Keywords
Bacteria, COVID-19, fungus, nitric oxide, nitrate, nitrite, protozoa, virus
This article is included in the Pathogens gateway.
Bacteria, COVID-19, fungus, nitric oxide, nitrate, nitrite, protozoa, virus
In response to the first reviewer, we have added text and references as follows: L-citrulline as a source of NO wherever dietary L-arginine is mentioned, including to table 1; the change to table 1 has meant renumbering the subsequent references to this table. We have added text and references addressing: nitrite/nitrate in animal models of wound healing (Afzali et al 2020); RRx-001 as a NO donor (Oronsky et al 2020); and text and two references relating to beet root juice improving insulin sensitivity and reducing blood glucose (Wootton et al 2014, Beals et al 2017).
In response to a comment from Dr Taylor-Robinsonwe, we have added text on the the importance of malaria and long history of research into the importance of nitric oxide with it having both protective and anti-immunological effects (Taylor-Robinson & Smith 1999, Shikani et al 2012).
See the authors' detailed response to the review by Khosrow Kashfi
Nitric oxide (NO), an inorganic molecule, is generated endogenously by prokaryotes and eukaryotes from L-arginine and L-citrulline by a family of NO synthase enzymes (NOS; Table 1.1, Table 1.2).1-3 In higher animals, it is also generated by reduction of dietary and endogenous nitrate (NO3−) to nitrite (NO2−) and thence NO (Table 1.3). NO is a pleiotropic signalling molecule involved in vascular, neuronal and metabolic regulation and has multiple physiological effects including lowering blood pressure, increasing exercise performance, and reversing metabolic syndrome. Underlying these processes, NO modulates multiple cell types including leucocytes,4 platelets,5 endothelial cells and smooth muscle cells, and neuronal, cardiac and renal function. Three isoforms of NOS exist in eukaryotes: neuronal (nNOS, NOS1), inducible (iNOS, NOS2) and endothelial (eNOS, NOS3). In multicellular organisms, NOS1-3 produces NO that broadly mediates neurotransmission, cyto-toxicity and vascular regulation respectively. Within cells, NO interacts with mitochondrial respiration, activates metabolic regulatory pathways and reduces oxidative stress.
As people age, endothelial-derived vascular NO levels fall and so vascular function declines causing relative endothelial dysfunction, pro-platelet and pro-inflammatory effects, and increased smooth muscle proliferation. Vascular NO levels are even lower in people with established vascular disease, e.g. those with stroke.6,7 Numerous viruses (including adeno, Coxsackie, coronavirus, cytomegalovirus, echovirus, herpes simplex, human T-cell leukaemia virus type-1, human immunodeficiency virus, influenza, measles, mumps, polio) and bacteria (Leptospira spp.) can infect8–10 and damage endothelium and so further reduce vascular NO levels. This also appears to occur in SARS-CoV-2 infection.11
Most physiological effects of NO are modulated by cyclic guanosine monophosphate (cGMP, second messenger),12,13 and terminated when cGMP is metabolised by phosphodiesterase-5 (PDE5). This combined L-arginine/nitrate-NO-cGMP-PDE5 system (or nitric oxide system) comprises one of two key vasculo-protective pathways, the other being the prostaglandin-cyclic adenosine monophosphate-phosphodiesterase-3 pathway (PG-cAMP-PDE3, or prostacyclin system, see below).
The NO system may be enhanced or stimulated exogenously with substrate (L-arginine, L-citrulline, organic nitrate, inorganic nitrite or nitrate), NO gas, and PDE5 inhibitors. These can be inhaled or administered via transdermal, sublingual, oral, intranasal or intravenous routes (see below). Since endogenous NO generated by inducible NOS plays a key role in defence against multiple microbial pathogens (including viruses, bacteria, protozoa and fungi/yeast), this raises the possibility that exogenous NO might have therapeutic potential as a broad-spectrum antimicrobial, and this is the topic of this review.
There are numerous publications in this research area and our intention was not to perform a systematic review of these; rather we present exemplars from the research field. We identified publications relating to the effect of NO on viruses, bacteria, protozoa and fungi/yeasts from searches of our own reference libraries, PubMed and Google, and reference lists given in earlier reviews and commentaries.14–22
The primary searches for relevant studies for inclusion were done through PubMed from inception to 4 May 2021, in English with the following disease terms: “microbe” or “virus” or “bacteria” or “protozoa” or “fungi” or “yeast”. The results of these searches were crossed with the drug terms “nitric oxide” or “nitrite” or “nitrate”, and the identified abstracts were screened by one or more researchers. Earlier studies, including published reviews, were also identified from the files of the senior author. Studies included in vitro, in vivo and clinical trials. Although other drug classes, such as statins and angiotensin-converting enzyme inhibitors, enhance endothelial production of nitric oxide, their main effects are mediated through other pathways, and we have not assessed them here even though they may attenuate microbial disease severity.
NO is a small diatomic hydrophobic colourless gas that diffuses easily and has a short half-life. With an unpaired electron (NO•), it is a free radical and so is chemically reactive and unstable in the presence of oxygen and superoxide producing reactive nitrogen oxide species (RNOS). In reality, the chemistry of NO is more complex and it exists in several redox forms: nitrosonium cation (NO+), NO (NO•) and nitroxyl anion NO−).23
As an inorganic molecule, its central role in biology as a signalling molecule was only discovered in the 1980s,24 the identification of which led to the Nobel Prize for Physiology and Medicine in 1998.25 Ironically, the medicinal use of NO in the form of glyceryl trinitrate (GTN) for angina prophylaxis antedates the modern understanding of the biological synthesis and role of NO by more than a century.26
Numerous preclinical in vitro studies have demonstrated that NO sources (stimulated endogenous NO, inorganic and organic nitrates, L-arginine) and PDE5-inhibitors can reduce replication in all seven types of virus as defined in the Baltimore classification (Table 2);27 this includes Class IV viruses (positive-sense RNA viruses) incorporating several coronaviruses28–33 including SARS-CoV-2.34–36 Most studies showed efficacy although neutral studies were reported for porcine reproductive and respiratory virus (an arterivirus, which is closely related to coronaviruses) and rhinovirus.37,38
| Year | Virus (Baltimore class) | Disease (human unless stated) | NO source | In vitro cell lines | In vivo | Results |
|---|---|---|---|---|---|---|
| dsDNA (I) | ||||||
| 202045 | Cytomegalovirus | In compromised immune systems | DETA/NO | MRC-5, ARPE-19 | Reduced replication | |
| 1993196 | Ectromelia | Mousepox | SNAP | 293 | C57BL/6NCR mice | Reduced replication |
| 1994197 | Epstein-Barr | Infectious mononucleosis, Burkitt lymphoma | Constitutive lymphocyte | Human B-lymphocytes | Reduced reactivation | |
| 1993196 | Herpes simplex-1 | Cold sores | SNAP | 293 | C57BL/6NCR mice | Reduced replication |
| 1993198 | Herpes simplex-1 | Cold sores | SNAP | RAW 264.7 | Reduced replication | |
| 2015132 | Human papilloma virus | Anogenital warts | Acidified NaNO2 | Human | Cure in 31% (active) vs 14% (control) | |
| 1999128 | Molluscipoxvirus | Molluscum contagiosum | Acidified NaNO2 | Human | Cure in 75% (active) vs 21% (control) | |
| 1993196 | Vaccinia | Human “pox” | SNAP | 293 | C57BL/6NCR mice | Reduced replication |
| 1995199 | Vaccinia | Human “pox” | L-arg | RAW 264.7 | Reduced replication | |
| 1995200 | Vaccinia | Human “pox” | iNOS | BSC-40, HeLa G | Reduced replication | |
| ssDNA (II) | ||||||
| 2017201 | Porcine circovirus-2 | Swine multisystemic wasting syndrome | GSNO | PK-15 | BALB/c mice | Reduced replication & infection |
| 2009202 | Porcine parvovirus | Swine embryonic/foetal death | SNAP, L-arg | PK-15 | Reduced replication | |
