Keywords
rubber, drought stress, water content, adaptation, abiotic stress
This article is included in the Agriculture, Food and Nutrition gateway.
rubber, drought stress, water content, adaptation, abiotic stress
We have revised to incoropreta Reviewer 's 1 comments and suggestions. We updated the aim of study in the Abstract and Introduction (the last paragraph). We also revised the first line of 2nd paragraph and 4th paragraph of Introduction. We revised in the 2nd paragraph of Discussion as well as 4th and 5th paragraph in the Discussion. The last revision is the Conclusion.
See the authors' detailed response to the review by Mohamed sathik Thirruvithamkottil
See the authors' detailed response to the review by Maryam Nazari
In rubber plants, drought can cause a delayed maturation phase, short tapping period, slow latex flow, dry latex, increased dry tapping grooves, and even tree death.1 Drought is one of the main abiotic stresses affecting yield and productivity in almost all crops.2 Hence, its significance overshadows that of other environmental factors because it interferes with plant growth and development and disrupts production and performance. Water is an important component of the protoplasm and makes up 85%–90% of the total weight of the plant tissue. Water is also a vital reagent in photosynthesis and hydrolysis reactions. Additionally, it acts as a solvent for salts, gases, and other substances transported between cell tissues to maintain cell growth and leaf shape stability.3
One of the primary sources of natural rubber producing plants was found in the Amazon basin, South America.4 Optimal conditions for the growth of rubber plants include high temperature (28 ± 2 °C), high humidity, and rainfall of 2000–4000 mm/year.5 Rubber plantations in marginal areas, such as Central India, southern China, northern and northeastern Thailand, and eastern Indonesia, experience occasional drought. Indonesia has a wide drought area of about 122.1 million ha, which is not optimally exploited due to limited water resources.
The response caused by drought with regard to plant is quite complex because it involves changes in morphology, physiology, and metabolism. The initial response to drought stress is the loss of turgor pressure, which results in reduced growth rate and leaf senescence. Drought changes the source–sink relationship and affects the translocation of photosynthate to produce fruit quickly for certain crops.6 The fastest response to a water deficit is the stomatal closure to protect plants from water shortages. Water deficit results in abscisic acid (ABA) biosynthesis, which triggers stomatal closure and causes a decrease in intracellular CO 2 levels and the inhibition of photosynthesis.7 Water shortages do not always promote these responses in all plant species. Lack of intracellular CO 2 due to prolonged stomatal closure leads to the accumulation of reactive oxygen and nitrogen species, which damages the photosynthetic apparatus.8 Besides that, the presence of osmoprotectants, such as proline, trehalose sugar, glycine betaine, donomitol, and mannitol maintain the growth and productivity of a plant experiencing drought stress.9–11 The presence of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), and glutathione reductase (GR), in cellular and cytoplasmic organelles play an important role in the detoxification of these reactive oxygen species (ROS) and enable plant cells to activate various stress sensors, which will then activate various signal paths.
Inhibited growth is a typical symptom of drought stress.12 The consequent physiological, biochemical, and molecular changes affect various cellular processes, thereby reducing the quantity and quality of the plant yield. The combined rise in atmospheric CO2 concentrations and global air tempeature may significantly alter plant water use.13 Pasaribu et al, reported based on bio-informatics, there are 20 proteins related to drought stress in rubber plants.14 This study aims to identify drought tolerant traits in order to select or identify drought-tolerant clones at juvenile stage.
Analysis of physiological characteristics was carried out at the Physiology and Protection Laboratory of the Unit Research SungeiPutih, Galang, Deli Serdang, North Sumatra. The study was carried out in a greenhouse during June 2020–May 2021. The materials used were red-yellow podzolic soil (water content = 3.9, pH = 4.5, C-organic = 0.92, N = 0.15, P2O5 = 2.13), compound fertilizer, Dithane M-45, and Triko-SP Plus. The tools used included polybags (18 × 35 cm and 50×60 cm), a hoe, a soil sieve, bucket, watering can, a 100 kg UK scale, an analytical balance, object glass, deck glass, binocular microscope, water bath, vortex, UV spectrophotometer, filter paper, test tube, gloves, mask, tissue, distilled water, mortar, beaker, micropipettes 1 ml and 100 μl, stirrer, 15-watt lamp, microcentrifuge, microwave, and others.
The plants age was 6-12 months, planted in polybags measuring 50 × 60 cm filled with 30 kg ultisol soil. The samples used were normal leaves and not affected by leaf fall disease such as Oidium, Colletotrichum, Corynespora and Pestalotiopsis. The leaves were taken between at 08.00-09.00 AM. The average temperature in the greenhouse during the six months observation was 25.9 °C (morning), 33.9 °C (midday) and 30.4 °C (afternoon). The average temperature outside the greenhouse was 22.9 C (morning), 28.1 °C (midday) and 25.5 °C (afternoon). The average humidity was 92.6% (morning), 62.9% (midday) and 69.2% (afternoon).
