Guo T, Deinhardt K and Wang Y. Single Nucleotide Polymorphisms in ANK3 and Psychiatric Risk: A Meta-Analysis, Systematic Review, and Quantitative Trait Locus Insights [version 1; peer review: 1 approved with reservations]. F1000Research 2025, 14:1020 (https://doi.org/10.12688/f1000research.169014.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Biological Sciences, Faculty of Environmental and Life Sciences, University of Southampton, Southampton, SO17 1BJ, UK 2University of Bremen, Cell Biology, Bremen, 28359, Germany 3Institute for Life Sciences, University of Southampton, Southampton, SO17 1BJ, UK 4NIHR Southampton Biomedical Research Centre, University Hospital Southampton, Southampton, England, SO16 6YD, UK
Tao Guo
Roles:
Data Curation,
Formal Analysis,
Investigation,
Methodology,
Software,
Validation,
Visualization,
Writing – Original Draft Preparation,
Writing – Review & Editing
Ankyrin 3 (Ankyrin G), encoded by the ANK3 gene, is a membrane-associated scaffold protein critical for neuronal signaling and highly enriched in axonal initial segments and nodes of Ranvier. ANK3 variants have been implicated in several neuropsychiatric disorders.
Objectives
To identify single nucleotide polymorphisms (SNPs) in ANK3 associated with psychiatric disorders, assess population-specific significance, determine variant locations, and explore regulatory effects through quantitative trait locus (QTL) analysis.
Methods
A meta-analysis and systematic literature review were conducted to identify disease-associated SNPs in ANK3. QTL analyses were performed using datasets from the eQTL Catalogue and GTEx v10 to evaluate regulatory effects in brain tissues.
Results
Meta-analysis identified significant associations between ANK3 SNPs and psychiatric disorders. In bipolar disorder, rs10994336 (OR = 1.28, P = 1.04×10−4) was significant in European+North American populations; rs9804190 was protective in Europeans (OR = 0.88, P = 0.020); and rs1938526 was significant in both European+North American (OR = 1.32, P = 7.26×10−12) and Asian populations (OR = 1.08, P = 4.68×10−12), with an overall effect (OR = 1.17, P = 0.003). In schizophrenia, rs10761482 showed opposite effects: protective in Chinese (OR = 0.74, P = 0.002) and risk-associated in Iranian cohorts (OR = 1.57, P = 0.025). rs10994359 was protective in major depressive disorder in Asians (OR = 0.69, P = 0.016), and three SNPs reduced PTSD risk (OR = 0.48, P = 0.045). All significant variants were intronic. QTL analyses showed rs10761482 increased ANK3 expression in neocortex (P = 0.001) and neurons (P = 0.038); three PTSD SNPs showed robust cerebellar splicing effects (FDR = 0.010); and apaQTLs indicated modest 3′-UTR effects in putamen.
Conclusion
Multiple intronic ANK3 SNPs are significantly associated with psychiatric disorders and display tissue-specific regulatory effects. Integrating QTL data provides insights into their potential functional roles in neuropsychiatric pathogenesis.
Keywords
ANK3, psychiatric disorders, bipolar disorder, schizophrenia, major depression, post‑traumatic stress disorder, single nucleotide polymorphism, QTL analysis
ANK3 (Ankyrin-G) is a membrane-associated scaffold protein encoded by the ANK3 gene on chromosome 10q21.2. It plays a fundamental role in organizing the axon initial segment (AIS) by anchoring voltage-gated sodium channels and adhesion molecules to the cytoskeleton, thereby ensuring proper neuronal polarity and efficient action potential initiation. Disruption of ANK3 function compromises AIS integrity and has been implicated in neuropsychiatric disorders such as bipolar disorder and schizophrenia, highlighting its importance in maintaining neuronal excitability and circuit stability.1 In the central nervous system, ANK3 is abundantly expressed in the neocortex and hippocampus—key regions involved in cognitive processing, emotional regulation, and behavioral control. Owing to its essential role in neuronal development, membrane organization, and signal transduction, ANK3 expressed as multiple isoforms arising from alternative splicing, which differ in both molecular weight and subcellular localization. The high-molecular-weight isoforms (~270–480 kDa) are predominantly enriched at the axon initial segment (AIS), where they interact with voltage-gated sodium channels and cytoskeletal components to maintain neuronal polarity and excitability.2 In contrast, shorter isoforms are typically localized at postsynaptic densities and are thought to contribute to dendritic spine formation and synaptic plasticity.3ANK3 has been repeatedly identified as a candidate gene in genetic and functional studies of neuropsychiatric disorders.4
Over the past decade, genome-wide association studies have identified significant associations between ANK3 single nucleotide polymorphisms (SNPs) and a range of psychiatric disorders, including bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD). These findings suggest that genetic variation within the ANK3 locus may contribute to shared pathophysiological mechanisms underlying multiple neuropsychiatric phenotypes.5,6 However, despite accumulating evidence, considerable inconsistencies remain regarding which ANK3 SNPs are most robustly associated with psychiatric disorders and whether these associations are consistent across diverse populations. Such uncertainties highlight the need for systematic synthesis of existing findings and comprehensive functional characterization of disease-associated variants.
To address this gap, the present study integrates meta-analysis with QTL-based transcriptomic analysis to comprehensively evaluate the role of ANK3 genetic variants in psychiatric disorders. We first conducted a meta-analysis of published association studies to identify statistically significant ANK3 SNPs across bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD), incorporating population stratification to examine potential ethnic differences. Candidate SNPs identified through this approach were subsequently subjected to functional investigation using large-scale transcriptomic and QTL datasets to elucidate their molecular regulatory effects.
