Introduction
l-ascorbic acid (AA, vitamin C) is an important antioxidant with multiple functions in many species. It serves as a scavenger of reactive oxygen species generated under adverse environmental conditions. However, AA also influences flowering time and senescence1–3, pathogen disease resistance2,4, the biosynthesis of various plant hormones5–7, and root development8–11. This suggests that AA and some of its intermediates have functions in addition to its antioxidant properties.
Ascorbic acid biosynthesis in plants occurs predominantly through the d-mannose/l-galactose pathway12,13. Given the multifaceted functions of AA in plants, there is a need to advance our understanding of how plants regulate the biosynthesis and accumulation of AA. Arabidopsis thaliana mutants deficient in AA have provided important insights into the breadth of molecular and physiological functions of AA. One of the Arabidopsis mutants, vtc1-1, contains a defect in the AA biosynthetic enzyme GDP-mannose pyrophosphorylase. The mutant was originally generated by ethyl methanesulfonate (EMS) mutagenesis of Col-0 wild-type seed14. The vtc1-1 mutant contains a point mutation in amino acid 22 that converts a conserved proline into a serine15. The VTC1 gene has recently been shown to be a determinant of ammonium sensitivity in plants. In the presence of ammonium, vtc1-1 mutants exhibit strongly reduced root growth in comparison to the wild type, a phenomenon that is independent of AA deficiency8–11. To better understand the mechanism through which VTC1 mediates conditional ammonium sensitivity, it is important to identify regulatory partners of VTC1. To accomplish this, we undertook a suppressor mutagenesis approach of vtc1-1 homozygous mutant seed in the hope of identifying vtc1-1 suppressor mutants that could then be isolated and studied.
One of the suppressor mutants isolated in the M0 generation, svt2 (suppressor of vtc1-1 2), contained wild-type AA levels and developed roots similar to the wild type in the presence of ammonium. However, while characterizing the mutant genotypically, we observed that it lost the original vtc1-1 mutation (i.e., svt2 contained the homozygous wild-type allele). Furthermore, we determined that svt2, although generated through EMS mutagenesis of Col-0 vtc1-1 mutant seed, was phenotypically and genotypically similar to Ler. Intriguingly, a small percentage of svt2 M1 plants produced offspring that have phenotypic and genotypic similarities to Col in the M2 generation. Even more remarkably, a small percentage of Col-like revertants in the M2 generation produced progeny that exhibited phenotypic and genotypic svt2 characteristics again in the M3 generation.
Phenotypic instability of Arabidopsis alleles affecting a disease resistance gene cluster has recently been reported16. In their work, Yi and Richards described that exposure to EMS or through the generation of different F1 hybrids induced phenotypic instability in the bal and cpr1 mutant alleles. The authors later proposed that the high phenotypic instability is caused by a genetic mechanism17.
The presented study focuses on describing and characterizing the Arabidopsis svt2 suppressor mutant and its phenotypic and genotypic behavior. After illustrating the phenotypic features of svt2, we investigate transgenerational changes in the phenome and genome of svt2 and provide evidence that svt2 is a true mutant and not the result of an experimental artifact or contamination. Finally, we discuss our experimental findings in respect to the vtc1-1 mutant background and other reports that previously described similar phenomena of genome instability and restoration, and we briefly speculate on possible mechanisms of phenome and genome instability in svt2.
Materials and methods
Plant material and growth conditions
Arabidopsis thaliana Col-0 wild type and the previously described vtc1-1 mutant14 (in the Col-0 background) were kindly provided by Patricia Conklin (SUNY Cortland, NY, USA). Ler-0 wild-type seed were obtained from The Arabidopsis Biological Resource Center (http://www.arabidopsis.org). Plants were grown in Metromix 360 potting soil at 23°C at both day and night with a 16-hour photoperiod at 160 μmol photons m-2 s-2 (fluorescent bulbs).
For assessment of root growth, seed of the wild types and mutant lines were surface-sterilized (see below) and grown on basal full strength 1× Murashige and Skoog (MS) medium without vitamins (Cat.# MSP01, Caisson Laboratories, Inc., North Logan, UT), containing 1% Phytoblend (Cat.# PTP01, Caisson Laboratories) in omni trays (Fisher Scientific, Pittsburgh, PA) as described11. Sucrose was omitted from the tissue culture medium. The pH of the medium was adjusted with KOH to 5.7. Trays were sealed with two layers of 3M micropore tape (Fisher Scientific), put in vertical orientation, and placed in the growth chamber under long days (16 h light, 8 h dark) at 23°C day and night, and 160 µmol photons m-2 s-1 in a growth chamber (Percival Scientific, Inc., Perry, IA). Each plate contained wild-type and mutant seed. Primary root length was measured in seven-day-old seedlings using a ruler.
To assess AA content in leaf tissue, seeds of wild type and mutants were randomly sown on MetroMix 360 soil (BFG supplies Co., Burton, OH) in the same flat under the growth conditions described above. When plants were three weeks old, whole rosettes were harvested for the AA assay.
Seed-surface sterilization
Seeds were soaked for 1 min in 50% ethanol, followed by washing the seeds in 50% bleach plus 0.01% sodium dodecyl sulphate for 6 min. Finally, seeds were rinsed six times with sterile water and stored in 0.1% sterile Phytoblend agar for 2 d at 4°C18.
Ethyl methanesulfonate mutagenesis
Seeds of homozygous vtc1-1 Arabidopsis thaliana (Col-0 background) were mutagenized with 0.2% ethyl methanesulfonate as described (Figure 1;18). Approximately, 1200 M0 seed were stratified for 4 days at 4°C in 0.1% agar, sown on MetroMix soil and grown as above. Plants were screened for wild-type AA levels using the nitroblue tetrazolium assay19. Additional suppressor mutants were isolated by pooling seeds generated from M1 plants. Putative mutants were isolated and allowed to self-pollinate to obtain seed.
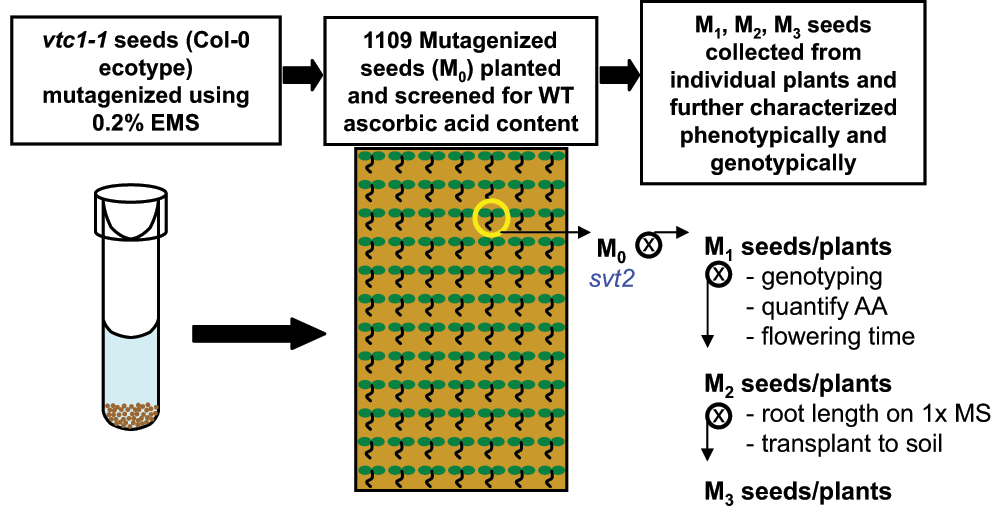
Figure 1. Isolation of svt2.
To isolate vtc1-1 suppressor mutants, homozygous vtc1-1 seed (in the Col-0 genetic background) were exposed to chemical mutagenesis using ethyl methanesulfonate (EMS). Over 1000 mutagenized seed (M0) were planted on soil and screened for wild-type levels of ascorbic acid. The only mutant isolated in the M0 generation containing recovered ascorbic acid levels was svt2. The mutant was allowed to self-fertilize and was characterized phenotypically and genotypically in subsequent generations.
Pollen grain analysis and microscopy
Pollen was taken from 4.5-week-old flowering plants of Col-0 and Ler wild type and vtc1-1 and svt2 M2 mutants, mounted in glycerol, and photographed using bright field settings on a Nikon E800 microscope equipped with a CoolSNAP cf CCD camera (Photometrics, Tuscon, AZ, USA).
Genomic DNA isolation
Genomic DNA was isolated from rosette leaves following a previously described protocol3. In case of genomic DNA isolation from vtc1-1 seeds, a small amount of dried seeds was crushed and the extraction procedure described previously3 was followed. Primers for the VTC1 gene and for the Insertion/Deletion (InDel) polymorphisms were designed using sequence data available on The Arabidopsis Information Resource (TAIR) database (http://www.arabidopsis.org). Polymerase chain reaction (PCR) was used to amplify fragments of the VTC1 gene for sequencing and to assess InDel polymorphisms. Sequences of primers used for sequencing and InDel analysis are summarized in Table 1. PCR reactions were run on 1.0% agarose gels stained with ethidium bromide.
Table 1. Forward (F) and reverse (R) sequences of primers used in analyzing the VTC1 gene and for amplifying five Col/Ler Insertion/Deletion (InDel) polymorphisms.
| Primer Name | 5´ 3´ |
|---|
| VTC1 G1 F | AAA AAT TCG TTC TAG ATG GAT GCT |
| VTC1 G1 R | ATG GCT GTA AAT TGG AAG AGA T |
| VTC1 G2 F | GAA CCC TTG TCT CTA AAA TA |
| VTC1 G2 R | CAA ATC CCA TAA TCT GTT CC |
| VTC1 G3 F | CAA TTT TGC TTA CTT CTC T |
| VTC1 G3 R | TGG ATG CAA CCG ACA CAA AAC AAT |
| VTC1 G4 F | ACA TTT TTA GCA GCT GGT ATT GAG |
| VTC1 G4 R | AGG TAA GAA CTG GCA GAC TAA AG |
| VTC1 G5 F | TCG CTT GAG ACC ATT GAC T |
| VTC1 G5 R | GAG GCT TCC CCA CCG TGA GAT TTG |
| VTC1 G6 F | CAA GCT GGA AAT CAA AAT CAC T |
| VTC1 G6 R | GCG CTG CTG CAA TCT TAG G |
| VTC1 G7 F | ACA AAT CTC ACG GTG GGG AAG C |
| VTC1 G7 R | TGG TTA ATT TGG CAG GAG A |
| VTC1 G8 F | CAA GGG CTC TAT GCT ATG GTG |
| VTC1 G8 R | GCG TTT TGA TTG ATG CTT ATT C |
| VTC1 G9 F | GCG TGT ATC TCG AGC AGT ATC AT |
| VTC1 G9 R | GTG GAG GGA AGT TAA GGG TAT TTT |
| InDel 1 450919 F | ATC GGT TTG TAA TCT CTG TCC A |
| InDel 1 450919 R | TAT GCG TTC CCA AAT TTG TTA TCT C |
| Indel 2 451470 F | GGA GAC CCA AAC TGC TAT TAC A |
| Indel 2 451470 R | AAC CGC CTC CAT TTG CAC CTT ATC |
| Indel 3 469762 F | GTC ACC GAG TTT TGC TTT GTT CAT |
| Indel 3 469762 R | CTC GTT TCT TTT CTG GGC TTG TAG |
| Indel 4 449053 F | GAA AGA AAG CAG CGA AAG ACA |
| Indel 4 449053 R | GCC CAT GCC CAT ACA CTG A |
| Indel 5 455100 F | ACT TGC TTA ATC GTT TCT TTG TA |
| Indel 5 455100 R | GCC CAC TCG TAT TCG CTT AG |
Gene copy analysis using qPCR
Quantitative PCR reactions were set up to measure gene copy number using 2.5 pmole gene-specific primers, 300 ng of genomic DNA diluted in DNase/RNase free water, and iQ SYBR Green supermix (Bio-Rad, Hercules, CA, USA) for a total volume of 10 μL. Reactions without template were used as negative controls. Each single copy reaction was set up in triplicate and run in a Bio-Rad iCycler for 40 cycles. Threshold cycles (CT) were calculated using iQ software (Bio-Rad).