The study was arranged based on a factorial randomized block design (RBD). The first factor was the type of clone, consisting of seven types, namely C1: IRR 425, C2: IRR 428, C3: IRR 429, C4: IRR 434, C5: IRR 440, C6: RRIC 100, and C7: BPM 24. The second factor was water content, consisting of three levels, namely: W1: 30%, W2: 60%, and W3: 90%. Each experimental unit was repeated three times, and as many as 63 samples were observed.
Observations were carried out six times on physiological characteristics, with an intervals of three weeks. If the test of variance obtained significantly different treatments, then the Tukey distance test of 0.5% was carried out.15 The characteristics observed were total sugar content,16 chlorophyll a, b, and total,17 proline,18–21 super peroxidase dismutase (SOD),22 peroxidase dismutase (POD),23 APX enzyme,24 and hydrogen peroxide (H2O2).22
A step-by-step description of the procedure to analyze sugar content, proline, chlorophyll a, b, and total, SOD, POD, H2O2, and APX has been deposited in protocols.io and is available at dx.doi.org/10.17504/protocols.io.5jyl8je1dg2w/v1.
The total sugar content analysis showed a significant effect in all the observations except the first one (Table 1).
The total sugar content (μM) in different clones, different water content (%), and interactions between clone type and water content (%).
The total sugar content in the six observations carried out on tested clones was consistent. The RRIC 100 clone had the highest total sugar content four times, and the IRR 425 clone had the lowest four times.
The total sugar content analysis at different water levels generally showed a significant effect, except for the initial observation. This indicates that water content affects the total sugar content of the tested clones (Table 1).
Analysis of the total sugar content due to the interaction between the clone type and water content level (30%, 60%, and 90%) showed significant differences, except in the first observation (Table 1).
What is interesting about these results is that the highest accumulation of total sugar was seen in the application of 30% water content. Meanwhile, the effects were quite diverse across the different clone types. The IRR 429 had the highest total sugar in three observations (second, fourth, and fifth). The RRIC 100 had the highest total sugar in two observations (third and sixth). The two clones, RRIC 100 and IRR 429, also had the lowest total sugar in the fifth and sixth observations, respectively.
Two forms of polynomial curves can be the effect of water content and can be shown by the orthogonal polynomial regression obtained from three levels of water content, namely linear and cubic curves. The results of the analysis show that the linear curve shows a real effect. Figure 1 presents the linear curve regression pattern formed in detail. It demonstrates that the lower the water content added to the growing media, the higher the total sugar content derived from the leaf analysis of several rubber clones of IRR 400 series, RRIC 100, and BPM 24.
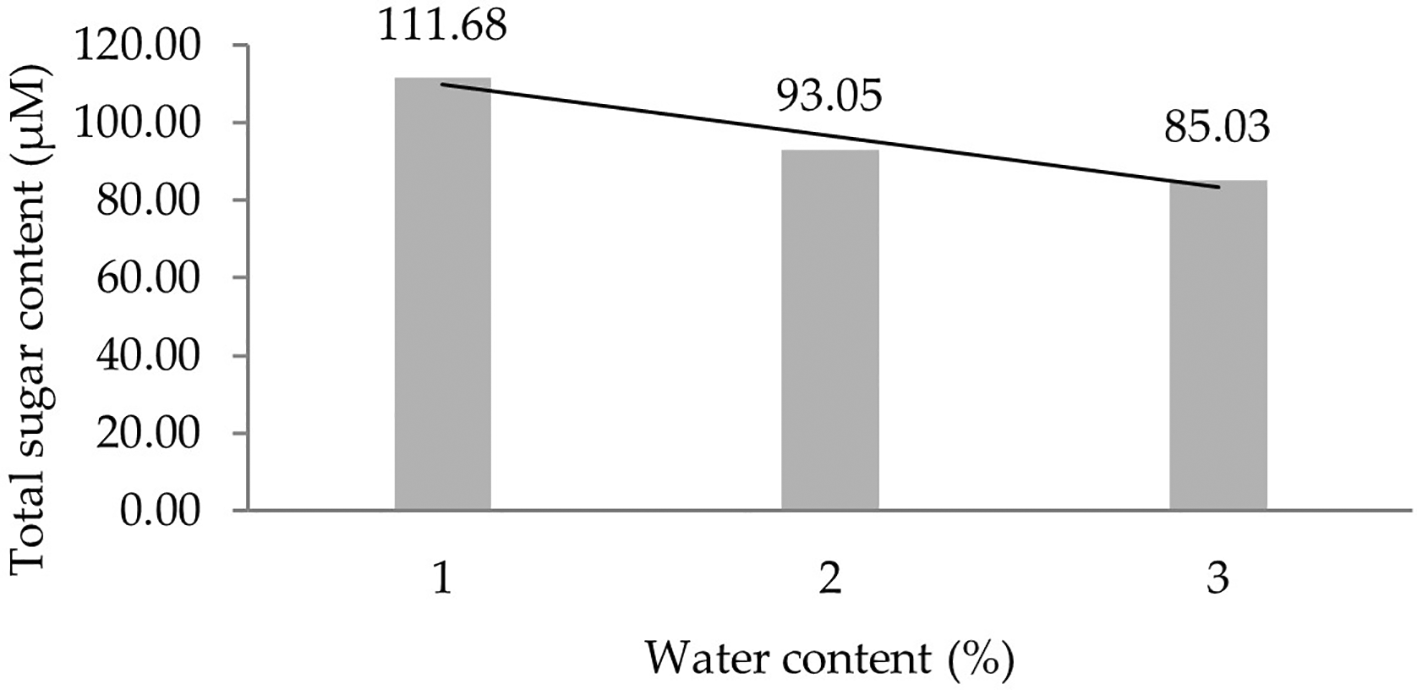
1: 30%; 2: 60%; 3: 90%.