Specifically, we performed expression quantitative trait locus (eQTL) analysis using the eQTL Catalogue to determine whether disease-associated ANK3 SNPs modulate gene expression levels in brain tissues. In parallel, alternative polyadenylation QTL (apaQTL) data from GTEx v10 were utilized to investigate the impact of these variants on ANK3 3′-end processing, which may influence transcript isoform usage, mRNA stability, and translational efficiency. To further assess regulatory effects on transcript structure, splice QTL (sQTL) data from GTEx v10 were analyzed to examine whether the identified SNPs affect alternative exon inclusion or splice junction usage within ANK3.7
By integrating statistical associations with multilayered transcriptomic evidence—including gene expression, alternative polyadenylation, and splicing—this study seeks to elucidate the functional relevance of ANK3 genetic variants in the pathogenesis of psychiatric disorders. Our findings reconcile prior inconsistencies in GWAS through meta-analysis and uncover potential mechanisms by which noncoding intronic SNPs contribute to disease via modulation of ANK3 expression and post-transcriptional regulation in a tissue-specific context. This comprehensive framework underscores the value of combining genetic epidemiology with functional genomics to advance our understanding of psychiatric disease biology and to inform the development of targeted therapeutic strategies.
Method
Criteria for inclusion
In this analysis, we included both randomized controlled trials (RCTs) and observational studies involving individuals diagnosed with neurological or psychiatric disorders across diverse geographic regions. For each patient cohort, a matched control group comprising healthy individuals from the same population and study was selected. Eligible studies were published in English between January 1, 2000, and recent years, and were required to report associations between ANK3 SNPs and disease risk. Risk estimates were extracted in the form of odds ratios (OR), while SNP relationships were assessed using either minor allele frequency (MAF) or specific genotype distributions. For cohorts stratified by region, meta-analysis was performed only when a consistent direction of effect was observed across all subgroups. SNPs reported in two or more independent studies were included in the quantitative meta-analysis, whereas those reported in a single study were synthesized narratively.
Criteria for exclusion
Studies employing hazard ratios (HR) for SNP risk estimation were excluded. We also omitted investigations not focused on neurological or psychiatric disorders, as well as those lacking clear information regarding the geographic or national origin of participants outside Europe, America, or Asia. Articles that did not specifically address risk associations with ANK3 SNPs were disregarded. In addition, animal studies, medical records, case reports, expert commentaries, and review articles were excluded. Only peer-reviewed articles published in English were included in the final dataset.
Literature retrieval and selection
In accordance with the predefined inclusion criteria, two independent researchers (TG and YW) systematically searched PubMed, Web of Science, and MEDLINE using combinations of the terms “genome-wide association study” with “ANK3”, as well as “nucleotide polymorphisms” with “ankyrin 3” or “Ankyrin-G”. Relevant studies were screened, and data were extracted independently by both reviewers using a standardized data extraction form. Any discrepancies were resolved through discussion with a third researcher until consensus was achieved. All analyses were subsequently conducted by two researchers. This multi-reviewer approach was implemented to minimize subjective bias and enhance the reproducibility of the findings.
Quality assessment of included studies
The studies with controlled trials were evaluated by the Newcastle-Ottawa Scale (NOS)8,9 by two different researchers independently. NOS include 1. Selection: 1) Is the case definition adequate? 2) Representativeness of the cases. 3) Selection of controls. 4) Definition of Controls. 2. Comparability. 1) Comparability of cases and controls based on the design or analysis. 3. Exposure. 1) Ascertainment of exposure. 2) Same method of ascertainment for cases and controls. 3) Non-Response rate. The total score is 9, and if a study can get over 5, it can be assessed as good quality. Those articles which did not meet the quality requirement were excluded.
Extraction of data
In accordance with our eligibility criteria, studies were evaluated based on study design, characteristics of patient and control cohorts, and clearly defined outcome measures. From each eligible study, we extracted key variables including lead author, year of publication, number of cases, participant nationality, age distribution, gender ratio, smoking status, reported odds ratio (OR), corresponding 95% confidence interval (CI), and associated p-value. The complete extracted dataset used for conducting the meta-analysis, including all OR and confidence intervals for each ANK3 SNP across psychiatric disorders, is available as Extended dataset 1 (Figshare, https://doi.org/10.6084/m9.figshare.29886620).10
Synthesis and analysis of data and assessment of publication bias
Utilizing the “meta (version 4.18-0)” package within R 4.0.1 and RStudio, we conducted a meta-analysis on the specified SNPs. Odds ratios (OR) and 95% confidence intervals (95%CI) were input to yield synthesized outcomes. For studies presenting results as MAF with case numbers for both patient and control groups, preliminary calculations for OR and 95%CI were necessary. We combined the funnel plot and the Egger test11 to assess the publication bias of the meta-analysis. As none of the meta-analyses included 10 or more studies, publication bias was not assessed, in accordance with Cochrane guidance.12
Following the meta-analysis of psychiatric trait SNPs, we conducted an integrative QTL examination to determine how the ten identified ANK3-associated variants influence transcript regulation in brain tissue. eQTL slopes and p values were retrieved from the eQTL Catalogue (https://www.ebi.ac.uk/eqtl/)14; when the same SNP–tissue pair appeared in more than one study, a fixed-effect inverse variance model was used to obtain a single β estimate. apaQTL (alternative polyadenylation) and sQTL (splicing) summary statistics were retrieved from the GTEx v10 resource (https://gtexportal.org/home/datasets)15 and filtered to chromosome 10 entries mapping to ANK3. For every brain tissue, we extracted the slope (β), its standard error, and the nominal p value.
To correct for multiple testing across tissues and regulatory layers, we applied Benjamini–Hochberg false discovery rate (FDR)16 correction to the nominal p-values within each QTL modality (eQTL, apaQTL, and sQTL). Associations with FDR < 0.05 were considered statistically significant. In cases where nominal significance (p < 0.05) was observed but FDR correction yielded non-significant results, the findings were interpreted as suggestive and reported with appropriate caution. All datasets were parsed and harmonised by rsID using custom Python 3.11 and Bash scripts. Data were visualised using GraphPad Prism 9 and matplotlib in Python. Throughout all analyses, β values are reported to indicate the direction and magnitude of allele effects on gene expression (eQTL), 3′-end/poly(A) site usage (apaQTL), or splice junction usage (sQTL).