Primer efficiencies (E) were calculated using cDNAs synthesized from RNA isolated from Col-0 plants as previously described11. cDNA samples were serially diluted across three orders of magnitude. Serial dilutions were amplified in triplicate using the same protocol as for the copy number experiment. The CTs of each triplicate were averaged and plotted against the dilution factor. A linear trend was fitted to the data and the slope of this trend was used to calculate E for each primer with the formula: E=10(1/-slope).
Copy number of VTC1 (AT2G39770) was calculated using the formula: Reported Quantity (RQ) = 1/ECT normalized to the RQ of a known single copy gene (PAD4, AT3G52430;20,21). VTC1 RQ was calculated from the average VTC1 RQ of three biological replicates per genotype and was normalized to the average RQ of PAD4 from three replicates of each respective genotype, all run in the same reaction plate.
Sequencing analysis
PCR products were purified using the Qiagen Miniprep Kit. Dye-terminator based DNA sequencing was performed at the Genomics Facility in the Department of Biology at West Virginia University. Sequence alignments were performed using the BioEdit program (http://www.mbio.ncsu.edu/bioedit/bioedit.html).
Ascorbic acid quantification
To screen mutants, AA levels were analyzed qualitatively in small pieces of two-week-old rosette leaves using the nitroblue tetrazolium assay previously described19. The AA content was determined in whole rosettes of three-week-old plants using the iron reduction assay4.
Statistical analysis
Experiments were performed at least three times. Figures represent individual experiments. Data were expressed as mean values ± SE. P values were determined by Student’s t test analysis.
Results
Isolation of svt2
Our laboratory is interested in understanding how the VTC1 gene, which is essential for the biosynthesis of GDP-mannose and AA, is regulated. This would help deciphering the pleiotropic phenotypes displayed by vtc1-1, including its hypersensitivity to ammonium8–11. We employed a gene suppressor analysis with the goal of identifying novel genes that interact or regulate VTC1. Seed of the vtc1-1 mutant, which is in the Col-0 genetic background14, were subjected to chemical mutagenesis using EMS18. One thousand and one hundred mutagenized vtc1-1 seeds (M0 generation) were planted onto soil and screened for recovered (wild-type) leaf AA content using the qualitative nitroblue tetrazolium test19. One of the mutants exhibited wild-type AA levels in the M0 generation. This mutant was named svt2 (suppressor of vtc1-1 2), isolated, and further characterized. The mutant was allowed to self-fertilize and seeds from the plant were collected (M1 generation) (Figure 1). Note that we isolated additional suppressor mutants by pooling M2 seed and by screening for long roots on 1× Murashige and Skoog (MS) medium containing ammonium. Six suppressor mutants were identified among 2000 plants. M3 seed were collected and screened for long roots again to test for segregation. M4 progeny of one line had all long roots, whereas the other five lines segregated in a ratio of three plants producing long roots, and one plant having short roots. Figure 2 summarizes data of four of these suppressor mutants, with D3–4 homogenously producing long roots, whereas D3–3, D3–7, and D3–15 developed long and short roots in a 3:1 ratio. As is illustrated in Figure 2A, these suppressor mutants developed roots that were significantly longer than those of the Col-0 wild type. Analysis of the total AA content revealed that the suppressor D3–4 had an AA content comparable to the Col-0 wild type, whereas that of vtc1-1 was only approximately 40% of that of the wild type (Figure 2B)14,15. Finally, sequence analysis of these four suppressor mutants demonstrated a lack of the vtc1-1 mutation (Figure 2C). Except for the assessments described above, these suppressor mutants were not yet characterized further.
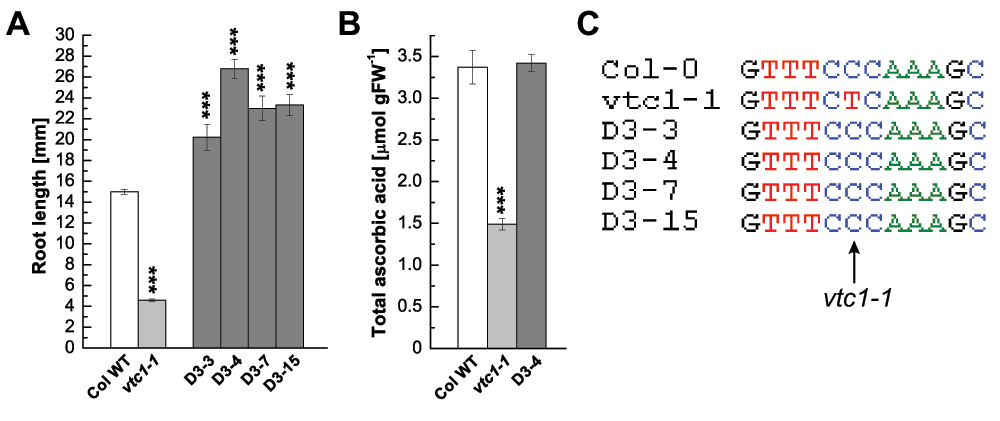
Figure 2. Phenotypic and genotypic characterization of additional vtc1-1 suppressor mutants.
(A) Root length in seven-day-old seedlings grown on 1× MS. Bars represent means ± SE of 18–73 individuals. Since D3-4 homogenously produced long roots, all individuals were included in the calculations. As D3-3, D3-7, and D3-15 developed long and short roots in an approximate 3:1 ratio, only individual seedlings that produced long roots were included in the calculations. (B) Total ascorbic acid content per gram fresh weight in whole rosettes of three-week-old plants. Bars represent means ± SE of three (Col-0 and vtc1-1) or 24 individual replicates. ***P < 0.001 by Student’s t-test indicates significant differences in comparison to the Col-0 wild type. (C) Sequences of the Col-0 wild type, the vtc1-1 mutant and four suppressor mutants. The arrow points to the vtc1-1 mutation, a conversion of cytosine to a thymine.
svt2 has similarities with the Ler phenotype, but has also phenotypic characteristics that are distinct from Ler
The first observation we made when characterizing svt2 M1 plants was that svt2 exhibited a phenotype reminiscent of the Ler ecotype with the characteristic round leaves and erect morphology when compared to Col (Figure 3A). However, svt2 also had features that were distinct from the Ler phenotype, including overall enlarged vegetative and reproductive morphology (insets of rosettes and flowers in Figure 3A). In addition, svt2 was strongly delayed in flowering compared to the Col-0 and Ler-0 wild types and the vtc1-1 mutant (Figure 3A, 3B). Primary inflorescences in four-week-old plants were 1.4-times significantly longer in the vtc1-1 mutant and approximately twice as long in the Ler-0 wild type compared to the Col-0 wild type. In svt2 mutant plants, however, buds of primary inflorescences only began to emerge when plants were four weeks old (Figure 3A, 3B). The flowering data are consistent with previous reports, with Ler-0 wild type entering the reproductive phase before Col-0 wild type. An early flowering phenotype of vtc1-1 has been reported previously3.
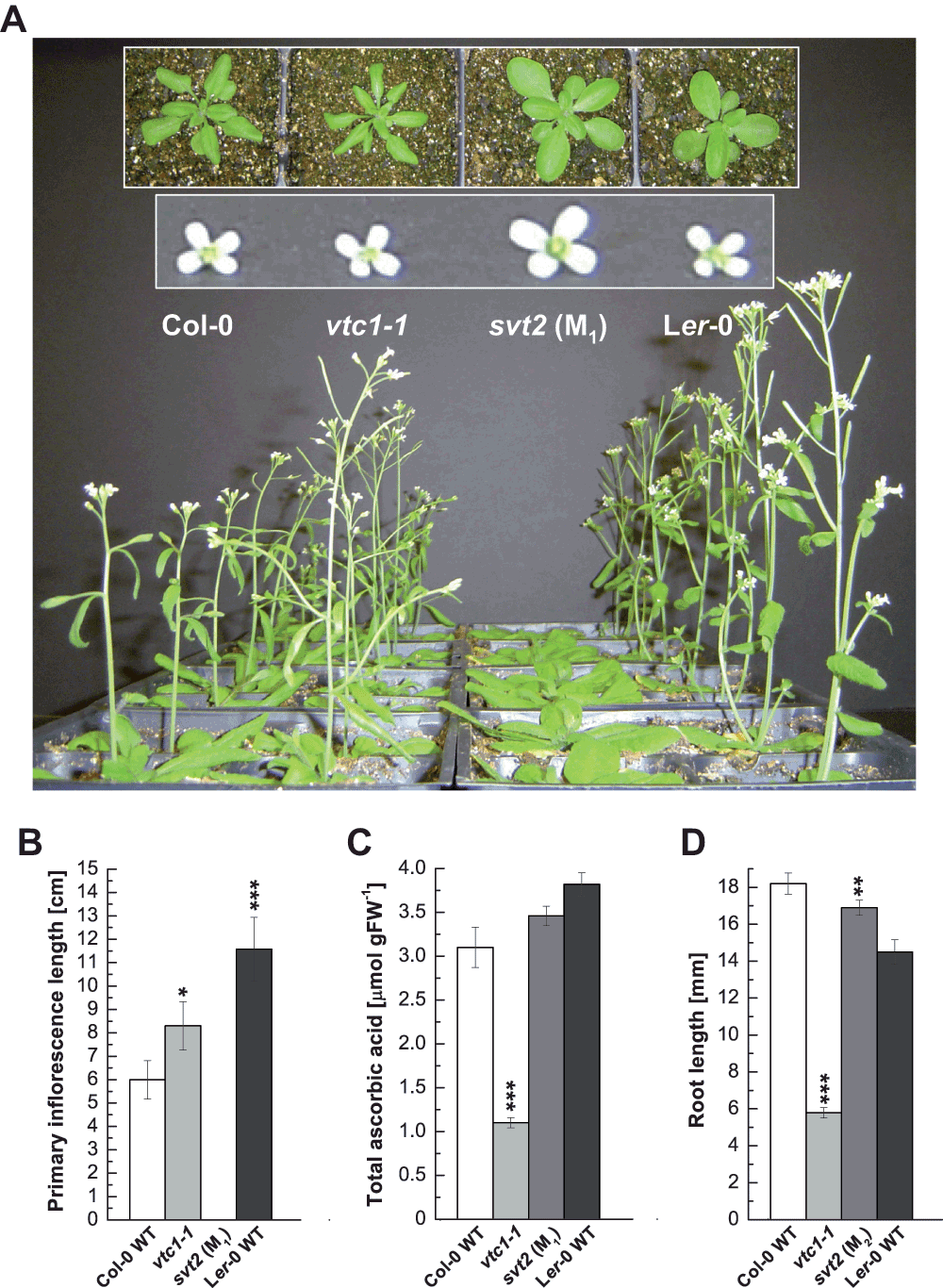
Figure 3. Phenotypic characterization of svt2.
(A) Flowering phenotype of four-week-old Col-0 wild type, the vtc1-1 and svt2 mutants and the Ler-0 wild type. Insets show rosette phenotypes of the four genotypes when plants were three weeks old and the flower phenotype of six-week-old plants, respectively. (B) Primary inflorescence length when plants were four weeks old. Bars represent means ± SE of eight individual replicates. (C) Total ascorbic acid content per gram fresh weight in whole rosettes of three-week-old plants. Bars represent means ± SE of three individual replicates. (D) Root length in seven-day-old seedlings grown on 1× MS. Bars represent means ± SE of 30–90 individuals. *P < 0.05, **P < 0.01, ***P < 0.001 by Student’s t-test indicate significant differences in comparison to Col-0 and Ler-0 wild type, respectively.