Table 2 depicts the proline analysis of clone types treated with different water content levels and shows significantly different effects in all observations.
Proline levels (mg g-1) in different clones, different water content (%), and interactions between clone type and water content (%).
The results of proline analysis at different water content levels showed significantly different effects in all observations (see Table 2).
The proline analysis caused by the interaction between rubber clones IRR 400 series, RRIC 100, and BPM 24 and given water content (30%, 60%, 90%) showed significantly different effects in all observations, as displayed in Table 2.
The assessment of orthogonal polynomial regression showed a linear curve, where the water content at the 30% level had the highest proline value. The orthogonal polynomial linear curve pattern of proline characteristics of several rubbers, namely, IRR 400 series, RRIC 100, and BPM 24, is shown in Figure 2.
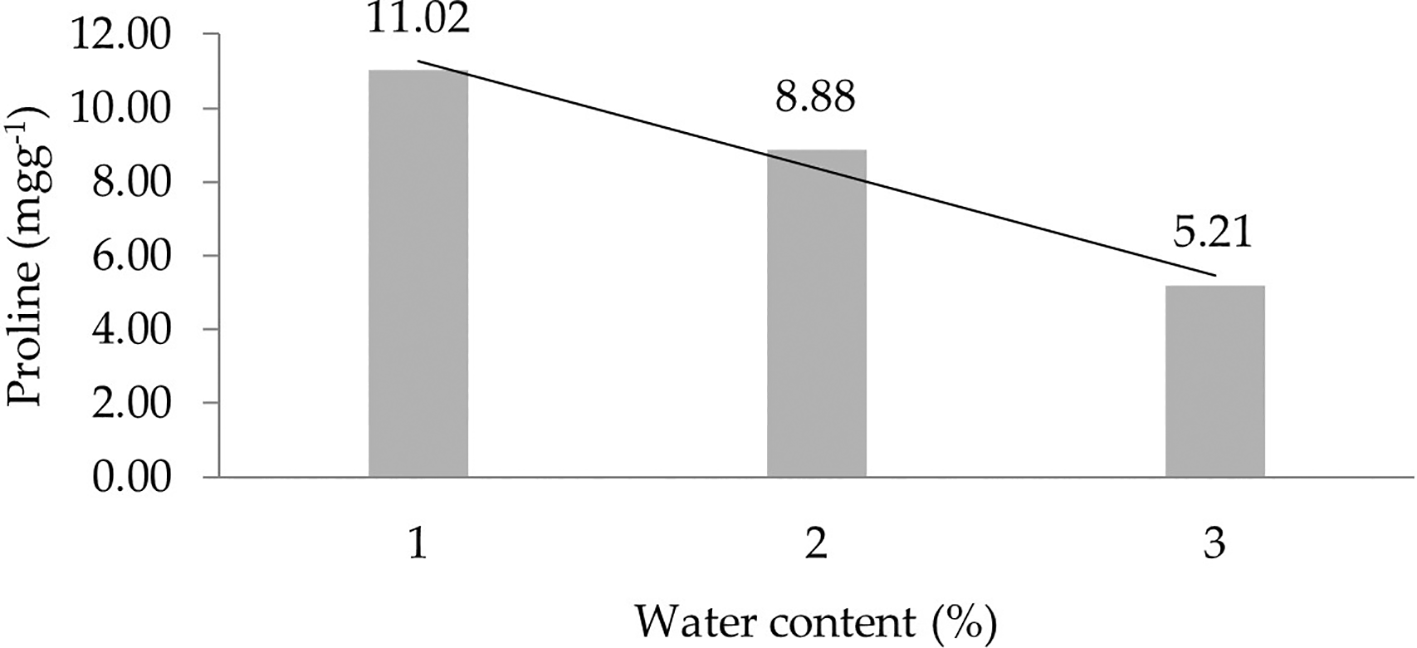
1: 30%; 2: 60%; 3: 90%.