Results
Literature extraction and quality assessment
Using the predefined search strategy, a total of 1,140 articles were retrieved from the selected databases. Following title and abstract screening, duplicate entries were removed. Full-text review resulted in the exclusion of 214 studies, primarily due to methodological insufficiencies or unclear study protocols. Ultimately, 27 English-language studies met the inclusion criteria and were retained for analysis. All included studies satisfied the quality standards as assessed by the Newcastle–Ottawa Scale. The key characteristics of the selected studies are summarized in Extended dataset 2 (https://doi.org/10.6084/m9.figshare.30090913),17 and the literature selection process is outlined in
Figure 1.
Figure 1. The process of literature screening.
*For exclusion, please refer to section Criteria for Exclusion.
Meta-analysis and synthetic review of the risk of ANK3 SNPs with bipolar disorder
The meta-analysis identified five ANK3 SNPs—rs10994336, rs9804190, rs10994397, rs1938526, and rs139972937—as loci of interest across psychiatric disorders. Rs10994336, included in nine studies from European and North American populations, emerged as a significant risk locus (OR = 1.28, 95% CI: 1.13–1.45, P = 1.04 × 10−4), but did not show significance in Asian populations (P = 0.600). Rs9804190 was reported in three European studies, where it showed a significant protective effect (OR = 0.88, 95% CI: 0.79–0.98, P = 0.020), though this association was not observed when extended to the combined European and North American cohort (P
= 0.173). Rs10994397 was identified as a significant risk variant in two European studies (OR = 1.35, 95% CI: 1.24–1.47, P = 4.71 × 10−12), but this effect was not replicated in the broader dataset (P = 0.254). In contrast, rs1938526 demonstrated consistent risk associations across both European + North American (five studies; OR = 1.32, 95% CI: 1.22–1.43, P = 7.26 × 10−12) and Asian populations (three studies; OR = 1.08, 95% CI: 1.04–1.12, P = 4.68 × 10−12), with pooled analysis confirming its significance (OR = 1.17, 95% CI: 1.05–1.29, P = 0.003). Additionally, rs139972937, although reported in a single European study, was significantly associated with bipolar disorder (P = 0.040) and was thus retained for synthetic review (
Figure 2 and Extended dataset 3 (https://doi.org/10.6084/m9.figshare.30090964)18).
Figure 2. Forrest plots of the risk of ANK3 SNPs in patients with bipolar disorder.
(A) Comparison of ANK3 (rs10994336) MAF risks in BP patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs10994336) MAF risks. (B) Comparison of ANK3 (rs9804190) MAF risks in BP patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs9804190) MAF risks. (C) Comparison of ANK3 (rs10994397) MAF risks in BP patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs10994397) MAF risks. (D) Comparison of ANK3 (rs1938526) MAF risks in BP patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs1938526) MAF risks.
Meta-analysis and synthetic review of the risk of ANK3 SNPs with schizophrenia
This segment of the meta-analysis focused on four ANK3 SNPs previously implicated in schizophrenia—rs1938526, rs10761482, rs10994336, and rs9804190. Across all included studies, none of these variants demonstrated a statistically significant association with schizophrenia in their respective populations (P > 0.05). To further explore potential population-specific effects, subgroup analysis was conducted for rs10761482 within the Asian population, which was stratified into three subgroups based on Hardy–Weinberg equilibrium (HWE) status and ethnicity. The first subgroup comprised Chinese cohorts not in HWE (details regarding HWE evaluation are presented in the Discussion), the second subgroup included Iranian participants, and the third subgroup consisted of Chinese cohorts with significant HWE conformity. Notably, in the Chinese subgroup conforming to HWE, rs10761482 exhibited a significant protective association (OR = 0.74, 95% CI: 0.61–0.90, P = 0.002), while in the Iranian subgroup, it was identified as a potential risk locus (OR = 1.57, 95% CI: 1.06–2.33, P = 0.025), suggesting possible ethnic or population-specific heterogeneity in the genetic architecture of schizophrenia (
Figures 3 & 4; Extended dataset 3 (https://doi.org/10.6084/m9.figshare.30090964)18).
Figure 3. Forrest plots of meta-analysis of the risk of ANK3 SNPs in patients with Schizophrenia.
(A) Comparison of ANK3 (rs1938526) MAF risks in SZ patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs1938526) MAF risks. (B) Comparison of ANK3 (rs10761482) MAF risks in SZ patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs10761482) MAF risks. (C) Comparison of ANK3 (rs10994336) MAF risks in SZ patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs10994336) MAF risks. (D) Comparison of ANK3 (rs9804190) MAF risks in SZ patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs9804190) MAF risks.
Figure 4. Forrest plots of subgroup meta-analysis of the risk of ANK3 rs10761482 in Asian patients with Schizophrenia.
Comparison of ANK3 (rs10761482) MAF risks in SZ patients and healthy control of Asian population. Forest plot showing meta-analysis of ANK3 (rs10761482) MAF risks.
Meta-analysis and synthetic review of the risk of ANK3 SNPs with major depression and posttraumatic stress disorder
This portion of the meta-analysis examined the association between ANK3 variants and two psychiatric conditions: major depressive disorder (MD) and post-traumatic stress disorder (PTSD). Specifically, the analysis focused on rs10994336 and rs10994359 in relation to MD, and on rs28932171, rs11599164, and rs17208576 in relation to PTSD. Rs10994336 did not demonstrate a significant association with MD in either North American or Asian populations (P > 0.05). In contrast, rs10994359 was identified as a significant protective variant in Asian populations (OR = 0.69, 95% CI: 0.51–0.93, P = 0.016), suggesting potential ethnic-specific effects. For PTSD, all three investigated variants—rs28932171, rs11599164, and rs17208576—were found to be significantly associated with reduced risk, each exhibiting the same odds ratio (OR = 0.48) and a nominally significant P-value (P = 0.045), indicating their potential role as protective loci (
Figure 5; Extended dataset 3 (https://doi.org/10.6084/m9.figshare.30090964)).18
Figure 5. Forrest plot of meta-analysis of the risk of ANK3 SNPs in patients with major depression.