| dsRNA (III) | ||||||
| 1996203 | Avian (ortho-) reovirus | Avian arthritis, tenosynovitis | LPS-stimulated macrophages | HD11 | Reduced replication | |
| (+)ssRNA (IV) | ||||||
| 1997204 | Coxsackievirus (B3) | Pleurodynia, myocarditis, pericarditis, and hepatitis | iNOS transfection, SNAP | HeLa | Reduced replication, RNA and protein synthesis | |
| 2006205 | Dengue virus-2 | Viral haemorrhagic fever | SNAP | LLC-MK2 monkey kidney | Reduced RNA and protein synthesis | |
| 1999206 | Human immunodeficiency virus | Acquired immune deficiency syndrome | SNAP | Human monocytes. U1 cells | Reduced replication in monocytes. Increased replication in U1 cells | |
| 1997207 | Japanese encephalitis virus | Encephalitis | SNAP, IFN-γ activated macrophages | Murine RAW 264.7 and N18 cells | Reduced replication | |
| 2005208 | Mengovirus | Acute fever | Dipyridamole | HeLa or L cells | Reduced replication | |
| 199728 | Murine coronavirus | Murine hepatitis | SNAP | OBL21a | Reduced replication | |
| 1998 | Poliovirus | Poliomyelitis | GTN | HeLa, U937 | Reduced replication | |
| 201037 | Porcine reproductive & respiratory virus | Swine reproductive failure, respiratory tract infection | SNAP, NAP | Marc-145 cells | NAP (but not SNAP) inhibited replication | |
| 201029 | Porcine respiratory coronavirus | Swine reproductive failure, respiratory tract infection | SNAP | STC | Reduced replication | |
| 202032 | OC43 coronavirus | Cold | NO gas | HCT-8 cells | Reduced replication | |
| 199938 | Rhinovirus type 23 | Common cold | SNAP, SNP, PAPA-NONOate | BEAS-2B cells, MRC-5 cells | No effect on replication | |
| 1996209 | Sindbis | Sindbis fever. Murine encephalomyelitis | SNAP, SNP, NOS | N18 | BALB/cJ, BALB/cByJ, scid/CB17 mice | Increased cell viability; less death |
| 200430 | SARS-CoV-1 | SARS | SNAP, SNP | Vero E6 | SNAP (not SNP) reduced replication | |
| 200433 | SARS-CoV-1 | SARS | NO gas | Human (n=14) | Improved arterial oxygenation, less lung radiological infiltrates | |
| 200531 | SARS-CoV-1 | SARS | SNAP, iNOS | Vero E6 | Reduced replication | |
| 202034 | SARS-CoV-2 | COVID-19 | SNAP | Vero E6 | Reduced replication, recombinant protease activity | |
| 202035 | SARS-CoV-2 | COVID-19 | Dipyridamole | Vero E6 | Human (n=31) | Reduced replication. Clinical improvement, increased Lϕ count |
| (-)ssRNA (V) | ||||||
| 1982210 | Influenza A 42/72, 1/79, A/fowl plague | ‘Flu | Dipyridamole | White mice | Reduced replication. Infection prevention | |
| 1999211 | Influenza A/B | ‘Flu | SNAP, SNP, SIN-1 | Mabin Darby | Reduced replication | |
| 2013212 | Influenza H1N1, H3N2, B HongKong | ‘Flu | NO gas | MDCK | Reduced infectivity. Inhibition of neuraminidase | |
| 2000213 | Lymphocytic choriomeningitis virus | Meningoencephalitis | IFN-γ | HBV transgenic/iNOS knockout mice | NO mediates antiviral activity of IFN-γ | |
| 2006214 | (Ortho)hantavirus | Haemorrhagic fever/pulmonary syndrome | SNAP, SIN-1 | Vero E6 | C57BL/6 (iNOS-/-, +/+) mice | Reduced replication |
| 2006215 | Parainfluenza virus | ‘Cold’ | DetaNONOate. SNAP. iNOS overexpression. | Cystic fibrosis epithelial cells | Reduced replication | |
| 2001216 | Rabies virus | Rabies | SNP + ascorbate | Neuroblastoma cells | Reduced replication | |
| 1995217 | Vesicular stomatitis | ‘Flu. Bovine oral ulcers | SNAP | NB41A3 | Reduced replication | |
| ssRNA-RT (VI) | ||||||
| 1995218 | Friend leukaemia + spleen focus-forming | Murine leukaemia | SIN-1, SNP, SNAP | Dunni | Reduced replication (but not with NaNO2) | |
| dsDNA-RT (VII) | ||||||
| 2000213 | Hepatitis B | Viral hepatitis | IFN-γ | HBV transgenic/iNOS knockout mice | NO mediates antiviral activity of IFN-γ |
Multiple studies have assessed the effect of NO on bacteria and inhibitory effects have been seen across a wide range of gram negative, gram positive and acid-fast bacteria (Table 3). NO sources included L-arginine, NO, nitrite, organic nitrates, and endogenously-generated NO. Multiple mechanisms for efficacy have been reported, as discussed below.
| Year | Bacteria | Disease (human unless stated) | NO source | In vitro | In vivo | Results |
|---|---|---|---|---|---|---|
| Gracilicutes (gram negative) | ||||||
| 2005219 | Acinetobacter baumanii | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 1993220 | Brucella abortus | Brucellosis | Activated macrophages (IFN-γ) | BALB-c murine J774A.1 | Reduced cfu | |
| 2003194 | Burkholderia pseudomallei | Melioidosis | Activated macrophages (IFN-β) | RAW 264.7 murine macrophages | Reduced intracellular bacteria | |
| 1993221 | Chlamydia trachomatis | Trachoma, pelvic inflammatory disease | Activated McCoy cells (IFN-γ) | Murine fibroblasts | Reduced infectivity | |
| 1992222 | Ehrlichia | Ehrilichiosis | Activated macrophages (L-arginine/IFN-γ). SNP | Murine macrophages | No viable bacteria. Dependent on iron (not cGMP) | |
| 2005219 | Enterobacter aerogenes | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 2005219 | Escherichia coli | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 1992223 | Francisella tularensis | Tularaemia | Activated macrophages (L-arginine/IFN-γ) | Murine macrophages | Suppressed growth | |
| 1998124 | Helicobacter pylori | Gastritis, gastric/duodenal ulcers | Acidified (pH 2) potassium nitrite | Bacterial cfu | No viable bacteria at KNO2 >500 μmol/L | |
| 2005219 | Klebsiella pneumoniae | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 1992224 | Legionella | Legionnaires/Pontiac fever | Activated macrophages (IFN-γ) | RAW 264.7/HL-60 | Few viable bacteria | |
| 2005219 | Pseudomonas aeruginosa | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 1993225 | Rickettsia | Spotted fever, typhus | Activated macrophages (IFN-γ/TNF-α) | Murine fibroblasts | Reduced infection | |
| 199540 | Salmonella enterica (Typhimurium) | Typhoid fever | SIN-1, GSNO, (diethylenetriamine-NO) | Suspension | SIN-1: oxygen-dependent cytostasis. GSNO: oxygen-independent cytostasis | |
| 2005219 | Serratia marcescens | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 2005219 | Stenotrophomonas maltophilia | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr | |
| 1992 de Giusti226 | Yersinia pestis | Plague (bubonic, pneumonic, septicaemic) | NaNO2, NaNO3. KNO3 | Pork meat | Reduced growth | |
| Firmicutes (gram positive) | ||||||
| 1981227 | Bacillus cereus | Gastroenteritis | Nitrosothiols (RSN=O) | Suspension of spores | Inhibition of spore germination | |
| 197646 | Clostridium perfringens | Gastroenteritis | Sodium nitrite | Reduced cfu, GAP-DH and aldolase activity, and free sulfhydryl groups | ||
| 1994228 | Listeria | Listeriosis | Sublethal inoculum | C57BL/6 mice | L-NMMA inhibition of NO worsened outcome | |
| 2005219 | Staphylococcus aureus | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr. | |
| 2012229 | Staphylococcus aureus | Wound infection | Probiotic NO gas patch | Ischaemic/infected (S. aureus) full thickness wounds in rabbits | Improved closure. | |
| 2013153 | Staphylococcus aureus | Experimental biofilms | Glyceryl trinitrate | Biofilm | Antimicrobial synergisation with citrate and ethanol | |
| 2018154 | Staphylococcus aureus | Experimental biofilms | Isosorbide mononitrate | Biofilm | Increased dispersal (conversion of sessile to planktonic cells) | |
| 2005219 | Streptococci (group B) | ICU organisms | NO gas (200 ppm) | Bacterial cfu | No viable bacteria by 4.8 (±1.3) hr. | |
| Acid fast | ||||||
| 1991230 | Mycobacterium avium | Atypical respiratory TB | Activated macrophages (TNF) | Human macrophages | Reduced growth | |
| 1991231 | Mycobacterium leprae | Leprosy | Activated macrophages (IFN-γ) | Murine macrophages | Reduced M. leprae metabolism | |
| 200350 | Mycobacterium tuberculosis | Respiratory TB | Adjuvant L-arginine | Smear positive TB | Improved outcome (weight, less cough) | |
| 2004232 | Mycobacterium ulcerans | Buruli skin ulcer | Acidified nitrite (40 mM) | In suspension | Bacteriocidal |
NO sources have been tested on both intracellular and extracellular protozoa (Table 4) with sources involving activated macrophages, sodium nitrite, glyceryl trinitrate, sodium nitroprusside (SNP) and S-nitroso-L-acetylpenicillamine (SNAP).