The chlorophyll-a analysis on the different clone types showed a significant effect, except for the first observation, as displayed in Table 3.
Chlorophyll a levels (μg mg-1) in different clones, different water content (%), and interactions between clones and water content (%).
Table 3 shows the chlorophyll-a analysis at different water contents, which demonstrated a significant effect in all six observations.
Analysis of chlorophyll-a levels due to the interaction between clones and water content (30%, 60%, 90%) showed significant differences in all six observations (Table 3).
The results of the chlorophyll b analysis with different clone types showed significantly different results, except for the first observation (Table 4).
Chlorophyll b levels (μg mg-1) in different clones, different water content (%) and interactions between clones and water content (%).
The results of the analysis of chlorophyll b levels at the given water contents showed a significant effect in all six observations (Table 4).
The analysis of chlorophyll b levels due to the interaction between clones and water content (30%, 60%, 90%) showed significant differences in all six observations, as shown in Table 4.
The assessment of the orthogonal polynomial regression showed a linear curve, where the water content at the 30% level had the highest chlorophyll b value. The orthogonal polynomial linear curve pattern of the chlorophyll b characteristics of several rubber clones of IRR 400 series, RRIC 100, and BPM 24 can be seen in Figure 3.
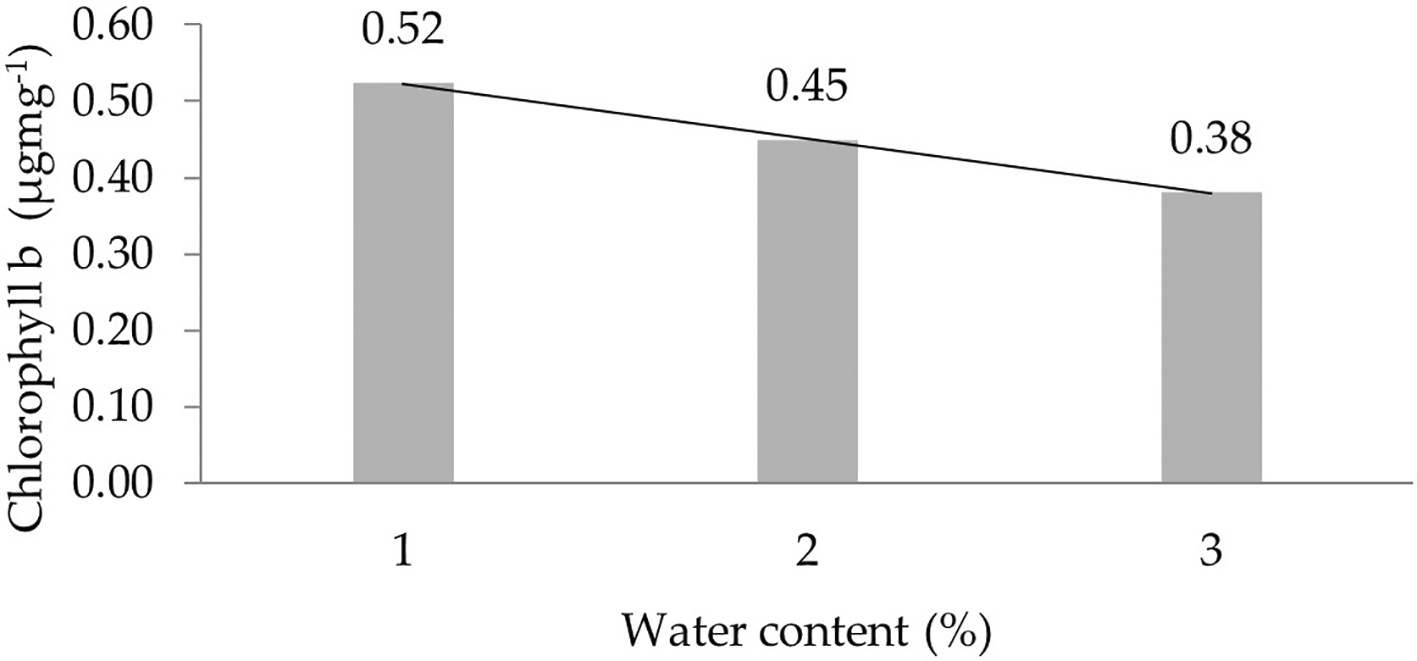
1: 30%; 2: 60%; 3: 90%.
The analysis results of total chlorophyll content with different clone types showed significantly different results except for one observation (Table 5).
Total chlorophyll content levels (μg mg-1) in different clones, different water content (%), and interactions between clones and water content (%).
The results of total chlorophyll content analysis with the given water content showed a significant effect in all six observations, as depicted in Table 5.