The AA content in svt2 was similar to levels quantified in Col-0 and Ler-0 wild types, whereas vtc1-1 contained approximately 30% of the AA content as expected14,15 (Figure 3C). Finally, we investigated whether svt2 also exhibits recovered root development in the presence of ammonium by growing the four genotypes in full strength 1× MS medium. The vtc1-1 mutant is conditionally hypersensitive to ammonium8,9,11. Figure 3D illustrates that root length in svt2 was the same as in Col-0 wild type, whereas root development was strongly inhibited in vtc1-1 as expected.
The enlarged morphology of svt2 raises the question as to whether svt2 is polyploid. In order to test this, we assessed the size of pollen grains from the Col-0 and Ler-0 wild-types and vtc1-1 and svt2 mutants. As is shown in Figure 4, pollen grains of the four genotypes are similar in size. In addition, using qPCR, we determined the number of VTC1 gene copies in the four genotypes. Our results revealed that VTC1 is present as a single copy gene in both the Col-0 and Ler-0 wild types and in the vtc1-1 and svt2 mutants (Table 2). Although an extensive chromosome analysis has not yet been performed in svt2, our results suggest that the mutant does not contain additional sets of chromosomes.
Table 2. Quantitative PCR to verify that VTC1 is a single copy gene in Col-0 and Ler-0 wild types and vtc1-1 and svt2 mutants.
Quantitative PCR was performed as described in Materials and Methods. The PAD4 gene is a known single copy gene. Therefore, an RQ/RQ ratio of approximately 1 indicates that VTC1 is present in similar quantity as PAD4, and therefore a single-copy gene.
| RQ/RQ | |
|---|
| VTC1/PAD4 |
|---|
| Col-0 WT | 0.3796 | VTC1
E=1.8 |
| vtc1-1 | 0.5843 |
| svt2 | 0.5504 |
| Ler-0 WT | 0.6329 |
| Col-0 | 0.3153 | VTC1
E=2 |
| vtc1-1 | 0.5292 |
| svt2 | 0.4946 |
| Ler-0 WT | 0.5807 |
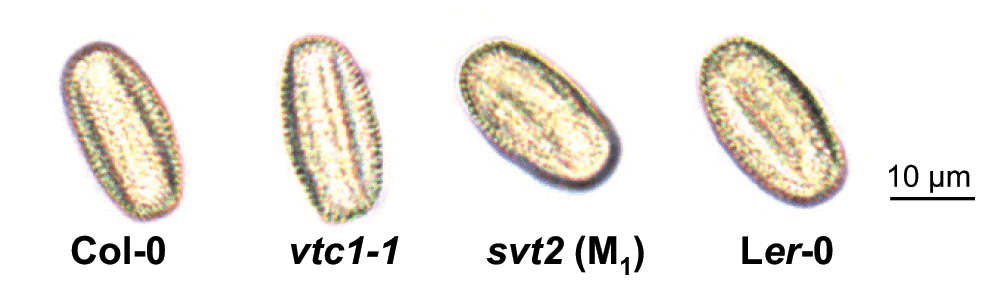
Figure 4. Pollen phenotype of Col-0 wild type, the vtc1-1 and svt2 mutants and the Ler-0 wild type.
Light images were taken when plants were 4.5 weeks old. Scale bar represents 10 µm.
Taken together, based on the phenotypic observations, our data suggest that svt2 represents a novel vtc1-1 suppressor mutant with recovered AA content and root development. Next, we characterized svt2 genotypically in order to determine whether svt2 represents an intragenic or extragenic suppressor.
svt2 shares genome similarity with Ler
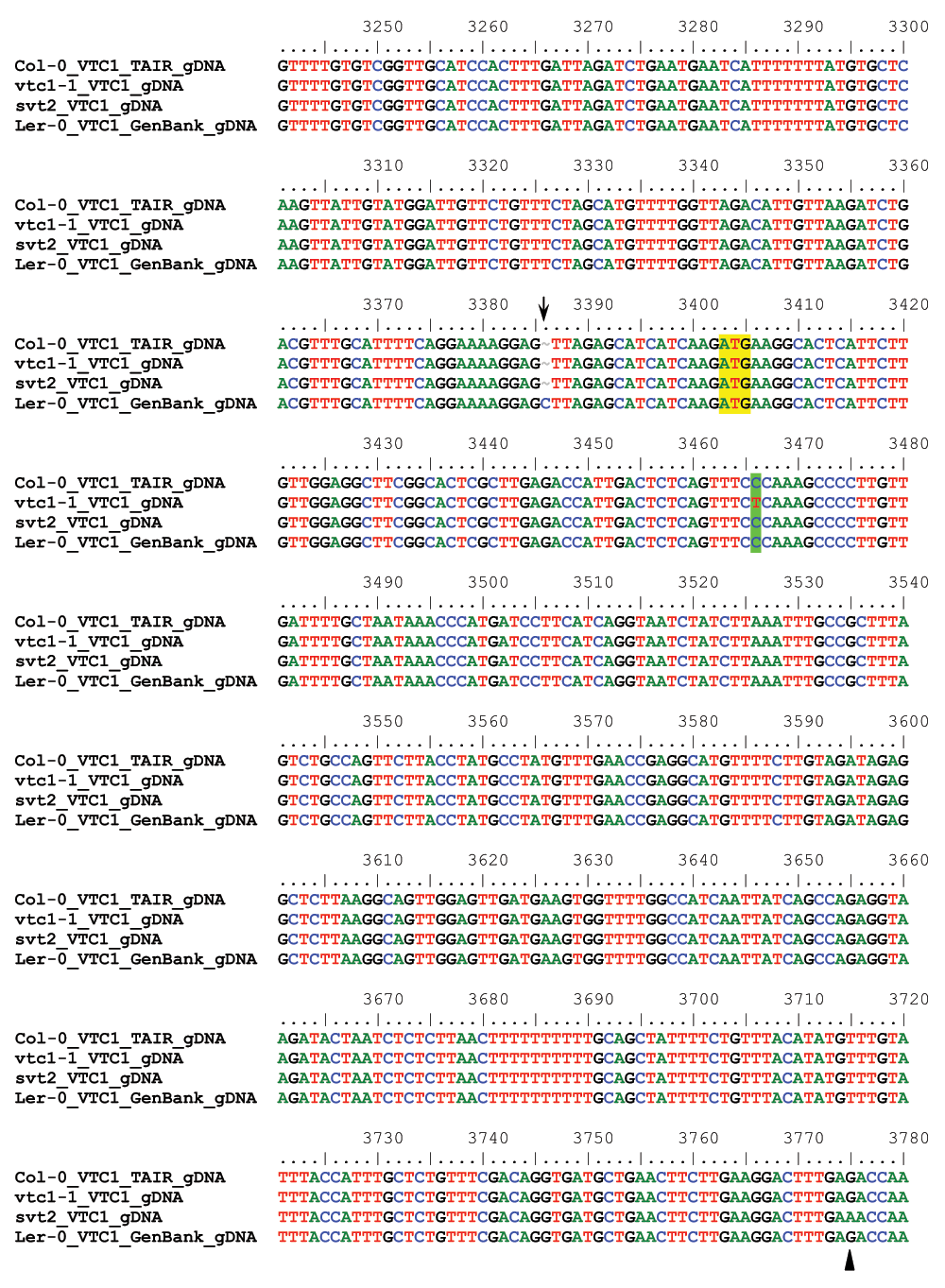
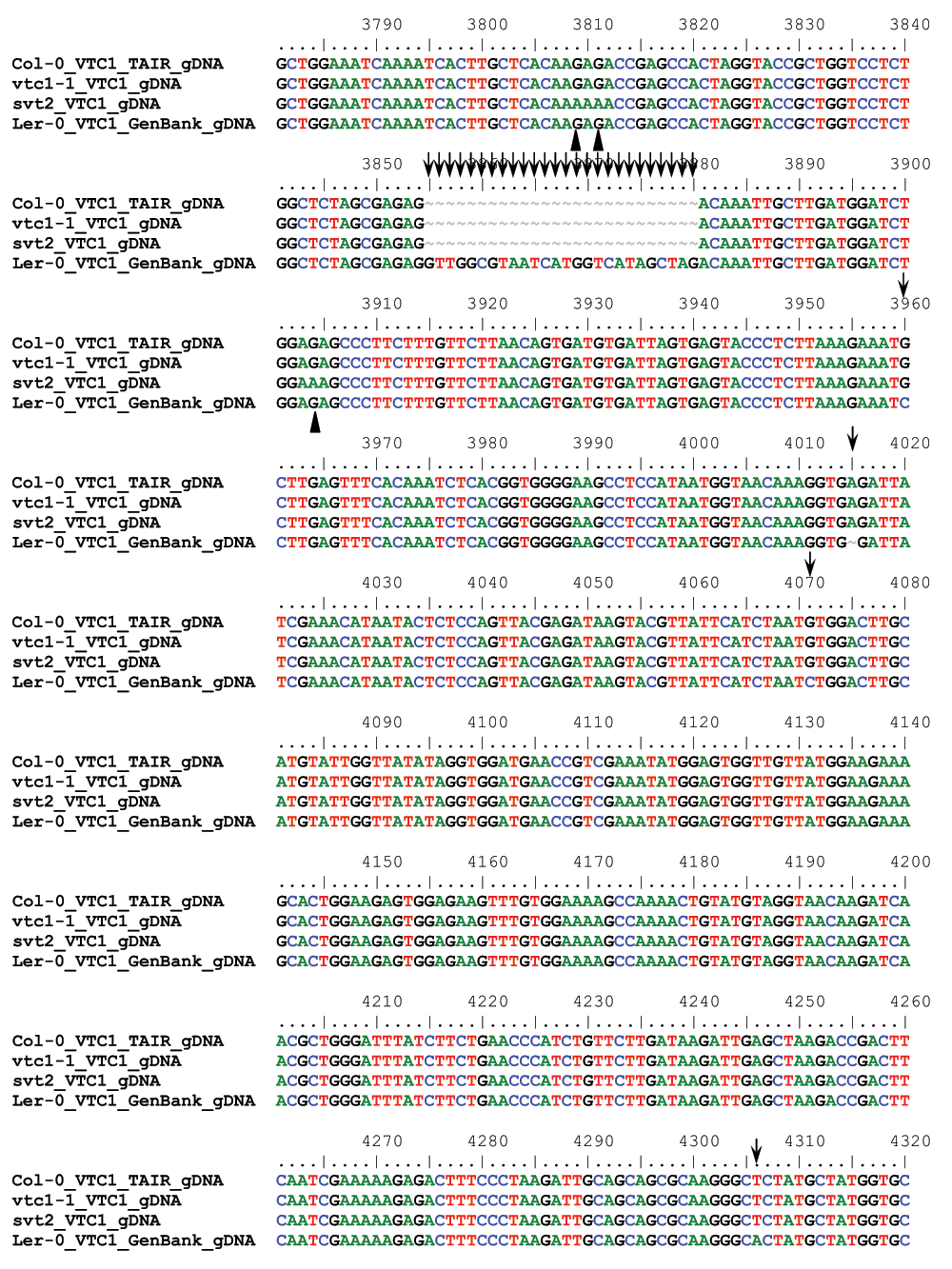
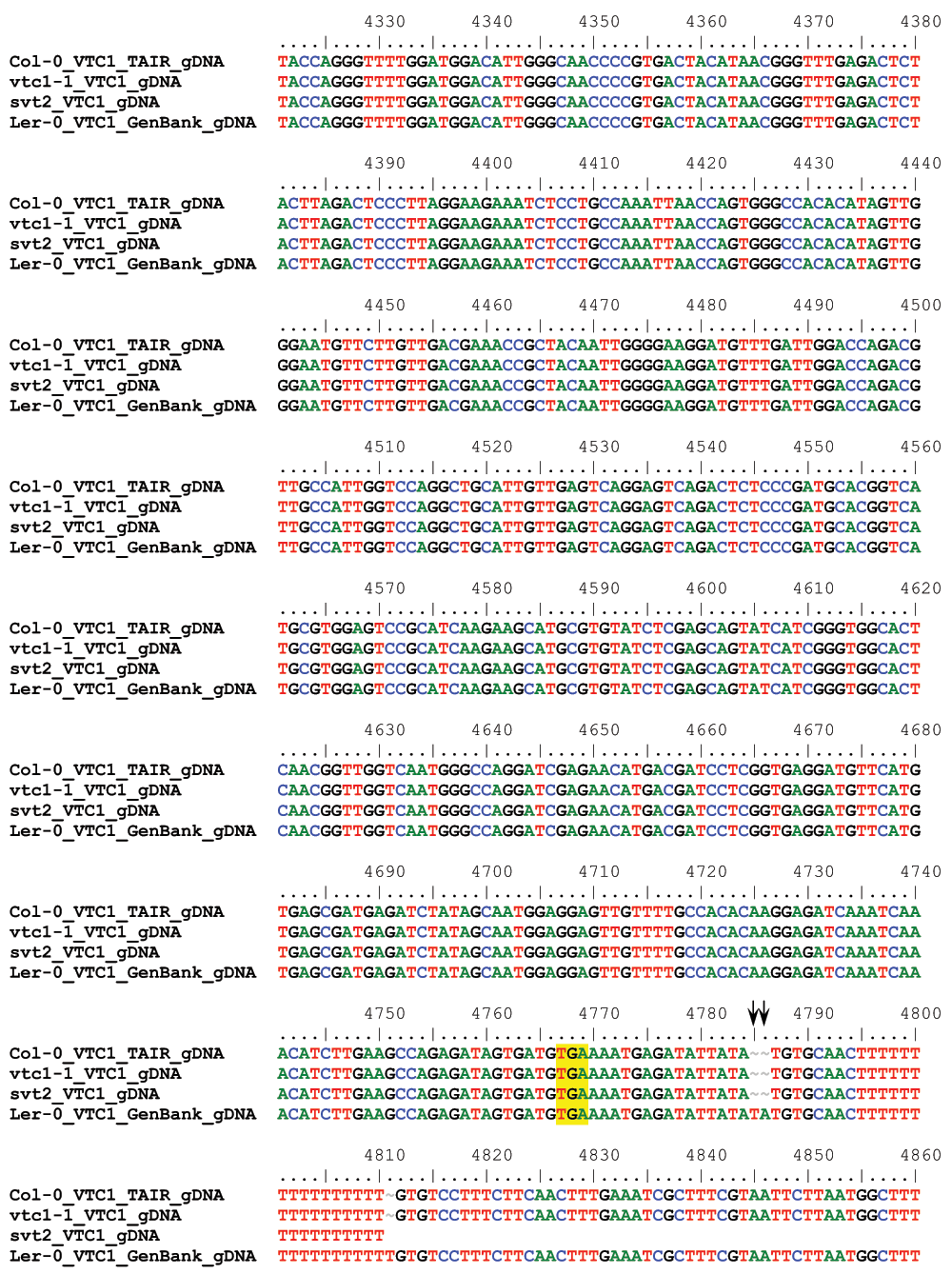
