all replicated chromosomes14.
Mps1 and the SAC, in brief
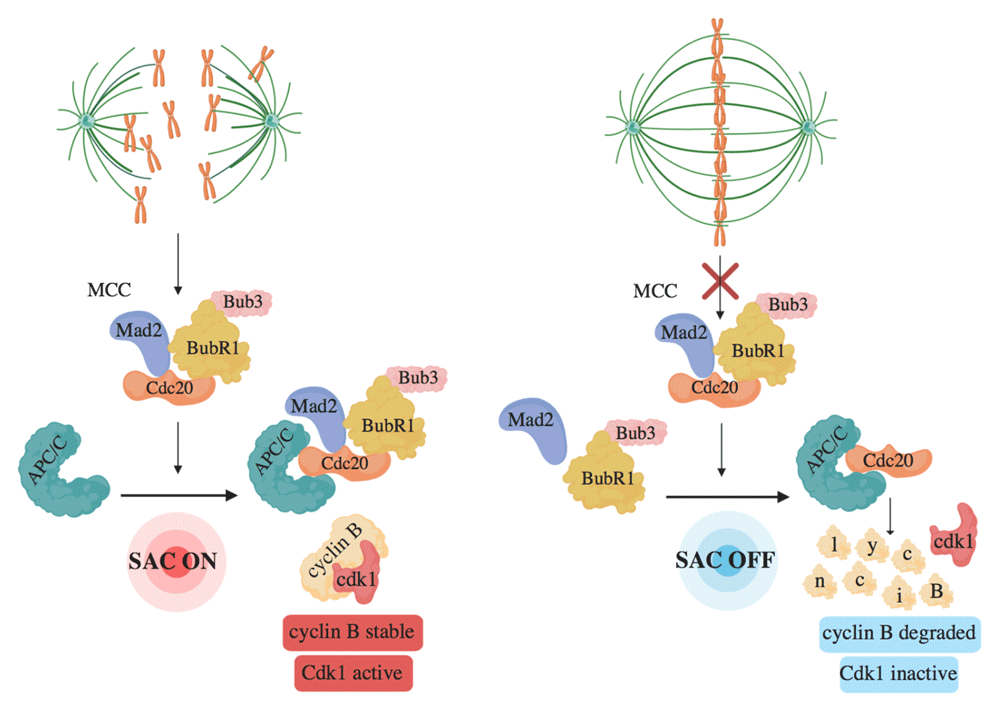
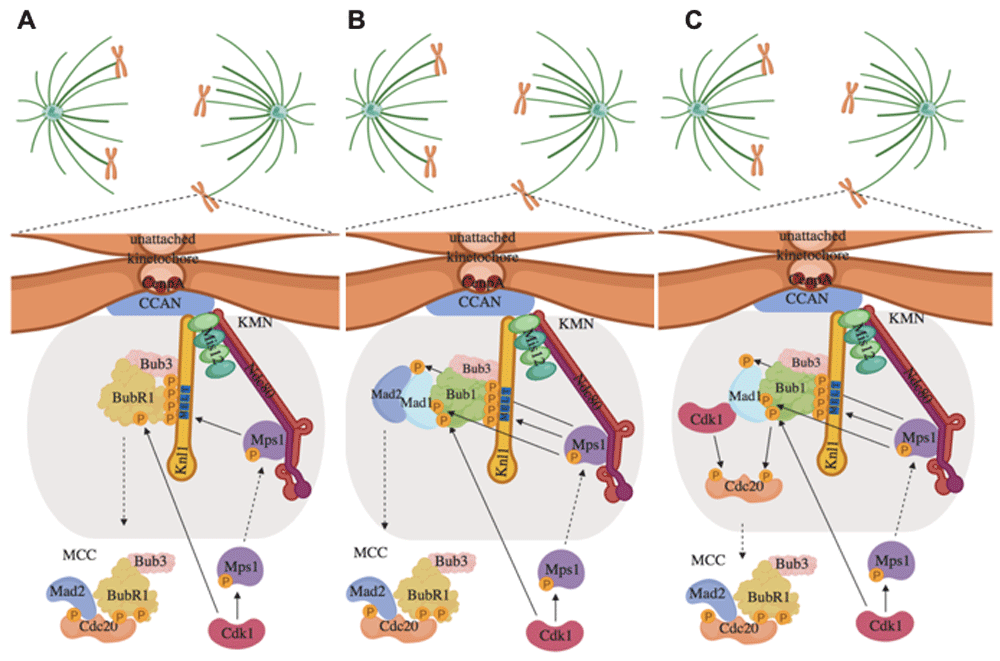
The SAC inhibits APC/CCdc20 activation by forming a diffusible Mitotic Checkpoint Complex (MCC), composed of the proteins Mad2, Bub3, BubR1, and Cdc20 itself, in which Cdc20 is restrained from activating APC/C14,34–37. MCC forms at unattached kinetochores, proteinaceous centromeric structures deputed to interact with spindle microtubules and permit chromosome segregation (Figure 1)14. MCC formation requires the action of crucial SAC kinases like Plk1, Aurora B, and Mps138–40. These kinases also have important roles in correcting faulty chromosome–microtubule interactions to promote correct, end-on, bipolar chromosome–microtubule attachments41. Here, however, we will primarily review recent advancements in the regulation of Mps1 in SAC control and its dependence on Cdk1 activity. Mps1 binds unattached kinetochores where it phosphorylates SAC proteins and activates them and then gets released from kinetochores upon stable microtubule binding, perhaps by competition mechanisms42–46. The bridge deputed to connect centromeres to microtubules is called the KMN network and is composed by the Knl1 complex, the Mis12 complex, and the Ndc80 complex46–50. The KMN, in the outer kinetochore, interacts with the inner kinetochore Constitutive Centromere Associated Network (CCAN), a protein network that assembles onto Cenp-A nucleosomes, a histone H3 variant found at centromeric nucleosomes51–53. Mps1 localizes