| Year | Protozoa | Disease (human unless stated) | NO source | In vitro | In vivo | Results |
|---|---|---|---|---|---|---|
| Extracellular | ||||||
| 1992233 | Entamoeba histolytica | Amoebiasis | Activated macrophages (IFN-γ/LPS) | Murine macrophages | Reduced infection | |
| 1992234 | Naegleria fowleri | Meningitis | BCG-activated macrophages | Female C57BL/6 mice | Destruction of amoebae | |
| 1994235 | Opisthorchis | Opisthorchiasis, cholangiocarcinoma | ||||
| 2013236 | Plasmodium berghei | Murine cerebral malaria | TD GTN | Prevention, and adjunctive treatment | Reduced infection, and improved outcome | |
| 1989237 | Schistosoma mansoni | Schistosomiasis, intestinal | Activated macro-phages (TNF) | Female C57BL/6 mice | Larval cytotoxicity | |
| 2017238 | Schistosoma japonicum | Schistosomiasis, intestinal | Endogenous from iNOS | Sprague-Dawley rats | Reduced granuloma formation | |
| Intracellular | ||||||
| 2000129 | Leishmania major/tropica | Leishmaniasis | Acidified sodium nitrite | BALB/c mice macrophages | Human cutaneous L. tropica | Reduced amastigotes and promastigotes. 28% patients improved, 12% cured. |
| 2000193 | Leishmania major | Leishmaniasis | Activated macrophages (IFN-α/β) | CD1/C57BL/6 mice macrophages | Reduced intracellular parasites | |
| 2016239 | Leishmania spp. | Leishmaniasis | SNP | BALB/c mice macrophages | Reduced amastigotes and promastigotes | |
| 1990240 | Toxoplasma gondii | Toxoplasmosis | Activated macrophages (IFN-γ/LPS) | Murine macrophages | Reduced growth | |
| 1996241 | Toxoplasma gondii (ME49) | Toxoplasmosis | Spleen cells | C57Bl/6 mice | Increased ocular inflammation with aminoguanidine | |
| 1992175 | Trypanosoma brucei gambiencei/brucei | African trypanosomiasis (sleeping sickness) | Activated macrophages (BCG-infected mice or IFN-γ/LPS) | Murine macrophages | BCG-infected mice | No proliferation (cytostasis). Reduced parasitaemia/prolonged survival |
| 1994176 | Trypanosoma brucei | African trypanosomiasis (sleeping sickness) | SNAP. Activated macrophages (IFN-γ) | Murine peritoneal macrophages | C3H.He mice | In vitro: NO inhibits proliferation. In vivo: NO reduced protozoa T-cell proliferative responses |
The effects of NO on several fungi and yeasts have been studied (Table 5). NO was donated exogenously through stimulating macrophages or as acidified nitrite. In vitro experiments demonstrated reduced replication whilst in vivo experiments in mice showed reduced infection.
| Year | Fungi/Yeasts | Disease (human unless stated) | NO source | In vitro | In vivo | Results |
|---|---|---|---|---|---|---|
| Fungi | ||||||
| 1999242 | Aspergillus fumigatus | Aspergillosis | Activated macro-phages (IFN-γ) | Rat alveolar macrophages | Reduced infection | |
| 1998127 | Epidermophyton floccosum | Tinea pedis | Acidified NaNO2 | Human (n=35) | Cure in 81% (active) vs 31% (control) | |
| 1994243 | Histoplasma capsulatum | Histoplasmosis (‘flu-like) | IFN-γ/LPS activated macrophages | C57BL/6 mice | Reduced infection | |
| 1999244 | Pneumocystis carinii | Pneumonia | IFN-γ activated macrophages via L-arginine | Sprague Dawley rats | Killed P. carinii | |
| 1998127 | Trichophyton rubrum, T. interdigitale | Tinea pedis | Acidified NaNO2 | Human (n=35) | Cure in 81% (active) vs 31% (control) | |
| Yeasts | ||||||
| 1993245 | Candida albicans | Candidiasis (oropharyngeal, vulvovaginal), candidaemia | Murine macrophages | C. albicans infection | Mice | Reduced infection |
| 1991246 | Cryptococcus neoformans | Cryptococcosis (pneumonia, meningitis, encephalitis) | Acidified NaNO2 | C. neoformans culture | Reduced replication | |
| 1999242 | Cryptococcus neoformans | Cryptococcosis | Activated macro-phages (IFN-γ) | Rat alveolar macrophages | Reduced infection | |
| 2018247 | Dermantophytes | Onychomycosis, tinea pedis | NVN1000 | Macrodilution broth test | 78-99% kill |
Whilst endogenous NO derived from eNOS and nNOS is physiologically active via its second messenger (cGMP), the antimicrobial effects of NO relate to its toxic effects when present at higher concentrations. Although it is technically challenging to measure free NO concentrations, studies suggest that NO concentrations derived from iNOS are 10–100× higher than those resulting from eNOS/nNOS (Table 6). NO concentrations resulting from exogenous administration lie between those from eNOS/nNOS and iNOS but approximate more closely to those from iNOS than eNOS. Importantly, much antimicrobial NO activity is likely to reflect the effects of derivative molecules rather than NO itself:
• Nitric oxide (NO•). In general, bacteria deficient in low molecular weight thiols such as glutathione (e.g. Staphylococci spp.) are sensitive to attack by NO whereas those with high thiol concentrations are resistant to NO.
• Peroxynitrite (OONO-, Table 1.4). The reaction between NO and superoxide means that NO synergises with the respiratory burst, another antimicrobial system present in phagocytic cells. Experimentally, this synergism can be inhibited with the addition of superoxide dismutase which converts superoxide into molecular oxygen and hydrogen peroxide (Table 1.5).
• Peroxynitrous acid (ONOOH, Table 1.6), e.g. toxic to Escherichia coli.39
• Nitrogen dioxide (NO2, Table 1.7), e.g. toxic to E. coli.39
• Dinitrogen trioxide (N2O3, Table 1.8).
• Dinitrogen tetroxide (N2O4, Table 1.9).
• S-nitrosothiols (RSNO, e.g. S-nitrosoglutathione), e.g. toxic to E. coli and Salmonella enterica serovar typhimurium.40 RSNO reacts with protein sulphydryl groups changing their function. Thiol concentrations do not appear to determine sensitivity to peroxynitrite and S-nitrosothiols.
• Dinitrosyl-iron ((2 RS)-Fe-(2 NO)). The reaction of NO with iron or iron–sulphur molecules can: inactivate enzymes such as aconitase (which converts citrate to isocitrate in the citric acid cycle), ribonucleotide reductase and ubiquinone reductase; increase free ferrous (Fe2+) which causes oxidative damage; and deplete iron stores.
• RNOS (especially auto-oxidised products of NO).
| NO | eNOS/nNOS | iNOS | Exogenous NO |
|---|---|---|---|
| Source | Endothelium, neurones | Intracellular, e.g. macrophages | Exogenous |
| Role | Cell signalling | Microbial killing | Vasodilation, antiplatelet |
| Synthesis | Constitutive, intermittent (“dripping tap”19), calcium-dependent, feedback controlled | Inducible, continuous (“fire hose”19), calcium-independent/cytokine-microbial dependent. Part of innate immunity | |
| Concentration | 0.1-5 nM248 | >10 μM248 | SNP, 52 nM249 |
| Targets | sGC (CcOX) | Aconitase, NADH dehydrogenase, succinate dehydrogenase, metalloenzymes, ribonucleotide reductase, DNA | sGC |
| Effects | Reversible | Irreversible. Nitrosation, nitration, oxidation. | Reversible |
Since these molecules differ in their stability, reactivity, location and cellular diffusivity, the overall effect of NO will depend on the molecular species involved and its location.
The targets for NO and associated reactive nitrogen species are multitudinous:
• DNA, through deamination of adenine, cytosine and guanine;41 cross-linking; breakage of strands; inhibition of DNA repair enzymes such as DNA alkyl transferases (and so preventing transfer of the guanine alkyl group to protein); and disruption of DNA replication by inhibition of ribonucleotide reductase;42 as in S. enterica and vaccinia virus.
• RNA, through disruption of RNA replication by inhibition of viral ribonucleotide reductase.
• Inhibition of mitochondrial function, specifically through inactivation of iron-sulphur complexes within respiratory chain enzymes.43
• Protein modification at cysteine, methionine, phenylalanine, tryptophan and tyrosine residues, e.g. by RNOS. Such protein effects will reduce enzyme activity, as seen for DNA, proteases44 and mitochondrial function, as in Coxsackievirus.44
• Limit late protein synthesis, e.g. through posttranslational modification of viral proteases. (Early protein translation/synthesis is not typically affected.)
• Induction of lipid peroxidation.
• Limit glutaminolysis by shuttling glutamine to glutathione synthesis, as in cytomegalovirus.45
• Interaction with sulfhydryl-containing constituents of the bacterial cell.46
• Disrupt zinc homeostasis, as in S. enterica.47
• Limit virion assembly/particle formation.
• Reduce bacterial adhesion to NO-releasing surfaces.48
Nitric oxide may also play an augmenting role as an antimicrobial agent. Examples include the adjuvant roles of NO when given with type I interferons in the treatment of DNA viruses49 and L-arginine when given with conventional chemotherapy in smear-positive TB.50
In addition, NO’s vasculo-active effects are likely to be beneficial in preventing infection and its severity, with NO:
• Reversing endothelial dysfunction and so potentially reducing endotheliitis,11 as occurs in COVID-19.9,10
• Reducing leucocyte function (e.g. adhesion, chemotaxis, phagocytosis);4 COVID-19 is associated with increased phagocyte counts.10,51
• Reducing platelet activation and platelet–leucocyte conjugation and so reducing micro- and macro-thrombosis, as seen in COVID-19.10,52
• Improving organ blood flow and perfusion through smooth muscle relaxation and vasodilation and so likely reversing infection-related vasoconstriction as seen in COVID-19,10 including in the pulmonary circulation.53
These actions of NO are all mediated via the second messenger cGMP.
