Keywords
p38, MAPK, inflammation, signalling
p38, MAPK, inflammation, signalling
p38α (originally named p38) was identified and cloned as a 38 kDa protein that was tyrosine-phosphorylated in response to LPS stimulation in mammalian cells1,2. Sequence comparison, on the day p38α was cloned, revealed that it belonged to the mitogen-activated protein kinase (MAPK) family and that a Saccharomyces cerevisiae osmotic response protein kinase HOG1 was a p38α homologue3–5. p38α was also named cytokine suppressive drug binding protein (CSBP) because it was identified as the target of a series of anti-inflammatory pyridinyl-imidazole compounds and as reactivating kinase (RK) because it phosphorylated and activated MK23–5. There are four members of the p38 group of MAPKs encoded by four different genes in mammals: p38α (MAPK14, chromosome 6p21.31 in humans), p38β (MAPK11, SAPK2b, Chr22q13.33)6, p38γ (MAPK12, ERK6, SAPK3, Chr22q13.33)7,8, and p38δ (MAPK13, SAPK4, Serk4, Chr6p21.31)9,10. As can be surmised from their chromosomal locations, MAPK14/p38α and MAPK13/p38δ are physically close and separated by just over 15 kb, as are MAPK12/p38β and MAPK11/p38γ, which are separated by less than 2 kb. All the p38s contain a conserved Thr–Gly–Tyr (TGY) dual phosphorylation motif within the kinase activation loop, and both Thr and Tyr phosphorylation are necessary to fully activate the kinase11. However, monophosphorylated p38α Thr180 has some kinase activity in vitro, but a different substrate specificity, when compared with dual-site phosphorylated p38α12. p38 group members are expressed ubiquitously, but p38γ and p38δ are enriched in certain cell types and tissues, such as p38γ in skeletal muscle and p38δ in the salivary, pituitary, and adrenal glands13. p38β shares more amino acid sequence identity with p38α (~70%), while p38γ and p38δ share ~60% identity with p38α. p38γ and p38δ also share high sequence homology with cyclin-dependent kinases (CDKs) and are sensitive to some CDK inhibitors14.
p38α is involved in the response to almost all stressful stimuli, including LPS, UV light, heat shock, osmotic shock, inflammatory cytokines, T cell receptor ligation, glucose starvation, and oncogene activation2,4,5,15–20. Under certain circumstances, it is also activated upon growth factor stimulation. It should be noted that the activation of p38 in some cases is cell type specific, since an activating stimulus in one cell type may inhibit p38 in other cell types21. The study of p38 group members other than p38α has been less intensive; however, where it has been examined, the other p38s are frequently co-activated with p38α22.
Like other MAPK signaling pathways, the activation of all p38s is mediated by a kinase cascade: MAPKKK (MAP3K), which activates MAPKK (MAP2K), which in turn activates MAPK. The MAP2K kinases MKK3 and MKK6 are the major upstream kinases for p38 activation23–25. Although MKK3 and MKK6 phosphorylate most p38 isoforms in vitro, selective activation and substrate specificity have been observed in vivo26. MKK4 has also been reported to phosphorylate p38α and p38δ in specific cell types9. A number of MAP3Ks have been reported to participate in p38 activation including TAK127, ASK128, DLK29, and MEKK429,30. Low-molecular-weight GTP-binding proteins in the Rho family, such as Rac1 and Cdc42, can activate p38 through binding to MEK1 or MLK1, which function as upstream activators of MAP3K31,32.
p38α can also be activated by MAP2K-independent mechanisms. TAB1 (TAK1-binding protein 1) directly interacts with p38α and can promote trans autophosphorylation on Thr180 and Tyr182 and thus full activation of p38α33. A subsequent study revealed that autophosphorylation of Thr180 and Tyr182 requires a conserved Thr185 residue34. TAB1-dependent p38α activation has been implicated in ischemic myocardial injury and T cell anergy35,36. TAB1 is also claimed to play a role in Sestrin-mediated p38α activation12. Another MAP2K-independent activation is mediated by ZAP70 after T cell receptor ligation. ZAP70 can directly phosphorylate p38α/β on Tyr32318, leading to autophosphorylation on Thr180, one of the dual phosphorylation sites. As discussed, mono-Thr180 phosphorylated p38 still has some kinase activity37, and loss of ZAP70-mediated p38 activation in p38αβY323F double knock-in mice reduces autoimmunity and inflammation in several autoimmune disease models38–40. Interestingly, p38α also phosphorylates ZAP70, resulting in a decrease in the size and persistence of the T cell receptor signaling complex, and therefore acts as a feedback regulator of ZAP7041.