at unattached kinetochores primarily by interacting with the Ndc80 complex54. At kinetochores, Mps1 phosphorylates the “MELT” repeats of Knl1, promoting kinetochore recruitment of the BubR1-Bub3 and Bub1-Bub3 complexes (Figure 2), while Knl1 dephosphorylation by PP1 appears involved in SAC silencing43,55–58. Mps1 also phosphorylates Bub1, further promoting kinetochore recruitment of Mad1, another crucial SAC protein needed for the activation of Mad259–61. Mad1 recruitment at kinetochores is also facilitated by the Rod-Zwilch-ZW10 (RZZ) complex62. In addition, phosphorylation of Mad1 by Mps1 helps the Mad1-dependent conversion of Mad2 into the functional conformation required to inhibit Cdc20 in the MCC59,60. Thus, Mps1 is a crucial effector of the SAC mechanism by promoting MCC formation (Figure 2).
Figure 1. Unattached or incorrectly attached chromosomes promote formation of the Mitotic Checkpoint Complex (MCC).
Until bipolar spindle assembly, the MCC, composed of Mad2, BubR1-Bub3, and Cdc20, forms, binds, and blocks APC/C action (SAC ON). Upon bipolar spindle assembly, MCC is dismantled and MCC-free Cdc20 activates APC/C (SAC OFF).
Figure 2. Paths to Mitotic Checkpoint Complex (MCC) formation.
Cdk1 phosphorylation of Mps1 helps kinetochore recruitment of Mps1 to (A) recruit BubR1-Bub3 complex for its incorporation into MCC, (B) recruit Bub1-Bub3 for Mad1-Mad2 docking and Mad2 incorporation into MCC, and (C) recruit Bub1-Bub3 for Mad1-Cdk1 docking for Bub1- and Cdk1-dependent phosphorylation of Cdc20 and incorporation into MCC.