The analysis of total chlorophyll content levels due to the interaction between IRR 400 series, RRIC 100, and BPM 24 and water content (30%, 60%, 90%) showed significant differences in all six observations (Table 5). Orthogonal polynomial regression shows a linear curve, where the water content at 30% has the highest total chlorophyll content value. The linear curve shows that the total chlorophyll content increases with the decreasing water content. The orthogonal polynomial linear curve pattern of total chlorophyll content of several clones of IRR 400 series, RRIC 100, and BPM 24 can be seen in Figure 4.
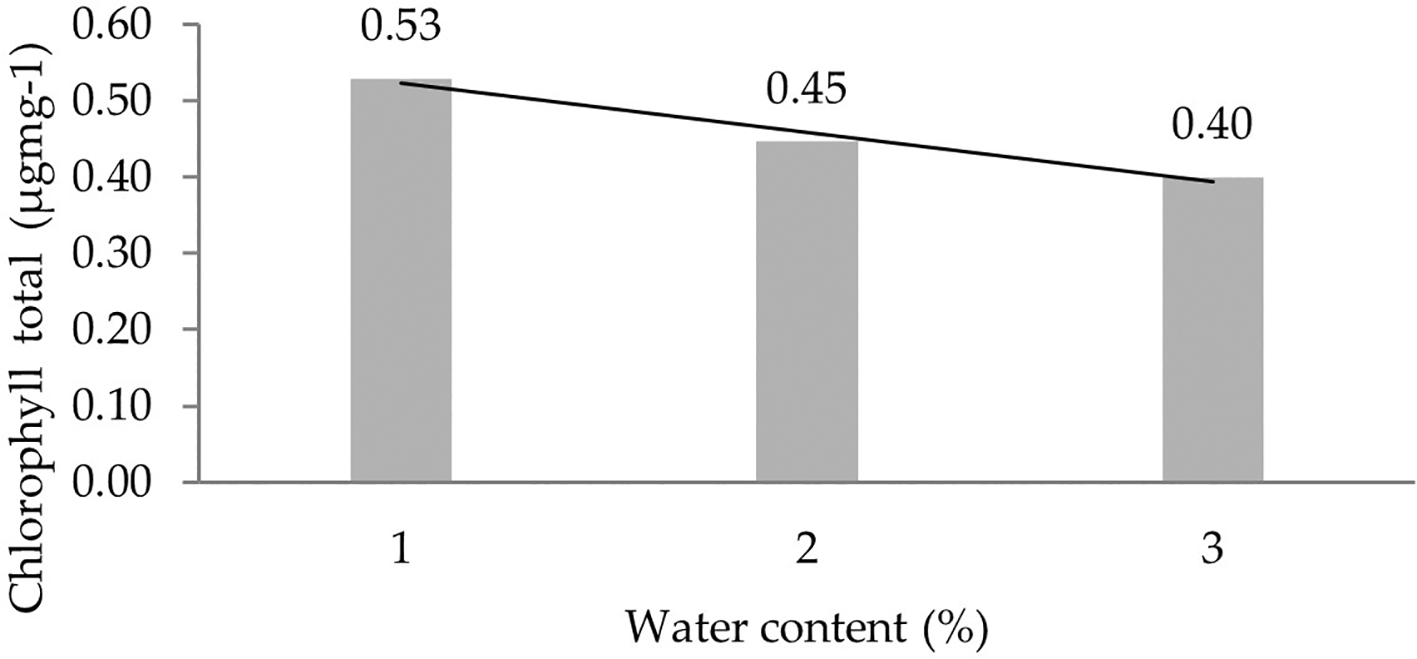
1: 30%; 2: 60%; 3: 90%.
The results of total the H2O2 analysis with different clone types showed a significantly different effect in two of the observations (third and fourth) (Table 6).
H2O2 levels (μmolg-1) in different clones, different water content (%), interactions between clones and water content (%).
The results of H2O2 analysis at different water content levels did not show significant differences in any of the observations (Table 6).
The analysis of H2O2 levels (μmol g-1) due to interactions between IRR 400 series, RRIC 100, and BPM 24 and given water content (30%, 60%, 90%) showed a significantly different effect in just one observation (fourth) (Table 6).
The effect of water content on the H2O2 characteristic shows a linear regression curve based on orthogonal polynomials. This indicates that the lower the water content, the higher the concentration of H2O2. The linear regression pattern between H2O2 content and water content can be seen in Figure 5.