Comparison of ANK3 (rs10994336) MAF risks in MD patients and healthy control. Forest plot showing meta-analysis of ANK3 (rs10994336) MAF risks.
Plotting of ANK3 significant SNPs
To further elucidate the potential functional relevance of ANK3 risk variants, we mapped the SNPs identified as significantly associated with bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD) onto the ANK3 gene structure. Genomic localization was determined using the GRCh38.p14 assembly (GCF_000001405.40), with chromosomal coordinates corresponding to NC_000010.11 and transcript ENST00000280772.7. Structural annotation revealed that rs9804190 and rs139972937 (associated with BD), as well as rs28932171, rs11599164, and rs17208576 (associated with PTSD), are all located within the same intronic region between exon 36 (ENSE00001736354) and exon 37 (ENSE00001671026). Notably, all SNPs showing significant associations across the four psychiatric disorders investigated reside within intronic regions of ANK3, suggesting that noncoding regulatory elements within introns may contribute to disease susceptibility through transcriptional or post-transcriptional mechanisms (
Figure 6;
Table 1).
Figure 6. Location of SNPs in the ANK3 gene significantly associated with psychiatric disorders.
The gene map was derived from the human genome reference sequence Assembly: GRCh38.p14 (GCF_000001405.40) with locus: NC_000010.11. The transcript ID referenced for intron and exon information is ENST00000280772.7. SNPs obtained from meta-analysis are labelled in red, whereas SNPs obtained by synthetic review are labelled in black at the corresponding positions.
Table 1. Summary of results of Localisation of SNPs in the ANK3 gene significantly associated with psychiatric disorders.
Exon No.
Exon/Intron
Start
End
Bipolar disorder
Schizophrenia
Major depression
Posttraumatic stress disorder
5' upstream sequence
//
2
ENSE00003669229
60,615,224
60,615,186
Intron 2-3
60,615,185
60,279,640
rs10994336,rs10994397,rs1938526
rs10761482
rs10994359
3
ENSE00002439064
60,279,639
60,279,538
//
36
ENSE00001736354
60,080,618
60,080,537
Intron 36-37
60,080,536
60,068,010
rs9804190,rs139972937
rs28932171,rs11599164,rs17208576
37
ENSE00001671026
60,068,009
60,067,935
//
QTL analysis of psychiatric risk SNPs at the ANK3 locus
Among the ten SNPs associated with psychiatric disorders within the ANK3 locus, we identified significant regulatory effects on gene expression, alternative polyadenylation (APA), and splicing in human brain tissues. In the expression quantitative trait locus (eQTL) analysis using the eQTL Catalogue, rs10761482 demonstrated a notable association with ANK3 expression in both neuron-enriched and neocortical cell populations. In neurons, the variant was associated with increased expression (β = 0.12, SE = 0.06, P = 0.038), although the signal did not remain significant following false discovery rate (FDR) correction (FDR = 0.999). In contrast, a stronger effect was observed in the neocortex (β = 0.14, SE = 0.04, P = 0.001), with marginal significance after multiple testing adjustment (FDR = 0.162). These findings suggest that rs10761482 may exert cell-type-specific regulatory effects on ANK3 expression, particularly within neuronal populations. (
Figure 7A, B;
Table 2).
Figure 7. Comparative analysis of eQTL and apaQTL effects of ANK3-associated SNPs across brain tissues and cell types.
(A) Heatmap displaying β values (slope estimates) of expression quantitative trait loci (eQTLs) for ten ANK3-associated SNPs across various brain regions and cell types. The eQTL data were obtained from the eQTL Catalogue, and included both bulk tissues and fine-grained cellular populations (e.g., neuron, neocortex, microglia). (B) Bar plot showing the β value ± standard error (SE) of rs10761482 in neuron and neocortex cell types, indicating differential eQTL effects across these populations. (C) Heatmap showing β values of alternative polyadenylation QTLs (apaQTLs) for the same SNP set across multiple bulk brain tissues. The apaQTL data were derived from GTEx v10 apaQTLs database. (D) Bar plot of β ± SE for four SNPs with significant apaQTL associations (p < 0.05) in the Putamen basal ganglia, illustrating their negative effect on ANK3 3' end usage. All SNPs were selected based on their reported association with psychiatric disorders and genomic proximity to ANK3 in meta-analyses. β values represent slope estimates from linear regression models. Significance was determined by nominal p-value < 0.05.
Table 2. Summary of ANK3 SNPs associated expression QTL (eQTL) effects on ANK3 in brain-related tissues.
Tissue_label
Rsid
Beta
SE
P-val_nominal
P-value_FDR
Neocortex
rs10761482
0.14
0.04
0.001
0.162
Neuron
rs10761482
0.12
0.06
0.038
0.999
In the alternative polyadenylation QTL (apaQTL) analysis using GTEx v10 data, four SNPs—rs28932171, rs11599164, rs17208576, and rs9804190—were found to significantly influence the 3′-end processing of ANK3 transcripts in the putamen, a region of the basal ganglia. All four variants exhibited negative slope values, indicating that the presence of risk alleles was associated with decreased usage of distal polyadenylation sites. Rs28932171, rs11599164, and rs17208576 showed identical effect sizes (β = –0.26, SE = 0.12, P = 0.033), while rs9804190 displayed a slightly weaker but still nominally significant association (β = –0.21, SE = 0.09, P = 0.018). However, none of these associations remained significant following false discovery rate (FDR) correction (FDR = 0.965), suggesting that the observed APA effects are modest and require further validation in independent datasets (
Figure 7C, D;
Table 3).
Table 3. Summary of ANK3 SNPs associated alternative polyadenylation (APA) QTL of ANK3 in brain-related tissues.