Cdk1 and the SAC
The observation that APC/C activity was promoted by Cdk1-dependent phosphorylation, while APC/C activation was inhibited by the SAC until spindle assembly, reinforced the idea that checkpoint mechanisms would oppose the forward trend of the basic cell cycle engine4. However, in 2003, a few independent observations, from yeast and vertebrates, changed this view by showing that Cdk1 was instrumental to the SAC action63–65. Indeed, in yeast, SAC-defective cdk1 mutants were described and Bub1 was shown to be phosphorylated by Cdk1 for SAC proficiency63,64. In the Xenopus egg extract system and in human somatic cells, Cdk1 activity was revealed to be required to sustain SAC-dependent arrest and the ability of MCC members to block the APC/C65,66. Cdk1-dependent phosphorylation of Cdc20 appeared to have a role in reducing Cdc20 affinity for APC/C while increasing that for other MCC proteins65–68. Thus, Cdk1, the cell cycle engine, though paving the way for its own inactivation by phosphorylating APC/C, was instrumental for the checkpoint SAC that would block APC/C activation until correct spindle assembly66. These observations also helped to explain why the SAC does not get reactivated at the onset of anaphase, when loss of chromatid cohesion causes loss of kinetochore tension, a condition that would have activated the SAC at earlier stages69,70. This was shown to be due to the concomitant reduction of Cdk1 because of the mentioned coupling of anaphase onset with degradation of cyclin B69,70. A few years later, the notion that Cdk1 was required for the SAC function was reinforced by the findings that, in the Xenopus egg extract system, Mps1 was phosphorylated by Cdk1 and that this phosphorylation substantially helped Mps1 activity in its fundamental role for the SAC71.
Very recently, through careful biochemical dissection, important observations have described in closer detail how Cdk1 is an integral part of the SAC mechanisms72,73. Indeed, it has been shown that kinetochore localization of Mps1, in human cells, greatly depends on direct phosphorylation by Cdk1; thus, Cdk1 controls activity and localization of Mps172. Mps1, in turn, helps kinetochore localization of Cdk173–76. As mentioned earlier, by phosphorylating Knl1, Mps1 creates a docking site for kinetochore localization of Bub1, and cooperative Cdk1- and Mps1-dependent phosphorylations of Bub1 are required to recruit Mad1 at kinetochores42,43,50,59,77. Kinetochore localization of Mad1 is crucial for its ability to convert Mad2 in the effective form that incorporates into the MCC61. However, it has also recently been shown that Mad1 stably interacts with Cdk1 and that Mps1, through kinetochore recruitment of Mad1, in turn, promotes kinetochore localization of Cdk1 (Figure 2)72,73. At kinetochores, Cdk1 may further phosphorylate other substrates to sustain the SAC like Cdc20 or BubR1 and possibly also help error correction and SAC resolution by favoring BubR1 interaction with the protein phosphatase PP2A-B5665,78–80. Recent evidence also indicated how the indirect downregulation of the protein phosphatase PP2A-B55 activity by Cdk1 is instrumental for the SAC-promoting action of Cdk1 itself72,81. In addition, it should be noted that kinetochore localization of Mps1 is favored by the activity of Aurora B, perhaps by phosphorylating members of the Ndc80 complex42. However, centromere localization of Aurora B depends on other components of the Chromosomal Passenger Complex (CPC), composed of survivin, borealin, INCENP, and Aurora B itself, and Cdk1 activity is required, directly and indirectly, for CPC centromeric localization82–84. Thus, even by mastering CPC localization, Cdk1 affects Mps1 and is fundamental for SAC action.