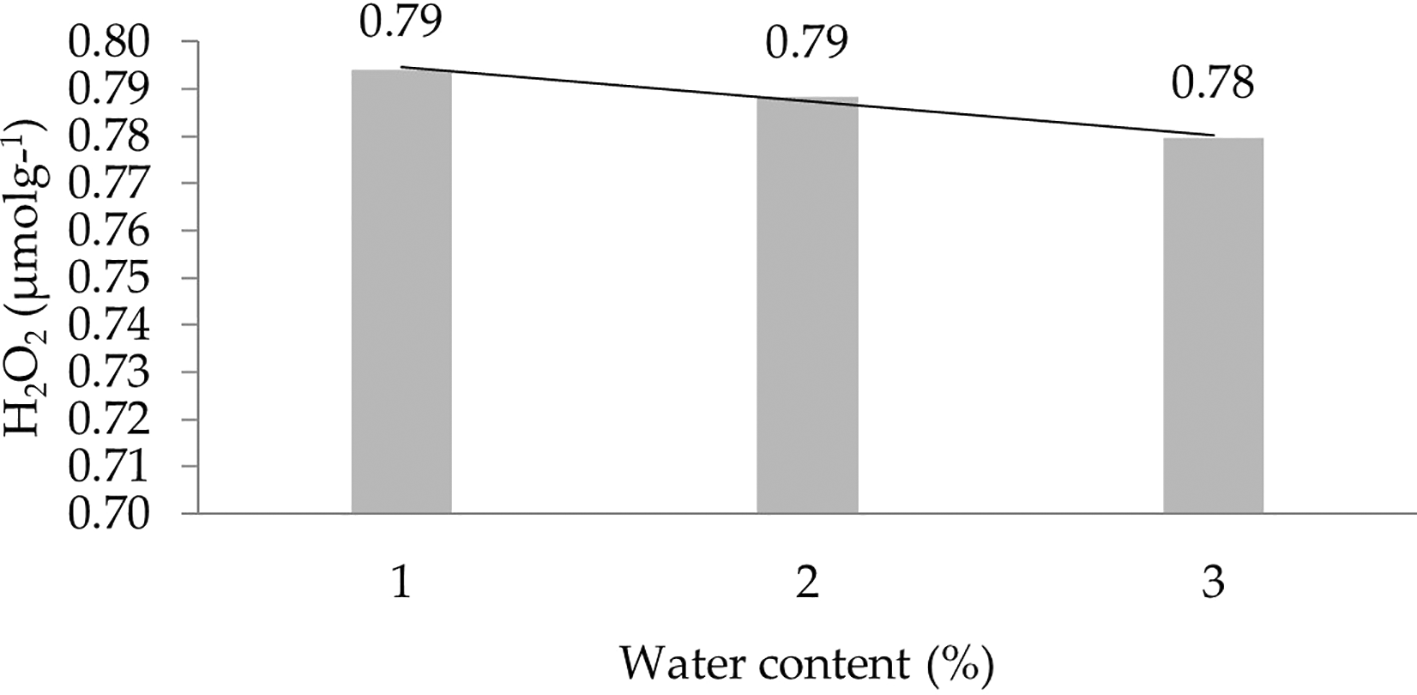
1: 30%; 2: 60%; 3: 90%.
The results of APX analysis with different clone types were not significantly different in any of the observations (Table 7).
APX levels (units mg-1) in different clones, different water content (%) and interactions between clones and water content (%).
The analysis results of APX at different water content levels were not significantly different in any of the observations (Table 7).
The analysis of APX levels (units mg-1) due to the interaction between IRR 400 series, RRIC 100, and BPM 24 and water content (30%, 60%, 90%) did not show any significant differences in any of the observations (Table 7).
The SOD analysis with clone types showed a significant difference in three of the observations (Table 8).
SOD levels (units mg-1) in different clones, different water content (%) and interactions between clones and water content (%).
Analysis of SOD (units mg-1) levels at different water content levels showed significant differences in three of the observations, as depicted in Table 8.
The analysis of SOD levels due to interaction between IRR 400 series, RRIC 100, and BPM 24 and water content (30%, 60%, 90%) showed significant differences in two observations (Table 8).
The POD analysis with different types of clones showed significant differences in two observations, as shown in Table 9.
POD levels (units mg-1) in different clones, different water content (%), interactions between clones and water content (%).
The analysis of POD levels at different water content levels showed a significant difference in one observation, as depicted in Table 9.
The analysis of POD levels due to interaction between IRR 400 series, RRIC 100, and BPM 24 and given water content (30%, 60%, 90%) showed a significant difference in just one observation (Table 9).
Plants respond and adapt to the change in environmental conditions such as drought stress by altering its physiological and biochemical activities to overcome such stressful situation.9,10
The plants respond to the environmental stress by altering its physiological activities so as to maintain its growth and development without succumbing to the strees by producing compounds that render tolerance. The accumulation of osmoprotectants is a key biochemical property in plants tolerant to abiotic stress,10, 21 and there is clear evidence that osmotic adjustment sustains crop yields under drought stress.9 Drought stress causes changes in amino acid metabolism. The accumulated solutes protect cellular proteins, organelles, membranes, and various enzymes against drought stress.