Tissue_label
Rsid
Beta
SE
P-val_nominal
P-value_FDR
Putamen_basal_ganglia
rs28932171
-0.26
0.12
0.033
0.965
Putamen_basal_ganglia
rs11599164
-0.26
0.12
0.033
0.965
Putamen_basal_ganglia
rs17208576
-0.26
0.12
0.033
0.965
Putamen_basal_ganglia
rs9804190
-0.21
0.09
0.018
0.965
In contrast to the more modest effects observed in eQTL and APAQTL analyses, the splice QTL (sQTL) analysis revealed robust associations between the same three SNPs—rs28932171, rs11599164, and rs17208576—and a specific splicing event within ANK3 in the cerebellum. These variants exhibited strong and consistent effect sizes (β = 0.67–0.70, SE = 0.15), accompanied by highly significant nominal P-values (P = 1.06 × 10−5 to 1.64 × 10−5) and false discovery rate (FDR) values of 0.010. These findings provide compelling evidence for the role of these SNPs in modulating alternative splicing of ANK3 in brain tissue (
Table 4). Collectively, the integration of eQTL, APAQTL, and sQTL analyses demonstrates that psychiatric disorder-associated ANK3 variants may exert regulatory effects through multiple layers of RNA regulation. Among these, splicing alterations in the cerebellum show the most statistically robust signal, whereas expression and polyadenylation effects appear more modest and context dependent. Full results for each QTL analysis are provided in the extended data: eQTL (Extended dataset 4, https://doi.org/10.6084/m9.figshare.29901494),19 APAQTL (Extended dataset 5, https://doi.org/10.6084/m9.figshare.29901641),20 and sQTL (Extended dataset 6, https://doi.org/10.6084/m9.figshare.29901662).21
Table 4. Significant ANK3 SNPs associated Splicing QTL (sQTL) of ANK3 in brain-related tissues.
Previous studies have demonstrated that ANK3 single nucleotide polymorphisms (SNPs) contribute to the genetic risk of various psychiatric disorders. However, a comprehensive meta-analysis integrating cross-disorder associations and functional mapping has been lacking. In this study, we systematically analyzed the relationship between ANK3 SNPs and four major psychiatric conditions—bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD)—using both meta-analysis and systematic review approaches. Several SNPs showed significant associations with disease risk, with notable differences in effect size and direction across populations. When mapped onto the ANK3 gene structure, all significantly associated variants were found within intronic regions. This intronic clustering suggests that noncoding regulatory mechanisms, rather than direct coding sequence alterations, may underlie the pathogenic effects of these variants—potentially through modulation of splicing, transcriptional regulation, or alternative polyadenylation.
The meta-analysis revealed that all the SNPs related to psychiatric disorders are concentrated in two introns. In particular, five SNPs related to BD and PTSD are found within the intron preceding the brain-specific exon 37. This exon 37 (ENSE00001671026) of ANK3 encodes a key element of the AIS targeting motif, influencing the subcellular localization of Ankyrin-G in neurons. Transcript structure analysis revealed that it is retained in longer isoforms such as ENST00000280772.7 (ANK3-201), but absent in several shorter splice variants, including ENST00000616444.4 (ANK3-228).13 It is partially or fully included in the transcripts giving rise to the AIS-localised protein isoforms of 270 kDa and 480 kDa, respectively, but excluded from the postsynaptic 190 kDa isoform of AnkyrinG. Loss of exon 37 affects action potential generation and neuronal signal integration.22 Interestingly, three of the SNPs significantly associated with psychiatric disorders—rs28932171, rs11599164, and rs17208576— that are located in the intronic region between exon 36 (ENSE00001736354) and exon 37 exhibit strong splicing QTL (sQTL) signals specifically in cerebellum tissue (
Table 4). These findings suggest that the risk variants may influence alternative splicing decisions that determine exon 37 inclusion. Such splice isoform shifts could lead to changes in the intracellular distribution of Ankyrin-G, potentially altering its ability to cluster ion channels at the AIS and thereby affecting neuronal polarity, excitability, or signaling, leading to cellular and circuit-level changes in neuronal signalling, and to disease in the long term.
Additionally, SNPs in the intron of ANK3 may also affect binding of miRNAs (microRNAs) or disrupt enhancer or suppressor elements that regulate gene expression.23,24 For example, it has been shown that a SNP in miR-137 is associated with an increased risk of schizophrenia. SNPs may alter the binding pattern of miRNAs, leading to overexpression or repression of key genes. The same may be true for ANK3. The concentration of SNPs in both introns could mean that these regions are evolutionarily conserved and therefore may have some functionality.25 So in summary, although the specific functions of these two regions are currently unknown, it would be interesting to explore the specific biological functions of these two introns in further studies.
In our meta-analysis, we found that the significance of associations between some SNPs in ANK3 and disease varied considerably in different populations. For example, some single nucleotide polymorphisms are significant in some populations in Europe but not in the study of both European and North American populations; or significant in European and American populations but not in Asian populations; or even the same SNPs show opposite trends or significance in different populations. This has a significant relationship with their different populations and genetic backgrounds. For example, Asian populations are very diverse and contain East Asians Central Asians and others. In our meta-analysis of schizophrenia, we found that the significance of rs10761482 in Chinese and Iranian populations showed completely opposite characteristics, one being a risk locus and the other a protective locus. There are some differences between the Chinese and Iranian populations, as most of the Chinese population belongs to East Asian ethnicity, while the Iranians belong to West Asian or Middle Eastern ethnicity. Our meta-analyses revealed greater heterogeneity within some subgroups, which may be related to the fact that there are still differences in the populations included in the same population subgroup. And as we mentioned in the research methodology, we can only further meta-analyse these subgroups if the different population subgroups show the same significance and are all significant. Otherwise, if one population shows significance and the other does not, then even if they show significance as a whole, it is meaningless and is a false positive.26–28
In our meta-analysis of rs10761482 for schizophrenia, we conducted further subgroup analyses of Asian populations into Chinese and Iranian, and one study of Chinese was separated into a separate group because it was significant for the HWE test. The HWE (Hardy-Weinberg Equilibrium) test is a statistical test based on the Hardy-Weinberg equilibrium law. The Hardy-Weinberg law of equilibrium describes how the gene and genotype frequencies of a population remain constant under certain conditions. Significant deviation from Hardy–Weinberg equilibrium (HWE) may indicate non-random mating, population stratification, genotyping error, selection pressure, or recent mutation or migration events that alter genotype frequencies. To minimize potential confounding from such factors, studies with significant HWE deviation were analyzed as a separate subgroup.29
We also conducted a targeted QTL analysis of ten pre-selected ANK3 SNPs previously associated with psychiatric disorders through meta-analysis. Although the scope of variant testing was limited, we applied false discovery rate (FDR) correction to account for multiple comparisons across tissues and regulatory modalities. Unlike genome-wide QTL studies, which necessitate stringent correction due to the extensive number of hypotheses tested, our focused design involved a comparatively small set of SNP–phenotype pairs. Nonetheless, implementing FDR correction in this context strengthens the statistical rigor of our analysis and increases confidence in the robustness and reproducibility of the identified associations.
