Keywords
Nanoplastic, toxicity, nervous system
Nanoplastic, toxicity, nervous system
Plastics (also known as synthetic polymers) have revolutionized modern everyday life, making the world without plastics almost unrecognizable. Plastics are chains of organic subunits (“monomers”) linked through strong covalent chemical bonds. In 1907, Belgian chemist Leo Baekeland developed the first ever true synthetic polymer, Bakelite, marking the commencement of the “Age of Plastics.” Mass plastic production began several years (in the 1940s and 1950s) after the discovery of Bakelite. At that time, plastic utilization became a heated discussion among industries. A book entitled ‘Plastics’ written by Yarsley & Couzens (1941/45) discussed the indispensable benefits of plastics in a wide variety of sectors.1 Since then, the development of plastics has gradually expanded, with at least 15 new classes of polymers being synthesized.2
To understand the magnitude of the benefits that plastics have introduced in the modern world, it is essential to comprehend the definition of plastics. Generally, plastics polymerize monomers derived from fossil oil, natural gas, or coal. Virgin plastic polymers are extremely rare, and most are synthesized with various chemical additives to enhance their potential. For instance, the addition of carbon and silica reinforces the polymer material.3 Occasionally, various antioxidants and stabilizers are added to plastics to increase their half-life, thereby slowing the process of environmental degradation. Plastics with different compositions and properties can be manufactured using these enhancers. Currently, plastics are grouped into 20 classes based on their grade, variety, and composition.4
The manufacture of plastics requires extreme precision because the materials to be engineered require high accuracy and specific physical properties. Plastics are developed under certain heat and pressure conditions, and their material properties are adjusted during and/or after manufacturing to attain desired characteristics such as strength and permeability. Furthermore, plastics can be molded into limitless shapes and sizes via rotation, extrusion, compression, blowing, or injection, thereby allowing the development of a vast array of products.5 Although extra effort is needed for their production, plastics remain inexpensive. Thus, they have gradually displaced materials such as rubber, wood, and metal in a wide variety of sectors owing to their unique and versatile properties: electrical, heat, chemical, and light resistance; strong and durable but meltable at specific temperatures; and cost-effective.2
With the advent of the plastic revolution, these versatile polymers have been mass-produced to accommodate rapid economic growth and urbanization. Statistics reveal that global plastic production has skyrocketed over the past 70 years, from 1.5 million metric tons (Mt) in the 1950s to approximately 367 million Mt in 2020.6
Although it is undeniable that plastics are important in the modern world, the increasing usage of these persistent and ubiquitous materials has led to an emerging threat to the environment. Similar to all consumer materials, plastic waste contributes to an increase in municipal solid waste and mostly to urban litter. This is because most plastics generated today are made from nonrenewable and nonbiodegradable petrochemicals, translating to persistent and prolonged plastic litter endurance.2
In addition, proper plastic waste management (PWM) failed to achieve public acclaim. A study conducted revealed that the practice of PWM from 1980 to 2015 included discarding (55%), recycling (20%), and incineration (25%).7 In the early days, most plastics were discarded through landfill, as the waste volume remained stable without generating significant levels of harmful leachates or gases. However, this method does not promote plastic waste deterioration, as even biodegradable waste, such as paper, requires an extended period to decay because of the lack of moisture and oxygen needed for the biotic process. The decreasing capacity of landfills and persistence of plastics in the environment could pose an issue in the future. Furthermore, a vast majority of commodity plastics are not readily biodegradable. In this context, the lifetime of plastics is highly variable, as degradation depends on their chemical composition and the condition of their immediate environment.8
Even with the development of new PWM methods, the responsibility for the recovery efforts still lies with the public. Inevitably, littering is a critical social issue that requires attention and resources to promote awareness of its grave consequences for the environment. Mismanaged plastic wastes accumulate and contaminate a wide range of terrestrial environments, including freshwater, seashores, the open ocean, and the seafloor.9 A study by Topçu et al. and Thiel et al.10,11 revealed that more than 80% of litter stranded on beaches are plastic-containing materials such as single-use bags and food containers, which are responsible for the massive quantities of plastic debris (~79%) in the environment.12
Despite being persistent recalcitrant materials, plastics ultimately fragment at an extremely slow rate. The term ‘degradation’ refers to the breakdown of polymer chains owing to alterations in their material properties. Plastics can be degraded either biologically (by microbial biodegradation) or environmentally (through wind, rain, and sunlight exposure). Commonly, sunlight is particularly effective in degrading plastic waste owing to solar ultraviolet (UV)-induced photodegradation reactions.13 Tiny plastic fragments are termed according to their sizes: secondary microplastics (MPs) and nanoplastics (NPs), the latter of which is the primary focus of this review.
NPs are one of the very persistent contaminants, particularly in marine ecosystems, and are essentially different from bulk-form polymer types.14 The concern about NPs as an emergent contaminant triggered burgeoning research on its sources, effects, and potential hazards, especially to human health. However, this evidence is starkly deficient. First, the definition of NPs has been intensely debated. NPs have been categorized as plastic particles of either < 20 μm,15 < 1 μm in at least one of their dimensions,16 or < 100 nm in size.17 This review adopts a broader approach to NP size by Hartmann et al.18 (1–1000 nm-sized particles).
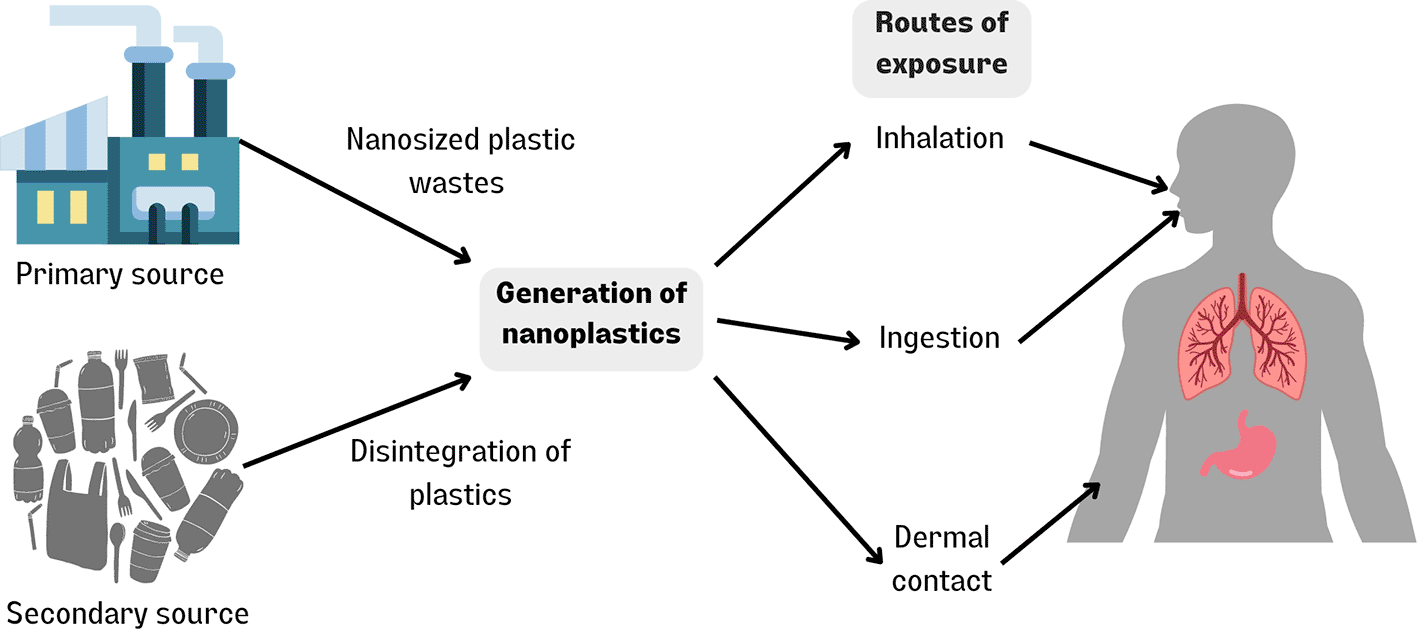
NPs can be derived from primary or secondary sources. Primary NPs are created by the deliberate production of polymer materials for specific purposes, such as in consumer products such as cosmetics, medicine, and raw materials.19–21 Secondary NPs are commonly mismanaged bulk plastic waste that are fragmented in the environment.
Convincing evidence suggests that the fate of NPs is often associated with the marine environment. Although information on how NPs are measured in aquatic systems is limited, it has been demonstrated that marine environments constitute the final recipients of plastic particles and NPs. For instance, plastics from land-based sources leak into the sea through freshwater and buoyed by the wind, rendering NPs mostly in aquatic environments.22 Thousands of particles/m2 have been detected in marine environments, further supporting this notion.23
In the marine environment, there are three main sources of NPs: (1) Primary NPs (polymer NPs that are produced deliberately for specific applications such as drug delivery, biomedical products, electronics),22 (2) Secondary NPs (degradation of bulk plastic wastes in the environment via mechanical action, UV photodegradation, hydrolysis, or microbial biodegradation),13 and (3) inadequate disposal infrastructure at Wastewater Treatment Plants (WWTP).