NO is produced by some bacteria, archaea and yeasts via several pathways including denitrification of nitrate to nitrite and then to NO54 and oxidation of L-arginine to NO and L-citrulline as catalysed by a bacterial nitric oxide synthase (bNOS), a process that can be inhibited by NOS inhibitors.55 Whilst eukaryotic NOS contains both catalytic and reductase domains, prokaryotic bNOS lacks the latter relying instead on other cellular reductases to generate NO; the one exception to this is the bNOS present in Sorangium cellulosum which does include a reductase module.
In contrast to the signalling role of NO in mammals, NO synthesis in bacteria has multiple functions which vary between antimicrobial species:56–59
• Protection against oxidative stress with NO limiting thiol reduction and so the formation of hydroxyl radicals (Bacillus anthracis/subtilis, Staphylococcus aureus56) (Table 1.10).
• Protection against oxidative stress with NO activating catalase (B. subtilis). Such defence will limit damage from phagocytic respiratory bursts.56
• Protection against oxidative stress by reducing endogenous NO synthesis and increasing the expression of NO dioxygenase to detoxify NO (Candida albicans).58
• Biosynthesis of toxins, e.g. thaxtomins (a phytotoxin) interfere with potato plant wall synthesis (Streptomyces turgidiscabies).56
• Activation of aerobic and nitrate respiration to optimise growth (S. aureus).59,60
• Protection against antimicrobial agents including amoxycillin, cefuroxime, gentamicin and novobiocin (B. anthracis/cereus/thuringiensis/weihenstephanensis, S. aureus),57 and azoles (Candida albicans).58
The production by some microbes of endogenous NO to protect against oxidative stress is ironic since hosts are using NO to try to destroy the microbe!
Microbial resistance to antibiotics is an increasingly common problem and has left some bacteria with few treatment options, e.g. drug-resistant Neisseria gonorrhoeae. Hence, it is vital to consider whether resistance to NO is innate in some microbes or can be acquired. As already highlighted, some microbes have an intrinsic ability to produce their own NO and so attenuate the effects of oxidative stress (e.g. B. anthracis/subtilis, C. albicans, S. aureus56,58), activate aerobic respiration (S. aureus59) or protect against antimicrobial agents (B. anthracis/cereus/thuringiensis/weihenstephanensis, C. albicans, S. aureus57,58).
Microbes may also have mechanisms for deactivating NO. One mechanism is via a NO reductase which reduces NO to nitrous oxide and then nitrogen, e.g. as occurs in fungi61 (Table 1.11). Bacteria have different NO reductases but similarly produce nitrous oxide,61 as seen in Pseudomonas aeruginosa.54 Loss-of-function mutations in NO reductase may be lethal, possibly because intracellular NO concentrations rise to toxic levels.
A second mechanism for detoxifying NO is via NO dioxygenase oxidation to nitrate (Table 1.12). The pre-eminent NO dioxygenase is flavohaemoglobin,62 as present in bacteria (e.g. Salmonella enterica, S. aureus, Vibrio cholerae, Yersinia pestis,22,63,64) and yeasts. A related haemoglobin, truncated haemoglobin, detoxifies NO in mycobacteria. Of note, Mycobacterium leprae has undergone reductive genome evolution losing more than 2,000 genes, including some that protect against RNOS; as a result, M. leprae has fewer defences against NO than Mycobacterium tuberculosis.65 Bacterial lactate dehydrogenase also detoxifies NO, as seen in S. aureus.22 Importantly, these detoxifying enzymes only cope with low levels of NO and are not protective against high NO levels.
As a result, microbes show differing sensitivities to NO, as seen for common airways pathogens where sensitivity was ranked (sensitivity most to least): P. aeruginosa ~ C. albicans > S. aureus > Klebsiella pneumoniae ~ Staphylococcus epidermis.66
However, there is little evidence that bacteria can acquire de novo resistance to NO, as confirmed in experiments on strains of E. coli, P. aeruginosa, S. aureus and Staphylococcus epidermidis.67 This property is unsurprising since NO has multiple mechanisms for antimicrobial activity and these are likely to be invoked orders of magnitude faster than any microbe can process metabolically, especially if protein synthesis is required. Equally, the main mechanisms for antibiotic resistance (drug inactivation, altered binding sites or metabolism and reduced drug permeability) are unlikely to be relevant to many NO sources. Whether viruses, protozoa and fungi can develop resistance to NO remains unclear.
In the presence of NOS, administration of L-arginine may enhance NO synthesis (Table 7) although intracellular L-arginine levels are not normally rate limiting and so administration may not have physiological effects.68 Although oral preparations of L-arginine are commercially available, consumption of high doses is associated with profuse diarrhoea (P Bath, personal observation). However, L-citrulline may be a more efficient method for delivering L-arginine since it has more efficient intestinal absorption, lower first pass metabolism and higher renal reabsorption, does not induce arginase and is safe and tolerable at high doses.4
| Intervention | Example | Administration | Licensed for use in (BNF) | Antimicrobial effects: target (disease) |
|---|---|---|---|---|
| L-arginine | Dietary: meat | Oral | N/A | None reported |
| Powder | N/A | Mycobacteria tuberculosis (pulmonary tuberculosis)50 | ||
| Liquid | Intravenous | N/A | None reported | |
| L-citrulline | Dietary | Oral | N/A | None reported |
| Inorganic nitrite | Acidified sodium nitrite (NaNO3) cream | Topical | Burkholderia cepacia, dermatophytes (tinea pedis),127 pox virus (molluscum contagiosum),128 Leishmaniasis,129 Mycobacterium ulcerans (Buruli ulcer),130 human papilloma virus (anogenital warts),132 Propionibacterium acnes, P. aeruginosa, S. aureus (including MRSA131).110 | |
| Oral | N/A | Food preservation:151 Clostridium botulinum | ||
| Intravenous | Cyanide poisoning (given with sodium thiosulfate)151 | None reported | ||
| Inorganic nitrate | Silver nitrate | Topical | Common wart (human papilloma virus) | Human papilloma virus (Common wart).133 |
| Dietary: beetroot, celery, rocket, spinach | Oral | N/A | Lactobacillus, Streptococcus (tooth decay), C. albicans (oral infection), Campylobacter, E. coli 0157, Salmonella enterica (Typhimurium), Shigella sonnei, Yersinia enterocolitica (gastroenteritis)119 | |
| Organic nitrate | Glyceryl trinitrate (GTN) | Topical patch | Prophylaxis of angina and phlebitis | Malaria (murine cerebral malaria).236 Non-specific infections (presenting as stroke mimics).156 |
| Ointment | Treatment of anal fissure | None reported | ||
| Sublingual | Prophylaxis-treatment of angina | None reported | ||
| Intravenous | Hypertension/myocardial ischaemia after cardiac surgery. Congestive heart failure. Unstable angina | E.coli, P. Aeruginosa, S. aureus in solution.250 S. aureus, MRSE, P. aeruginosa, C. albicans in biofilms153 | ||
| Isosorbide dinitrate (ISDN) | Oral | Prophylaxis/treatment of angina. Left ventricular failure | None reported | |
| Sublingual | Prophylaxis/treatment of angina | None reported | ||
| Intravenous | Prophylaxis/treatment of angina. Left ventricular failure | None reported | ||
| Isosorbide mononitrate (ISMN) | Oral | Prophylaxis of angina. Adjunct in congestive heart failure. | S. aureus in biofilms154 | |
| Spontaneous nitric oxide donors | Sodium nitroprusside (SNP) | Intravenous | Hypertensive emergencies. Controlled hypotension. Acute/chronic heart failure | E.coli, P. Aeruginosa, S. aureus in solution.250 Bacillus licheniformis, Candida albicans, Escherichia coli BW20767, Fusobacterium nucleatum, Serratia marcescens MG1, S. epidermidis, Vibrio cholerae 92A1552.251 Leishmania spp.239 |
| Nitric oxide | Nitric oxide (NO) gas | Gas | (Neonatal pulmonary hypertension) | E. coli, P. aeruginosa, S. aureus.252 SARS-CoV-2 (ongoing trials: NCT04290871, NCT04305457, NCT04312243). |
| Probiotic patch | N/A | E. coli, S. aureus, MRSA, P. aeruginosa, T. mentagrophytes, T. rubrum.229,253 | ||
| NO releasing solution | N/A (in development) | Propionibacterium acnes, T. mentagrophytes, T. rubrum. SARS-CoV-2 (ongoing trial: NCT04337918) | ||
| PDE5 inhibitor | Dipyridamole | Oral | Post-stroke prophylaxis. | Picornaviridae, Togaviridae, Orthomyxoviridae, Paramyxoviridae, Herpetoviridae and Poxviridae.254 Mengovirus.208 SARS-CoV-2 (COVID-19 clinical trial).35 |
| Sildenafil | Oral | Erectile dysfunction, pulmonary arterial hypertension | Adenovirus, Chikungunya, Cytomegalovirus, Dengue, Enterovirus 71, Influenza virus, Measles, Mumps, Rabies, Respiratory syncytial virus, Rubella, West Nile, Yellow Fever; Methicillin-resistant Staphylococcus epidermidis.255,256 SARS-CoV-2 (ongoing trial: NCT04304313). |
Gaseous NO may be inhaled with the aim of improving pulmonary haemodynamics and killing microbes. Multiple trials are underway for COVID-19 prevention and treatment (Table 9). NO may also be created in real time by combining sodium nitrite and citric acid and administering this either as a nasal spray (for local therapy) or via nebuliser (for combined nasal and bronchial therapy).