The application of FDR correction enabled us to differentiate nominal associations from those with stronger statistical support after accounting for multiple testing. In the eQTL analysis, rs10761482 demonstrated nominal significance in both neurons and neocortex, with only the neocortex association approaching FDR significance (FDR = 0.162), suggesting moderate evidence for a cell type–specific regulatory effect. In contrast, apaQTL signals for rs28932171, rs11599164, rs17208576, and rs9804190 in the putamen were all nominally significant (P < 0.05), yet none withstood FDR correction (FDR = 0.965), indicating that these intronic variants may influence 3′-end transcript processing, but further validation is required. Most notably, sQTL analysis revealed strong and FDR-significant associations (FDR = 0.010) between rs28932171, rs11599164, and rs17208576 and a specific ANK3 splicing event in the cerebellum, providing robust evidence that these SNPs exert a functional impact on transcript structure in a brain region–specific manner.
It is important to recognize that nominally significant but FDR-nonsignificant results may still carry biological relevance, particularly within hypothesis-driven, low-dimensional analyses. While FDR correction is essential for controlling the expected proportion of false discoveries across multiple comparisons, its statistical stringency can obscure true signals—especially when effect sizes are moderate or sample sizes are limited. In our study, the consistency of effect direction across tissues and regulatory modalities—for instance, the increased ANK3 expression associated with rs10761482 in both neurons and neocortex, and the concordant negative APA effects of four SNPs in the putamen—supports the plausibility of underlying regulatory activity. These findings, although not FDR-significant, may reflect genuine biological effects and warrant further validation through functional assays or replication in independent cohorts.
Moreover, the brain regions exhibiting the strongest regulatory signals in our QTL analysis—namely, the neocortex and cerebellum—are well-established contributors to the pathophysiology of psychiatric disorders. The neocortex plays a central role in higher-order cognitive functions, emotional regulation, and sensory integration, and structural and functional abnormalities in this region have been consistently reported in schizophrenia, bipolar disorder, and major depressive disorder. The observed association between rs10761482 and ANK3 expression in neocortical cells thus offers a plausible molecular link between genetic risk and cortical dysfunction. Similarly, although historically considered a motor coordination center, the cerebellum is increasingly recognized for its involvement in mood regulation, cognitive processing, and social behavior. The robust sQTL effects observed in the cerebellum suggest that ANK3 splicing regulation in this region may contribute to psychiatric vulnerability, aligning with growing evidence implicating cerebellar dysfunction in conditions such as schizophrenia and autism spectrum disorder.
Together with our findings, a previous methylation QTL (meQTL) study,30 which reported that rs10994336 was significantly associated with ANK3 methylation levels in the prefrontal cortex, further reinforcing the regulatory relevance of disease-associated ANK3 variants. Together, these results suggest that intronic ANK3 SNPs may modulate disease risk not only through genetic association but also via transcriptional and post-transcriptional regulatory mechanisms.
Although there is no previous comprehensive meta-analysis and systematic review specifically addressing the relationship between ANK3 variants and multiple psychiatric disorders, prior studies have performed meta-analyses focused on individual SNPs—such as rs10994336 in bipolar disorder.31 Compared to these, our study incorporates a broader scope of psychiatric phenotypes, includes a wider range of population data, and adopts stricter criteria for population-level meta-analysis. Specifically, we argue that meta-analysis across populations should only be conducted when consistent significance is observed in each population, as combining heterogeneous signals risks generating false positives.
A key strength of this study lies in the integration of multi-layered QTL analyses to functionally annotate SNPs identified through meta-analysis. By leveraging publicly available datasets from the eQTL Catalogue and GTEx v10, we systematically assessed the impact of disease-associated ANK3 variants on gene expression (eQTL), transcript splicing (sQTL), and 3′UTR processing (apaQTL) in brain tissues and neuron-enriched cell types. This functional annotation framework, which has been largely absent in prior ANK3-focused genetic studies, enabled the identification of regulatory effects for several intronic SNPs. Nonetheless, our study has limitations. We did not perform genotype-stratified analyses, which could refine population-specific effects, nor did we incorporate epigenetic QTLs—such as methylation QTLs—that may modulate ANK3 expression. For instance, rs10994336 has been previously linked to ANK3 methylation in brain tissue. Future studies integrating epigenetic data and expanding functional validation may offer a more comprehensive understanding of the regulatory networks through which ANK3 contributes to psychiatric disease risk.
Ethical considerations
This study was based entirely on publicly available summary-level genomic data (e.g., GWAS, QTL datasets), and does not involve any human participants, animal experiments, or the use of identifiable personal information. Therefore, no ethical approval or informed consent was required. All data used comply with relevant institutional and national guidelines for research integrity and data usage.
TG gratefully acknowledges the invaluable guidance and support provided by his supervisors and research team throughout the course of his PhD studies. TG also extends his sincere appreciation to the China Scholarship Council for their financial support.
2.
Zhou DX, Lambert S, Malen PL, et al.:
Ankyring is required for clustering of voltage-gated Na channels at axon initial segments and for normal action potential firing.
Mol. Biol. Cell.
1998; 143: 1295–1304. Publisher Full Text
3.
Bird KM, Jenkins PM:
Regulation of neuronal ankyrin localization and function by post-translational modifications.