The first source (primary NPs) originates from nanoparticle-sized plastics produced for specific applications. These include biodegradable nanoparticles, nanospheres, and nanocapsules used in the medical field,24 and plastic particles of approximately 4 μm in size that are incorporated into cosmetic products such as exfoliating scrubs and skin cleaners.25
The second presumed source is the degradation of macroplastics in the environment. As previously mentioned, the breakdown process occurs because of biotic and/or abiotic processes. Biotic processes include microbial biodegradation, whereas abiotic processes include mechanical effects (wave forces), wind, rain (hydrolysis of polymers), UV radiation, and photooxidation.13 Such processes contribute either solely or jointly to decreasing the size of particles, from macro, micro, and ultimately to nano dimensions.26 The degradation of plastics was successfully demonstrated under the experimental conditions. For example, research by Zhang et al. on the thermal cutting process of polystyrene foam was able to generate nanosized polymer particles with a range between 22 and 220 nm.27 Lambert and Wagner et al. demonstrated that polystyrene coffee cup lids disintegrated into particles of 1.26 × 108 particles/mL with an average size of 224 nm in only 56 days. Overall, it can be concluded that the degradation depends greatly on the size of the initial plastics to achieve nanoscale dimensions.
Plastic fragmentation occurs because of a drastic reduction in the average polymer molecular weight. Polymers consist of a mixture of chains with variable lengths chemically linked by weak bonds (i.e., hydrogen or van der Waals) or physical interactions between chains. The weak interactions contribute to the brittleness of the bond at low-energy levels. In combination with external forces such as friction, plastics can be fragmented easily – a postulation reinforced by a study done by Shim et al., where expanded polystyrene beads disintegrated into micro- and nanosized particles through glass beads and sand mechanical abrasion in a month.28 This experiment mimicked the conditions at seashores or riverbanks, where abrasion by sand particles against plastic litter for a prolonged duration can eventually lead to the formation of NPs. In addition, the combination of UV radiation, elevated temperature, and humidity accelerated the rate of fragmentation. However, the association between physical abrasion and NP formation remains unclear.
Photodegradation is a widely accepted cause of NP production is photodegradation.29 This is because exposure to oxygen and sunlight initiates polymer fragmentation. Therefore, it can be deduced that the rate of degradation increased with physical abrasion, photodegradation, and the combined effects of both. There are two types of nano-fragmentation: (1) direct fragmentation of macro-or microplastics and (2) further gradual size reduction through degradation.22 The formation of NPs in oceans is based on the second type of nano-fragmentation and is estimated to be slower due to the lack of factors such as oxygen and light.
The third source of NPs is WWTP. Biosolid and effluent waters containing plastic particles from WWTP may leak into the marine environment without proper disposal methods. Talvitie et al. conducted a comparative study on the concentration of microplastics in wastewater from a WWTP located in the Baltic Sea and in seawater samples. It was shown that the particles per litre were 25 times higher than those of seawater samples.30 Another example of the presence of NP particles in synthetic fibers showed that > 1900 fibers per item could leak into sewage during each washing of clothes.31 Therefore, plastic particles entering the marine environment without proper disposal are a grave concern.
In most studies, detection and quantification methods for MPs in various environmental samples have been successful. However, analytical methods for detecting NPs in the environment have not yet been established. There are 33 studies on developing established protocols for pre-treatment (digestion and preconcentration), isolating, identifying, and quantifying NPs.18 Unfortunately, most were only able to detect the presence of NPs under laboratory conditions but failed to isolate and quantify environmental NPs from original field samples.
UV-VIS spectrometry, electron microscopy, field flow fractionation (FFF), and dynamic light scattering (DLS) are some techniques used in nanoparticle assessment.32 Presently, it is presumptuous to think that these methods are appropriate for NPs, as each technique has limitations. Shim et al. successfully identified the presence of NPs under mechanical abrasion against plastics using scanning electron microscopy and energy-dispersive X-ray spectroscopy (SEM-EDS).28
Other analytical methods such as Coulter Counter and nanoparticle tracking analysis (NTA) characterize particles in environmental samples.33 Coulter Counter is a conductivity-based method that measures particles of 0.4 – 1200 μm in diameter using an aperture. This method is highly dependent on the size of the aperture used.34 In this technique, the sample is placed in a weak electrolyte solution to be diluted, and then allowed to pass through the aperture, which is concurrent with an electrical current. As the particles in the sample passed through the aperture, they displaced their own electrolyte and changed the impedance. Alterations in the impedance generate a pulse that is directly proportional to the volume of particles displaced and converted to a signal. This signal was then passed to an instrument to assess and measure the volume and size distribution of the particles.35,36 NTA allows the visualization, measurement, and characterization of particles ranging in size from 30 to 2000 nm.37 This technique uses a laser beam to illuminate a single particle in the sample of interest; the particle size is proportional to the rate of Brownian motion.38 NTA has been previously used in studies by Lamber and Wagner for the degradation of polystyrene.39
In general, published studies have described a series of procedures before the identification and quantification of NPs in real field samples. These include sampling, pre-treatment, imaging (for identification), and quantification (of its concentration and size distribution). Sampling involves the collection of a portion of real field samples, whereas pre-treatment is a process of retrieving nanoparticles from environmental samples such as freshwater, wastewater, sediments, soils, and food.40,41
As NPs are highly heterogeneous and can undergo aggregation with organic and inorganic materials, pretreatment is important to separate impurities while retaining the amount and essential properties of the NPs.42 Three types of pretreatments, which are performed solely or jointly, include digestion of the matrix, preconcentration, and separation. The identification of NPs involves chemically identifying whether the nanosized particles are plastic. The methods used are spectroscopy (e.g., SEM-Raman spectroscopy, X-ray photoelectron spectroscopy (XPS),43–46 mass spectrometry (e.g., pyrolysis-gas chromatography-mass spectrometry (Py-GC-MS),47–50 thermal desorption-proton transfer reaction-mass spectrometry (TD-PTR-MS),51 and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS).52
Quantification of NPs is the final procedure of the analytical methods used to detect NPs in environmental samples. Usually, quantitative data, such as particle size distribution, particle concentration, and mass concentration, are considered for the assessment of the potential risks of NPs to the environment. However, the quantification of NPs has yet to be established and discovered. Some of the methods used include SEM, TEM, DLS, and NTA.18
To date, only five studies have reported the detection of environmental NPs in field samples. One study isolated, quantified, and identified nanosized plastic particles in seawater samples from the North Atlantic subtropical gyre using ultrafiltration, DLS, pyrolysis, thermal desorption, and thermochemolysis coupled with GC-MS.53 Another study used membrane filtration and TD-PTR-MS to analyze polyethylene terephthalate (PET) in alpine snow samples. The other two studies investigated NPs in sand and agricultural soil samples, respectively, using Py-GC-MS.49,50
Despite the findings discussed here, analytical methods for NPs in field samples remain scarce, suggesting that the occurrence of NPs may be speculative.54 Nevertheless, recent literature has highlighted the presence of nanometric particles produced by degradation. It is assumed that NPs are present particularly on the ocean surface, with intense exposure to sunlight and abundant oxygen. The distributions of NPs depend on their buoyancies; for instance, NPs with higher densities sink and accumulate in sediments. Less dense NPs remain on the surface of the sea and face biofouling, entrain with biological aggregates, or degrade further.55
The understanding of NPs and their interaction with the environment is vague owing to methodological challenges. Currently, NPs in environmental samples remain largely unquantified. Although NPs have recently evolved to a heated topic in research communities, many characteristics of NPs have been assumed through the data obtained from MPs owing to their similar composition and sources. Some may even assume that the interactions of NPs are analogous to those of MPs because of their similar constituents and sources. However, both plastics are distinguishable based on their size distribution, transport properties, bioavailability, interactions with environmental materials, and potential toxicity towards organisms.
Hence, it is essential to define ‘MPs’ and ‘NPs.’ Although studies on NPs have been inconsistent with their exact size threshold compared to MPs, the US National Nanotechnology Initiative defines ‘nano’ as having at least one dimension in the range of 1 to 100 nm. MPs are plastic particles that are less than 5 mm in size. However, the more obvious differences lie in their distinct characteristics. In terms of sedimentation and buoyancy, a study found that NPs have a higher dominance of Brownian motion in suspensions. This is because the NPs are more likely to be homogeneously dispersed. Furthermore, the ability of NPs to heteroaggregate or adsorb with other natural colloids (e.g., natural organic matter (NOM)) is higher because of their small sizes. However, this largely depends on environmental conditions such as porewater pH, ionic strength, and hardness. As NPs heteroaggregate with natural colloids, they affect the environmental fate, as NPs are smaller than most microbes. Heteroaggregates formed with NPs are relatively difficult to discriminate from their plastic components owing to microbial interactions. Additionally, most plastics are chemically modified and NPs have short scales in diffusive transport, which accelerates the release of these additives.