Conversely, de-phosphorylation of both threonine and tyrosine residues in the activation loop inactivates MAPKs, and this is mainly carried out by dual-specificity phosphatases of the MAPK phosphatase (MKP)/dual specificity phosphatase (DUSP) family42. Although several MKPs have been reported to dephosphorylate p38α, MKP1/DUSP1, MKP5/DUSP10, MKP8/DUSP26, and DUSP8 are more potent inhibitors of p38α and JNK than ERK43. A recent report showed that DUSP12 is also a p38α phosphatase44. While there are a number of p38α DUSPs, no DUSP for p38γ or p38δ has been reported, and these two p38s are resistant to several known p38α MKPs such as MKP1, 3, 5, and 745. p38α-dependent upregulation of MKP1 was reported and is believed to be part of a negative feedback loop of p38α activation46. Other types of phosphatases have also been reported to target p38 MAPKs, such as CacyBP/SIP47, Wip148, and PP2C49,50. The substrate specificity between p38 and phosphatases and the related physiological functions in vivo still need further investigation. p38γ has also been reported to be degraded by a p38/JNK/ubiquitin-proteasome-dependent pathway, which represents an additional mechanism by which p38 kinases may cross regulate each other51. Yet other ways of regulating p38 are suggested from studies in Caenorhabditis elegans, where a genetic screen for resistance against bacterial infection identified RIOK-1, an atypical serine kinase and human RIO kinase homolog, as a suppressor of the p38 pathway52. As RIOK-1 is a transcriptional target of the p38 pathway in C. elegans, this suggests that RIOK-1 is part of a negative feedback loop. A brief summary of the p38 pathway is shown in Figure 1.
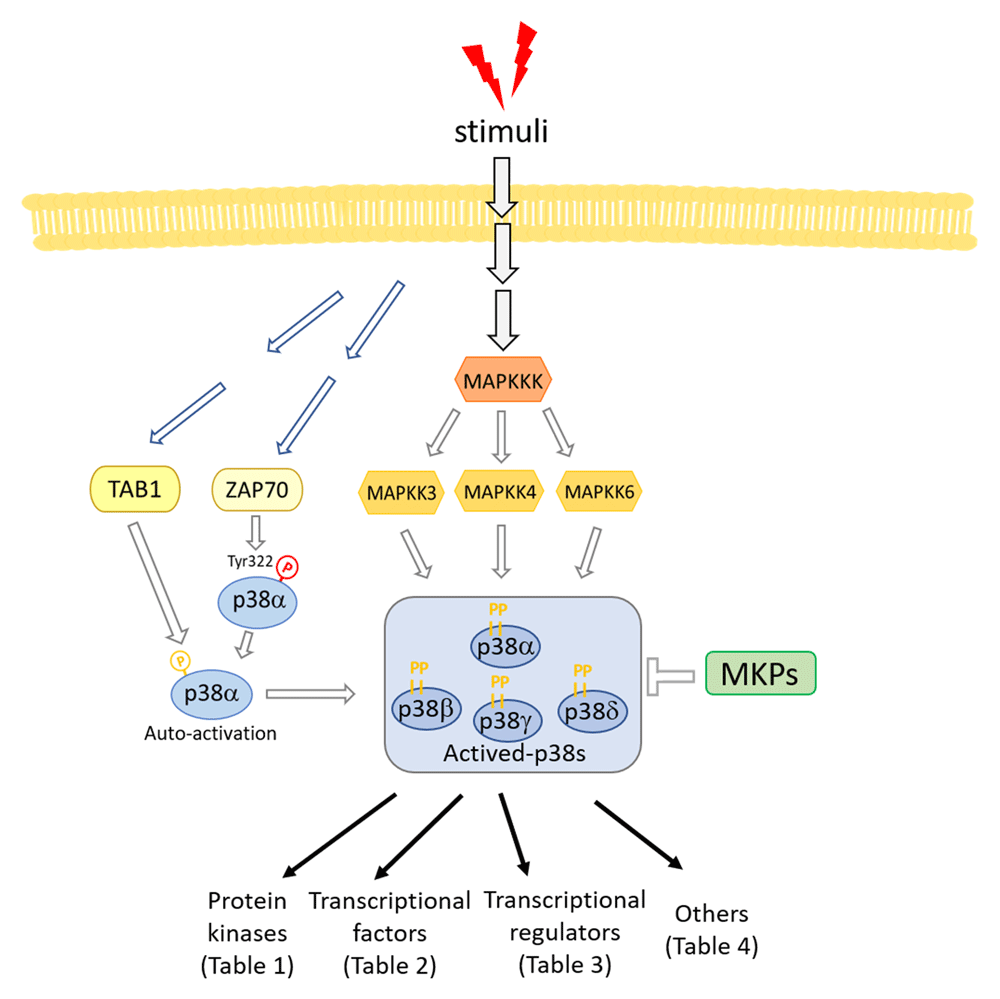
MKP, mitogen-activated protein kinase phosphatase; TAB1, TAK1-binding protein 1; Tyr, tyrosine.
The p38 MAPK cascade does not end at p38. Members of the MAPK-activated protein kinase (MAPKAPK) family such as MK2, MK3, and MK5 (PRAK) are all p38 substrates3,4,53–55. The MKs have a broad range of substrates that extend the range of functions regulated by p38 kinases. Mitogen- and stress-activated protein kinase-1/2 (MSK1/2), which are important for CREB activation and chromosome remodeling, have also been identified as substrates of p38α56. MNK1/2, kinases that phosphorylate the eukaryotic initiation factor-4e (eIF-4E), are phosphorylated by p38α57,58. p38α has also been reported to inactivate murine GSK3β by phosphorylating Ser389, and since GSK3β is required for the continuous degradation of β-catenin in the Wnt signaling pathway, this can lead to an accumulation of β-catenin59,60. It was also reported that p38δ negatively regulates insulin secretion by catalyzing an inhibitory phosphorylation of PKD161. A number of p38 protein kinase substrates are summarized in Table 1.