Concluding remarks and further questions
The recent advancements, reviewed here, in the mechanisms of mitotic exit and in particular in how Cdk1 mechanistically serves the SAC, suggest that Cdk1 is an integral part of the SAC system. Thus, perhaps the cell cycle engine, Cdk1, and the checkpoint, SAC, are not to be viewed any longer as separate mechanisms but rather as integrated systems that ensure correct execution of complex biological tasks. Important hints have also been recently provided on how the SAC can be silenced, such as on priming mechanisms for protein phosphatases that would reverse SAC-activating phosphorylations upon bipolar chromosome attachments, in addition to the notion that the MCC itself undergoes proteasome-dependent turnover for rapid SAC silencing79,84–88. Nevertheless, major phosphatases like PP1 and PP2A are directly or indirectly inhibited by Cdk1 activity22. Thus, it is still unclear whether chromosome attachment and kinetochore tension are sufficient to dislodge kinases and let phosphatases take the upper hand for SAC silencing or whether these conditions also affect the activity of crucial SAC kinases40. Based on our previous observations, we hypothesize in this regard that Cdk1 activity could be locally downregulated by non-proteolytic means upon bipolar chromosome attachment and that this would lead to SAC silencing89–92. If this were true, a proteolysis-independent negative control of Cdk1 would be required for SAC silencing, ahead of and for final, proteolysis-dependent, Cdk1 inactivation and mitotic exit.
The authors acknowledge Associazione Italiana per la Ricerca sul Cancro (AIRC) for support and all the relevant works on this topic apologizing for not having cited them all.
3.
de Wolf B, Kops GJPL:
Kinetochore Malfunction in Human Pathologies.
Adv Exp Med Biol.
2017; 1002: 69–91. PubMed Abstract
| Publisher Full Text
4.
Hartwell LH, Weinert TA:
Checkpoints: controls that ensure the order of cell cycle events.
Science.
1989; 246(4930): 629–34. PubMed Abstract
| Publisher Full Text
5.
Murray AW:
Creative blocks: cell-cycle checkpoints and feedback controls.
Nature.
1992; 359(6396): 599–604. PubMed Abstract
| Publisher Full Text
10.
Sorger PK, Dobles M, Tournebize R, et al.:
Coupling cell division and cell death to microtubule dynamics.
Curr Opin Cell Biol.
1997; 9(6): 807–14. PubMed Abstract
| Publisher Full Text
11.
Jacotot E, Ferri KF, Kroemer G:
Apoptosis and cell cycle: distinct checkpoints with overlapping upstream control.
Pathol Biol (Paris).
2000; 48(3): 271–9. PubMed Abstract
12.
Clarke PR, Allan LA:
Cell-cycle control in the face of damage--a matter of life or death.
Trends Cell Biol.
2009; 19(3): 89–98. PubMed Abstract
| Publisher Full Text
14.
Musacchio A, Salmon ED:
The spindle-assembly checkpoint in space and time.
Nat Rev Mol Cell Biol.
2007; 8(5): 379–93. PubMed Abstract
| Publisher Full Text
16.
Minshull J, Pines J, Golsteyn R, et al.:
The role of cyclin synthesis, modification and destruction in the control of cell division.
J Cell Sci.
1989; 12: 77–97. PubMed Abstract
| Publisher Full Text
17.
Atherton-Fessler S, Hannig G, Piwnica-Worms H:
Reversible tyrosine phosphorylation and cell cycle control.
Semin Cell Biol.
1993; 4(6): 433–42. PubMed Abstract
| Publisher Full Text
19.
Perdiguero E, Nebreda AR:
Regulation of Cdc25C activity during the meiotic G2/M transition.
Cell Cycle.
2014; 3(6): 733–7. PubMed Abstract
| Publisher Full Text
21.
Domingo-Sananes MR, Kapuy O, Hunt T, et al.:
Switches and latches: A biochemical tug-of-war between the kinases and phosphatases that control mitosis.
Philos Trans R Soc Lond B Biol Sci.
2011; 366(1584): 3584–94. PubMed Abstract
| Publisher Full Text
| Free Full Text
22.
Hunt T:
On the regulation of protein phosphatase 2A and its role in controlling entry into and exit from mitosis.