In terms of biological consequences, the minute size of NPs increases their bioavailability and accessibility in biological organisms through biouptake, translocation, and transportation across membranes. As NPs are approximately the same size as natural proteins, they can pass through biological membranes through passive diffusion or other pathways, such as endocytosis. Thus, increased bioavailability may indicate higher toxicity. Rist et al. conducted a toxicity study of MPs and NPs in Daphnia magna by assessing the feeding rates and reproduction of crustaceans. The results revealed that nano-polystyrene particles caused significantly decreased feeding rates in plants compared to MPs, and also deduced that nanoparticles produce reactive oxygen species after interacting with subcellular components.56 NPs also have a higher surface reactivity than MPs owing to their exposure to specific molecules. This results in a series of consequences for the surface chemistry of biological systems.
In conclusion, both nano- and microplastics are hazardous to most organisms, including humans, owing to their high specific surface area and hydrophobic nature, which allows them to adsorb easily on certain surfaces.57 Furthermore, their miniature size allows them to act as vectors for both chemical pollutants and pathogens. A study by Wu et al. proved that MPs provide a microhabitat that supports the growth of certain bacteria, thereby increasing opportunistic human pathogen resistance to antibiotics.58 Alarmingly, plastic particles with sizes of MPs or smaller can translocate across biological membranes through endocytic-like mechanisms, which results in cell and/or tissue internalization and adversely affects health. In contrast, plastic particles larger than 100 μm are generally non-toxic.
As studies on NPs and their toxicity towards human health are limited, the data suggest that NPs are more hazardous to human health than MPs. Jeong et al. and Rist et al. provided factual evidence that smaller MP show higher bioavailability and retention time in the body compartments, which implies higher toxicity levels.59,60 Furthermore, NPs show a higher potential for bioaccumulation in organisms and can pass through biological barriers. In addition, NPs have a higher surface-to-volume ratio and aggregation potential than MPs,61 which allows them to interfere with most biological and chemical interactions in the body.62
As NPs have only recently commanded the spotlight, it is not surprising that information on the potential effects of NPs is still scarce. In contrast to NPs, the routes for MPs entering the human body have been relatively well studied and thus form the basis for work on NP transport and bioaccumulation. In this regard, NPs can enter the human body through ingestion, inhalation, and skin contact in three ways.
Ingestion. The primary route of human exposure to NPs is through ingestion of contaminated food or drinks. In this context, the entry of NPs into food and beverages may be due to the migration of NPs from packaging materials or via the food chain. The initial analysis to verify ingestion as the route of NP internalization was by studying the presence of plastic particles excreted in human stool samples. The study also reaffirmed that humans experience continuous exposure to NPs through the uptake of food and water.63
The most well-defined route is drinking water, especially in countries where water is consumed directly from pipes. This is because water networks are connected from household to household through plastic-based plumbing. As expected, NPs are produced through long-term, high levels of fluid force on the piping walls, which eventually wind up in consumers’ gastrointestinal (GI) tracts.64 Additionally, packaging can be used as a source of plastics. A study reported that billions of NPs are ingested by consumers from a single cup of tea per plastic teabag.65
Common food products, such as sugar, salt, bottled water, and aquatic organisms, such as bivalves, fish, and crustaceans, are known to be impregnated with MPs and NPs. However, studies on MP absorption through the diet are more prevalent than those on NPs. For instance, a study on table salt consumption per person (which is higher in China66 than in Europe67) measured MPs that were inadvertently mixed with table salt and showed that 37 and 100 plastic nanoparticles were consumed per person in Europe and China, respectively.66,67
The presence of MPs and NPs in food, particularly seafood, was reported by the ESPA Panel on Contaminants in the Food Chain (CONTAM). In 2016, they quantified the amount of MPs that entered the human digestive system by extracting, detecting, quantifying, and characterizing plastics using Fourier-transform infrared spectroscopy or Raman spectroscopy. The average number of MPs found in fish is 1-7 particles/g and bivalves is 0.2-4 particles/g, while data on NPs are limited due to technical challenges in analytical methods.
Another study revealed that MPs and NPs can be ingested by Pacific oyster larvae,68 further supporting the presence of plastic particles in food. The entry of MPs and NPs through this pathway causes pathological changes in the digestive tract, including gut barrier dysfunction, intestinal inflammation, and dysbiosis of gut microbiota.69–72
The transfer of plastic particles between trophic levels may play a significant role in the presence of small plastic particles in food. Evidently, the human consumption of aquatic organisms can be linked to the food chain, whereby bioaccumulation and subsequent transfer of NPs through trophic levels occur.68,73 Other routes include deposition of NPs in the gills or skin of aquatic organisms.74
There are three assumptions regarding the fate of NPs after entering the GI tract: whether they remain in the gut lumen, translocate across the gut epithelia into systemic circulation, or are excreted from the body. In comparison to MPs, NPs were able to pass through the pores at paracellular tight junction channels with a maximum size of 1.5 nm.67 Other routes for NPs to enter systemic circulation are through the lymphatic system or phagocytosis by microfold (M) cells in Peyer’s patches.75
Two studies conducted on the absorption of polystyrene NPs in vivo and in vitro suggested that the oral bioavailability of polystyrene NPs was 10 times higher than that of MPs.76,77 However, the rate of NP absorption varies greatly, and there is no correlation between the size and chemical structure of NPs. The absorption rates of NPs in the GI tract are of great interest because NPs undergo transformation post-ingestion, which in turn affects their absorption rate and ability.
The heteroaggregation ability of NPs enables them to interact with organic molecules such as proteins, lipids, and carbohydrates.78 Eventually, the particle surface is bound by clusters of proteins, forming a coating known as the “corona”.79 This coating critically affects the biological identity of NPs, including their biodistribution and therapeutic and pathophysiological effects.80 The presence of a corona enhances potential NP toxicity in humans, as protein-coated NPs can interact more easily between tissues and organs than virgin NPs.81 Gopinath et al. revealed that protein-coated NPs led to higher cytotoxicity and genotoxicity in an in vitro study of human blood cells in comparison with bare NPs.82 This could be due to the formation of a biomolecular corona on the surface of NPs, which prevents them from being detected by immune cells and thus persists longer in the circulation.
Inhalation. Humans can also be exposed to NPs through unintended inhalation or occupational exposure.83 Airborne NPs originate from either outdoor (rubber tyres, fertilizers, and sea salt aerosols) or indoor environments (synthetic textiles and agriculture). A study revealed that NPs are present in atmospheric fallout.84 Dris et al. showed that most MPs from atmospheric fallout in both urban and suburban areas of Paris were derived from synthetic fiber particles. The atmospheric flux from urban areas is twice the average amount and exceeds that of suburban communities. A further study by Dris et al. indicated that MPs are higher in indoor samples (1-60 fibres/m3) than in outdoor samples (0.3-1.5 fibres/m3).85,86 However, information on the amount, source, and concentration of airborne NPs is limited.
The entry of NPs into an organism can lead to many health consequences, either through the toxicity of the particle itself and/or its chemical constituents, or by introducing pathogens and parasite vectors.87 Inhaled NPs are destined to be deposited in the lungs, which is a part of the complex network that comprises the respiratory system. Approximately 300 million alveoli facilitate gas exchange via diffusion, encompassing a surface area of approximately 150 m2 and a fine tissue barrier of less than 1 μm.88 Air in the alveoli is in proximity to the flowing blood, as the lumen and alveoli are only separated by an epithelial and endothelial layer.
Under normal circumstances, foreign particles deposited on the lungs are removed as they enter the respiratory tract or bronchial tract via mucus and cilia. However, fine particles (< 2.5 μm) such as NPs are transported into the alveoli along with air. Mucociliary transport mechanisms are absent in the alveoli; thus, trapped foreign particles are removed via macrophage engulfment and transported back to the bronchial regions for eradication. Some studies have shown that a small proportion of nanoparticles (<0.05%) can be transported into the bloodstream by overcoming the air-blood barrier. Although this circumstance is uncommon and is dependent on the physicochemical properties of the NPs, this suggests that uptake into the lungs is possible at high concentrations.