| Substrate | Kinase | Function | References |
|---|---|---|---|
| MAPKAPK2 (MK2) | p38α, p38β, p38γ, p38δ | Activates the kinase substrate | Freshney NW et al., Cell, 19944 Rouse J et al., Cell, 19943 |
| MAPKAPK3 (MK3) | p38α, p38β, p38γ, p38δ | Activates the kinase substrate | McLaughlin MM et al., J Biol Chem, 199654 |
| MNK1/2 | p38α | Activates the kinase substrate | Fukunaga R et al., EMBO J, 199758 Waskiewicz AJ et al., EMBO J, 199757 |
| MSK1/2 | p38α | Activates the kinase substrate | Deak M et al., EMBO J, 199856 Pierrat B et al., J Biol Chem, 199877 |
| PAK6 | p38α | Activates the kinase substrate | Kaur R et al., J Biol Chem, 200578 |
| PIP4Kb | p38α | Inactivates the kinase substrate | Jones DR et al., Mol Cell, 200679 |
| RPAK (MK5) | p38α, p38β | Activates the kinase substrate | New L et al., EMBO J, 199855 |
| PKCε | p38α, p38β | Completes cytokinesis | Saurin AT et al., Nat Cell Biol, 200880 |
| GSK3β | p38α | Inactivates the kinase substrate, activates Wnt pathway. | Bikkavilli RK et al., J Cell Sci, 200860 Thornton TM et al., Science, 200859 |
p38 targets a large number of transcription factors, including myocyte-specific enhancer factor 2 (MEF2) family members, cyclic AMP-dependent transcription factor 1, 2, and 6 (ATF-1/2/6), CHOP (growth arrest and DNA damage inducible gene 153, or GADD153), p53, C/EBPβ, MITF1, DDIT3, ELK1/4, NFAT, and STAT1/4. p38 phosphorylation of transcription factors predominantly leads to enhanced transcriptional activity. However, in some cases, it represses transcription, and this is summarized in Table 2. Transcription factor phosphorylation by p38 is often stimulus and cell type dependent and plays a role in the cellular response to inflammation, DNA damage, metabolic stress, and many other stresses62–76. The effects of p38 on transcription seem to constitute the major part of p38’s responses to stress stimuli.
| Substrate | Kinase | Function | References |
|---|---|---|---|
| ATF2 | p38α, p38β, p38γ, p38δ | Enhances transcriptional activity | Cuenda A et al., EMBO J, 199781 Jiang Y et al., J Biol Chem, 19979 |
| C/EBPα | p38α | Enhances transcriptional activity | Qiao L et al., J Biol Chem, 200682 |
| C/EBPβ | p38α | Enhances transcriptional activity | Engelman JA et al., J Biol Chem, 199883 |
| C/EBPε | p38α | Enhances transcriptional activity | Williamson EA et al., Blood, 200584 |
| CHOP | p38α, p38β | Enhances transcriptional activity | Wang XZ et al., Science, 199668 |
| E2F4 | p38α | Enhances transcriptional activity | Morillo SM et al., Mol Cell Biol, 201285 |
| Elk-1 | p38α | Enhances transcriptional activity in specific cell types | Janknecht R et al., EMBO J, 199767 Whitmarsh AJ et al., Mol Cell Biol, 199766 |
| ERα | p38α | Enhances nuclear localization and transcriptional activity | Lee H et al., Mol Cell Biol, 200286 |
| Fos | p38α, p38β, p38γ, p38δ | Enhances transcriptional activity | Tanos T et al., J Biol Chem, 200587 |
| FOXO3a | p38α | Enhances nuclear relocalization | Ho KK et al., J Biol Chem, 201288 |
| GR | p38α | Enhances transcriptional activity | Miller AL et al., Mol Endocrinol, 200589 |
| IUF1 | p38α, p38β | Enhances transcriptional activity | Macfarlane WM et al., J Biol Chem, 199790 |
| JDP2 | p38α | N/D | Katz S et al., Biochem J, 200291 |
| c-JUN | p38α, p38β, p38γ | Enhances transcriptional activity | Humar M et al., Int J Biochem Cell Biol, 200792 |
| MafA | p38α, p38β, p38γ, p38δ | Enhances transcriptional activity | Sii-Felice K et al., FEBS Lett, 200593 |
| MEF2A | p38α, p38β, p38δ | Enhances transcriptional activity | Zhao M et al., Mol Cell Biol, 199994 |
| MEF2C | p38α, p38β p38γ, p38δ | Enhances transcriptional activity | Han J et al., Nature, 199762 |
| MEF2D | p38α | Enhances recruitment of Ash2L to muscle-specific promoters | Zhao M et al., Mol Cell Biol, 199994 Rampalli S et al., Nat Struct Mol Biol, 200773 |
| MITF | p38α | Enhances transcriptional activity | Mansky KC et al., J Biol Chem, 200295 |
| MRF4 | p38α | Represses transcriptional activity | Suelves M et al., EMBO J, 200496 |
| NFATc1 | p38α | Enhances transcriptional activity and interaction with PU.1 | Matsumoto M et al., J Biol Chem, 200497 |
| NFATc4 | p38α, p38β p38γ | Represses nuclear localization and transcriptional activity | Yang TT et al., Mol Cell Biol, 200298 |
| NR4A | p38α | Enhances transcriptional activity | Sekine Y et al., J Cell Sci, 201199 |
| Nur77 | p38α | Disrupts interaction with p65 and represses transcriptional activity | Li L et al., Nat Chem Biol, 2015100 |
| Osterix | p38α | Enhances recruitment of coactivators | Ortuño MJ et al., J Biol Chem, 2010101 |
| p53 | p38α | Increases protein stability and apoptosis | Bulavin DV et al., EMBO J, 199969 |