To determine whether svt2 represents an intragenic suppressor, i.e., to test whether the suppressor mutation is present within the VTC1 gene, we designed nine overlapping PCR primer sets spanning the entire VTC1 gene and approximately 500 bp of the promoter region directly upstream of the first base in the 5’ UTR (Table 1, Figure 5A). PCR products were generated from genomic DNA extracted from Col-0 and Ler-0 wild types, and vtc1-1 and svt2 mutants. In eight of the nine primer pairs covering the entire VTC1 gene, the PCR products generated using svt2 genomic DNA had the same electrophoretic mobility as those generated using Col-0 wild-type genomic DNA. However, this was not the case for the first primer set. The G1F/G1R primer set, used to amplify the VTC1 promoter region, generated a larger PCR product in svt2 than in the wild type (Figure 5B, Figure 6). The PCR product in the wild type was 567 bp, whereas that in svt2 had a size of approximately 850 bp, suggesting that svt2 contained an approximately 300 bp insertion in this region. We repeated the PCR analysis of the VTC1 promoter region using the G1F-G1R and the G1F-G2R primer sets that should generate a PCR product of 567 bp and 751 bp, respectively (Figure 5A). The expected size was obtained for the Col-0 wild type and the vtc1-1 mutant. However, approximately 300 bp larger PCR products were detected in the svt2 mutant and the Ler-0 wild type (Figure 5B), suggesting a Ler insertion polymorphism. Thus, these data imply that svt2 shares both phenotypic and genotypic similarities with Ler.
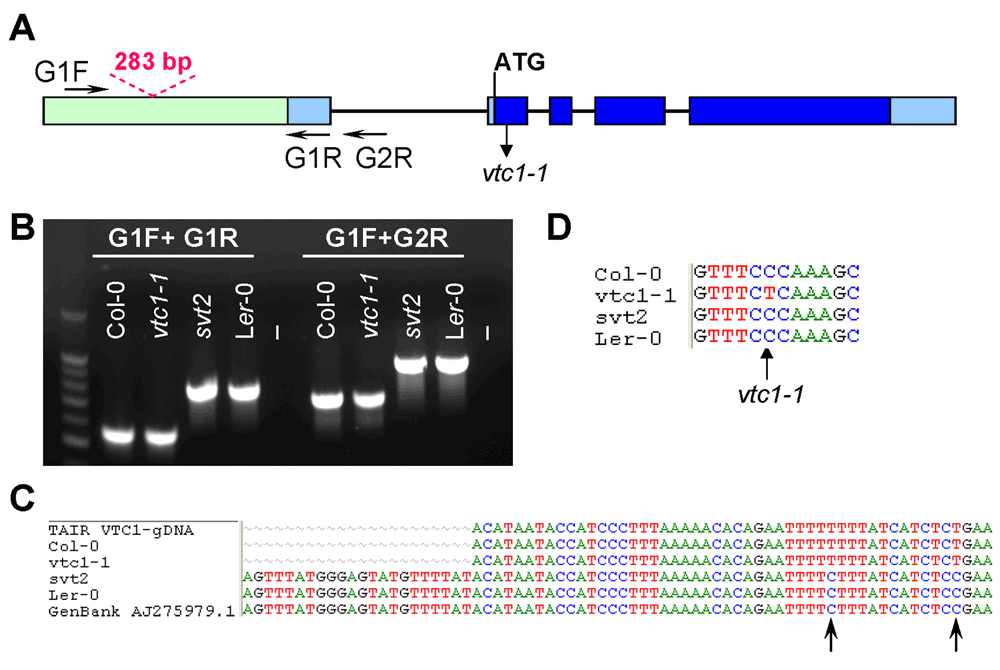
Figure 5. Genotypic characterization of svt2.
(A) VTC1 Col-0 gene model. Light green box indicates VTC1 gene promoter region, light blue rectangles indicate 5´ and 3´ UTRs, dark blue rectangles indicate exons, and lines indicate introns. Shown is the location of the vtc1-1 mutation within the first exon, primer locations, and polymorphism insertion of 283 bp in Ler-0 VTC1. (B) PCR amplification of the VTC1 promoter region in the Col-0 wild type, vtc1-1 and svt2 mutants and Ler-0 wild type. (-) indicates negative control, no DNA. (C) Partial sequence alignment of the VTC1 promoter region from the TAIR database (Col-0), sequenced Col-0 wild type, vtc1-1 and svt2 mutants, sequenced Ler-0 wild type and the Ler-0 sequence obtained from GenBank. The alignment shows the sequence insertion in the svt2 mutant, the Ler-0 wild type and the GenBank sequence. Arrows indicate single nucleotide polymorphisms between the Ler-0 and Col-0 sequence. (D) Point mutation in vtc1-1, a conversion from a cytosine to a thymine.
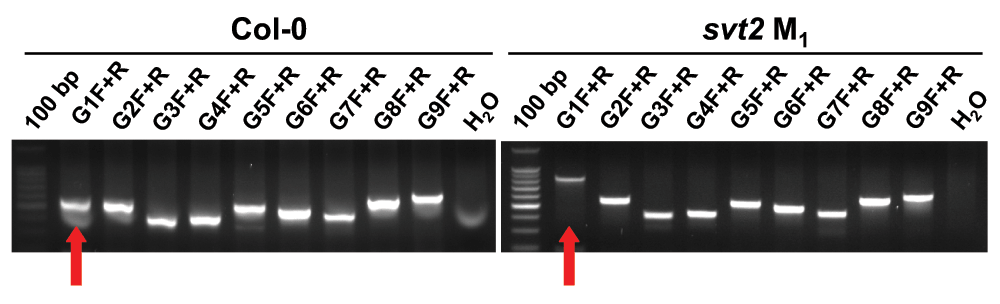
Figure 6. Molecular characterization of svt2.
Amplification of the VTC1 gene including ~500 bp of the promoter region using a series of nine, overlapping primers (G1F+R through G9F+R) in both Col-0 wild type and svt2 M1 mutant genomic DNA. The last lane in each gel contained a negative control (water instead of DNA). Red arrows indicate the different sized PCR products using the same primer set.