Furthermore, there are two fates for plastic particles that enter the respiratory system depending on their physical properties (size and density): deposition or penetration deeper into the lungs. NPs with lower densities and smaller sizes can be embedded deep into the lungs, deposited on the alveolar surface, or translocated to other body parts.89,90 The deposition of NPs on alveolar surfaces can trigger clearance through macrophage action and eventually translocate to the circulatory and lymphatic systems.91
Several studies have reported that NPs may induce inflammatory responses in the lung tissue. Xu and colleagues demonstrated that internalized polystyrene NPs (PS-NPs) interfere with gene expression in A549 cells, resulting in the induction of inflammation and apoptosis. Cell viability is dependent on the NP concentration and size.92 Another study by Shi et al. investigated the toxicity exerted by PS-NPs on the same cell line in terms of cell viability, oxidative stress, and inflammatory reactions.93 Taken together, the absorption of NPs via inhalation can result in lung damage. Several factors, including hydrophobicity, surface charge, functionalization, and size, affect the rate and ability of NP absorption in the lungs.94 Absorption rates of NPs through the lungs were found to correlate with occupational exposure and comorbidity in in vivo models, such as pulmonary inflammation and cancer.95
Generally, respirable NPs may trigger a series of reactions such as oxidative stress, inflammation, and genotoxicity in the respiratory, cardiovascular, and nervous systems. However, the available literature on these mechanisms is inconsistent because of variations in the NP sizes used. Because NPs have a large specific surface area, they contribute to the formation of free oxygen radicals or reactive oxygen species (ROS), such as superoxide anions and hydroxyl radicals. Large quantities of ROS generated in cells create havoc on cellular components such as lipids, proteins, and DNA, causing long-term cell damage in the long run. ROS are formed through one of several mechanisms: (1) direct formation on the surfaces of the NPs, (2) triggered by NPs through mitochondrial damage and disruption of balance in the respiratory chain, or (3) triggered by macrophages and neutrophils activated by foreign particles. These radicals trigger inflammation if they are not eradicated immediately via binding to endogenous antioxidants (e.g., Vitamin C) or degraded by antioxidative enzymes.
Dermal contact. Another route of NP exposure is through dermal contact. The human body is protected from injuries, chemicals, and microbial agents by the stratum corneum, which is the outermost layer of skin. The stratum corneum consists of hydrophilic lipids, such as ceramide, long-chain free fatty acids, and cholesterol.96 Owing to these properties, the entry of hydrophobic NPs into the body seemed improbable; however, the skin barrier only restricted the entrance of particles with sizes above 100 nm.
The dermal route comprises contact via cosmetic and healthcare products or NP-contaminated water. The introduction of NPs through direct contact with the skin is uncommon, but they may invade the body through sweat glands, skin wounds, or hair follicles.97 Alvarez-Roman et al. studied the entry and distribution of plastic particles throughout skin tissue, whereby fluorescent polystyrene particles with diameters of 20 and 200 nm were placed on skin tissues from a pig. They revealed that a higher amount of 20 nm polystyrene NPs was concentrated in the hair follicles in comparison with the 200 nm polystyrene NPs using a confocal laser scanning microscope. However, neither NPs penetrated the stratum corneum. In contrast, Campbell et al. provided evidence that 20-200 nm NPs can penetrate a depth of 2-3 μm into the top layers of the skin, and Vogt et al. suggested that transcutaneous NPs were absorbed by Langerhans cells.98–100 One facilitating factor for NPs to enter the skin is UV radiation, as UV rays can damage and weaken the skin barrier.101
The entry of NPs into keratinocytes may induce cytotoxic reactions, such as oxidative stress, trigger inflammatory factors, or even result in cell death, either by apoptosis or necrosis. A previous study suggested that polystyrene particles can interact with biological membranes, allowing the penetration of lipid bilayer membranes, alteration of cell membrane structure, and ultimately impeding normal cellular function.102
Figure 1 depicts the sources of nanoplastic and their routes of exposure in human.
Once NPs enter the human body, they enter the circulatory system and are then distributed throughout the body. The concentration and interaction of NPs with their target molecules (e.g., proteins, phospholipids, and carbohydrates) depend greatly on the size, surface chemistry, and charge of the target elements. Furthermore, the high free surface energy of NPs allows their adsorption onto biomolecules when they cross complex biological fluids. The interaction between NPs and biomolecules leads to the formation of ‘biomolecular coronas,’ where NPs are encompassed by a collection of biomolecules. The formation of biomolecular coronas depends on the physicochemical properties of the NPs and their exposure time to biological components in the environment.80 These biomolecular coronas modify the biological traits of NPs, such as the enhanced rate of translocation, by altering the form of NPs to adapt to the surrounding biological environment. This is because the corona can lower the NP surface energy, thereby promoting dispersion. In some circumstances, it also promotes the interaction between NPs and cell surfaces, thereby increasing the toxicity potential of NPs. In contrast, some NPs lose their targeting abilities owing to corona formation.103
There are two types of coronas: ‘hard’ and ‘soft.’ ‘Hard’ coronas are a somewhat reversible but tightly bound layer of biomolecules formed through adsorption of biomolecules on bare nanoparticle surfaces immediately after contact. On the other hand, ‘soft’ corona consists of a more loosely bound interaction of biomolecules on the surfaces of NPs, and they are readily exchangeable if exposed with other nanoparticles with higher energy surfaces in the environment. Unlike ‘hard’ corona, any subsequent encounter of the nanoparticles and different biomolecules in a pristine environment may result only in partial displacement of the original ‘soft corona by the new biomolecules. Biomolecules that are not displaced would serve as a memory of the corona to the previous environment to which the nanoparticles are exposed. In general, the composition of the corona depends on the environment through which the nanoparticles pass. Furthermore, NPs can travel across every component within organisms because of their small sizes and potentially retain biomolecules in their corona from every environment they encounter. If the nanoparticles are retained in the corona for a long duration, the nanoparticle-corona complex will be recognized by the cell membrane receptor.80 The consequences of corona formation include the biodistribution of NPs and their potential pathophysiological effects within organisms.
Cellular uptake of NPs. There are several routes by which NPs can be taken up by the cells. The main route of NPs uptake is through endocytic pathways, where adhesive interactions between NPs and channel or transport proteins occur. These pathways include phagocytosis, micropinocytosis, clathrin-mediated endocytosis, and caveolae-mediated endocytosis pathways. Phagocytosis is a process in which particles (> 0.5 μm) are ingested by cells. After ingestion, the particles are enclosed in membrane-bound vesicles, also known as phagosomes, to allow them to be targeted by lysosomes for enzymatic degradation. The process is initiated when the particles bind to specialized receptors on cell membranes. After binding, polymerization occurs at the site of ingestion, leading to the formation of pseudopodal extensions, which are extensive deformations of the plasma membrane. Pseudopodia encompass particles in a cup-shaped structure. As the particles are completely internalized, depolymerization of actin filaments begins at the base of the phagocytic cup, allowing them to seal up into phagosomes. As phagosomes mature, they undergo a series of membrane fusion and fission events with endosomal compartments, eventually binding with lysosomes and forming microbicidal phagolysosomes.104
Macropinocytosis is an endocytic pathway involving nonspecific uptake of extracellular materials, including soluble molecules. The process involves inward folding of cells due to the fusion of the cell surface with the basal membrane to form vesicular structures and macropinosomes. The initiation of this actin-dependent process depends on the stimulation of growth factors such as colony-stimulating factor 1 (CSF-1), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF). Thereby, the polymerization of actin filaments increases at the cell membrane, allowing membrane ruffling to occur. Some lamellipodia-induced ruffles continue to fold inward and fuse with the basal membrane, forming macropinosomes. The maturation process of macropinosomes is cell type-dependent. For instance, the maturation process in macrophages occurs by shrinking macropinosomes, which eventually fuses with lysosomes for enzymatic degradation.105
Clathrin-mediated endocytosis is a type of vesicular transport that involves the use of clathrin, a triskelion-shaped scaffold protein. In this process, clathrin assembles and polymerizes to form a coat around the cytoplasmic region of the invaginated membrane. The coat serves as a reinforced mold and can dissociate rapidly after the vesicle splits from the membrane. However, this process can only occur in the presence of phosphatidylinositol-4,5-biphosphate (PIP2) and adaptor proteins. The growth of clathrin-coated pits requires the BAR (Bin/amphiphysin/Rvs) domain protein and reorganization of the actin network.106
Caveolar-mediated endocytosis is clathrin-independent endocytosis, where it forms a bulb-shaped plasma membrane invagination called caveolae. The formation of caveolae depends on the presence of the integral membrane protein caveolins and peripheral membrane protein caving. During this process, caveolae bud off from the plasma membrane and form endosomes.107 It was shown that 44 nm polystyrene particles were mainly absorbed through clathrin-independent endocytosis into human colon fibroblasts and bovine oviductal epithelial cells.108 Dos Santos et al. studied the cellular absorption of 40 and 200 nm carboxylated polystyrene NPs in several tumor cell lines and deduced that NPs were absorbed into cells via active transport. This suggests that the uptake of NPs differs depending on the cell type.109 Still, the absorption of NPs into human cells is greatly dependent on their size and surface chemistry. Furthermore, another study demonstrated the absorption of NPs into J774A.1 macrophage cells through micropinocytosis, phagocytosis, and clathrin-mediated pathways, whereas in A549 cell lines, NPs enter via caveolae- and clathrin-mediated endocytic pathways.