| Pax6 | p38α | Enhances transcriptional activity | Mikkola I et al., J Biol Chem, 1999102 |
| PPARα | p38α | Enhances transcriptional activity | Barger PM et al., J Biol Chem, 2001103 |
| SAP1 | p38α, p38β p38γ, p38δ | Enhances transcriptional activity | Janknecht R et al., EMBO J, 199767 |
| Smad3 | p38α | Enhances nuclear translocation | Hayes SA et al., Oncogene, 2003104 |
| Snail | p38α | Increases protein stability and transcriptional activity | Ryu KJ et al., Cancer Res, 2019105 |
| STAT1 | p38α, p38β | Enhances transcriptional activity | Kovarik P et al., Proc Natl Acad Sci U S A, 1999106 |
| STAT4 | p38α | Enhances transcriptional activity | Visconti R et al., Blood, 2000107 |
| TEAD4 | p38α | Enhances cytoplasmic translocation and suppresses transcriptional activity | Lin KC et al., Nat Cell Biol, 201776 |
| Twist1 | p38α | Increases protein stability and transcriptional activity | Hong J et al., Cancer Res, 2011108 |
| USF1 | p38α | Enhances transcriptional activity | Galibert MD et al., EMBO J, 200171 |
| Xbp1s | p38α | Enhances nuclear translocation and transcriptional activity | Lee J et al., Nat Med, 201175 |
ATF2, activating transcription factor 2; C/EBP, CCAAT/enhancer binding protein; CHOP, CCAAT/enhancer-binding protein homologous protein; ER, estrogen receptor; GR, glucocorticoid receptor; IUF1, insulin upstream factor 1; JDP2, Jun dimerization protein 2; MEF, myocyte-specific enhancer factor; MITF, microphthalmia transcription factor; MRF, muscle regulatory factor; NFAT, nuclear factor of activated T cells; Pax6, paired box 6; PPARα, peroxisome proliferator-activated receptor alpha; TEAD4, TEA domain family transcription factor 4; USF1, upstream transcription factor 1; Xbp1s, spliced form of X-box binding protein 1.
A large number of transcriptional regulators, including epigenetic enzymes, are substrates of p38, and these are summarized in Table 3. The SWI–SNF complex subunit BAF60 is phosphorylated and inactivated by p38 during skeletal myogenesis109,110, and EZH2, the catalytic component of the Polycomb Repressive Complex 2 (PRC2), was also found to be phosphorylated by p38, particularly in ER-negative breast cancer samples111. Besides its transcriptional function, dATF-2 is also involved in heterochromatin formation, and stress-induced phosphorylation of dATF-2 by p38 disrupts heterochromatin in Drosophila112.
| Substrate | Kinase | Function | References | |
|---|---|---|---|---|
| Chromatin remodeling regulators | BAF60c | p38α, p38β | Activates transcription of MyoD- target genes | Simone C et al., Nat Genet, 2004109 Forcales SV et al., EMBO J, 2012110 |
| RNF2 | p38α | Modulates gene expression and histone 2B acetylation | Rao PS et al., Proteomics, 2009124 | |
| EZH2 | p38α | Promotes cytoplasmic localization | Anwar T et al., Nat Commun, 2018111 | |
| dAFF2 | p38α, p38β | Disrupts heterochromatin formation | Seong K-H et al., Cell, 2011112 | |
| Other regulators | CRTC2 | p38α | Enhances nucleocytoplasmic transport and represses transcription activity | Ma H et al., Mol Cell Biol, 2019125 |
| E47 | p38α, p38β | Enhances the formation of MyoD/ E47 heterodimers | Page JL et al., J Biol Chem, . 2004126 Lluís F et al., EMBO J, 2005127 | |
| HBP1 | p38α | Increases protein stability and represses transcription | Xiu M et al., Mol Cell Biol, 2003128 | |
| p18(Hamlet) | p38α, p38β | Increases protein stability and enhances transcription | Cuadrado A et al., EMBO J, 2007129 | |
| PGC-1α | p38α, p38β | Increases protein stability and enhances transcription | Puigserver P et al., Mol Cell, 2001130 | |
| Rb1 | p38α, p38γ | Induces Rb degradation and cell death; suppresses Rb activity and promotes the G0-to-G1 transition | Delston RB et al., Oncogene, 2011131 Tomás-Loba A et al., Nature, 201914 | |
| SRC-3 | p38α | Induces SRC-3 degradation and suppresses RARα-dependent transcription | Giannì M et al., EMBO J, 2006132 |
Given the wide range of responses that p38 is involved in, it is not surprising that many p38 substrates cannot be so easily categorized into groups, and these miscellaneous substrates are summarized in Table 4. Some of them are involved in metabolism such as Raptor phosphorylation by p38β, which enhances mTORC1 activity in response to arsenite-stress113, and DEPTOR (mTOR-inhibitory protein) phosphorylation by p38γ and p38δ, leading to its degradation and mTOR hyperactivation114. p38α phosphorylation of Tip60 at Thr158 promotes senescence and DNA-damage-induced apoptosis115,116. Some p38 substrates are cell death regulators. In the ER stress response, p38α locates to the lysosome and phosphorylates the chaperone-mediated autophagy (CMA) receptor LAMP2A, leading to activation of CMA and thus protecting cells from ER stress-induced death117.