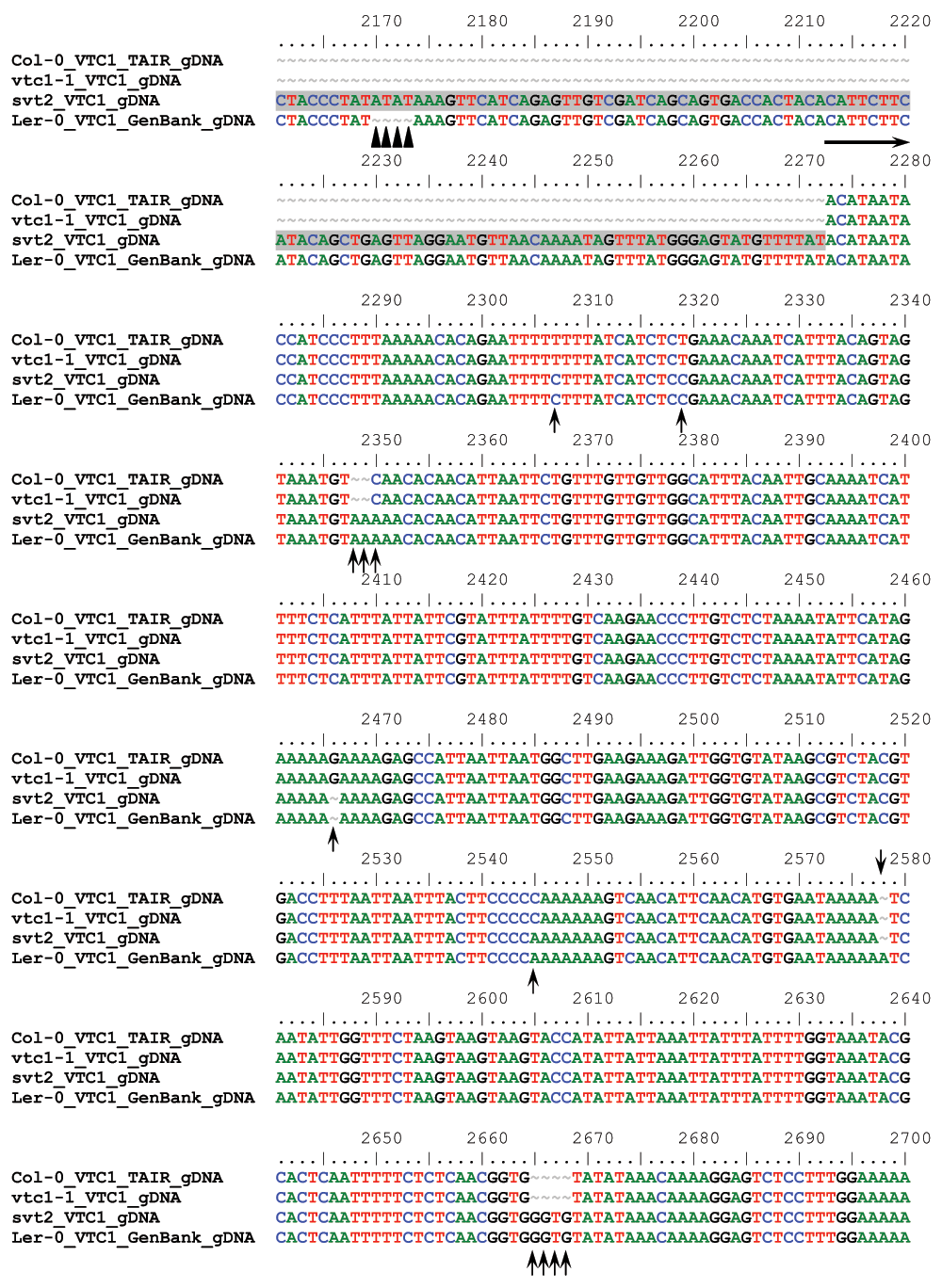
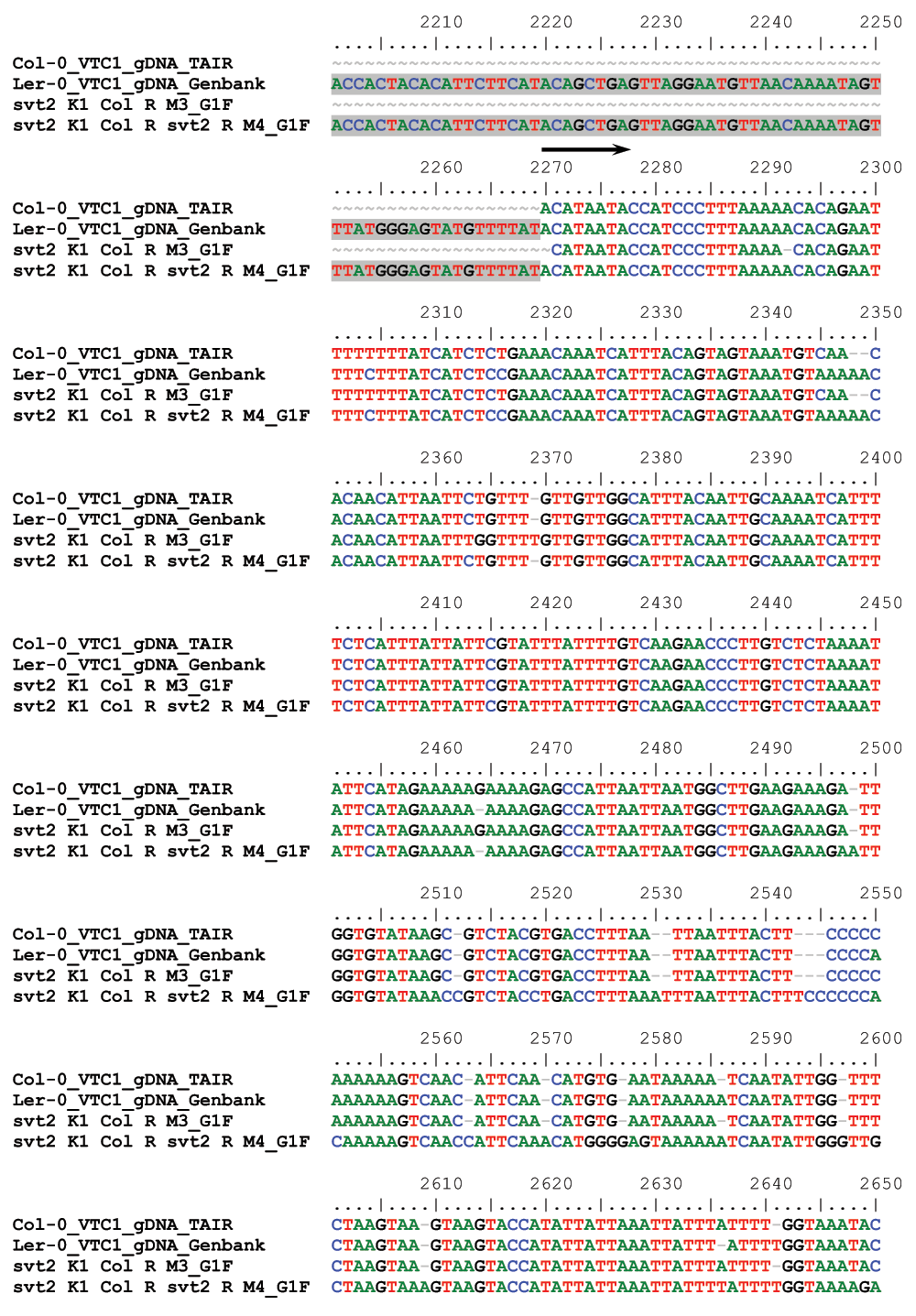
We therefore assessed five additional Insertion/Deletion (InDel) polymorphisms randomly chosen across the five Arabidopsis chromosomes (Table 1) in svt2 compared to the Col-0 and Ler-0 wild types and sequenced the entire VTC1 gene and the promoter region tested. Our data show that the PCR products generated for those five InDels using svt2 genomic DNA had the same electrophoretic mobility as those produced from Ler-0 genomic DNA (Figure 7). Moreover, sequence analysis of the VTC1 gene and promoter region revealed that svt2 contained a 283 bp insertion in the VTC1 promoter (Figure 5C). The insertion is highlighted in gray in Figure S1. Note additional single nucleotide polymorphisms as indicated by upright arrows in Figure 5C and Figure S1. When we aligned the VTC1 gene sequence obtained from svt2 with that of the vtc1-1 mutant, the VTC1 Col-0 gene sequence deposited in the TAIR database, and the VTC1 Ler GenBank sequence, the VTC1 gene sequence in svt2 shared similarities with Ler (upright arrows in Figure 5C, Figure S1) and Col (arrows pointing down in Figure S1). However, note that there are sequences that are unique to svt2 and are not shared between Col, vtc1-1 or Ler (arrowheads in Figure S1). Finally, note the overlap in sequences between Col, vtc1-1, svt2 and Ler on the 5´ end of the sequence flanking the insertion (at approximately base pair 1990); see left-facing horizontal black arrow in Figure S1 compared to the sequence flanking the 3´ end of the DNA sequence insertion (starting at base pair 2273); see right-facing horizontal black arrow in Figure S1.
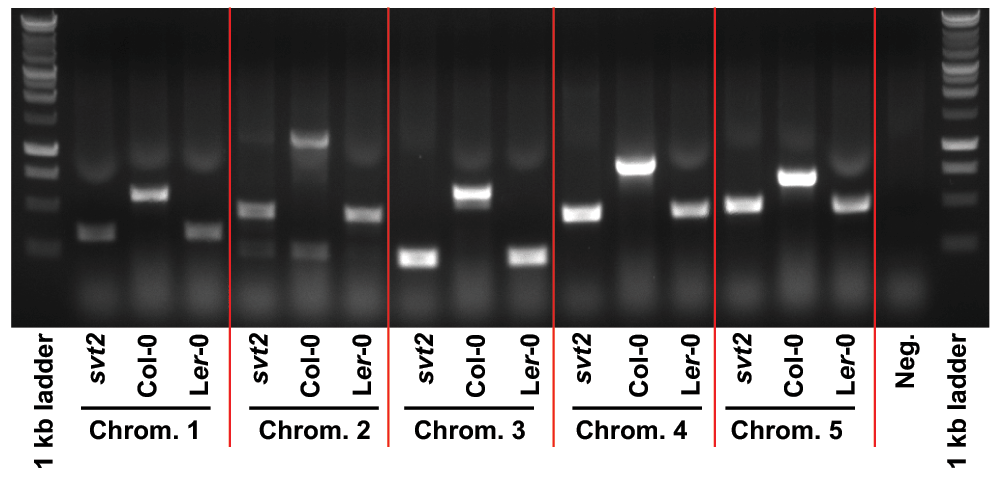
Figure 7. Insertion/Deletion polymorphism analysis in svt2, Col-0 and Ler-0.
Primers were designed for five randomly selected InDel polymorphisms across the five Arabidopsis chromosomes. The polymorphisms represent insertions in Col-0 and deletions in Ler.
Finally, most intragenic suppressor mutants still contain the original mutation in addition to the suppressor mutation. Therefore, we expected that the vtc1-1 mutation is still present in svt2. However, our sequencing analysis demonstrated that svt2 did not contain the vtc1-1 mutation anymore and that the mutation reverted back to the homozygous wild-type allele (Figures 5D; green shading in Figure S1).
In summary, our data demonstrate that svt2 shares DNA sequence similarity with Col and Ler, but also contains DNA sequences that are unique to this mutant. This is particularly remarkable because svt2 was generated in the vtc1-1 Col-0 background. Also, svt2 did not contain the original vtc1-1 mutation anymore. Although our data already argue against svt2 being a result of an artifact of the experiment or a contamination with Ler, we analyzed subsequent svt2 generations and discovered additional characteristics that are unique to svt2.
svt2 exhibits phenotypic and genotypic instability
Our initial observations revealed that approximately 10% of svt2 M2 plants displayed a Col-like phenotype. Therefore, we planted svt2 M1, M2, and M3 progeny to check whether this result could be repeated and to determine segregation ratios (Table 3). Additionally, we investigated whether svt2 progeny that were phenotypically Col-like revertants would produce svt2 (Ler-like) offspring in the next generation.
Table 3. Summary of revertant data.
The table summarizes the number of plants screened in each of three svt2 generations (M1, M2 and M3), screens of revertant progeny from Col-like revertants (A8, G7, K1), and the revertant progeny of a Ler-like line (K1 Col R svt2 R). The percent reversion is shown in the last column. Although the number of progeny plants tested is relatively large, some lines did not give rise to revertant progeny. R denotes revertant. *indicates mutant plants that were also analyzed genotypically (see Table 4).
| Experiment | Generation | Total # of plants | # of phenotypic revertants | % reversion |
|---|
| 1 | svt2 M1 | 63 | 0 | 0 |
svt2 M2, 3 of 7 revertants tested further:
svt2 A8 Col R M3
svt2 G7 Col R M3
svt2 K1 Col R M3*
svt2 K1 Col R svt2 R M4* | 78
64
64
63
96 | 7 (Col phenotype)
0
0
1 (svt2 phenotype)
0 | 8.97
0
0
1.58
0 |
| svt2 M3 | 96 | 0 | 0 |
| 2 | svt2 M1 | 96 | 1 (Col phenotype) | 1.04 |
svt2 M2, 2 of 5 revertants tested further:
svt2 Col R1 M3*
svt2 Col R4 M3* | 62
88
96 | 5 (Col phenotype)
20 (svt2 phenotype)
0 | 8.06
22.73
0 |
| 3 | svt2 M2 | 96 | 10 (Col phenotype) | 10.42 |
As summarized in Table 3, revertants could only be detected when a relatively large population was planted. In the svt2 M1 generation, only 1% of Col-like revertants were detected. In contrast, 8–10% of svt2 M2 plants displayed a Col-like phenotype, whereas no revertants were detected in the svt2 M3 generation. These Col-like revertants were isolated and seeds were collected from individual plants and the phenotype of the progeny in the M3 generation was assessed in some examples. In most cases, reversion appeared to be stable, i.e., once svt2 plants reverted, displaying a Col-like phenotype in the M2 generation, their M3 progeny continued to appear as Col-like plants. This was the case for the M3 progeny of the A8 and G7 plants listed in Table 3. However, out of 63 progeny from the K1 revertant plant, one reverted back to a svt2-like phenotype (Table 3), i.e., the K1 double revertant switched from svt2 phenotype in the M1 generation to a Col-like phenotype in the M2 generation, and then reverted back to a svt2-like phenotype in the M3 generation. Note that only a small number of progeny was planted. In a second experiment, the svt2 Col R1 revertant produced 20 individuals displaying a svt2-like phenotype (Table 3). This represents a larger reversion percentage than in the K1 double revertant (22.7% vs. 1.6%). This may be explained by the genotypic make-up of the Col-like reverted parents and will be presented in the next section. Figure 8 illustrates the phenotypic appearance of three examples of svt2 → Col single revertants (Col R1, Col R2, K1 Col R) and a svt2 → Col → svt2 double revertant (K1 Col R svt2 R).