Adv Biol Regul.
2013; 53(2): 173–8. PubMed Abstract
| Publisher Full Text
23.
Evans T, Rosenthal ET, Youngblom J, et al.:
Cyclin: a protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division.
Cell.
1983; 33(2): 389–96. PubMed Abstract
| Publisher Full Text
24.
Murray AW, Solomon MJ, Kirschner MW:
The role of cyclin synthesis and degradation in the control of maturation promoting factor activity.
Nature.
1989; 339(6222): 280–6. PubMed Abstract
| Publisher Full Text
25.
King RW, Peters JM, Tugendreich S, et al.:
A 20S complex containing CDC27 and CDC16 catalyzes the mitosis-specific conjugation of ubiquitin to cyclin B.
Cell.
1995; 81(2): 279–88. PubMed Abstract
| Publisher Full Text
26.
Michaelis C, Ciosk R, Nasmyth K:
Cohesins: chromosomal proteins that prevent premature separation of sister chromatids.
Cell.
1997; 91(1): 35–45. PubMed Abstract
| Publisher Full Text
27.
Ciosk R, Zachariae W, Michaelis C, et al.:
An ESP1/PDS1 complex regulates loss of sister chromatid cohesion at the metaphase to anaphase transition in yeast.
Cell.
1998; 93(6): 1067–76. PubMed Abstract
| Publisher Full Text
29.
Shteinberg M, Protopopov Y, Listovsky T, et al.:
Phosphorylation of the cyclosome is required for its stimulation by Fizzy/cdc20.
Biochem Biophys Res Commun.
1999; 260(1): 193–8. PubMed Abstract
| Publisher Full Text
30.
Rudner AD, Murray AW:
Phosphorylation by Cdc28 activates the Cdc20-dependent activity of the anaphase-promoting complex.
J Cell Biol.
2000; 149(7): 1377–90. PubMed Abstract
| Publisher Full Text
| Free Full Text
38.
Nigg EA:
Mitotic kinases as regulators of cell division and its checkpoints.
Nat Rev Mol Cell Biol.
2001; 2(1): 21–32. PubMed Abstract
| Publisher Full Text
39.
Suijkerbuijk SJ, Kops GJ:
Preventing aneuploidy: the contribution of mitotic checkpoint proteins.
Biochim Biophys Acta.
2008; 1786(1): 24–31. PubMed Abstract
| Publisher Full Text
42.
Santaguida S, Tighe A, D'Alise AM, et al.:
Dissecting the role of MPS1 in chromosome biorientation and the spindle checkpoint through the small molecule inhibitor reversine.
J Cell Biol.
2010; 190(1): 73–87. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Yamagishi Y, Yang CH, Tanno Y, et al.:
MPS1/Mph1 phosphorylates the kinetochore protein KNL1/Spc7 to recruit SAC components.
Nat Cell Biol.
2012; 14(7): 746–52. PubMed Abstract
| Publisher Full Text
44.
Hiruma Y, Sacristan C, Pachis ST, et al.:
CELL DIVISION CYCLE. Competition between MPS1 and microtubules at kinetochores regulates spindle checkpoint signaling.
Science.
2015; 348(6240): 1264–7. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
45.
Ji Z, Gao H, Yu H:
CELL DIVISION CYCLE. Kinetochore attachment sensed by competitive Mps1 and microtubule binding to Ndc80C.
Science.
2015; 348(6240): 1260–4. PubMed Abstract
| Publisher Full Text
47.
Varma D, Wan X, Cheerambathur D, et al.:
Spindle assembly checkpoint proteins are positioned close to core microtubule attachment sites at kinetochores.
J Cell Biol.
2013; 202(5): 735–46. PubMed Abstract
| Publisher Full Text
| Free Full Text
48.
Petrovic A, Keller J, Liu Y, et al.:
Structure of the MIS12 Complex and Molecular Basis of Its Interaction with CENP-C at Human Kinetochores.