Previous studies have shown that the interaction between plastic particles and human cells is influenced by their high specific surface areas, charges, structures, and types of targeted cells.110
Recent laboratory and field studies on aquatic organisms and other commercial species have suggested that NPs are mainly absorbed in the GI tract and are distributed to other parts of the body. Once NPs enter the blood circulation, their nanosize facilitates translocation across biological barriers, and ultimately across the blood-brain barrier (BBB). Ding et al. revealed that MPs could enter the brain of Oreochromis niloticus through the circulatory system.111 Geiser et al. further supported the hypothesis that NPs are present in the brain of fish through the ingestion of water or food across the BBB in an in vitro study.112 Mattsson et al. showed that both 53 and 180 nm sized polystyrene plastic particles can enter the brain of crucian carp (Carassius carassius).113 Studies have shown that nanoparticles can reach the brain directly via nerve fibers in the olfactory epithelium (nervus olfactorius) located at the roof of the nose. This method has been demonstrated using transsynaptic transport. Although this method allows nanoparticles to bypass the blood-brain barrier, its occurrence is extremely rare.114
As it has been proven that NPs can cross the BBB, it is essential to evaluate the toxic effects of NPs on brain tissue. Several studies have proposed that NPs can accumulate in these tissues, exerting multiple adverse effects on the nervous system. In this context, the toxic effects of NPs on the brain depend on the exposure duration, size, and type of NPs, as well as indirect mechanisms, such as oxidative stress.115 Wan et al.116 revealed that neurotoxicity caused by exposure to polystyrene MPs in zebrafish larvae may be associated with some metabolic changes. Ding et al. demonstrated decreased levels of amino acids phenylalanine and tyrosine in adult tilapia after exposure to polystyrene NPs. These amino acids play a significant role in neurological function. Ding et al. concluded that NPs can interact with these amino acids, which eventually alters their metabolic pathways, and thus alters the formation of various neurotransmitters.115
Furthermore, NPs can induce oxidative stress-related damage in the brain. It has been reported that 1 to100 μm sized particles exert more severe stress than 0.3 μm polystyrene MPs, which indicates that larger particles, though in the micro range, can be more harmful. This may be explained by the hypothesis that medium-sized MPs can travel more freely across cells while inducing mechanical injuries and alterations in biochemical pathways, including oxidative damage and inflammatory responses.115 The brain is extremely sensitive to oxidative stress due to several factors. Oxidative stress is a biological condition in which redox homeostasis, which is an equilibrium between oxidant and antioxidant levels maintained in vivo, fails to occur. The imbalance in this equilibrium between free radical production and cellular antioxidants can be due to several disorders and/or diseases, such as carcinogenesis, inflammation, and aging. Chen et al. developed methods to evaluate the role of oxidative stress in neurotoxicity via specific biomarkers of oxidative stress, such as catalase (CAT), glutathione, peroxidase activity, and reduced glutathione.117,118
First, the brain is one of the most metabolically active organs and consumes an approximate amount of 20-30% of inspired oxygen, despite it only accounting for 2% of the body weight (approximately 1300 g).119 The need for high oxygen levels in the brain is because of the substantial amount of adenosine triphosphate (ATP) required in ion channels for activities such as action potentials and neurosecretion to maintain neuronal intracellular ion homeostasis.120
Second, most areas of the brain (e.g., substantia nigra, caudate nucleus, putamen, and globus pallidus) are rich in iron. Iron plays a key role in oxygen transport. For instance, the redox cycle between ferrous and ferric ions is essential for various electron transfer reactions, which are important for several cellular processes, including neurotransmitter synthesis, myelination of neurons, and mitochondrial function.121 Hence, defective iron homeostasis can result in the production of neurotoxic reactive species (e.g., such as reactive oxygen species (ROS)), eventually leading to oxidative damage and cell death in the central nervous system. Because ROS are highly unstable, they can easily react with a majority of macromolecules, such as proteins, nucleic acids, and lipids.121 Moreover, this reaction is further exacerbated, especially in the brain, because neuronal cells are unable to neutralize free radicals.119 Several studies have demonstrated that NPs can cause ROS accumulation.
Additionally, neuronal membranes are rich in polyunsaturated fatty acids (PUFAs) such as arachidonic acid (AA), docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA).120 PUFAs are especially vulnerable to oxidative stress because of their unsaturated double bonds. Recent discoveries have revealed that ROS can cause lipid peroxidation (LPO), a process that involves the disruption of oxidants on lipids that consist of C=C double bonds, such as PUFAs. In this context, ROS attack cellular membrane PUFAs and cause functional and/or structural impairments. Consequently, a group of highly reactive α, β-unsaturated aldehydes (e.g., 4-hydroxy-2-noneal (HNE), malondialdehyde (MDA), and acrolein) were generated.121 These highly reactive aldehydes are highly diffusive, allowing them to attack or generate covalent linkages with other cellular constituents, eventually leading to extensive cellular damage.
There are three types of mechanisms that result in LPO production-free radical-mediated oxidation: free radical-independent, nonenzymatic oxidation, and enzymatic oxidation.120 In the case of NP exposure, it is presumed that LPO occurs following free-radical-mediated oxidation. This mechanism involves a cascade of processes in the oxidative deterioration of lipids, such as initiation, propagation, and termination.121 In the last step, termination of propagation steps in LPO can only be performed with the complete utilization of the substrates or through interaction with antioxidants such as vitamin E and C.121 Otherwise, these cyclized fatty acids will only proliferate further while damaging the components of the cellular membrane, such as lipids and proteins, and further generate by-products of LPO.
In general, LPO affects several functions of the central nervous system (CNS) through membrane rigidity, decreased enzymatic activity in the membrane, damage to membrane receptors, and altered permeability of biological membranes.121 Consequently, these reactions lead to altered cellular function and severe cytotoxicity, resulting in uncontrolled cellular growth or apoptosis. In the CNS, PUFAs disrupt neuronal homeostasis, eventually causing brain dysfunction. After NP exposure, Barboza et al.122 reported higher levels of LPO and acetylcholinesterase (AChE) activity in the brains of Dicentrachus labrax, Trachurus, and Scomber colias. The increased level of AChE activity can be explained by the higher levels of LPO damaging acetylcholine-containing vesicles, thus causing a rise in neurotransmitter levels released in the synaptic clefts.
AChE is a cholinergic enzyme that is mainly produced at the postsynaptic neuromuscular junctions. AChE breaks down the neurotransmitter acetylcholine (ACh) into acetic acid and choline to ensure the termination of cholinergic neurotransmission in neuromuscular junctions and cholinergic brain synapses.123 In general, AChE is important for the regulation of brain function and is thus considered a biomarker for neurotoxicity. A study on zebrafish revealed that NP exposure can decrease AChE levels and inhibit neurotransmitters, such as dopamine, melatonin, aminobutyric acid, and serotonin in zebrafish.124 This is further supported by other studies on Medaka, O. latipes, and Pormatoschistus microps after exposure to MPs and NPs.
Inhibition of AChE activity can lead to several adverse effects, such as paralysis and, in severe cases, death.123 Furthermore, inhibition of AChE activity can be determined by developmental and motility alterations. For instance, the exposure of zebrafish to NPs suppresses locomotor ability, which leads to the deduction that decreased AChE activity is due to the binding of important channels, thus preventing AChE production. In addition, most studies have scrutinized the exposure of MPs and NPs to neurotoxicity through the evaluation of behavioral anomalies.125 Uncontrolled movements after exposure to NPs are inferred by the altered expression of essential neurotransmitters, such as ACh, glutamate, and γ-aminobutyric acid, or defective activation of neurons.
Recent discoveries have been made regarding the association between gene deregulation and exposure to NPs. For instance, the expression of glial fibrillary acidic protein (GFAP) and alpha-1 tubulin (α1-tubulin) in the CNS during the initial stages of zebrafish development is an important biomarker of neurotoxicity. In this context, α1-tubulin is essential for the formation of microtubules, whereas GFAP is important for astrocyte development. According to the US Environmental Protection Agency (EPA), upregulation of GFAP indicates neurotoxicity.124
To date, there have been a very limited number of studies on the neurotoxic potential of NPs in vitro. In vitro studies have largely focused on the extent of NP absorption and its implications on cells, such as proinflammatory effects and genotoxicity, based on particle size. Furthermore, cell models allow the study of NP-induced neurotoxic mechanisms, including oxidative stress, routes of NPs integrating into mitochondria, immune responses after exposure to NPs, interaction between NPs and enzymes, and alteration of receptor or channel function of neuronal cells. Generally, in vitro studies have revealed the many effects of NPs on fundamental cellular functions, including proliferation, differentiation, and apoptosis, to determine their neurotoxicity.126 However, the mechanism of neurotoxicity greatly depends on the surface area, composition, type, and shape of NPs, as well as the absorptivity of NPs into the body.