Several physiological characteristics were analyzed to see the effect of water content on IRR 400 series, RRIC 100, and BPM 24 rubber clones. Some of the dissolved substances assessed in this study were total sugar, proline, and chlorophyll (a, b, total). The correlation of total sugar content to each clone showed different effects. Each clone had altered levels of total sugar content under stress conditions. The RRIC 100 is a drought tolerant clone in the field. The increase in total sugar content was seen in most observations of the water content treatment. The interaction between clone type and water content can increase total sugar content, especially when the water content added is 30%. Initial hypotheses suggest that each clone has the ability to adapt to water shortages. The accumulation of soluble sugars in plant cells subjected to drought stress is responsible for the osmotic adjustment.25 Sugar accumulation in drought-stressed plants is controlled by several mechanisms that affect soluble sugar formation and transfer in leaves.26 Similar results of increased total sugar accumulation have been observed in drought-stressed soybeans26 and sugarcane.27
This study showed different proline values among the tested clones. All six observations indicate that clones have the ability to survive drought. The IRR 425 clone had the highest proline levels in three observations. Meanwhile, the IRR 440 had the lowest proline levels in four observations. Assessing the proline characteristic, the initial assumption was that IRR 425 had a stronger adaptation compared with other clones, especially the IRR 440. Regarding different water contents (30%, 60%, 90%), the proline levels at 30% were greater than at 60% and 90%. This indicated that a higher amount of proline accumulated when the water content was lower in the growth medium. Proline is an important amino acid as it is an osmotic-compatible molecule and has the potential to form a defense system to increase drought tolerance. Proline acts as an antioxidative defense molecule and causes stress signaling.12 It is classified as an osmoprotectant, as it increases hyperosmolarity and antioxidant enzymes’ activity.28 Increased proline content in drought-stress plants can maintain turgor for continued growth.29 Hence, proline accumulation correlates with osmoprotection.30 The interaction between clone types and moisture content indicated that each clone showed a different effect in all six observations. The clones had high proline levels when treated with 30% water content. This shows that the clonal factor still has to be planted in the field which is classified as a drought environment. The proline content has been shown to increase about 10-fold in mungbean,31 maize,32 millet,12,33,34 nyamplung,35 and soybean26 under drought stress.
Chlorophyll is the main pigment found in chloroplasts.36 The three main functions of chlorophyll in the photosynthesis process are harnessing solar energy, triggering CO 2 fixation to produce carbohydrates, and providing energy for the ecosystem as a whole. Chlorophyll a and chlorophyll b absorb the most light in the red part (600–700 nm) and absorb the least in the green part (500–600 nm).35–37 In this study, it was seen that chlorophyll a, b, and total levels at 30% were higher than at 90%. The genotype BC678 and BC404 of corn cultivars showed resistant to drought stress have highest chlorophyll index and higher potential yield.38
This is presumably because the rubber plant is a perennial plant that is able to adapt when water conditions are on a small scale due to the root structure of the rubber plant. It is a bud-grafted clonal material (of each varuet/genotype) which does not have a tap root to mine water from the deeper soil. In addition, when stressed, the lateral roots will grow even more to take advantage of the existing water on the surface.39
This study was conducted in greenhouse conditions where the water provided to the plants will not be lost as water treated gradually. Even though the polybags made small holes to drain excess water during its pre-treatment conditions. Besides that, the polybag surface is also covered by plastic to minimize the occurrence of evapotranspiration from the planting medium. Another factor that can be used as a cause of high chlorophyll levels is ability to adjust osmolyte content (sugar, proline, glycine betaine, ABA, ethylene, and others), and the nature of the rubber plant itself (plants that will shed leaves naturally every year).
Antioxidants are active substances that naturally detoxify free radicals (ROS). The presence of oxidative stress and an abundance of antioxidants are important activities for metabolic protection when plants are under stress. ROS in the form of free radicals and peroxides are molecules derived from oxygen metabolism. The toxic effects of ROS can be countered by antioxidant enzymatic as well as non-enzymatic systems, such as SOD, CAT, APX, GR, ascorbic acid (AsA), tocopherols, glutathione and phenolic compounds, and others. Typically, each cellular compartment contains more than one enzymatic activity that detoxifies ROS. The presence of these enzymes in almost all cells plays an important role in ROS detoxification for plant survival.40
H2O2 has several important roles in various biochemical and physiological processes. Long plant life and long growth processes result in H2O2 crossing cellular membranes and potentially acting as a signal in the signal transduction pathway of stress. This pathway triggers various responses of the adaptation process in the environment where the plant is cultivated.41 High levels of H2O2 cause oxidative stress, which then causes cell damage and death.42 However, optimal levels of H2O2 can increase tolerance to abiotic stresses through modulation of various physiological processes, including photosynthesis, opening and closing of stomata, osmotic adjustment, and ROS detoxification.41,42 ROS detoxification is very important in maintaining the structural and membrane integrity of cellular organelles and keeping them fully functional under stress. The accumulation of optimal amounts of H2O2 triggers the occurrence of chitinase proteins that can produce calcium homeostasis, ion channels, phosphatases, transcription factors, and abscisic acid (ABA), signaling responses to stress.43
APX in ascorbate–glutathione (AsA–GSH) cycling enzymes is responsible for the decomposition of H2O2 produced by SOD in different cellular organelles. APX plays a key role in both drought stress response and recovery after drought.43,44 APX is an integral component of the (ASC–GSH) cycle. APX performs the same function in the cytosol and chloroplasts. APX reduces H2O2 to H2O and docosahexaenoic acid (DHA), using AsA as a reducing agent.