Figure 8. Phenotype of svt2 revertants.
Plants were three weeks old when photographs were taken. Top row represents controls, Col-0 wild type, vtc1-1 and svt2 mutants, and Ler-0 wild type. Bottom row represents three Col-like revertants, svt2 Col R1 M3, svt2 Col R2 M3, svt2 K1 Col R M3, and a double revertant, svt2 K1 Col R svt2 R M4. R stands for revertant.
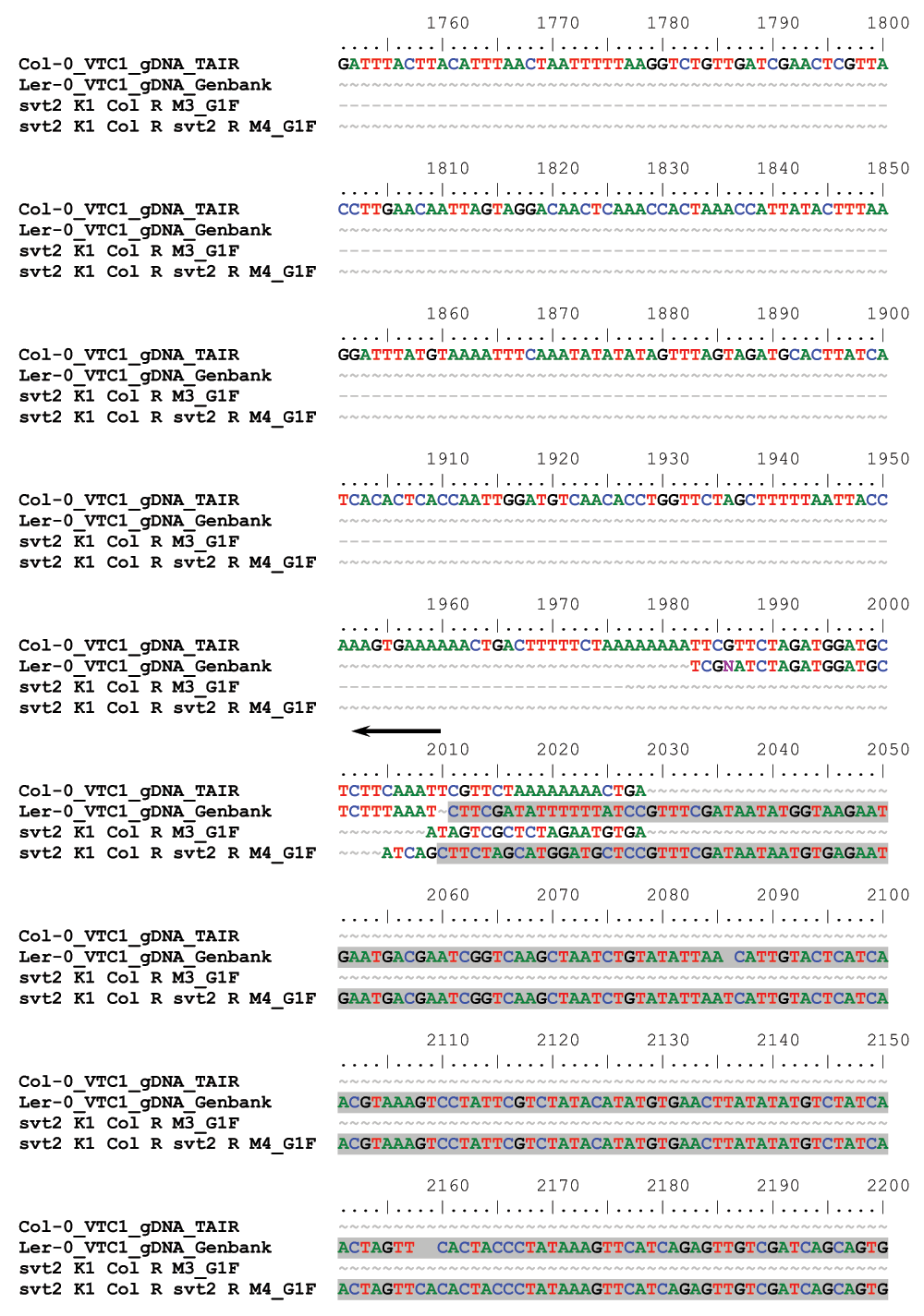
Next we tested whether a Col-like revertant phenotype correlated with a Col-like genotype. Likewise, we would expect that a svt2 → Col → svt2 double revertant phenotype corresponds with svt2-like genomic markers. To check this we isolated genomic DNA from Col-0 and Ler-0 wild types, svt2, vtc1-1 and revertant mutants, and PCR-amplified the five randomly selected InDel polymorphisms plus the InDel polymorphism in the VTC1 promoter (Table 1). In all cases but the svt2 M2 Col R1 revertant, the svt2-like revertant plants (labeled svt2 M2 Col revertants 1 through 5) produced PCR products that where of the same electrophoretic mobility as the PCR products generated using Col-0 wild-type genomic DNA. In contrast, svt2 M1 plants and svt2 M2 plants that displayed an svt2 phenotype, gave rise to PCR products that were of the same electrophoretic mobility as those of the Ler wild type (Table 4, Figure 9). In addition, the double revertant plant K1 (labeled svt2 M2 K1 Col R) was genotyped in both its M2 and M3 generations. The K1 plant produced InDel PCR products similar to those of the Col-0 wild type in the M2 generation. However, the M3 generation that displayed svt2-like morphology produced PCR products that were comparable to the InDel PCR products generated using Ler genomic DNA (Table 4). The svt2 M2 Col R1 (highlighted in red in Table 4 is intriguing, because it appears to contain DNA that is similar to both Col and Ler genomic DNA. This suggests the presence of chimeric genome sectors, which may explain the higher percentage of Col-like revertants compared to svt2 M2 K1 Col R. Note that the PCR results are in line with the sequencing analysis of the revertants. That is, Col-like revertants and svt2-like revertants share sequence similarity with Col-0 and Ler wild type, respectively (Figure S2).
Table 4. Summary of PCR-based molecular genotypes.
With the exception of svt2 Col R1 M2, where Col and Ler markers and one heterozygous marker were found (highlighted in red), phenotype matched genotype. That is, a Col-like phenotype correlated with the presence of Col polymorphisms, while a Ler-like phenotype correlated with Ler polymorphisms. C, L, and H refer to Col, Ler, or heterozygous, respectively. R denotes revertant. n.d., not detected.
| Genotype | InDel 1
450919 | InDel 2
451470 | InDel 3
469762 | InDel 4
449053 | InDel 5
455100 | G1F +
G2R VTC1 |
|---|
| Col-0 WT | C | C | C | C | C | C |
| vtc1-1 | C | C | C | C | C | C |
| Ler-0 WT | L | L | L | L | L | L |
| svt2 M1 | L | L | L | L | L | L |
| svt2 M2 | L | L | L | L | L | L |
| svt2 Col R1 M2 | C | L | C | H | C | C |
| svt2 Col R2 M2 | C | C | C | C | C | C |
| svt2 Col R3 M2 | C | C | C | C | n.d. | C |
| svt2 Col R4 M2 | C | C | C | C | C | C |
| svt2 Col R5 M2 | C | C | C | C | C | C |
| svt2 K1 Col R M2 | C | C | C | C | C | C |
svt2 K1 Col R
svt2 R M3 | L | L | L | L | L | L |

Figure 9. Insertion/Deletion polymorphism analysis in svt2, Col-0, Ler-0, and revertants.
PCR amplification of the Col/Ler VTC1 promoter polymorphism in svt2 plants and svt2 revertant (R) plants, amplified with the VTC1 G1F and G2R primers. (-) indicates negative control, no DNA.
Taken together, these data suggest (i) transgenerational phenotypic and genotypic instability in svt2, and that (ii) svt2 offspring do not segregate in a Mendelian fashion. In an attempt to obtain first insights toward a mechanism that is causing this genotypic instability, we investigated whether transgenerational epigenetic inheritance could play a role.
Genome instability in svt2 does not appear to be triggered by a transgenerational epigenetic mechanism
To investigate whether genome instability is caused by transgenerational epigenetic inheritance in the svt2 mutant, we performed reciprocal crosses between svt2 mutants and Col-0 wild-type plants. It is possible that through the EMS mutagenesis of vtc1-1 seeds, genes involved in the regulation of epigenetic alterations were altered, whereby their activity was affected. There is increasing evidence in both plants and animals that epigenetic marks are not always cleared between generations. Incomplete erasure at genes associated with a measurable phenotype results in unusual patterns of inheritance from one generation to the next, termed transgenerational epigenetic inheritance22,23. Therefore, analysis of the progeny of the reciprocal crosses is expected to provide some first insights on the possibility of transgenerational epigenetic inheritance that is transmitted maternally. If this were the case, only progeny of crosses with a maternal svt2 donor should have a svt2-like phenotype. To determine the genotypes of the F1 progeny of the reciprocal crosses, we performed another InDel polymorphism assay as described above. In addition, progeny were also screened using the VTC1 InDel promoter polymorphism. Table 5 contains a summary of the InDel screen for progeny from each reciprocal cross. In all but six of the progeny from the reciprocal crosses, PCR products similar to those obtained using Col and Ler genomic DNA, respectively, were generated, suggesting that the F1 of the reciprocal crosses were heterozygous. A similar result was obtained for the VTC1 promoter polymorphism marker in all reciprocal crosses. Note, however, that for some polymorphisms and irrespective of whether svt2 or Col-0 served as female or male donor, respectively, PCR products comparable to those obtained using Ler-0 wild-type DNA were prevalent (highlighted in red in Table 5). This is surprising because heterozygosity was expected at all loci. This suggests that some parts of the genome were not inherited equally from both parents. Taken together, these results suggest that maternal epigenetic inheritance may not be the cause of genome instability in svt2. However, at some loci svt2-like alleles dominate over Col-0.
Table 5. Reciprocal crosses between svt2 and Col-0 wild-type lines.
Molecular analysis of the InDel polymorphism markers showed evidence of cryptic but persistent homozygosity, irrespective of the direction of the sexual cross (highlighted in red). However, heterozygosity was expected at all loci.