Human cell lines. A 3-dimensional (3D) in vitro model of a human embryonic stem cell (hESC)-derived (WA09) was used to test the potential developmental neurotoxicity exerted by NPs because of its ability to imitate conditions in vivo.126 The WA09 cells were differentiated into PAX6+ neural precursor cells. Methylmercury, a developmental neurotoxicant, was used to test the sensitivity of the 3D model in detecting developmental neurotoxicity (DNT). After exposure to methylmercury, a significant decrease in the expression of neuronal precursor and neuronal marker genes indicated that the 3D model reacted sensitively with methylmercury.
Polyethylene nanoplastics (PE-NPs) (30 nm, anisotropic nanocrystals, ζ potential -30mV) were used because of their ability to remain inert under physiological conditions, including retaining non-aggregation in the culture medium for a prolonged period (>3 months).126 This allows for the investigation of the biological effects of bio-persistent NPs upon chronic exposure. The acute and chronic toxicities exerted by the fluorophore-incorporated PE-NPs were recorded using flow cytometry. Acute toxicity was measured after 48 h in HeLa cells, with an EC50 value of 450 μg/ml, in which the ATP content decreased with increasing concentrations of PE-NPs.
In addition, LUHMES (human neuronal precursors) cells, a model closer to neural tissue, were used to incubate PE-NPs instead of cervical tissue-derived HeLa cells. Similarly, LUHMES cells were incubated for 48 h and an EC50 value of 257 μg/ml was determined. As the concentration of PE-NPs increased, the ATP content decreased. However, unlike in HeLa cells, the initial decrease in ATP content was constant but minimal. A significant decrease in ATP content was observed only when the PE-NPs reached a concentration of 90 μg/ml, which is likely indicative of the cellular oxidative stress exerted by PE-NPs. These findings strongly suggest that PE-NPs can initiate stress in mitochondria by reducing ATP content in a dose-dependent manner.126
Before chronic toxicity was measured, the incorporation of PE-NPs into neurospheres was also assessed. Fluorophore-labelled PE-NPs were incorporated into neurospheres at increasing concentrations and the results were recorded after 24 and 48 h. Unsurprisingly, PE-NPs were clearly recorded inside the neurospheres at a concentration of 360 μg/ml at the 48-hour mark. To confirm this, flow cytometric analysis of single-cell suspensions showed that PE-NPs could penetrate and incorporate into the neurospheres, as most of the cells (84 ± 6%) were positive at the highest concentration used (1440 μg/ml). An ATP concentration of 696 μg/ml (EC50) and decreasing ATP content with increasing PE-NP concentration were noted. Together with increased malondialdehyde concentrations, these findings may indicate increased oxidative stress levels in the cells and highlight the ability of PE-NPs to pass through the structural barrier into the cells without using altered forms.126
The chronic toxicity of PE-NPs was assessed after 18 days using cell viability assessment. EC50 values of 296 and 191 μg/ml were recorded using the resazurin reduction and ATP content as parameters, respectively. For both parameters, a similar trend of decreasing cell viability with increasing PE-NP concentration was observed. Another study on the differentiation and maturation process of neural precursor cells was conducted by exposing neurospheres to 22.5 μg/ml PE-NPs (a non-cytotoxic concentration). The results showed that PE-NPs interfered with the downstream expression of NOTCH pathway genes (HES5 and NOTCH1), NEUROD1, ASCL1, and FOXG1. PAX6 was not affected, suggesting that PE-NPs only affected later neuronal progenitors.126 In summary, this study revealed that PE-NPs caused severe impairment of the nervous system (DNT) due to dysregulated gene expression. Remarkably, all affected genes play a vital role in neurodevelopment; for instance, the NOTCH pathway ensures proper embryonic neural development and adult brain plasticity.127 Additionally, ASCL1 regulates neurogenesis by deleting defects in the ventral telencephalon,128 and FOXG1 is responsible for repressing specific transcription during early telencephalon development. PE-NPs can affect progenitor cells and neuronal development by affecting the genes responsible for neurogenesis. Furthermore, increasing concentrations of PE-NPs may cause cytotoxicity in neural cells, thereby manifesting as DNT.126
Another study used varying concentrations of NPs to determine their direct effect on cells in the nervous system. In this study, polystyrene NPs (50 nm, negatively discharged sulfonated surface, spherical) were added to human neuroblastoma cells SH-SY5Y (seeding density of 1.3×104 cells/cm2, cultured for 82 h), and the effects were compared to those of acrylamide, a potent neurotoxin (polystyrene NP concentration: 2, 10, and 50 μl/ml; acrylamide concentration: 0.8, 20, and 500 μg/ml). The viability of cells in polystyrene NP-containing medium at concentrations of 10 and 50 μl/ml was lower than that at 2 μl/ml. The morphology of cells in the polystyrene NP-containing medium differed from that in the acrylamide-containing medium with the appearance of decreasing processes, fuzzy cell bodies, and more granular substances (a few μm in size) surrounding the cells. Furthermore, the granular substances consisted of nucleic acid fragments, indicating necrosis, where the integrity of the organelle and plasma membrane had been breached, leading to the release of cytosolic and intracellular components into the surrounding environment. Thus, this study demonstrated the induction of cytotoxicity in SH-SY5Y cells when exposed to polystyrene NPs, whose toxicity might be on par with or exceed that of acrylamide. Morphological alterations, swelling of nuclei, release of organelle components, and shrinkage of neurite outgrowth deduced the neurotoxicity of NPs in vitro.129 This study also provides insight into the concentration of polystyrene NPs required to induce cytotoxicity. Based on these results, higher concentrations caused more apparent adverse effects on the neuronal cells. However, the dose or amount required to trigger cytotoxicity based on experimental models may differ from the threshold of the human brain. This is because NPs must overcome several obstacles, including the blood brain barrier (BBB), glial cells, and phagocytes of the central nervous system. Hence, the concentration of NPs used in this study may not reflect the manifestation in the actual human brain tissue.
Rodent cell lines. A study investigated neurotoxicity of NPs using different concentrations of polystyrene (PS) NPs (nanoscale-PS latex beads, 100 nm diameter, ζ potential -40.60 ± 3.96 mV) towards three different primary cell types from CD-1 (ICR) mice, including mouse embryonic fibroblasts (MEFs), mixed neuronal cells isolated from embryonic cortex, and cortical astrocytes.130 The experiment was conducted on the cell types by measuring two parameters, including cellular viability and level of the apoptosis marker cleaved caspase-3 (CC3). To test cellular viability, cells were incubated with different concentrations of PS-NPs (0, 50, 100, and 200 mg/L) for two days and the MTT assay was performed. The results revealed that MEFs and astrocytes were not affected, with no change in viability at concentrations of 200 and 100 mg/L, respectively. However, the viability of mixed neuronal cells significantly decreased at a concentration of 100 mg/L. Apoptosis markers, analyzed using immunofluorescence, revealed that CC3 increased upon exposure to PS-NPs at 100 mg/L, with higher levels in mixed neuronal cells after 2 days, but there was no change in CC3 levels in MEFs and astrocytes. However, the levels of lipocalin-2 (Lcn2) and proinflammatory cytokines, rarely detected under standard culture conditions, were elevated upon exposure to PS-NPs. This indicated the induction of reactive astrocytosis, but astrocytes did not undergo apoptosis.
The study also revealed that increasing concentrations of PS-NPs and plasma membrane permeability greatly enhanced the deposition and accumulation of PS-NPs, which in turn reduced cellular viability and increased apoptosis observed from the neuronal marker gene expression, Tubb3, and glial marker, GFAP. Tubb3 decreased, whereas GFAP increased at a concentration 100 mg/L of PS-NPs in mixed neuronal cells, suggesting that exposure to NPs leads to defective neuronal development, neuronal death, and accumulation of neuronal cell bodies. In astrocytes, GFAP decreased after exposure to PS-NPs, whereas the proinflammatory cytokines TNFa and IL-1b increased, indicating reactive astrocytosis.130 To summarize, this study revealed the toxicity of PS-NPs on neuronal cells through apoptosis. Furthermore, this investigation provides the insight that NPs neurotoxicity is dependent on cell type, as evident from the difference in PS-NP deposition and accumulation in different cells.
In contrast to in vitro studies, the neurotoxic potential of NPs has been extensively investigated in animal models. In vivo studies have highlighted the entry of NPs into experimental modules via multiple routes, including dietary, respiratory, or epidermal pathways. The behavioral changes in the experimental animals were also emphasized, providing more comprehensive data on the absorption, distribution, metabolism, and excretion of NPs in the body.