| Substrate | Kinase | Function | References | |
|---|---|---|---|---|
| Cell-cycle regulators | Cdc25A | p38α | Increases protein stability | Goloudina A et al., Cell Cycle, 2003133 |
| Cdc25B | p38α | Increases protein stability | Lemaire M et al., Cell Cycle, 2006134 | |
| Cyclin D1 | p38α | Causes ubiquitination and degradation of cyclin D1 | Casanovas O et al., J Biol Chem, 2000135 | |
| Cyclin D3 | p38α, p38β p38γ, p38δ | Causes ubiquitination and degradation of cyclin D3 | Casanovas O et al., Oncogene, 2004136 | |
| p57kip2 | p38α | Enhances interaction with CDKs and inhibits CDKs | Joaquin M et al., EMBO J, 2012137 | |
| Cell-death regulators | Bax | p38α | Prevents Bcl-2–Bax heterodimer formation, enhances apoptosis | Min H et al., Mol Carcinog, 2012138 |
| BimEL | p38α | Enhances apoptosis | Cai B et al., J Biol Chem, 2006139 | |
| Caspase-3 | p38α | Inhibits caspase-3 activity and apoptosis | Alvarado-Kristensson M et al., J Exp Med, 2004140 | |
| Caspase-8 | p38α | Inhibits caspase-8 activity and apoptosis | Alvarado-Kristensson M et al., J Exp Med, 2004140 | |
| Caspase-9 | p38α | Inhibits caspase-9 activity and apoptosis | Seifert A et al., Cell Signal, 2009141 | |
| DNA/RNA binding proteins | Cdt1 | p38α, p38β | Increases protein stability | Chandrasekaran S et al., Mol Cell Biol, 2011142 |
| Drosha | p38α | Enhances nuclear export and degradation | Yang Q et al., Mol Cell, 2015143 | |
| FBP2 | p38α | Promotes prothrombin mRNA 3' end processing | Danckwardt S et al., Mol Cell, 2011144 | |
| FBP3 | p38α | Promotes prothrombin mRNA 3' end processing | Danckwardt S et al., Mol Cell, 2011144 | |
| H2AX | p38α, p38β | Promotes serum starvation-induced apoptosis | Lu C et al., FEBS Lett, 2008145 | |
| H3 | p38α | N/D | Zhong SP et al., J Biol Chem, 2000146 | |
| HuR | p38α, p38β | Enhances cytoplasmic accumulation and increases mRNA stability | Lafarga V et al., Mol Cell Biol, 2009147 | |
| KSRP | p38α, p38β | Prevents KSRP-mediated ARE-directed mRNA decay | Briata P et al., Mol Cell, 2005148 | |
| Rps27 | p38α | N/D | Knight JD et al., Skelet Muscle, 2012149 | |
| SPF45 | p38α | Inhibits Fas alternative splicing (exon 6 exclusion) | Al-Ayoubi AM et al., Mol Cell Biol, 2012150 | |
| Endocytosis regulators | EEA1 | p38α | Promotes recruitment to endocytic membranes and enhances MOR endocytosis | Macé G et al., EMBO J, 2005151 |
| Rabenosyn-5 | p38α | Promotes recruitment to endocytic membranes and enhances MOR endocytosis | Macé G et al., EMBO J, 2005151 | |
| GDI-2 | p38α | Enhances GDI:Rab5 complex formation and modulates endocytosis | Cavalli V et al., Mol Cell, 2001152 | |
| MAPK pathway regulator | JIP4 | p38α | Enhances p38 activity | Kelkar N et al., Mol Cell Biol, 2005153 |
| Tip60 | p38α | Enhances the pro-senescent function of Tip60 | Zheng H et al., Mol Cell, 2013115 | |
| TAB1 | p38α | Inhibits TAK1 activity | Cheung PC et al., EMBO J, 2003154 | |
| TAB3 | p38α | Inhibits TAK1 activity | Mendoza H et al., Biochem J, 2008155 | |
| FRS2 | p38α | Downregulates FGF1-induced signaling | Zakrzewska M et al., Int J Mol Sci, 2019156 | |
| Membrane proteins | EGFR | p38α | Induces EGFR internalization | Winograd-Katz SE et al., Oncogene, 2006157 |
| FGFR1 | p38α | Regulates translocation of exogenous FGF1 into the cytosol/nucleus | Sørensen V et al., Mol Cell Biol, 2008158 | |
| Nav1.6 | p38α | Promotes interaction with NEDD-4 and protein degradation | Gasser A et al., J Biol Chem, 2010159 | |
| NHE1 | p38α | Induces intracellular alkalinization | Khaled AR et al., Mol Cell Biol, 2001160 | |
| PLA2 | p38α | N/D | Börsch-Haubold AG et al., J Biol Chem, 1998161 | |
| TACE | p38α, p38β | Increases TACE-mediated ectodomain shedding and TGF-alpha family ligand release | Xu P et al., Mol Cell, 2010162 | |
| ZAP70 | p38α | Phosphorylation of ZAP70 increases stability of T cell receptor | Giardino Torchia ML et al., Proc Natl Acad Sci U S A, 201841 | |
| Structure proteins | Caldesmon | p38α | N/D | Hedges JC et al., Am J Physiol, 1998163 |
| Hsp27 | p38α | N/D | Knight JD et al., Skelet Muscle, 2012149 | |
| Keratin 8 | p38α | Regulates cellular keratin filament reorganization | Ku NO et al., J Biol Chem, 2002164 | |
| Lamin B1 | p38α | Enhances lamin B1 accumulation | Barascu A et al., EMBO J, 2012165 | |
| Paxillin | p38α | Required for NGF-induced neurite extension of PC-12 cells | Huang C et al., J Cell Biol, 2004166 | |