Organic nitrates such as GTN, isosorbide dinitrate (ISDN) and isosorbide mononitrate (ISMN) are widely used in vascular medicine for the prevention and treatment of angina, treatment of chest pain in unstable angina and myocardial infarction, treatment of severe heart failure, and blood pressure lowering after cardiac surgery and in acute stroke (Table 7). There is increasing concern that chronic use of organic nitrates may cause major adverse cardiac events and death,69 reduce daily activity,70 and not improve quality of life or exercise capacity.70 Potential explanations include the development of tolerance, and induction of endothelial dysfunction and cell damage through oxidative stress, e.g. production of free radicals/peroxynitrite.71,72
Importantly, organic nitrates only release NO in cells and tissues expressing mitochondrial aldehyde dehydrogenase-2.73 For example, SNP and SIN-1 inhibit monocyte chemotaxis whilst organic nitrates (ISDN, GTN and molsidomine) do not;74 this contrasts with smooth muscle cells which vasodilate with all five agents. Since aldehyde dehydrogenase-2 suffers from use-inactivation, nitrate tolerance (tachyphylaxis) and endothelial dysfunction develops72 and bioconversion only restarts following a nitrate-free period. Several in vitro studies have demonstrated the potential antimicrobial effects of organic nitrates (Table 7). Other non-organic nitrates include pentaerythritol tetranitrate and erythrityl tetranitrate.
Therapeutic use of inorganic nitrite is limited with intravenous administration used in cyanide poisoning (British National Formulary). Topical acidified sodium nitrite has been shown to reduce cutaneous infections secondary to a variety of viruses and bacteria although its general use is probably limited by skin irritation and erythema (Table 7). A recent study found that acidified nitrite improved wound healing in rats with diabetes.75
NO may also be produced from dietary inorganic nitrate, as is present in high concentrations in green leafy and some root vegetables, e.g. spinach, lettuce, rocket, beetroot, celery, fennel, radish and Chinese cabbage.76 Nitrate is absorbed from the proximal gastrointestinal tract, excreted by salivary glands, reduced to nitrite by oral bacteria and then absorbed in the gastrointestinal tract. A number of bacterial species situated on the dorsal surface of the tongue perform this conversion via nitrate reductases.77 (In the absence of oxygen, nitrate and nitrite are commonly used by bacteria as terminal electron acceptors for respiration.78) Through this symbiotic relationship, the mammalian host provides the nutrients and the environment in return for nitrite production by bacteria.79
Absorbed and circulating nitrite is then further reduced to NO, a process that is enhanced in hypoxic or acidic conditions and by multiple mechanisms including deoxyhaemoglobin, deoxymyoglobin, xanthine oxidoreductase and endothelial nitric oxide synthase.80,81 As such, most effects of dietary nitrate will be vascular and perivascular. The beneficial vascular protective effects of vegetable consumption are very clear epidemiologically, as present in the classical Japanese diet,82 the Dietary Approaches to Stop Hypertension (DASH) diet,83 and the Mediterranean diet.84,85 Further, vegetable-derived nitrate may reduce the risk of gastrointestinal cancer.76,86,87 The benefit on cancer is at variance with oral consumption of nitrite. Although nitrite is not carcinogenic per se, the processing and cooking of nitrite-cured meat can form carcinogens such as N-nitroso compounds and heterocyclic aromatic amines. In contrast, carcinogens are not formed when eating raw vegetable-derived nitrate. A recent meta-analysis showed an increased risk gastric cancer with oral nitrite but reduced risk with oral nitrate.88 Dietary nitrate is known to modify the oral and gastric biome (Table 7).
High dietary intake of nitrate is associated with many mechanisms that may have beneficial vascular, and potentially, antimicrobial effects. Experimentally, beetroot juice is often used as a potent source of dietary nitrate since dosing can be controlled and a nitrate-free placebo version is available for use in randomised controlled trials. Studies have shown that beetroot juice increases plasma nitrate and nitrite concentrations,89,90 that most vascular effects are mediated via the second messenger cGMP,90 tolerance does not develop (unlike with organic nitrates)76 and inorganic nitrate does not lead to free radical formation. In clinical studies, beetroot juice has been given over weeks and months89–94 and has been shown to have multiple effects with improved exercise performance (hence use by elite athletes)95; improved cognitive performance in older people95; vasodilation with reduced blood pressure92,93,95–98; antiplatelet and anti-leucocyte effects and reduced platelet-leucocyte conjugation89,92; improved endothelial function; reduced left ventricular volume;94 improved metabolic profile; and improved oral health.92 Beyond anti-inflammatory effects on blood cells, nitrite or nitrate reduce soluble pro-inflammatory factors including C-reactive protein, chemokine (C-X-C motif) ligand-1/2, endothelin-1, interleukins-1β/6/10/12p70, interferon-γ, monocyte chemoattractant protein and tissue necrosis factor-α.99 Dietary nitrate has profound metabolic effects and appears to have the potential for reversing the metabolic syndrome and have anti-diabetic effects through improving insulin sensitivity and lowering blood glucose levels.100-102 Overall, the pharmacological effects of beetroot juice have been demonstrated in younger and older people, and in people with cardiovascular disease, e.g. diabetes, obesity, hypertension, hypercholesterolaemia, heart failure and stroke.103 Importantly, inorganic nitrate (given as beetroot juice) may be taken by pregnant women.104 Experimentally, watermelon juice and chard gel may be used as an alternative source of dietary nitrate.105,106
PDE5-inhibitors, such as dipyridamole and sildenafil, enhance the physiological effects of NO as mediated by cGMP. Whether these agents should have antimicrobial effects is unclear since they do not enhance NO levels per se; nevertheless, both drugs have exhibited antimicrobial activity (Table 7) and are being tested in COVID-19 trials (Table 9).107,108
Endogenous NO production may also be stimulated externally. First, nasal breathing promotes the production of NO from the paranasal sinuses and this has bronchodilatory, vasodilatory and potential antimicrobial activities.109 This natural defence mechanism may be attenuated with mouth breathing, as occurs with increasing age and obesity. Second, ultraviolet radiation (UVA and UVB) stimulates the release of NO from both keratinocytes and melanocytes; NO has multiple effects including attenuation of free radical damage, melanogenesis, blood pressure lowering110 and potentially protection against skin infections.
Recent research has focussed on the development of new antimicrobial NO delivery systems and some examples are listed:
• NO microspheres, e.g. biodegradable poly (lactic-co-glycolic acid) spheres loaded with S-nitroso-N-acetyl-D-penicillamine.111
• NO-releasing nanoparticles, with activity against Acinetobacter baumanii, C. albicans, Enterococcus faecalis, E. coli, K. pneumoniae, P. aeruginosa, S. aureus (MRSA), S. epidermidis, Trichophyton menatgrophytes.110,112
• Modified chitosan, e.g. against Trypanosoma cruzi,113 E. coli, S. aureus, Streptococcus mutans.114
• NO–metal complexes (zeolites), with activity against B. subtilis, Clostridium difficile, E. coli, P. aeruginosa, S. aureus (including MRSA).110
• NONOoates (diazeniumdiolates), e.g. with activity against C. albicans, E. coli.110,115
• NO coating of medical device surfaces and tubing,116,117 e.g. using S-nitroso-N-acetyl-D-penicillamine to kill Staphylococcus aureus and P. aeruginosa.
• RRx-001, a small molecule nitric oxide donor.118
NO sources can also be categorised by whether administration is local (e.g. cutaneous nitrite or intranasal preparations), systemic (e.g. dietary nitrate, L-arginine or L-citrulline, oral isosorbide or sildenafil, sublingual GTN, intravenous GTN or SNP) or mixed local and systemic (transdermal GTN). Local administration allows high and potentially cidal concentrations of NO to be achieved without unwanted systemic effects. Intravenous formulations might allow for systemic infections to be treated.
Most microbial studies presented above and in Tables 2-5 were performed in vitro and involved either inducing the L-arginine/NO pathway with cytokines (e.g. interferon gamma [IFN-γ] and/or lipopolysaccharide [LPS]) or with NO sources (such as NO gas, nitrite, 3-morpholinosydnonimine, S-nitroso-L-acetylpenicillamine or sodium nitroprusside). However, the inhibitory effect of NO on microbes in vitro does not represent the complex biochemical environment that they face in vivo including the presence of NO derivatives such as peroxynitrite, microbial production of NO, microbial resistance to NO and excess NO synthesis. Nevertheless, there are many ex vivo and clinical examples where NO has been effective. These issues are now discussed.