Rodents. One study in rodents investigated the neurotoxicity of pristine polystyrene NPs (25 and 50 nm) on neurobehavioral effects in animals. Four different dosages (1, 3, 6, and 10 mg/kg of body weight) of NPs were administered to four groups of adult Wistar male rats (6-8 weeks old; 190 ± 10 g) for 35 days through oral gavage using a 5 ml gavage needle to the pharyngeal region to avoid particle loss. At the end of the final week, several neurobehavioral tests were performed on six consecutive days to assess behavioral domains, including total locomotor activity, spatial working memory, anxiety-related behavior, motor coordination, and passive avoidance memory performance.
The results from the different tests were mixed: total locomotor activity in the open field, Y-maze test to assess spatial working memory, and passive avoidance test to investigate memory retention were normal for each rat in the four treatment groups. The elevated plus maze test to analyze anxiety-like behaviors in the rats was also normal, except for rats that received 10 mg polystyrene NPs/kg of body weight/day, which showed greater open arm entries, indicating reduced anxiety. The coordination and balance of rats receiving 3 mg polystyrene NP/kg of body weight/day, as assessed by the rotarod test, decreased compared to baseline values.131 Overall, this study suggests that pristine polystyrene NPs did not cause any significant adverse effects on the behavior of adult rats.
Invertebrates. In one neurotoxicity study, Caenorhabditis elegans was exposed to different concentrations of PS-NPs (25, 50, and 100 nm) for 72 h.132 This study highlighted the effect of PS-NPs on the growth and locomotion of C. elegans, induction of dopaminergic loss and mitochondrial damage, as well as oxidative stress and apoptosis. To assess growth and locomotion, C. elegans was exposed to different concentrations (10 and 100 μg/L) for 72 h, and parameters such as body length, head thrashes, and body bending were recorded. The results revealed that increasing concentrations of PS-NPs stunted body length, and the effect was more significant for larger particles (50 and 100 nm). In addition, 50 and 100 nm PS-NPs exerted obvious locomotor deficits.
In terms of head thrashes and body bends, the results revealed a reduction of 17.2% and 28.3% at 10 μg/L 50 nm PS-NP exposure and 29.0% and 37.0% at 100 μg/L 50 nm PS-NPs, respectively. As for 100 nm PS-NPs at the concentrations of 10 and 100 μg/L, head thrashes reduced by 17.4% and 29.2%, and body bends by 20.3% and 36.3%, respectively. These results indicated that PS-NPs exerted neurotoxicity by affecting C. elegans locomotion, and the effects were influenced by the size and concentration of PS-NPs.132
To further assess the neurotoxicity of PS-NPs, induction of dopaminergic loss and mitochondrial damage in C. elegans was evaluated. In this study, BZ555 strain transgenic C. elegans was exposed to a range of PS-NP concentrations, and fluorescently labelled dopaminergic neurons were measured. Researchers found that only 100 μg/L showed downregulation of dopaminergic content by 8.2%, 11.3%, and 12.2% at 25, 50, and 100 nm NPs, respectively. Furthermore, mitochondrial dysfunction was observed by labelling the mitochondria with green fluorescent proteins in the transgenic PD4251 strain C. elegans. A reduced fluorescence signal was noted after exposure to PS-NPs, and 100 nm particles revealed a lower fluorescence intensity at 10 and 100 μg/L when compared with those at 25 and 50 nm. Mitochondrial damage affects the mechanisms required for neurodevelopment, implying that PE-NPs exert neurotoxicity.
Finally, previous study evaluated oxidative stress and apoptosis in C. elegans following exposure to PS-NPs. Mitochondria play a significant role in ensuring a delicate balance of reactive oxygen species (ROS) levels, and excess production of ROS results in several disorders. In this test, oxidative stress induced by PS-NPs was measured using the DCFH-DA assay. Although there was no change in C. elegans exposed to 10 μg/L of 25 nm PS-NPs, an increased intracellular ROS level was noted in 100 μg/L of 25 nm PS-NPs was noted. Furthermore, both 50 and 100 nm NPs at concentrations of 10 and 100 μg/L showed a significant increase in ROS levels, with 100 nm PS-NPs recording the highest value. Apoptosis levels were measured based on lipofuscin and lipid peroxidation levels (oxidative stress markers). The results showed that higher concentrations of larger PS-NPs caused increased lipofuscin levels, which indicated a higher rate of apoptosis.132 In conclusion, PS-NPs exerted significant neurotoxicity in C. elegans depending on their concentration, and the extent of neurotoxicity was inversely proportional to the size of the NPs.
Another study was conducted on exposure to PS-NPs and its neurotoxicity in nauplii of brine shrimp Artemia fransiscana. To further understand the neurotoxicity of PS-NPs, A. fransiscana was considered a suitable model for planktonic species. In this study, 50 nm cationic amino-modified PS-NH2 was used at concentrations of 0.1, 1, 3, and 10 μg/mL, and each concentration was tested by incubating in a short- (48 h) and long-term exposure duration (14 days). The neurotoxic potential of PS-NPs was determined in accordance with physiological, chemical, and molecular responses. Physiological endpoints include growth, development, and feeding behaviors, whereas at the molecular level, the parameters are processes such as biotransformation, neuronal transmission, oxidative stress, and stress response. All endpoints were evaluated based on the activity of carboxylesterases (CbE), cholinesterase (ChE), glutathione-S-transferase (GST), CAT, LPO, and heat shock protein.
Results demonstrated that the highest mortality of over 70% was observed at highest concentration (10 μg/mL) and short-term exposure (48 h) evaluated using enzymatic activities. In this context, CbE was significantly inhibited, whereas ChE was unaffected in a concentration-dependent manner. There was a significant reduction in total body length at 1 and 10 μg/mL PS-NPs, but no reduction was observed upon exposure of 0.1 μg/mL PS-NPs. In comparison with long-term exposure (14 days), mortality rates were significantly different with increasing concentrations of PS-NPs. Developmental alterations were observed after 3 days of exposure to 1 and 3 μg/mL PS-NPs. Feeding behaviours, evaluated as the rate of ingestion and filtration, did not change after exposure to 0.1 and 1 μg/mL PS-NPs for 14 days. As for the oxidative response and detoxification, both GST and CAT levels were significantly decreased upon exposure. No differences were observed in GST between 0.1 and 1 μg/mL, and only CAT levels were significantly affected by exposure to 1 μg/mL.133
In summary, short-term exposure to PS-NPs may affect the growth and development of the brine shrimp Artemia fransiscana in a concentration-dependent manner, with an increase in oxidative stress. After long-term exposure to NPs, the survival of the test models was impaired due to neurotoxic effects. The presence of LPO, together with decreased ChE activity, indicates an accumulative effect.
Table 1 summarizes the neurotoxicity of NPs in different models.