Female × Male
crosses | InDel 1
450919 | InDel 2
451470 | InDel 3
469762 | InDel 4
449053 | InDel 5
455100 | G1F + G2R
VTC1 |
|---|
| svt2 × Col-0 F1 1 | H | H | L | H | H | H |
| svt2 × Col-0 F1 2 | H | H | L | H | H | H |
| svt2 × Col-0 F1 3 | H | H | H | H | H | H |
| svt2 × Col-0 F1 4 | H | H | H | H | H | H |
| Col-0 × svt2 F1 1 | H | H | H | L | H | H |
| Col-0 × svt2 F1 2 | H | H | L | H | H | H |
| Col-0 × svt2 F1 3 | H | H | H | L | H | H |
| Col-0 × svt2 F1 4 | H | H | L | H | H | H |
Discussion
The svt2 mutant was initially identified as a putative suppressor of the AA-deficient Arabidopsis mutant vtc1-1, as was evident in wild-type levels of AA (Figure 3C) and recovered root development in the presence of ammonium (Figure 3D). However, svt2 manifests other interesting characteristics, including genotypic and phenotypic instability. These unique features could aid in our understanding of the complex mechanisms controlling genome instability and restoration.
svt2 is a novel Arabidopsis mutant and not a result of an experimental artifact, seed contamination, or outcrossing
Several lines of evidence support our findings that svt2 is a novel mutant. First, svt2 was the only suppressor mutant isolated among over 1000 EMS-mutagenized M0 seeds to show unique phenotypic characteristics. Astonishingly, our genetic analysis revealed that both maternal and paternal alleles were affected in five randomly selected InDel polymporphism loci, the newly discovered InDel polymporphism in the VTC1 promoter, and additional SNPs (Figure 5B–D, Figure 6, Figure S1). These data demonstrate that svt2 has acquired new characteristics, presumably as a result of EMS mutagenesis, and that svt2 is neither Col nor Ler. These data also argue against svt2 being an experimental or PCR artifact.
Second, a number of data provide strong arguments against seed contamination. (1) With high reproducibility, descendents of the original svt2 mutant produce offspring revertants with Col-like features (Table 3, Table 4; Figure 8, Figure 9). (2) One of the Col-like revertants, svt2 Col R1 M3, exhibited heterozygosity at some of the InDels tested (Table 4). (3) One of those Col-like revertants, svt2 K1 Col R M3, produced progeny that reverted back to svt2-like plants (Table 3, Table 4; Figure 8, Figure 9). (4) We were unable to obtain true F1 heterozygotes in all svt2/Col-0 reciprocal crosses (Table 5). (5) Delayed flowering and enlarged morphology phenotypes argue against the fact that svt2 is a result of a Ler-0 wild-type seed landing on the flat during the initial planting of the vtc1-1 M0 mutagenized population. There is the possibility of a Ler seed contamination of the vtc1-1 seed stock used for EMS mutagenesis. Although we have sequenced the vtc1-1 seed stock used for this experiment and confirmed that it is homozygous for the vtc1-1 mutation, one could argue that sequencing the seed stock may not be a sensitive enough method to rule out contamination with a few Ler seed. We performed many other experiments using this very same seed stock and never observed Ler-like plants among the vtc1 population. However, arguments (1) through (4) above speak most compellingly against seed contamination.
Third, the following experimental evidence argues against the possibility that svt2 was generated by cross pollination of vtc1-1 mutant plants with Ler wild-type plants. (1) If svt2 were generated by Ler cross-pollination, the InDel polymorphism markers tested using svt2 genomic DNA should have indicated heterozygosity. This, however, was not the case (Table 4). (2) While svt2 shares phenotypic and genotypic characteristics with Ler and Col, it also has unique features (Figure 3A, Figure S1). (3) svt2 exhibits phenotypic and genotypic instability, causing the appearance of revertants with persistent reproducibility. (4) Ler plants were not grown in our growth chambers at the time of the mutagenesis experiment. Furthermore, svt2 was isolated by placing Aracons over the mutant plant to allow self-fertilization and seed production.
Possible causes of genome instability in svt2
Our results are indicative of genome instability in svt2. Genome instability may be a result of polyploidy24. Polyploids can arise from genome duplication (autopolyploids) or interspecific hybridization (allopolyploids). Our data suggest that svt2 does not contain multiple sets of chromosomes, because VTC1 occurs as a single copy gene in svt2 and vtc1-1 mutants as well as the Col-0 and Ler-0 wild-type controls (Table 2). Furthermore, extra DNA must be replicated with each cell division. Therefore, enlarged cell size is often associated with polyploids25. The chemical mutagenesis of vtc1-1 seed could have resulted in mutations, which may have led to increased ploidy levels in one, two, or all three meristem layers, L1, L2, and L3. However, only mutations in the L2 layer, which gives rise to the reproductive organs, are inherited. Polyploidy in the L2 layer is reflected in pollen size. While svt2 has an overall enlarged morphology (Figure 3A), its pollen size is comparable to that of the other three genotypes (Figure 4). This suggests that svt2 anthers are not polyploid. Finally, allopolyploids often display a greater degree of heterozygosity25, low fertility, and low embryonic viability26–28. This, however, is not the case in svt2. The fact that svt2 is fertile and that its enlarged morphology is heritable from one generation to the next suggests that svt2 is neither a somatic nor a gametic polyploid. Thus, it is therefore unlikely that polyploidy in svt2 contributes to genome instability. This is supported by Ruffio-Chable and co-workers, who reported that between 5% and 21% of F1 hybrids in Brassica oleracea showed aberrant leaf phenotypes, despite normal ploidy levels29.
Instead, we hypothesize that genome instability of svt2 was further aggravated by exposing the already instable genome of vtc1-1 mutants to EMS. It has recently been shown that plants impaired in certain aspects of protection against reactive oxygen species have a higher incidence of spontaneous double-strand breaks30. The AA-deficient vtc1-1 mutant has a three-fold higher spontaneous homologous recombination frequency and has a higher incidence of double-strand breaks (see below). Similar results were reported for the Arabidopsis thaliana flavonoid-deficient mutants tt4 and tt530. One may speculate that through the high level of stress induced by EMS, a yet unknown mechanism of genome restoration was turned on. In fact, genome alterations in soybean and flax in response to environmental stress have been reported previously31,32. In the process of soybean cell culture, massive specific changes in numerous genome-wide loci were observed31. It was suggested that this genetic variation is a consequence of specific recombinational events. Similarly, in flax a single-copy 5.7 kilobase DNA fragment that was not present in the parent line appeared in genotrophs in response to particular growth conditions32.
Possible mechanisms of genome restoration in svt2
The experimental evidence described in this work raises the question as to what mechanism is responsible for the loss or reintroduction of genomic DNA sequences in the original svt2 mutant and its revertant offspring. Several mechanisms may be considered: activity of transposable elements, random mutations, unequal crossing over, gene conversion, double-strand breaks and recombination, and activity of an RNA cache.
Transposons are DNA elements capable of moving around the genome; movement is often associated with chromosome breaks and formation of unstable mutations, which revert frequently but often give rise to new phenotypes. Movement of transposable elements often occurs during meiosis and mitosis and is accelerated by genome damage33. These represent conditions that are present in svt2. However, transposons have a variety of molecular features that do not apply to svt2. Transposons exist as multiple copies in the genome. A blast search of the VTC1 promoter insertion in svt2 did not return any other hits, indicating that the DNA sequence is not present in its entirety anywhere else in the genome. Additionally, transposon termini represent inverted repeats. This, however, is not the case in svt2 (Figure S1). A short, direct repeat of genomic DNA often flanks the transposon, leaving a “footprint”. Our sequencing analysis of the VTC1 promoter region in svt2 did not reveal any footprints, suggesting that transposon activity is not responsible for the insertion or loss of novel sequences in svt2 (Figure S1).
Random mutations caused by EMS mutagenesis could have activated an unknown mechanism in vtc1-1 seeds, giving rise to the phenome and genome instability in svt2. This may explain the novel SNPs we detected in svt2 that are distinct from the vtc1-1 mutant and Col-0 and Ler-0 wild types (Figure S1). The disappearance of the vtc1-1 mutation in svt2 (Figure 5D, Figure S1) may also be explained by the introduction of a random mutation. However, it is possible that exposure of vtc1-1 seeds to EMS could have reversed the original vtc1-1 mutation to the wild-type sequence, as vtc1-1 was initially isolated in an EMS screen15. Interestingly, Conklin and co-workers previously isolated two vtc1 alleles, vtc1-1 and vtc1-2, containing the exact same single cytosine to thymine point mutation at amino acid position 64 relative to the start codon, despite the fact that vtc1-1 and vtc1-2 mutants were isolated independently from different EMS-mutagenized pools15. The authors suggested that a limited number of mutations are tolerable in the VTC1 enzyme GDP-d-mannose pyrophosphorylase without causing embryo lethality. This is supported by the fact that several independently isolated cyt mutant alleles containing different amino acid mutations in VTC1 are embryo lethal34. To date, only the vtc1-115 and hsn1 mutations8 have been isolated and reportedly do not cause embryo lethality. This suggests some form of allelic constraint that has been reported in Arabidopsis previously35,36. Furthermore, in the EMS screen in which the svt2 mutant was isolated, several other vtc1-1 suppressor mutants with restored root development in the presence of ammonium were identified. Sequencing analysis revealed that in all of these mutants the vtc1-1 mutation was restored to the wild-type allele, while the suppressor mutants neither exhibited a svt2-like phenotype nor did they produce revertants in the subsequent generation (Kempinski et al., unpublished data).
Exposure to EMS or γ–radiation has been reported to induce high frequency phenotypic instability in the Arabidopsis disease resistance genes CPR1 and BAL, which map to the RPP5 locus16. Yi and Richards reported destabilization of phenotypes in both the bal and cpr1 mutants in more than 10% of EMS-treated plants in the M1 generation. They also identified exceptions to simple Mendelian inheritance in the M2 generation. Phenotypic instability was also observed in bal × cpr1 F1 hybrids. The authors suggested that the high degree of phenotypic instability in bal and cpr1 mutants is due to the fact that the RPR5 locus can adopt different metastable genetic or epigenetic states, whose stability is highly susceptible to mutagenesis and pairing of different alleles. Yi and Richards later reported that the phenotypic instability of bal mutants is caused mainly by gene duplication and hypermutation of the SNC1 gene17.
As observed in the cpr1 and bal mutants, we hypothesize that EMS treatment has destabilized the genome of svt2 by interrupting one or more mechanisms involved in genomic inheritance. A combination of unequal crossing over, gene conversion, double-strand breaks, DNA recombination, and/or the presence of an RNA cache template may explain the loss and reappearance of DNA sequences in svt2. Genome-wide non-Medelian inheritance of extra-genomic information in Arabidopsis was reported in the hothead (hth) Arabidopsis mutant37. Self-fertilization of homozygous mutant plants resulted in approximately 10% hth revertants, which were hth/HTH heterozygous, suggesting that the HTH gene was altered in the progeny. However, the authors also detected rare homozygous revertants HTH/HTH embryos, which must have inherited one of their two wild-type HTH genes from the maternal parent and could not have been a result of outcrossing. Inheritable genome-wide high-frequency gene homozygosity in early generations in rice has also been reported38. Lolle et al. postulated that these genetic restoration events are the result of a template-directed process that utilizes an ancestral RNA-sequence cache37. This hypothesis is supported by observations reported by Xu and co-workers38. Therefore, our genetic and phenotypic svt2 data, in conjunction with the observed higher occurrence of double-strand breaks and spontaneous homologous recombination frequency in vtc1-1, are in support of the RNA cache theory. Additional studies are needed to provide experimental support for this hypothesis.