Biochem. Soc. T.
2025; 53(2): 497–507. PubMed Abstract
| Publisher Full Text
| Free Full Text
5.
Prata DP, Costa-Neves B, Cosme G, et al.:
Unravelling the genetic basis of schizophrenia and bipolar disorder with GWAS: A systematic review.
J. Psychiatr. Res.
2019; 114: 178–207. PubMed Abstract
| Publisher Full Text
6.
Li M, Li T, Xiao X, et al.:
Phenotypes, mechanisms and therapeutics: insights from bipolar disorder GWAS findings.
Mol. Psychiatry.
2022; 27(7): 2927–2939. PubMed Abstract
| Publisher Full Text
7.
Jamann TM, Balint-Kurti PJ, Holland JB:
QTL mapping using high-throughput sequencing.
Methods Mol. Biol.
2015; 1284: 257–285. Publisher Full Text
9.
Stang A:
Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses.
Eur. J. Epidemiol.
2010; 25(9): 603–605. PubMed Abstract
| Publisher Full Text
10.
Guo T, Deinhardt K, Wang Y:
Extended data Table 1. Extracted dataset for the meta-analysis of psychiatric disorder-associated ANK3 SNPs. Dataset.
figshare.
2025.Publisher Full Text
11.
Bowden J, Smith GD, Burgess S:
Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression.
Int. J. Epidemiol.
2015; 44(2): 512–525. PubMed Abstract
| Publisher Full Text
| Free Full Text
12.
Cumpston M, Li T, Page MJ, et al.:
Updated guidance for trusted systematic reviews: a new edition of the Cochrane Handbook for Systematic Reviews of Interventions.
Cochrane Database Syst. Rev.
2019; 10(10): ED000142. PubMed Abstract
| Publisher Full Text
14.
Kerimov N, Alasoo K:
eQTL Catalogue: A Compendium of Uniformly Processed Human Gene Expression and Splicing QTLs.
Hum. Hered.
2020; 84(4-5): 204–205.
16.
Yang L, Wang P, Chen J:
2dGBH: Two-dimensional group Benjamini-Hochberg procedure for false discovery rate control in two-way multiple testing of genomic data.
Bioinformatics.
2024; 40(2). PubMed Abstract
| Publisher Full Text
| Free Full Text
17.
Guo T, Deinhardt K, Wang Y:
Extended data Table 2. Characteristics of the studies included. Dataset.
figshare.
2025.Publisher Full Text
18.
Guo T, Deinhardt K, Wang Y:
Extended data Table 3. Summary of results of meta-analysis and synthetic review of the risk of ANK3 SNPs with psychiatric disorders. Dataset.
figshare.
2025.Publisher Full Text
19.
Guo T, Deinhardt K, Wang Y:
Extended data Table 4. Complete eQTL analysis results for ANK3 SNPs in human brain tissues. Dataset.
figshare.
2025.Publisher Full Text
20.
Guo T, Deinhardt K, Wang Y:
Extended data Table 5. Complete apaQTL analysis results for ANK3 SNPs in human brain tissues. Dataset.
figshare.
2025.Publisher Full Text
21.
Guo T, Deinhardt K, Wang Y:
Extended data Table 6. Complete sQTL analysis results for ANK3 SNPs in human brain tissues. Dataset.
figshare.
2025.Publisher Full Text
22.
Jenkins PM, Kim N, Jones SL, et al.:
Giant ankyrin-G: a critical innovation in vertebrate evolution of fast and integrated neuronal signaling.
Proc. Natl. Acad. Sci. USA.
2015; 112(4): 957–964. PubMed Abstract
| Publisher Full Text
| Free Full Text
23.
Zhang D, Lu W, Zhuo Z, et al.:
Construction of a breast cancer prognosis model based on alternative splicing and immune infiltration.
Discov. Oncol.
2022; 13(1): 78. PubMed Abstract
| Publisher Full Text
| Free Full Text
24.
Rueckert EH, Barker D, Ruderfer D, et al.:
Cis-acting regulation of brain-specific ANK3 gene expression by a genetic variant associated with bipolar disorder.
Mol. Psychiatry.
2013; 18(8): 922–929. PubMed Abstract
| Publisher Full Text
| Free Full Text
25.
Edinger RS, Coronnello C, Bodnar AJ, et al.:
Aldosterone regulates microRNAs in the cortical collecting duct to alter sodium transport.
J. Am. Soc. Nephrol.
2014; 25(11): 2445–2457. PubMed Abstract
| Publisher Full Text
| Free Full Text
26.
Foster MW, Sharp RR:
Beyond race: towards a whole-genome perspective on human populations and genetic variation.
Nat. Rev. Genet.
2004; 5(10): 790–796. PubMed Abstract
| Publisher Full Text
27.
Bamshad MJ, Wooding S, Watkins WS, et al.:
Human population genetic structure and inference of group membership.
Am. J. Hum. Genet.
2003; 72(3): 578–589. PubMed Abstract
| Publisher Full Text
| Free Full Text
30.
Tang L, Liu J, Zhu Y, et al.:
ANK3 Gene Polymorphism Rs10994336 Influences Executive Functions by Modulating Methylation in Patients With Bipolar Disorder.
Front. Neurosci.
2021; 15: 682873. PubMed Abstract
| Publisher Full Text
| Free Full Text
31.
Roby Y:
ANK3 gene polymorphisms and bipolar disorder: a meta-analysis.
Psychiatr. Genet.
2017; 27(6): 225–235. PubMed Abstract
| Publisher Full Text
32.
Dedman A, McQuillin A, Kandaswamy R, et al.:
Sequencing of the ANKYRIN 3 gene (ANK3) encoding ankyrin G in bipolar disorder reveals a non-conservative amino acid change in a short isoform of ankyrin G.
Am. J. Med. Genet. B Neuropsychiatr. Genet.
2012; 159b(3): 328–335. Publisher Full Text
33.
Takata A, Kim SH, Ozaki N, et al.:
Association of ANK3 with bipolar disorder confirmed in East Asia.