| Models | Nanoplastics used | Time of exposure | Neurotoxic effects | References |
|---|---|---|---|---|
| Human embryonic stem cell (hESC)-derived (WA09 line) 3-dimensional (3-D) in vitro model | Polyethylene NPs (30 nm, anisotropic nanocrystals, ζ potential -30 mV) | 126 | ||
| Human neuroblastoma cells SH-SY5Y | Polystyrene NPs (50 nm, negatively discharged sulfonated surface, spherical) *Concentrations used: 2, 10 and 50 μl/ml | Not listed | 129 | |
| Primary cell types of CD-1 (ICR) mice: | Polystyrene NPs (nanoscale-PS latex beads, 100 nm diameter, ζ potential -40.60 ± 3.96 mV) *Concentrations used: 0, 50, 100, 200 mg/L | 2 days (48 h) | 130 | |
| Wistar male rats (6-8 weeks old; 190 ± 10 g) | Pristine polystyrene NPs (25 and 50 nm) *Concentrations used: 1, 3, 6 and 10mg polystyrene NPs/kg of body weight/day | 5 weeks (35 d) | 131 | |
| Caenorhabditis elegans | Polystyrene NPs (25, 50 and 100 nm) *Concentrations used: 10 and 100 μg/L | 3 days (72 h) | 132 | |
| Brine shrimp Artemia franciscana | PS-NH2 (50 nm, cationic amino modified) *Concentrations used: 0.1, 1, 3 and 10 μg/mL | Short term duration: 2 days (48 h) and Long term duration: 14 days | 133 |
To understand the association of NPs with neurodegenerative diseases, it is important to understand the interaction of NPs with proteins. As mentioned previously, the internalization of NPs allows interaction with biomolecules in the cytoplasm (in particular, proteins). The binding mechanism between proteins and NPs mainly involves weak interactions, including hydrophobic interactions, hydrogen bonds, van der Waals attraction forces, and electrostatic forces. The interaction of NPs with proteins can result in three consequences: (1) protein corona formation, (2) protein-induced coalescence of NPs, and (3) conformational changes in the protein secondary structure.134 Formation of a protein corona allows NPs to escape immune system surveillance and further impede the functions of cellular and molecular processes.134 Furthermore, the presence of a protein corona allows NPs to interact with each other through protein-protein interactions, thereby promoting coalescence and aggregation.135
The effects of NPs on the secondary structure of proteins are critical. This is because the function of a protein is dependent on its three-dimensional structure, and a slight alteration in protein structure may lead to protein misfolding and functional loss. Holloczki et al.136 demonstrated that NPs can modify the protein structure and ultimately lead to protein misfolding. The interaction between NPs and proteins is dependent on the presence of apolar side chains of amino acids, such as phenylalanine and tryptophan, which are more susceptible to adsorption on the surface of NPs due to hydrophobic interactions. Furthermore, the team investigated the interactions between NPs and small peptides with different secondary structures (such as α-helices and β-sheets). In this context, it was demonstrated that polyethylene NPs promote α-helix formation, whereas nylon NPs increase the presence of β-sheets and unfold helical domains. This indicates that NPs can result in protein misfolding.135
There is little evidence to imply that NPs can induce neurodegenerative diseases. However, studies related to nanomaterials have been conducted to study the potential risks that nanoscale objects can introduce in neurodegenerative diseases. Most nanoparticles can easily access bodily compartments, including the brain. An in vitro study conducted to understand how nanoparticles promote the formation of amyloid fibrils (by assembling proteins through nucleation mechanisms) showed that nanoparticles are encompassed by proteins once they are in contact with biological fluids and protein coronas are formed. This state can either denature the protein or remain unchanged, depending on the surface charge, hydrophobicity, and intrinsic stability of the proteins and nanoparticles.137
The binding of proteins to nanoparticles with large surface-to-volume ratios can eventually increase the chances of partial unfolding of proteins. As the levels of these abnormal proteins increase, so do the chances of association with one another, and this process may form new radical protein clusters. Furthermore, nanoparticles can affect protein self-assembly reactions, leading to the disruption of certain essential biological processes and eventually inducing protein misfolding-associated diseases. Amyloid fibrils are formed through the assembly and aggregation of misfolded proteins, forming insoluble fibers that are resistant to degradation. Neurodegenerative diseases accompanying amyloid fibril formation include Alzheimer’s disease (AD) and Creutzfeldt-Jacob disease.137
AD is a neurodegenerative disorder characterized by impaired cognition and behavior that can significantly affect a patient’s daily function. AD is initiated by the accumulation of senile plaques (SPs), also referred to as beta-amyloid plaques. SPs accumulate before the clinical onset of AD, and this is followed by a loss of cognitive function when neurofibrillary tangles (NFTs) are formed and eventually the loss of neurons. SPs mainly consist of insoluble deposits of proteins and cellular components surrounding neurons. This plaque is constructed of beta-amyloid proteins snipped from a larger protein, namely, the amyloid precursor protein. The fragments subsequently aggregated to form plaques.
Eventually, plaques are deposited in the hippocampus and in other areas of the cerebral cortex. In a healthy person, the microtubules in their neurons are stabilized by a type of protein, tau, to ensure proper transportation of nutrients from the cell body all the way to the end of the axon, and vice versa. However, tau proteins in patients with AD are chemically altered; they are tangled with each other and accumulate in neurons. Eventually, distorted tau proteins cannot bind to microtubules, resulting in disintegration and impaired communication between neurons, further causing neuronal death and thus forming NFTs. It is still unknown if the formation of SPs is a by-product of AD or if the plaques are the initial trigger for AD. However, beta-amyloid fibrillar proteins are extremely neurotoxic to neurons. Furthermore, the fibrillar form of beta-amyloid proteins has been shown to affect the phosphorylation state of the tau protein.
Another speculation is that NFTs lead to the formation of SPs, as the absence of normal tau proteins destabilizes microtubular systems, eventually distorting the function of the Golgi apparatus, forming abnormal proteins, and increasing amyloid-beta protein production. NPs could be a risk factor for inducing the formation of NFTs or SPs because of their ability to enhance the probability of protein misfolding and promote the self-assembly of proteins into amyloid fibrils.138
Liang et al. suggested that NPs can potentially trigger Parkinson’s disease (PD)-like neurodegeneration in mice based on brain single-nucleus transcriptomics. Several approaches were applied to understand the potential neurotoxicity of NPs by exposing 50 nm polystyrene NPs at doses ranging from 0.25-250 mg/kg on mice orally for 28 days. The evaluation methods include neurobehavior, magnetic resonance imaging (MRI), neuropathological examination, and transcriptomics. Moreover, transcriptional changes were observed in the mouse brain using single-nucleus RNA sequencing (snRNA-seq) techniques, providing insights into the transcriptomic analysis of cell populations at the single-cell level. This method was further confirmed by evaluating the levels of misfolded proteins, inflammation, and adenosine triphosphate (ATP) in different areas of the mouse brain (midbrain, hippocampus, striatum, and cortex).139
First, researchers investigated the ability of polystyrene NPs to penetrate the BBB and enter the mouse brain tissue through Evans blue staining. The intensity of staining was found to increase with increasing concentrations of polystyrene NPs, and these were dispersed throughout the brain, including the cortex, hippocampus, substantia nigra pars compacta (SNc), and striata.
To evaluate the effect of polystyrene NP on neurobehavior, such as the spontaneous activity of mice, locomotion, and anxiety levels, the rotarod test (for coordination and balance), grip strength, and distance travelled in an open field were assessed. The results demonstrated that the mice exhibited shorter travelled distances, weakened grip strength, and increased chances of falling off the rod after exposure to increasing concentrations of polystyrene NPs. Although neurobehavioral deficits were detected, symptoms of PD such as resting tremors and muscle rigidity were not observed. Furthermore, studies have revealed that Nissl bodies and dopaminergic neurons are lost in the SNc and the striatum.
An snRNA-seq test revealed that polystyrene NPs can induce cell-specific responses, with the responses mainly linked to disorders in energy metabolism and failure in mitochondrial function, especially in excitatory neurons. This test was further verified by the decreased ATP content and expression levels of ATP-associated genes and proteins. Inflammatory instability can be observed in astrocytes and microglia, with further dysregulation of synaptic functions and proteins in astrocytes, oligodendrocytes, and endotheliocytes.
All the above-mentioned complications are indications of PD-like neurodegeneration, especially with the induction of diminished motor function. Implications such as decreased Nissl bodies, dopaminergic neurons, or level of ATP content mainly occur in the SNc and striatum, which are the main lesion sites of PD. Dopaminergic neurons could not be distinguished by snRNA-seq, but inhibition of the ATP metabolic gene and mitochondrial dysfunction were mainly manifested in excitatory neurons, including dopaminergic neurons. This could be a major cause of dopaminergic neuron depletion, thereby promoting PD-like neurodegeneration.139
The use of plastics is inevitable in daily life. The growing production and usage of plastics have contributed to the accumulation of plastic waste. Unfortunately, many levels of society are still oblivious to proper management of plastic waste, resulting in environmental pollution. Plastic waste can be degraded into smaller fragments, such as MPs, and further into NPs if exposed to factors such as UV radiation, weathering, and biodegradation under prolonged exposure.
Humans are then exposed to MPs and NPs, which are accessible because of their minute dimensions, enabling them to travel to any anatomical region without further resistance. Scientists have proven the possibility of MPs and NPs entering the human body via the ingestion of food or drinking water, inhalation, or contact. Moreover, several articles have also highlighted the various toxicities that MPs and NPs can induce in humans, in particular neurotoxicity. Compared to MPs, NPs have become intensely investigated in emerging research due to their enhanced capacity to move freely across biological components and increased toxicity due to their nano-characteristics.
Most concerns on NP toxicity lean towards the central nervous system, and NPs are regarded as a worse ‘extension’ of MPs in many circumstances. Evidence shows that the ability of NPs to pass through the BBB into the brain tissue may lead to neurotoxic mechanisms, such as induction of oxidative stress and inhibition of AChE activity in vitro and alteration of neurotransmitter levels or behavioral changes in vivo. However, clinical studies of human exposure are lacking.
Many questions remain to be answered in the quest to understand the impact of NPs on the human body. Most research has been conducted on cell lines, which overlook physiological processes, such as the body’s immune reaction to the entry of foreign substances. Detection methods or instruments to accurately measure the concentration of NPs in the environment and within the body are yet to be defined, as the techniques are predominantly experimental. Even the definition of ‘NPs’ are not well understood, as a relatively novel research field. While NPs are evidently present in the aquatic environment, data are scarce on the threshold levels that will impact the aquatic system and food chain, causing bioaccumulation and potential risks to humans.
Evidence of NP neurotoxicity is even less explicit, and most studies have depended on in vitro experiments. Future work should strive to elucidate the effect of leached chemicals and plastic materials, the concentration of NPs in the environment, chronic exposure, and risk assessment of NPs in humans. The focus should also include NPs from terrestrial environments in addition to marine environments.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)