Conclusions
We have isolated a novel Arabidopsis mutant that is capable of restoring genetic information that was not present in the chromosomal genome of its parents. We suggest that this ancestral information is present in some cryptic form that is accessible under extreme stress conditions. Genome restoration could be advantageous to plants that encounter environmental changes for which ancestral genes were better adapted. However, the mechanisms responsible for triggering and executing genome restoration remain to be determined. Double strand breaks, DNA recombination, and/or the activity of an RNA cache may be contributing factors. In the future, svt2 may serve as a model to study non-Mendelian inheritance and could provide insight into the evolution and diversification of Arabidopsis ecotypes.
Abbreviations
AA, ascorbic acid; EMS, ethyl methanesulfonate; InDel, Insertion/Deletion; MS, Murashige and Skoog.
Author contributions
CB and CFK conceived the study and designed the experiments. CFK, SVC, CS and CB conducted the experiments and analyzed the data. CB and CFK prepared the first draft of the manuscript. All authors were involved in the revision of the draft manuscript and have agreed to the final content.
Competing interests
No relevant competing interests disclosed.
Grant information
This work was supported by a start-up grant of West Virginia University to CB.
Acknowledgements
We would like to thank Dr. Patricia Conklin for providing vtc1-1 mutant and Col-0 wild-type seed. We also wish to thank Dr. Karen Weiler for allowing us to use her microscope and Dr. Rosana Schafer for providing a plate reader.
Supplementary material
Figure S1. Sequence alignment of the VTC1 gene sequence of the Col-0 TAIR database, the vtc1-1, svt2 mutants, and the Ler-0 GenBank database.
Horizontal arrows denote 5´ respectively 3´ flanking regions of the sequence insertion, which is highlighted in grey, in the VTC1 promoter region (between base pairs 1990 and 2273). Upright arrows indicate sequences shared between svt2 and Ler. Arrows pointing down denote sequences shared between svt2 and Col. Arrowheads point to sequences unique to svt2. Highlighted in yellow are the start and stop codons, respectively. Highlighted in green is the vtc1-1 mutation.
Figure S2. Sequence alignment of the VTC1 promoter InDel polymorphism sequence of the Col-0 TAIR database, the Ler-0 Genbank database, the svt2 K1 Col R M3 revertant (Col-like phenotype) and the svt2 K1 Col R svt2 R M4 revertant (svt2-like phenotype).
Horizontal arrows denote 5´ respectively 3´ flanking regions of the sequence insertion, which is highlighted in grey, in the VTC1 promoter region in Ler-0 and svt2 K1 Col R svt2 R M4, which exhibits an svt2-like phenotype. The svt2 K1 Col R M3 mutant has a Col-like phenotype and share sequence similarities with the Col-0 sequence. R denotes revertant.
Faculty Opinions recommendedReferences
- 1.
Conklin PL, Barth C:
Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens, and the onset of senescence.
Plant Cell Environ.
2004;27(8): 959–970. Publisher Full Text
- 2.
Pavet V, Olmos E, Kiddle G, et al.:
Ascorbic acid deficiency activates cell death and disease resistance responses in Arabidopsis.
Plant Physiol.
2005; 139(3): 1291–1303. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 3.
Kotchoni SO, Larrimore KE, Mukherjee M, et al.:
Alterations in the endogenous ascorbic acid content affect flowering time in Arabidopsis.
Plant Physiol.
2009; 149(2): 803–815. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 4.
Mukherjee M, Larrimore KE, Ahmed NJ, et al.:
Ascorbic Acid Deficiency in Arabidopsis Induces Constitutive Priming That is Dependent on Hydrogen Peroxide, Salicylic Acid, and the NPR1 Gene.
Mol Plant Microbe Interact.
2010; 23(3): 340–351. PubMed Abstract
| Publisher Full Text
- 5.
Arrigoni O, De Tullio MC:
The role of ascorbic acid in cell metabolism: between gene-directed functions and unpredictable chemical reactions.
J Plant Physiol.
2000; 157(6): 481–488. Publisher Full Text
- 6.
Arrigoni O, De Tullio MC:
Ascorbic acid: much more than just an antioxidant.
Biochim Biophys Acta.
2002; 1569(1–3): 1–9. PubMed Abstract
| Publisher Full Text
- 7.
Pastori GM, Kiddle G, Antoniw J, et al.:
Leaf vitamin C contents modulate plant defense transcripts and regulate genes that control development through hormone signaling.
Plant Cell.
2003; 15(4): 939–951. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 8.
Qin C, Qian W, Wang W, et al.:
GDP-mannose pyrophosphorylase is a genetic determinant of ammonium sensitivity in Arabidopsis thaliana.
Proc Natl Acad Sci USA.
2008; 105(47): 18308–18313. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 9.
Barth C, Gouzd ZA, Steele HP, et al.:
A mutation in GDP-mannose pyrophosphorylase causes conditional hypersensitivity to ammonium, resulting in Arabidopsis root growth inhibition, altered ammonium metabolism, and hormone homeostasis.
J Exp Bot.
2010; 61(2): 379–394. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 10.
Li Q, Li BH, Kronzucker HJ, et al.:
Root growth inhibition by NH4+ in Arabidopsis is mediated by the root tip and is linked to NH4+ efflux and GMPase activity.
Plant Cell Environ.
2010; 33(9): 1529–1542. PubMed Abstract
| Publisher Full Text
- 11.
Kempinski CF, Haffar R, Barth C, et al.:
Toward the mechanism of NH4+ sensitivity mediated by Arabidopsis GDP-mannose pyrophosphorylase.
Plant Cell Environ.
2011; 34(5): 847–858. PubMed Abstract
| Publisher Full Text
- 12.
Wheeler GL, Jones MA, Smirnoff N, et al.:
The biosynthetic pathway of vitamin C in higher plants.
Nature.
1998; 393(6683): 365–369. PubMed Abstract
| Publisher Full Text
- 13.
Dowdle J, Ishikawa T, Gatzek S, et al.:
Two genes in Arabidopsis thaliana encoding GDP-l-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability.
Plant J.
2007; 52(4): 673–689. PubMed Abstract
| Publisher Full Text
- 14.
Conklin PL, Williams EH, Last RL, et al.:
Environmental stress sensitivity of an ascorbic acid-deficient Arabidopsis mutant.
Proc Natl Acad Sci U S A.
1996; 93(18): 9970–9974. PubMed Abstract
| Free Full Text
- 15.
Conklin PL, Norris SR, Wheeler GL, et al.:
Genetic evidence for the role of GDP-mannose in plant ascorbic acid (vitamin C) biosynthesis.
Proc Natl Acad Sci U S A.
1999; 96(7): 4198–4203. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 16.
Yi H, Richards EJ:
Phenotypic instability of Arabidopsis alleles affecting a disease Resistance gene cluster.
BMC Plant Biol.
2008; 8: 36. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 17.
Yi H, Richards EJ:
Gene duplication and hypermutation of the pathogen Resistance gene SNC1 in the Arabidopsis bal variant.
Genetics.
2009; 183(4): 1227–1234. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 18.
Weigel D, Glazebrook J:
Arabidopsis: A Laboratory Manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press. 2002, 354. Reference Source
- 19.
Conklin PL, Saracco SA, Norris SR, et al.:
Identification of ascorbic acid-deficient Arabidopsis thaliana mutants.
Genetics.
2000; 154(2): 847–856. PubMed Abstract
| Free Full Text
- 20.
De Preter K, Speleman F, Combaret V, et al.:
Quantification of MYCN, DDX1, and NAG gene copy number in neuroblastoma using a real-time quantitative PCR assay.
Mod Pathol.
2002; 15(2): 159–166. PubMed Abstract
| Publisher Full Text
- 21.
Duarte JM, Wall PK, Edger PP, et al.:
Identification of shared single copy nuclear genes in Arabidopsis, Populus, Vitis and Oryza and their phylogenetic utility across various taxonomic levels.
BMC Evol Biol.
2010; 10: 61. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 22.
Molinier J, Ries G, Zipfel C, et al.:
Transgeneration memory of stress in plants.
Nature.
2006; 442(7106): 1046–1049. PubMed Abstract
| Publisher Full Text
- 23.
Jablonka E, Raz G:
Transgenerational epigenetic inheritance: prevalence, mechanisms, and implications for the study of heredity and evolution.
Q Rev Biol.
2009; 84(2): 131–176. PubMed Abstract
| Publisher Full Text
- 24.
Wang Y, Jha AK, Chen R, et al.:
Polyploidy-associated genomic instability in Arabidopsis thaliana.
Genesis.
2010; 48(4): 254–263. PubMed Abstract
| Publisher Full Text
- 25.
Ranney TG:
Polyploid: From Evolution to New Plant Development.
Combined Proceedings International Plant Propagators’ Society.
2006; 56: 137–142. Reference Source
- 26.
Soltis DE, Soltis PS:
The dynamic nature of polyploid genomes.
Proc Natl Acad Sci U S A.
1995; 92(18): 8089–8091. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 27.
Comai L, Tyagi AP, Winter K, et al.:
Phenotypic instability and rapid gene silencing in newly formed Arabidopsis allotetraploids.
Plant Cell.
2000; 12(9): 1551–1568. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 28.
Schranz ME, Osborn TC:
Novel flowering time variation in the resynthesized polyploid Brassica napus.
J Hered.
2000; 91(3): 242–246. PubMed Abstract
| Publisher Full Text
- 29.
Ruffio-Chable V, Chatelet P, Thomas G, et al.:
Developmentally “aberrant” plants in F1 hybrids of Brassica oleracea.
Acta Hort.
2000; 539: 89–94. Reference Source
- 30.
Filkowski J, Kovalchuk O, Kovalchuk I, et al.:
Genome stability of vtc1, tt4, and tt5 Arabidopsis thaliana mutants impaired in protection against oxidative stress.
Plant J.
2004; 38(1): 60–69. PubMed Abstract
| Publisher Full Text
- 31.
Roth EJ, Frazier BL, Apuya NR, et al.:
Genetic variation in an inbred plant: variation in tissue cultures of soybean [Glycine max (L.) Merrill].
Genetics.
1989; 121(2): 359–368. PubMed Abstract
| Free Full Text
- 32.
Chen Y, Lowenfeld R, Cullis CA, et al.:
An environmentally induced adaptive (?) insertion event in flax.
Int J Biochem Mol Biol.
2009; 1(3): 038–047. Reference Source
- 33.
Lankenau DH, Volff JN:
Transposons and the Dynamic Genome. Berlin, Heidelberg: Springer. 2009, 200. Reference Source
- 34.
Lukowitz W, Nickle TC, Meinke DW, et al.:
Arabidopsis cyt1 mutants are deficient in a mannose-1-phosphate guanylyltransferase and point to a requirement of N-linked glycosylation for cellulose biosynthesis.
Proc Natl Acad Sci U S A.
2001; 98(5): 2262–2267. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 35.
Li J, Last RL:
The Arabidopsis thaliana trp5 mutant has a feedback-resistant anthranilate synthase and elevated soluble tryptophan.
Plant Physiol.
1996; 110(1): 51–59. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 36.
Kreps JA, Ponappa T, Dong W, et al.:
Molecular basis of alpha-methyltryptophan resistance in amt-1 a mutant of Arabidopsis thaliana with altered tryptophan metabolism.
Plant Physiol.
1996; 110(4): 1159–1165. PubMed Abstract
| Publisher Full Text
| Free Full Text
- 37.
Lolle SJ, Victor JL, Young JM, et al.:
Genome-wide non-mendelian inheritance of extra-genomic information in Arabidopsis.
Nature.
2005; 434(7032): 505–509. PubMed Abstract
| Publisher Full Text
- 38.
Xu PZ, Yuan S, Li Y, et al.:
Genome-wide high-frequency non-Mendelian loss of heterozygosity in rice.
Genome.
2007; 50(3): 297–302. PubMed Abstract
| Publisher Full Text



Comments on this article Comments (0)