Cell.
2016; 167(4): 1028–1040.e15. PubMed Abstract
| Publisher Full Text
| Free Full Text
49.
Umbreit NT, Gestaut DR, Tien JF, et al.:
The Ndc80 kinetochore complex directly modulates microtubule dynamics.
Proc Natl Acad Sci U S A.
2012; 109(40): 16113–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
51.
Hori T, Amano M, Suzuki A, et al.:
CCAN makes multiple contacts with centromeric DNA to provide distinct pathways to the outer kinetochore.
Cell.
2008; 135(6): 1039–52. PubMed Abstract
| Publisher Full Text
53.
Hori T, Shang WH, Takeuchi K, et al.:
The CCAN recruits CENP-A to the centromere and forms the structural core for kinetochore assembly.
J Cell Biol.
2013; 200(1): 45–60. PubMed Abstract
| Publisher Full Text
| Free Full Text
54.
Stucke VM, Baumann C, Nigg EA:
Kinetochore localization and microtubule interaction of the human spindle checkpoint kinase Mps1.
Chromosoma.
2004; 113(1): 1–15. PubMed Abstract
| Publisher Full Text
56.
Primorac I, Weir JR, Chiroli E, et al.:
Bub3 reads phosphorylated MELT repeats to promote spindle assembly checkpoint signaling.
eLife.
2013; 2: e01030. PubMed Abstract
| Publisher Full Text
| Free Full Text
57.
Zhang G, Mendez BL, Sedgwick GG, et al.:
Two functionally distinct kinetochore pools of BubR1 ensure accurate chromosome segregation.
Nat Commun.
2016; 7: 12256. PubMed Abstract
| Publisher Full Text
| Free Full Text
62.
Caldas GV, Lynch TR, Anderson R, et al.:
The RZZ complex requires the N-terminus of KNL1 to mediate optimal Mad1 kinetochore localization in human cells.
Open Biol.
2015; 5(11): pii: 150160. PubMed Abstract
| Publisher Full Text
| Free Full Text
63.
Kitazono AA, Garza DA, Kron SJ:
Mutations in the yeast cyclin-dependent kinase Cdc28 reveal a role in the spindle assembly checkpoint.
Mol Genet Genomics.
2003; 269(5): 672–84. PubMed Abstract
| Publisher Full Text
64.
Yamaguchi S, Decottignies A, Nurse P:
Function of Cdc2p-dependent Bub1p phosphorylation and Bub1p kinase activity in the mitotic and meiotic spindle checkpoint.
EMBO J.
2003; 22(5): 1075–87. PubMed Abstract
| Publisher Full Text
| Free Full Text
66.
D'Angiolella V, Grieco D:
Attach first, then detach: a role for cyclin B-dependent kinase 1 in coordinating proteolysis with spindle assembly.
Cell Cycle.
2004; 3(2): 132–3. PubMed Abstract
| Publisher Full Text
68.
Labit H, Fujimitsu K, Bayin NS, et al.:
Dephosphorylation of Cdc20 is required for its C-box-dependent activation of the APC/C.
EMBO J.
2012; 31(15): 3351–62. PubMed Abstract
| Publisher Full Text
| Free Full Text
69.
Rattani A, Vinod PK, Godwin J, et al.:
Dependency of the spindle assembly checkpoint on Cdk1 renders the anaphase transition irreversible.
Curr Biol.
2014; 24(6): 630–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
70.
Vázquez-Novelle MD, Sansregret L, Dick AE, et al.:
Cdk1 inactivation terminates mitotic checkpoint surveillance and stabilizes kinetochore attachments in anaphase.
Curr Biol.
2014; 24(6): 638–45. PubMed Abstract
| Publisher Full Text
| Free Full Text
75.
Allan LA, Reis M, Liu Y, et al.:
Cyclin B1 scaffolds MAD1 at the corona to activate the spindle assembly checkpoint.