| Stathmin | p38δ | N/D | Parker CG et al., Biochem Biophys Res Commun, 1998167 | |
| SAP97 | p38γ | Modulating the association of this protein with other cytoskeleton proteins | Sabio G et al., EMBO J, 2005168 | |
| Tau | p38α, p38γ, p38δ | Enhances formation of paired helical filaments Inhibits amyloid-β toxicity in Alzheimer's mice | Reynolds CH et al., J Neurochem,1997169 Ittner A et al., Science, 2016170 | |
| Tensin1 | p38α | Regulates the binding specificity of tensin1 to different proteins | Hall EH et al., Mol Cell Proteomics, 2010171 | |
| Others | DEPTOR | p38γ, p38δ | Enhances degradation and mTOR hyperactivation | González-Terán B et al., Nat Commun, 2016114 |
| GS | p38β | Required for subsequent phosphorylation to inhibit enzyme activity | Kuma Y et al., Biochem J, 2004172 | |
| LAMP2A | p38α | Activates chaperone-mediated autophagy | Li W et al., Nat Commun, 2017117 | |
| Parkin | p38α | Decreases its interaction with PINK1 and suppresses mitophagy | Chen J et al., Cell Death Dis, 2018173 | |
| p47phox | p38α | Promotes NADPH oxidase activation and superoxide production | Makni-Maalej K et al., J Immunol, 2012174 | |
| p62 | p38γ, p38δ | Enhances mTORC1 activity | Linares JF et al., Cell Rep, 2015175 Koh A et al., Cell, 2018176 | |
| Raptor | p38β | Enhances mTORC1 activity in response to arsenite stress | Wu X-N et al., J Biol Chem, 2011113 | |
| Rpn2 | p38α | Inhibits proteasome activity | Lee SH et al., J Biol Chem, 2010177 | |
| Siah2 | p38α | Increases Siah2-mediated degradation of PHD3 | Khurana A et al., J Biol Chem, 2006178 |
CDK, cyclin-dependent kinase; EGFR, epidermal growth factor receptor; FBP1, far upstream binding protein; FGF1, fibroblast growth factor 1; FGFR1, fibroblast growth factor receptor 1; FRS2, fibroblast growth factor receptor substrate 2; GDI, GDP dissociation inhibitor; KSRP, hnRNPK-homology type splicing regulatory protein; MAPK, mitogen-activated protein kinase; mTORC1, mammalian target of rapamycin complex 1; NADPH, nicotinamide adenine dinucleotide phosphate; NGF, nerve growth factor; NHE1, Na+/H+ exchanger isoform 1; PHD3, prolyl hydroxylase 3; PLA2, phospholipase A2; SAP97, synapse-associated protein 97; TAB, transforming growth factor-β-activated protein kinase-1-binding protein; TACE, tumor necrosis factor-alpha-converting enzyme; TAK1, transforming growth factor β-activated kinase 1; TGF, transforming growth factor.
p38α is required for embryo development, since the mouse Mapk14–/– embryo dies between embryonic days (E) 10.5 and 12.5118–121. Mutant mice with a single Thr180 to Ala mutation or with the double T180A Y182F mutation are also embryonic lethal122,123. Surprisingly, given the importance of the dual phosphorylation for complete p38 activation, substitution of Tyr182 with Phe results in mice that have reduced p38 signaling but are nevertheless viable123, although this is consistent with previous studies showing that the p38 phosphorylated on Thr180 alone retains some activity in vitro37. Histological analysis demonstrates that p38α is required for placental angiogenesis, but not embryonic cardiovascular development, and tetraploid rescue of the placental defect in Mapk14–/– embryos confirmed that p38α is essential for extraembryonic development120,121. Given the important role that p38 and MK2 plays in regulating TNF-induced cell death179–182, it is intriguing that the Mapk14–/– embryonic lethal phenotype is very similar to that observed in other mice with defects in the TNF death pathway. Caspase-8, FADD, and cFLIP knock-out mice also die at E10.5, and this is due to TNF-dependent endothelial cell death and disruption of the vasculature in the yolk sac183,184. Other p38 isoforms are not necessary for embryo development, but p38α and p38β have overlapping functions, as Mapk14loxp/loxpMapk11–/–Sox2-Cre embryos die before E16.5 with spina bifida that correlates with neural hyperproliferation and increased apoptosis in the liver, which was not observed in Mapk14∆/∆Sox2-Cre embryos185. Remarkably, p38α appears to have a very specific function during embryogenesis because when p38α was replaced by p38β in the Mapk14 chromosomal locus, which thereby placed p38β under the control of the endogenous p38α promoter, it was unable to rescue the embryonic lethality induced by loss of p38α185.