The APX family consists of five isoforms based on different sites of amino acid formation, such as the cytosol, mitochondria, peroxisomes, and chloroplastids (stroma and thylakoids).45 APX is widely distributed and has a better affinity to H2O2, especially in terms of more efficient uptake of H2O2 in times of stress.45–48
Though the SOD levels in each clone showed a significant effect due to water content, it was limited to a few observations because drought affects the metabolic activity of clones. Likewise for the levels of SOD at a given water content. Water content of 30% showed relatively the same SOD activity as 60% and 90% in all observations. This indicates that SOD were formed in low levels in the observations and therefore cannot be used as a marker of tolerance for these tested clones. SOD is one of the key components of cell protection against oxidative stress. The SOD has three different isoenzymes distributed between organelles. Cu/Zn-SOD is predominantly located in the chloroplasts, cytosol, and peroxisomes, whereas FeSOD and MnSOD are mostly found in chloroplasts and mitochondria, respectively.49 POD and SOD activities increased sharply in rubber seedlings after being subject to drought stress. This suggests that the photosynthetic activity and lipid integrity of the cell membranes are rapidly attenuated by drought stress. SODs are metalloenzymes that play an important role in ROS reactions, or, in other words, are able to neutralize the negative effects of ROS. The decrease in substrate binding affinity to SOD as well as a decrease in one isozyme band of SOD under drought conditions may be responsible for the resistance. Plants that have a higher induced SOD activity show more tolerance to abiotic stresses. Numerous studies have shown that plants are able to better eliminate the negative effects of ROS produced under stressful situations when their SOD activity is higher, provided there are more SOD isoenzymes present.
POD had low values in all six observations of some clones. The low POD indicated that the effect of some water content percentages given during the six observations on several different clones did not have a significant effect. This indicates that the POD characteristics cannot be used as a reference for plant tolerance to drought stress. Plants that produce more POD under conditions of drought stress will be able to survive by eliminating the effects of ROS. In general, the activity of POD and other antioxidant enzymes will automatically have a higher value in tolerant clones/varieties and will have a lower value in susceptible clones/varieties. This indicates that drought-tolerant clones/varieties will be more efficient in removing H2O2 to produce optimal protection. The tolerance of some genotypes to environmental stresses has been associated with higher antioxidant enzyme activity. Drought-tolerant species of pigeon pea (Cajanus cajan),50 wheat (Triticum aestivum),51,52 and black bean (Phaseolus mungo)49 have higher SOD, POD, and CAT activities than drought-sensitive species. The results of this study indicate that ROS enzymes, which play a crucial role in the drought-tolerance mechanism under the drought treatment, have been identified in clones IRR 425, IRR 428, IRR 429, IRR 434, IRR 440, RRIC 100, and BPM 24. Based on the findings, several analyses have been carried out on physiological characteristics to determine the effects of water content on a greenhouse scale.
Based on the physiological and biochemical parameters made in this investigation to determine the effect of water content in a greenhouse scale, the findings indicate that only few parameters are reliable as indicators of drought stress tolerance. For e.g. the parameters such as total sugar content and proline were exhibiting strong correlation with drought stress tolerance in the IRR 400 series. They were produced at higher levels in plants treated with lesser water content. However, the parameters such as chlorophyll a, b, and total, H2O2, APX SOD, and POD did not show any correlation to use as drought tolerance indicators in this experiment.
This project contains the following underlying data:
Figshare: Data set: Physiological Characters of IRR 400 Series Rubber Clones (HeveabrasiliensisMuell. Arg.) on Drought Stress. https://doi.org/10.6084/m9.figshare.21708230.v252
• Biochemistry characters.xlsx (datasets for total sugar, chlorophyll (a, b, total), proline, H 2O 2, APX, SOD, POD)
Figshare: Data set: Physiological Characters of IRR 400 Series Rubber Clones (HeveabrasiliensisMuell. Arg.) on Drought Stress. https://doi.org/10.6084/m9.figshare.21708275.v253
Figshare: Data set: Physiological Characters of IRR 400 Series Rubber Clones (HeveabrasiliensisMuell. Arg.) on Drought Stress. https://doi.org/10.6084/m9.figshare.21645116.v554
This project contains the following extended data:
Data are available under the terms of the Creative Commons Attribution 4.0 International license (CC-BY 4.0).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)