As already highlighted, oral bacteria (e.g. Corynebacterium pseudodiphtheriticum, Fusobacterium nucleatum, Nocardia spp., Prevotella melaninogenica, S. aureus, S. epidermidis, Veillonella spp.) are vital for the reduction of salivary nitrate to nitrite as part of the entero-salivary circulation; nitrite is further reduced to NO.119 This represents a symbiotic relationship between bacteria and the mammalian host; the host provides the nutrients and the environment in return for nitrite production,79 as in the absence of oxygen, nitrate and nitrite are commonly used by such bacteria as terminal electron acceptors for respiration.78
Oral consumption of nitrate and the resulting increase in nitrite in the oro-pharynx leads to salivary alkalinisation (pH ~7.0 to 7.5)120 and so reduction in detrimental bacteria and caries.77 Similarly, nitrate supplementation was associated with increased oral Rothia spp. and Neisseria spp, and diminished oral Prevotella spp. and Veillonella spp.; in parallel, plasma nitrite levels rose and systemic blood pressure fell.121 Salivary nitrite production is related to the abundance of oral-nitrate-reducing bacteria.122 In contrast, bacteria and yeast, in particular Lactobacillus spp., Streptococcus spp. and C. albicans, are key to the development of dental caries through the production of acid. Equally, antibiotics that kill nitrate-reductase-containing bacteria inhibit oral nitrite production and so increase the risk of oral thrush.123 Acidified nitrite has antibacterial activity against Helicobacter pylori in vitro,124 an experiment that likely mimics the scenario seen by these bacteria in the stomach after a nitrate/nitrite-rich meal.
The skin is a potent source of nitric oxide and production is increased with exposure to sunlight (specifically ultraviolet radiation) sufficient to lower blood pressure.125,126 Hence, skin-derived NO may form a natural dermatological antimicrobial defence. Numerous studies have demonstrated that topical acidified sodium nitrite reduces cutaneous infections due to a variety of viruses and bacteria (Table 7) although prophylaxis had to be continued in some cases since NO suppressed replication without necessarily being viro-toxic.127–132 Inorganic nitrate has been used for the treatment of human papilloma virus.133 Phase II clinical trials have found that acidified nitrite in cream reduced Leishmania major/tropica amastigotes and promastigotes with a reduction in cutaneous leishmaniasis129 and increased cure rates in tinea pedis.127 Novel NO agents are in development to treat skin conditions (Table 8).
| Target organism/disease | Commercial company | |
|---|---|---|
| NO gas for inhalation | Beyond Air | |
| NO gas for inhalation | INOmax | |
| NO-releasing solution for nebulisation (sodium nitrite and citric acid) | COVID-19 | 30 Technology |
| NO released from acidified nitrite via a semi-permeable membrane | Cutaneous S. aureus, E. coli257 | |
| NO macromolecular scaffolds,48,258 e.g. NO-releasing cyclodextrins259 | P. aeruginosa | Vast Therapeutics |
| Polymer-based chronic NO delivery systems e.g. for treatment of biofilms,260 | Cutaneous viruses (human papilloma virus, molluscipoxvirus), dermatophytes (Epidermophyton floccosum, Fusarium solani, T. rubrum, T. menatgrophytes) or yeast (Candida albicans/tropicalis, Malassezia furfur)247 | Novan |
| NO-releasing solution/gel (NORS2791) NO-releasing solution (NORS6491) NO-releasing nasal spray (NORS1002) NO-releasing nasal lavage (NORS4002) | Acne (Propionibacterium), fungal nail infections/onychomycosis (T. rubrum, T. interdigitale) Athlete’s foot (T. rubrum and T. mentagrophyte) Cold, ‘flu, COVID-19 Sinusitis | SaNOtize |
| NO-stimulating nasal spray (GLS-1200) | GeneOne Life Science | |
| Nitric oxide generating lozenges (sodium nitrite) | COVID-19 | Nitric Oxide innovations |
| NO-generating probiotic patches, e.g. based on Lactobacilli conversion of glucose to lactic acid, and acidification of sodium nitrite229,253 | S. aureus | McGill University, Canada |
| Nitroreductase-activated release of NO, e.g. by O2-(4-Nitrobenzyl) diazeniumdiolate261 or nitroaromatic-protected piperazine diazeniumdiolate262 | E. coli | Indian Institute of Science Education and research, Pune, India Colorado State University, Fort Collins, USA |
In animal and human experiments, NO substrate (L-arginine) and a NO donor (SNP) has been shown to improve the mucociliary activity of the upper respiratory tract134 suggesting a modulatory role for NO in nasal barrier function and clearance. Novel NO agents building on this observation are in development (Table 8):
Endogenous NO has potent pulmonary haemodynamic and bronchodilator effects physiologically. The importance of endogenous NO in preventing infection is apparent experimentally where inhibition of NO results in increased susceptibility to microbes including Leishmania spp., Mycobacterium spp. and Plasmodium spp.135 Similarly, NO sources are used therapeutically, for example sildenafil in the management of pulmonary hypertension (Table 7). In respect of airway epithelial cells, nitrite reduced P. aeruginosa biofilm growth.136 In infection, NO reduces pulmonary vascular resistance and intrapulmonary shunt, and improves oxygen partial pressure in patients with acute severe pneumonia.137 More specifically, inhaled NO improves arterial oxygenation enabling a reduction in inspired oxygen therapy and airway pressure support, and reduces lung infiltrates, in patients with severe acute respiratory syndrome (SARS).33 These findings continue after termination of NO therapy suggesting that NO has both pulmonary vasodilator and anti-SARS effects. Small uncontrolled clinical studies have suggested that iNO may be beneficial in COVID-19.138–142 iNO and novel NO agents are in development, primarily for COVID-19 at present (Tables 8, 9).
Dipyridamole, a phosphodiesterase 5 inhibitor, may also have similar beneficial effects in severe COVID-19.35 A phase II clinical trial found that L-arginine might have beneficial effects when given on top of conventional therapy for tuberculosis (Table 7).50
There may also be a role for dietary nitrate/inorganic nitrite in the prevention and treatment of urinary tract infections. Many of the lower urinary tract opportunistic organisms (e.g. E. coli) possess nitrate reductases, this forming the basis of urine dipstick detection of nitrite. In acidic urine conditions, nitrite is reduced to NO with toxicity to bacteria; for example, transferring nitrite-rich urine containing E. coli to a more acidic environment (e.g. pH 5.5) dose-dependently inhibited bacterial growth,143 an effect potentiated by vitamin C. The antibacterial potency is comparable to conventional antibiotics such as trimethoprim and nitrofurantoin.
This approach has been tested by filling urinary catheter retention balloons with nitrite and ascorbic acid, resulting in measurable amounts of NO outside the membrane and effectively killing two strains of E. coli in the surrounding urine.144 A similar approach found decreased bacterial counts and prevented biofilm formation by P. aeruginosa, K. pneumoniae, and Enterobacter cloace (but not E. coli or S. aureus).145
Last, instillation of bacillus Calmette-Guerin (BCG, an attenuated strain of Mycobacterium bovis) into the bladder is used for the treatment of superficial/non-muscle invasive bladder cancer and carcinoma in situ. BCG induces long-term increases in NOS activity in urothelial cells146,147 and the formed NO is toxic to the malignant cells. The use of BCG to provide non-specific protection against SARS-CoV-2 is to be tested148,149 although vaccination in infancy does not appear to protect against COVID-19 in adults.150
Nitrate (usually KNO3) and nitrite (NaNO2) have been used for millennia to preserve food, especially meat and fish.151 Food preparation leads to reduction of nitrate to nitrite, and nitrite inhibits bacterial growth, especially Clostridium botulinum, a key and severe cause of neurotoxin poisoning. Additionally, nitrite adds colour to food,60,152 flavour (in part by overcoming rancid tastes) and is an antioxidant.
NO donors have also been investigated for eradicating or dispersing biofilm organisms. For example, GTN synergises with citrate and ethanol in eradicating biofilms (related to S. aureus, MRSE, P. aeruginosa and C. albicans) in an experimental catheter lock model.153 Similarly, isosorbide mononitrate synergised with antibiotics to disperse then kill S. aureus.154 An NO-releasing contact lens has been developed to treat microbial keratitis due to P. aeruginosa and S. aureus).155
GTN may have improved outcome after infection in participants enrolled into the RIGHT-2 trial, a study where paramedics recruited patients with suspected stroke and randomised them to GTN versus sham. Overall, the trial was neutral.156 However, in a planned subgroup analysis of those participants with a final diagnosis of a non-stroke mimic, functional outcome was better with GTN.156 In a post hoc analysis of participants in this subgroup, GTN was associated with a better outcome in those with a final diagnosis of infections of the respiratory and urinary tracts which raises the possibility that NO was treating the infectious cause underlying the stroke mimic diagnosis.
As with the NO system, the prostaglandin-cyclic adenosine monophosphate-phosphodiesterase-3 (PG-cAMP-PDE3) system has similar vasculo-protective roles with anti-leucocyte, antiplatelet and anti-smooth muscle, and pro-endothelial effects. It is therefore interesting to note that prostaglandins (PGA1, PGJ2), including prostacyclin (PGI2 and analogues), have been reported to have antiviral effects.157–160 Whether drugs based on these161 or the PDE3 inhibitor, cilostazol, have efficacy against SARS-CoV-2 remains to be investigated. Further, endogenous NO and PGI2 work together in the vascular tree, and it is conceivable that their potential antimicrobial effects will similarly synergise. Their combination, in the forms of ISMN and cilostazol, have been tested after stroke162 but not yet reported for the prevention or treatment of infection.