p38 is activated by many inflammatory stimuli, and its activity is important for inflammatory responses. Macrophage-specific deletion of Mapk14 inhibits inflammatory cytokine production and protects mice from CLP-induced sepsis186. p38α controls the production of inflammatory cytokines, such as TNF and IL-6, at many levels. It directly phosphorylates transcription factors, such as MEF2C62,186, and regulators of mRNA stability, such as hnRNPK-homology (KH) type splicing regulatory protein (KSRP)187. MEF2C appears to play an anti-inflammatory role in endothelial cells in vivo188. Via MK2/MK3, p38 also upregulates cytokine mRNA transcription by the serum response transcription factor (SRF)189, and similarly, via MK2/MK3, p38 regulates mRNA stability by phosphorylating and inactivating TTP/Zfp36, a protein that promotes rapid turnover of AU-rich mRNAs, many of which are cytokine mRNAs187,190. p38 activation also induces the expression of inflammatory mediators such as COX-2, MMP9, iNOS, and VCAM-1, which are involved in tissue remodeling and oxidation regulation191–194. The p38 pathway also regulates adaptive immunity. p38α participates in antigen processing in CD8+ cDCs195, and ZAP70-mediated p38α/β activation is important for T cell homeostasis and function18. In B cells, p38α is important for CD40-induced gene expression and proliferation of B cells196, and the p38α–MEF2c axis is believed to be necessary for germinal center B (GCB) cell proliferation and survival197,198. Excessive activation of p38α has been observed in many inflammatory diseases, such as inflammatory bowel disease (IBD), asthma, rheumatoid arthritis, and steatohepatitis199–201. The other members of the p38 family also play roles in immune responses. For example, p38γ and p38δ are required for neutrophil migration to damaged liver in non-alcoholic fatty liver disease202 and inhibition of eukaryotic elongation factor 2 in LPS-induced liver damage203. p38δ is required for neutrophil accumulation in acute lung injury204. These observations, and the role that p38s play in TNF production, led to enormous pharmaceutical efforts to develop p38 inhibitors to treat chronic inflammatory diseases. However, unfortunately, these drugs were not efficacious in these diseases205.
p38 has been implicated in G1 and G2/M phases of the cell cycle in several studies. The addition of activated recombinant p38α caused mitotic arrest in vitro, and an inhibitor of p38α/β suppressed activation of the checkpoint by nocodazole in NIH3T3 cells206. G1 arrest caused by Cdc42 overexpression is also dependent on p38α in NIH3T3 cells207. Besides, p38γ is specially required for gamma-irradiation-induced G2 arrest208. The link between p38 and cell cycle control has been proposed through the regulation of several p38 substrates. Both p38α and p38γ regulate cell cycle progression via Rb but in opposite directions14,209. HBP1 represses the expression of cell cycle regulatory genes during cell cycle arrest in a p38-dependent manner210; p53 and p21 activation by p38α prevented G1 progression through blockade of CDK activity211,212. The p38 pathway is also involved in cell cycle progress, as it is essential for self-renewal of mouse male germline stem cells213 and its regulation of G1-length plays a role in cell size uniformity214.
Participation of p38 in cell differentiation has been reported in certain cell types. p38α activity is essential for neuronal differentiation in PC-12 cells and EPO-induced differentiation in SKT6 cells20,215. Treatment of 3T3-L1 fibroblasts with specific p38α/β inhibitors prevents their differentiation into adipocytes by reducing C/EBPβ phosphorylation83, and p38α-dependent phosphorylation of MEF2C and BAF60 is critical for myogenic differentiation110,216. Intestinal epithelial cell-specific deletion of p38α also influences goblet cell differentiation in a Notch-dependent manner200.
p38 group members participate in many cellular events related to metabolism. The p38β–PRAK axis specifically phosphorylates Rheb and suppresses mTORC1 activity under energy depletion conditions22. DEPTOR, an inhibitor of mTORC, can be phosphorylated by p38γ and p38δ, leading to its degradation123. Meanwhile, p38δ directly phosphorylated p62 to enhance mTORC1 activity in response to amino acids175. In brown adipocytes, p38α functions as a central mediator in β-adrenergic-induced UCP1 expression217,218, while in white adipocytes, p38α inactivation leads to elevated white-to-beige adipocyte reprogramming and resistance to diet-induced obesity219,220. In hepatocytes, p38α controls lipolysis and protects against nutritional steatohepatitis. Thus, mice with hepatocyte-specific loss of p38α developed more severe steatohepatitis than wild type mice when fed high-fat or -cholesterol diets. Intriguingly, macrophage specific deletion of p38 had the opposite effect in the same high-fat diets and resulted in less steatohepatitis than in wild type mice, which probably reflects the inflammatory role of p38 in macrophages199. p38α also directly phosphorylates Xbp1s to enhance its nuclear migration for maintaining glucose homeostasis in obesity75. However, p38α also functions as a negative regulator of AMPK signaling in maintaining gluconeogenesis, and hepatic p38α could be a drug target for hyperglycemia221. It was also reported that p38γ directly phosphorylated p62 under imidazole propionate stimulation to promote mTORC1 activity in hepatocytes176. Interestingly, AMPK also triggers the recruitment of p38α to scaffold protein TAB1 for p38α autoactivation in human T cells222.