BioRxiv.
2019; 726224. Publisher Full Text
76.
Jackman M, Marcozzi C, Pardo M, et al.:
Cyclin B1-Cdk1 binding to MAD1 links nuclear pore disassembly to chromosomal stability.
BioRxiv.
2019; 701474. Publisher Full Text
78.
Wong OK, Fang G:
Cdk1 phosphorylation of BubR1 controls spindle checkpoint arrest and Plk1-mediated formation of the 3F3/2 epitope.
J Cell Biol.
2007; 179(4): 611–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
79.
Kruse T, Zhang G, Larsen MS, et al.:
Direct binding between BubR1 and B56-PP2A phosphatase complexes regulate mitotic progression.
J Cell Sci.
2013; 126(Pt 5): 1086–92. PubMed Abstract
| Publisher Full Text
81.
Diril MK, Bisteau X, Kitagawa M, et al.:
Loss of the Greatwall Kinase Weakens the Spindle Assembly Checkpoint.
PLoS Genet.
2016; 12(9): e1006310. PubMed Abstract
| Publisher Full Text
| Free Full Text
82.
Ruchaud S, Carmena M, Earnshaw WC:
Chromosomal passengers: conducting cell division.
Nat Rev Mol Cell Biol.
2007; 8(10): 798–812. PubMed Abstract
| Publisher Full Text
87.
Visconti R, Palazzo L, Grieco D:
Requirement for proteolysis in spindle assembly checkpoint silencing.
Cell Cycle.
2014; 9(3): 564–9. PubMed Abstract
| Publisher Full Text
88.
Varetti G, Guida C, Santaguida S, et al.:
Homeostatic control of mitotic arrest.
Mol Cell.
2011; 44(5): 710–20. PubMed Abstract
| Publisher Full Text
89.
D'Angiolella V, Palazzo L, Santarpia C, et al.:
Role for non-proteolytic control of M-phase-promoting factor activity at M-phase exit.
PLoS One.
2007; 2(2): e247. PubMed Abstract
| Publisher Full Text
| Free Full Text
90.
Visconti R, Palazzo L, Della Monica R, et al.:
Fcp1-dependent dephosphorylation is required for M-phase-promoting factor inactivation at mitosis exit.
Nat Commun.
2012; 3: 894. PubMed Abstract
| Publisher Full Text
| Free Full Text
91.
Visconti R, Della Monica R, Palazzo L, et al.:
The Fcp1-Wee1-Cdk1 axis affects spindle assembly checkpoint robustness and sensitivity to antimicrotubule cancer drugs.
Cell Death Differ.
2015; 22(9): 1551–60. PubMed Abstract
| Publisher Full Text
| Free Full Text
92.
Della Monica R, Visconti R, Cervone N, et al.:
Fcp1 phosphatase controls Greatwall kinase to promote PP2A-B55 activation and mitotic progression.
eLife.
2015; 4: pii: e10399. PubMed Abstract
| Publisher Full Text
| Free Full Text
1
CEINGE, Naples, 80145, Italy 2
DMMBM, University of Naples Federico II, Naples, 80131, Italy 3
Department of Pharmacy, University of Naples Federico II, Naples, 80131, Italy
This work was supported by a grant from AIRC (IG grant 2017; Id. 19851 to DG).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Serpico AF and Grieco D. Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.12688/f1000research.21185.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Raaijmakers J. Reviewer Report For: Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.5256/f1000research.23320.r59261)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Raaijmakers J. Reviewer Report For: Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.5256/f1000research.23320.r59261)
Nilsson J. Reviewer Report For: Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.5256/f1000research.23320.r59260)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Nilsson J. Reviewer Report For: Recent advances in understanding the role of Cdk1 in the Spindle Assembly Checkpoint [version 1; peer review: 2 approved]. F1000Research 2020, 9(F1000 Faculty Rev):57 (https://doi.org/10.5256/f1000research.23320.r59260)
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)