The interaction between diet, nutrition state and vaccine effectiveness has been assessed in multiple studies, principally in low–middle income countries where vaccination is paramount, especially in children, and yet where malnutrition may be widespread. In a systematic review and meta-analysis of observational studies and randomised controlled trials, there was little suggestion that malnutrition had any effect on vaccine responses163; similarly, supplementation with vitamins and D, and iron and zinc, did not appear to modify responses. In preclinical studies, protein-energy malnutrition had limited influence on vaccine efficacy in mice.164 The effect of dietary nitrate levels on vaccine efficacy is unstudied.
If nitric oxide derivatives attenuate microbial infections, then the efficacy of vaccines based on live attenuated viruses and bacteria (such as measles, poliovirus, BCG) might be attenuated by treatment with NO. Although there are many factors known to alter vaccine effectiveness (e.g. age), the effect of NO has not been studied.
Many infections cause long-term morbidity with chronic fatigue syndrome (CFS) and symptoms including fatigue, tiredness, myalgia, cognitive impairment and depression. Example associated microbes include Borrelia burgdorferi (Lyme disease), Chlamydia pneumoniae (community acquired pneumonia), Epstein–Barr virus (infectious mononucleosis), human herpes virus 6 (exanthema subitem), human immunodeficiency virus (AIDS), polio virus, SARS-CoV-1 virus (SARS), SARS-CoV-2 (long-COVID) and West Nile virus (fever).165–167 Although CFS may represent chronic or latent infection, it is more likely to reflect the presence of post-infectious chronic inflammation. Hypothetically, these patients might benefit from inorganic nitrates in view of their positive effects on exercise performance (elite athletes take beetroot juice for this purpose) and cognition,76,95 and potentially antimicrobial effects, a question that needs addressing (Table 7). A phase II trial of L-citrulline is studying this approach in patients with post-polio syndrome.168
During severe infection, sepsis (defined as “life-threatening organ dysfunction caused by a dysregulated host response to infection”) often develops. Septic shock is a subset of sepsis and is a leading cause of death worldwide.169 It manifests as hyper- or hypo-pyrexia, altered mental state, hypotension, tachycardia, tachypnoea, hypoxia, anuria and/or lactataemia. This can occur with many infections due to:
• Gram negative bacteria: Bacteroides fragilis, C. pneumoniae, Enterobacter spp., E. coli, Haemophilus influenzae, Klebsiella spp., Legionella spp., Neisseria meningitidis, Proteus spp., P. aeruginosa, Yersinia pestis.63
• Gram positive bacteria: Clostridium spp., Enterococcus spp., Listeria monocytogenes, Staphylococcus spp., Streptococcus agalactiae/pneumoniae/pyogenes
• Viral: Adenovirus, Coronaviruses, Dengue viruses, Ebola virus, Enteroviruses, human immunodeficiency virus, Influenza virus (A and B), haemorrhagic fever viruses, Parechoviruses.170,171
• Fungi: Candida spp.172
• Protozoa: Plasmodium falciparum, Schistosoma mansoni.173
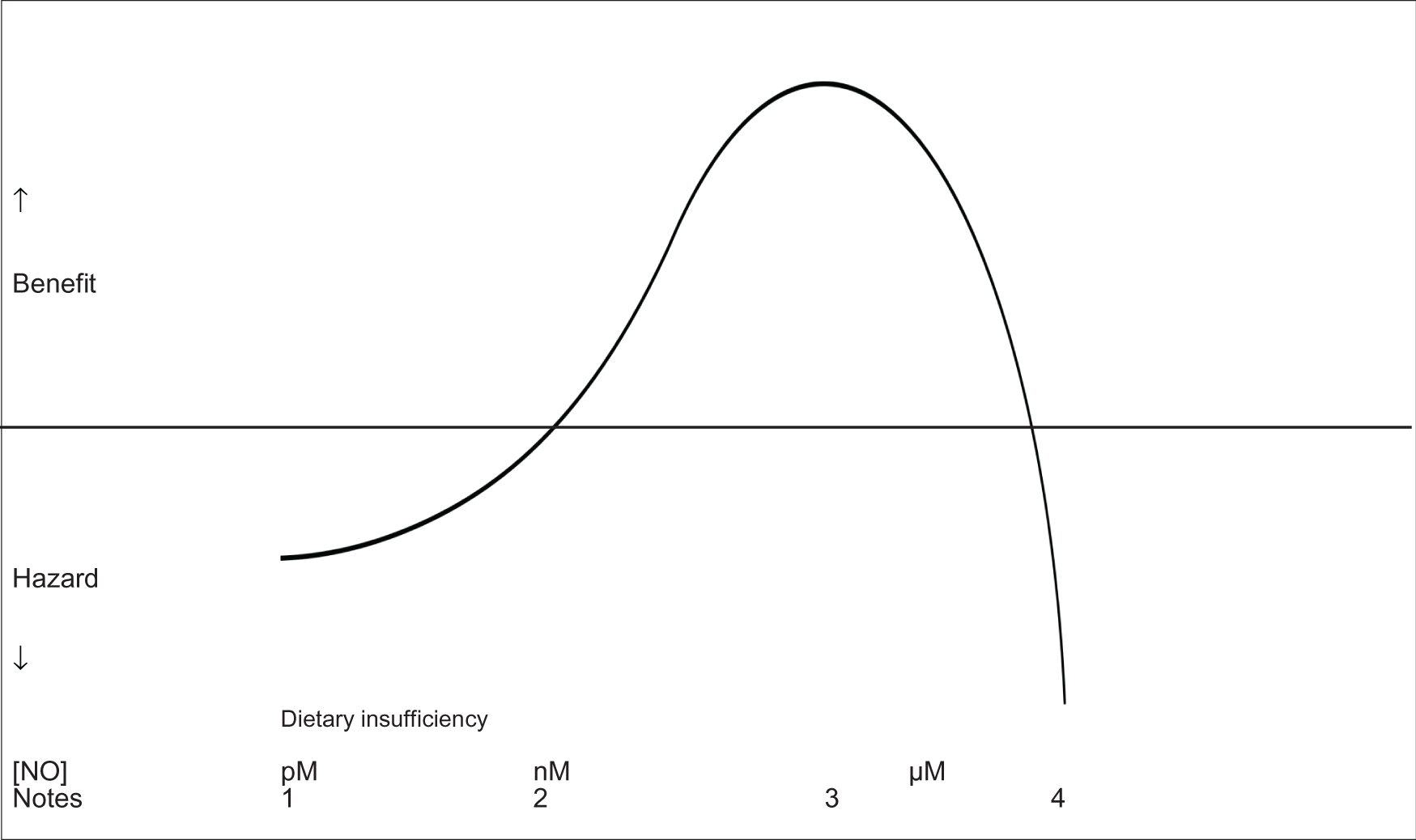
Typically, autoamplification of circulating cytokines (so-called cytokine storm) leads to excess NO synthesis, mostly derived from inducible NOS, leading to high circulating NO levels and the development of septic shock. In these circumstances, treatment with exogenous NO might be inappropriate. Trials of inhibiting endogenous NO synthesis with NOS-inhibitors in critically ill patients with sepsis have been reported although, disappointingly, did not improve outcome; indeed, the non-specific NOS-inhibitor, NG-methyl-L-arginine hydrochloride (L-NMMA, 546C88), was associated with increased death.174 It is not clear why inhibiting NO synthesis was ineffective but non-selective NOS inhibitors were used meaning that both toxic (iNOS) and beneficial (eNOS) sources of NO were inhibited; pharmacologically, such inhibitors will have reduced cardiac output, organ perfusion and tissue oxygenation. In the absence of licensed selective iNOS inhibitors, perhaps the analogous approach used in the management of hyperthyroidism using block (with carbimazole) and replace (thyroxine) might be effective, i.e. block NOS activity and replace with a low dose of a NO donor. That excess NO is dangerous does not mean that pharmacological doses of NO cannot be effective (Figure 1, Tables 6, 11) since all effective interventions in medicine have an inverted “U” dose response.
Some infections have opposing in vitro and in vivo responses to NO. For example activated macrophage-derived NO or NO donors such as SNAP reduced Trypanosoma brucei proliferation in vitro175,176 whereas endogenous iNOS-derived NO suppressed protozoa-antigen specific T-cell proliferative responses and so worsened infection, at least in infected mice.176 Intracellular protozoal infections are unlikely to be affected in this manner since macrophage-derived NO would be able to act directly on pathogens such as Leishmania major.177
Other infections do not appear, at least in vitro, to induce iNOS. For example, Cryptococcus neoformans failed to induce NOS in primed macrophages,178 apparently due to a lack of TNF-α secretion, probably because the polysaccharide capsule masked the signal for TNF-α secretion. Interestingly, non-encapsulated mutants of C. neoformans did induce endogenous NOS.