p38α appears to play a pivotal role in senescence. Constitutive activation of the p38 pathway by active MKK3 or MKK6 induces senescence in several cell types223,224, and p38α activity is responsible for senescence induced by multiple stimuli, such as telomere shortening225,226, H2O2 exposure227,228, and chronic oncogene activation19,223,229. p38α/β-specific inhibitors have been successfully used to prevent cellular senescence in cultivated human corneal endothelial cells230. Since cellular senescence is considered a defense strategy against oncogene activation, the p38 pathway plays important roles in tumorigenesis231. Meanwhile, p38α activity is important for senescence-associated secretory phenotype (SASP), and its inhibition markedly reduces the secretion of most SASP factors, suggesting multiple roles for the p38 pathway in senescence232–235.
The role of the p38 pathway in cell fate is cell type and stimulus dependent. For example, p38α becomes activated upon NGF withdrawal in PC-12 cells, and p38α activated by overexpression of MKK3 induced apoptosis in NGF differentiated PC-12 cells211. Similarly, inhibition of p38 with PD169316 blocked NGF withdrawal-induced apoptosis in PC-12 cells236,237. The interplay between the p38 pathway and caspases, the central regulators/executors of apoptosis, is complicated because p38α activity can be elevated in a caspase-dependent manner in death stimulus treated cells238,239, and caspase activity can also be elevated in MKK6E (dominant active form) overexpressed cells239,240. In contrast, inhibition of caspase-8 and caspase-3 by p38α-mediated phosphorylation in neutrophils was also reported140. Recent studies show that p38-activated MK2 directly phosphorylates RIPK1 in TNF-treated cells or pathogen-infected cells, limiting TNF-induced cell death180–182. This represents an interesting link between cytokine production induced by TNF and cell death because TNF-induced MK2/MK3 phosphorylation of tristetraprolin/Zfp36 inactivates it and leads to increased stability of cytokine mRNAs190. Aberrant p38α activity is observed in many tumor cells, and inhibition of p38α/β enhances cell death in these cells241,242.
p38 is one of the most researched of all proteins, let alone kinases, and a search in PubMed for p38 MAPK or p38 kinase returns more than 36,000 publications, which is a higher number than some proteins listed in a review of the "top 10" most studied genes243. By contrast, searches for the kinases Raf and Src return about 17,000 and 25,000 hits, respectively. In 2018, there were more than 2,000 publications that mention p38, and it is clearly impractical to summarize such a vast amount of literature. As might be surmised from the preceding commentary, the studies are on a wide range of topics; however, the publications are more concentrated in some areas than others. The role of the p38 pathway in cancers (>10,000)244–246, inflammation (>8,000)247–249, and infections (>3,600)250,251 was intensively studied. About 1,600 publications include the specific term "p38 inhibitor". This reflects the previously mentioned enormous interest of the pharmaceutical industry in developing p38 inhibitors to treat chronic inflammatory diseases, such as rheumatoid arthritis. Yet other publications report natural products that can activate or inhibit p38, with the ultimate aim of using them clinically252–258. In 2011, the European Commission approved Esbriet (pirfenidone), which was described as a p38γ inhibitor, for the treatment of idiopathic pulmonary fibrosis259. However, when this drug was approved by the FDA in 2014 for treating the same disease, it was described as a compound that acts on multiple pathways. In 2008, there were 27 clinical trials listed testing the use of p38 inhibitors in inflammatory disease settings205, while a search today for p38 inhibitors in clinicaltrials.gov returns 44 studies for conditions as diverse as pain, asthma, cognitive impairment, rheumatoid arthritis, cancer, myelodysplastic syndrome, and depression (Table 5). This indicates that there remains clinical interest in targeting the pathway and that there is therefore a need for more specific inhibitors of each of the p38 group members and more basic research to fully understand how the pathway, especially how each member of the p38 family, is utilized and regulated.
One consequence of the massive pharmaceutical effort over the last 20 years is a large number of very specific, well-tolerated, and readily bioavailable drugs that can enable such basic research. For example, one study using a boutique panel of kinase inhibitors was able to demonstrate that 11 potent and specific p38 inhibitors synergized with Smac-mimetic drugs to kill a subset of AML leukemias, providing the strongest evidence implicating p38 in Smac-mimetic-induced killing179. Since several of these p38 inhibitors had already been clinically trialed, this presents an opportunity to fast-track such combinations into the clinic. In our opinion, it is likely that this is where the future of p38 research and p38 inhibitors lies, in revealing the intricate web of inter-connections and inter-dependencies of this core and central regulator of cell stress. We also believe that greater efforts to genetically assess the role of p38 and p38 isoforms in the pathophysiology of inflammatory and other diseases need to be made in order to push forward the clinical application of our burgeoning knowledge.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)