High levels of iNOS activation were antimicrobial in studies of malaria. Based on monocyte-derived mRNA levels in circulating blood, uncomplicated malaria was associated with increased levels of iNOS activation in contrast to patients with severe malaria who had lower levels.179 The dual effects of NO in malaria, i.e. both low and high levels appear to be hazardous, are further reviewed.180,181
Over recorded history, most epidemics and pandemics have resulted from viral infections including Ebola (viral haemorrhagic fever), influenza (H1N1, H2N2, H3N2, H3N8), HIV-1 (AIDS), polio (poliomyelitis), smallpox, yellow fever, zika or corona (OC43, MERS-CoV, SARS-CoV-1/2) viruses. Bacterial pandemics have resulted from Vibrio cholerae (cholera), S. enterica (typhoid fever) and Yersinia pestis (plague). Studies in vitro have reported findings suggesting that NO can reduce infection for some of these pathogens (Tables 2, 3) but information appears to be lacking for smallpox, yellow fever, zika and cholera (Table 10).
With multiple pandemics over the last 100 years, it is only inevitable that further ones will occur and some, like COVID-19, will comprise a “global catastrophic biological risk”.182 Global pandemics will most likely be caused by a respiratory-spread virus that crosses over from animals such that humans have no inherent immunity to it. Likely candidates include orthomyxoviruses (especially influenza A viruses such as H7N9), paramyxoviruses (e.g. measles, mumps, croup), pneumovirus (e.g. human metapneumovirus), coronaviruses and picornaviruses (especially rhinoviruses and enteroviruses). All of these have had strains that have crossed from animals to humans. This emphasis on RNA viruses is because DNA viruses tend to have lower mutation rates and, therefore, evolve more slowly and are less likely to escape the human immune system within the first rounds of infection. Nevertheless, DNA viruses, such as pox or herpes viruses from great apes or monkeys, do have the potential to jump species. Non-viral causes of pandemics are less likely since most bacteria will be treatable with broad-spectrum antibacterial agents, most fungi are thermally restricted, and prions would require massive food contamination (and only spread slowly).182 Protozoa are usually thermally restricted although global warming may allow malaria to spread more widely in temperate zones.
Unfortunately, pandemics/epidemics may co-exist as seen with SARS-CoV-2 and dengue in Brazil,183 and both with S. enterica in Pakistan;184,185 in part, this reflects increasing travel with aircraft providing a portal for numerous microbes.186 Of theoretical concern was the potential for COVID-19 and epidemic influenza to co-exist during winter in the Northern hemisphere, this possibly leading to a dramatic increase in deaths.187 Nevertheless, ‘flu rates were very low in both southern and northern hemisphere 2020 winters, presumably due to hands, face, space, mask and fresh air measures. All-in-all, the absence of a true broad-spectrum of antiviral agents is a major concern188 and a potential agent such as NO with antimicrobial effects that extend beyond viruses would be most welcome.
One possible explanation for the observation that COVID-19 outcomes are worse in older people, males, black or Asian ethnicity, and those with co-morbidities such as diabetes, hypertension, stroke and chronic lung disease,189 is that these groups have lower vascular NO activity6,7,11 and so mount a sub-optimal host response against infection. Increasing NO availability is therefore a potential therapeutic strategy. Several NO sources have potential relevance to preventing and treating COVID-19. L-arginine, sodium nitrite, GTN, SNP, NO and dipyridamole each have clinical antimicrobial activity and can be administered, variously, orally, intravenously or as NO gas in the intensive care unit. Transdermal GTN, and oral ISMN, dipyridamole and sildenafil may be administered in the community or hospital. Of these, NO gas, dipyridamole and sildenafil are already being tested for preventing or treating COVID-19 (Table 9). It remains to be determined if increasing dietary nitrate may be a cost effective and safe intervention of widespread health relevance for the prevention of COVID-19 and, indeed, other emerging, pandemic, epidemic or endemic infections. Recent trial evidence provides indirect supporting evidence for the potential anti-SARS-CoV-2 effect of NO. First, dexamethasone and tocilizumab reduced death in patients in intensive care units,190,191 and these agents and NO share anti-inflammatory effects. And second, interferon-ß reduced the need for intensive care in COVID-19 patients;192 type I interferons increase iNOS activity and so have antimicrobial effects, as seen with L. major and Burkholderia pseudomallei.193,194
Nitric oxide is a fundamental molecule with wide-ranging and potent vascular, anti-platelet, anti-inflammatory and tumoricidal effects. Further, there is a large volume of literature spanning the last 30+ years demonstrating that NO also has potent in vitro antimicrobial effects on a wide variety of viruses, bacteria, protozoa, fungi and yeasts; these are supported by a modest number of in vivo studies. Further, several positive clinical phase II trials of NO have been reported in viral, bacterial, protozoa and fungal infections, these relating particularly to skin and respiratory infections administered by cream and gas respectively. Although not from randomised trials, there is also evidence that dietary nitrate modifies the oral biome and so reduces dental caries.
However, these results cannot be considered persuasive on their own. First, few neutral or negative studies have been reported suggesting that there may be a risk of publication bias. Second, conflicting data in some dual-protocol studies with positive in vitro and neutral/negative in vivo data suggest that although NO is antimicrobial per se, the local tissue environment may overcome or reverse this effect. Third, organic nitrates can suffer from tolerance and may lead to the generation of reactive NO species such as peroxynitrite and S-nitrosothiols which might exacerbate rather than attenuate infection. Fourth, resistance may develop although this seems unlikely to be a generic issue, not least because NO levels can change, and be changed, much faster than any microbe can raise defensive mechanisms. Fifth, some microbes can produce their own NO and use this to resist the oxidative stress induced by external NO and its derivatives. Sixth, excess NO production is associated with the development of septic shock which might suggest that any NO is ineffective. Potentially, unsuccessful trials of non-selective NOS-inhibitors in severe sepsis may have confused the issue, perhaps by suggesting that treatment with NO is not important in infection. Seventh, positive clinical studies have been performed in environments where very high local concentrations of NO can be achieved and without the risk of reactive responses, in particular on the surface of tissues such as cream on the skin, dietary nitrate in the mouth, nitrite in the stomach, NO gas in the lungs and nitrite in the bladder; whether NO is effective as an antimicrobial within tissues and the vascular tree remains to be determined.
There are many sources of NO suitable for studying the prevention and treatment of milder infections in the community and hospital (e.g. topical sodium nitrite, oral NO donors such as ISMN, or oral PDE5 inhibitors such as dipyridamole or sildenafil), and treatment of serious infections in hospital (e.g. intravenous L-arginine, sodium nitrite or NO donors such as GTN or SNP, of NO gas). NO may also be delivered via a high nitrate diet, thus offering a widely available and inexpensive public health approach to potentially reducing and attenuating the severity of infections worldwide. This approach has the added advantage that such diets are already known to reduce vascular disease and some cancers, and possibly other inflammatory diseases and dementia.
In summary, the wealth of in vitro data suggest that NO has generic antimicrobial effects. However, some data suggest that NO may be ineffective or even hazardous and these reinforce our view for the need for large scale clinical trials of NO donors in the community and hospitals to prevent and treat infections. Although such studies need to focus urgently on the COVID-19 pandemic (especially with the lack of broad spectrum antiviral agents188), other pathogens also need to be targeted. However, patients with established septic shock should not be administered NO donors to avoid exacerbating vascular collapse. One utopian vision would be demonstration that high dietary nitrate intake produces pre- or post-exposure prophylaxis against infections and their severity in the community whilst NO donors are effective antimicrobial treatments for use by general practitioners and in hospitals.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (2)
Much is known of the protective - and pathological - effects triggered by NO metabolites in the mammalian immune response to asexual plasmodia, both during uncomplicated infection of the peripheral blood (PB) and cerebral malaria (CM) sequalae of parasitised erythrocyte sequestration.
Investigation of NO during malaria infection extends back to the late 1980s and throughout the 1990s, principally in murine models including P. berghei (mostly for CM) and P. chabaudi (mostly for PB). In many aspects, the pioneering research of the groups of Langhorne, Phillips, Stephenson, Clark and Hunt, among others, led the field for what was then known about the role of NO in immunity to protozoan infection, as well as more broadly.
The implications of this collective body of work have been extrapolated extensively to other intracellular pathogens as they were reached by exploiting a tractable system for dissecting the evidently subtle balance between immune protection and immunopathology. The main findings remain very relevant to a contemporary consideration of the part played by NO in prevention and treatment of infectious diseases.
Much is known of the protective - and pathological - effects triggered by NO metabolites in the mammalian immune response to asexual plasmodia, both during uncomplicated infection of the peripheral blood (PB) and cerebral malaria (CM) sequalae of parasitised erythrocyte sequestration.
Investigation of NO during malaria infection extends back to the late 1980s and throughout the 1990s, principally in murine models including P. berghei (mostly for CM) and P. chabaudi (mostly for PB). In many aspects, the pioneering research of the groups of Langhorne, Phillips, Stephenson, Clark and Hunt, among others, led the field for what was then known about the role of NO in immunity to protozoan infection, as well as more broadly.
The implications of this collective body of work have been extrapolated extensively to other intracellular pathogens as they were reached by exploiting a tractable system for dissecting the evidently subtle balance between immune protection and immunopathology. The main findings remain very relevant to a contemporary consideration of the part played by NO in prevention and treatment of infectious diseases.