Am. J. Med. Genet. B Neuropsychiatr. Genet.
2011; 156b(3): 312–315. PubMed Abstract
| Publisher Full Text
34.
Yuan A, Yi Z, Wang Q, et al.:
ANK3 as a risk gene for schizophrenia: new data in Han Chinese and meta analysis.
Am. J. Med. Genet. B Neuropsychiatr. Genet.
2012; 159B: 997–1005. Publisher Full Text
35.
Cho CH, Kim S, Geum D, et al.:
Association analysis of ANK3 variants with bipolar disorder in the Korean population.
Nord. J. Psychiatry.
2017; 71(4): 245–249. PubMed Abstract
| Publisher Full Text
36.
Zhang C, Cai J, Zhang J, et al.:
Genetic modulation of working memory deficits by ankyrin 3 gene in schizophrenia.
Prog. Neuro-Psychopharmacol. Biol. Psychiatry.
2014; 50: 110–115. PubMed Abstract
| Publisher Full Text
37.
Green EK, Hamshere M, Forty L, et al.:
Replication of bipolar disorder susceptibility alleles and identification of two novel genome-wide significant associations in a new bipolar disorder case-control sample.
Mol. Psychiatry.
2013; 18(12): 1302–1307. PubMed Abstract
| Publisher Full Text
| Free Full Text
38.
Smith EN, Bloss CS, Badner JA, et al.:
Genome-wide association study of bipolar disorder in European American and African American individuals.
Mol. Psychiatry.
2009; 14(8): 755–763. PubMed Abstract
| Publisher Full Text
| Free Full Text
39.
Fiorentino A, O’Brien NL, Locke DP, et al.:
Analysis of ANK3 and CACNA1C variants identified in bipolar disorder whole genome sequence data.
Bipolar Disord.
2014; 16(6): 583–591. PubMed Abstract
| Publisher Full Text
| Free Full Text
41.
Hori H, Yamamoto N, Teraishi T, et al.:
Cognitive effects of the ANK3 risk variants in patients with bipolar disorder and healthy individuals.
J. Affect. Disord.
2014; 158: 90–96. PubMed Abstract
| Publisher Full Text
42.
Kondo K, Ikeda M, Kajio Y, et al.:
Genetic variants on 3q21 and in the Sp8 transcription factor gene (SP8) as susceptibility loci for psychotic disorders: a genetic association study.
PLoS One.
2013; 8(8): e70964. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Athanasiu L, Mattingsdal M, Kähler AK, et al.:
Gene variants associated with schizophrenia in a Norwegian genome-wide study are replicated in a large European cohort.
J. Psychiatr. Res.
2010; 44(12): 748–753. PubMed Abstract
| Publisher Full Text
| Free Full Text
44.
Lee MT, Chen CH, Lee CS, et al.:
Genome-wide association study of bipolar I disorder in the Han Chinese population.
Mol. Psychiatry.
2011; 16(5): 548–556. PubMed Abstract
| Publisher Full Text
45.
Lim CH, Zain SM, Reynolds GP, et al.:
Genetic association of LMAN2L gene in schizophrenia and bipolar disorder and its interaction with ANK3 gene polymorphism.
Prog. Neuro-Psychopharmacol. Biol. Psychiatry.
2014; 54: 157–162. PubMed Abstract
| Publisher Full Text
46.
Ferreira MA, O’Donovan MC, Meng YA, et al.:
Collaborative genome-wide association analysis supports a role for ANK3 and CACNA1C in bipolar disorder.
Nat. Genet.
2008; 40(9): 1056–1058. PubMed Abstract
| Publisher Full Text
| Free Full Text
47.
Tesli M, Koefoed P, Athanasiu L, et al.:
Association analysis of ANK3 gene variants in nordic bipolar disorder and schizophrenia case-control samples.
Am. J. Med. Genet. B Neuropsychiatr. Genet.
2011; 156b(8): 969–974. PubMed Abstract
| Publisher Full Text
48.
Logue MW, Solovieff N, Leussis MP, et al.:
The ankyrin-3 gene is associated with posttraumatic stress disorder and externalizing comorbidity.
Psychoneuroendocrinology.
2013; 38(10): 2249–2257. PubMed Abstract
| Publisher Full Text
| Free Full Text
49.
Sklar D, Chang B, Hoffman BD:
Commentary: experience with resident unions at one institution and implications for the future of practicing physicians.
Acad. Med.
2011; 86(5): 552–554. PubMed Abstract
| Publisher Full Text
50.
Dashtban S, Haj-Nasrolah-Fard F, Kosari Z, et al.:
ANK3 and ZNF804A intronic variants increase risk of schizophrenia in Iranian population: An association study.
Gene Rep.
2022; 26: 101511. Publisher Full Text
51.
Karimian SS, Akbari MT, Sadr SS, et al.:
Association of Candidate Single Nucleotide Polymorphisms Related to Candidate Genes in Patients With Schizophrenia.
Basic Clin. Neurosci.
2020; 11(5): 595–608. PubMed Abstract
| Publisher Full Text
| Free Full Text
52.
Scott LJ, Muglia P, Kong XQ, et al.:
Genome-wide association and meta-analysis of bipolar disorder in individuals of European ancestry.
Proc. Natl. Acad. Sci. USA.
2009; 106(18): 7501–7506. PubMed Abstract
| Publisher Full Text
| Free Full Text
53.
Schulze TG, Detera-Wadleigh SD, Akula N, et al.:
Two variants in Ankyrin 3 (ANK3) are independent genetic risk factors for bipolar disorder.
Mol. Psychiatry.
2009; 14(5): 487–491. PubMed Abstract
| Publisher Full Text
| Free Full Text
54.
Frazier TW, Youngstrom EA, Frankel BA, et al.:
Candidate gene associations with mood disorder, cognitive vulnerability, and fronto-limbic volumes.
Brain Behav.
2014; 4(3): 418–430. PubMed Abstract
| Publisher Full Text
| Free Full Text
55.
Mühleisen TW, Leber M, Schulze TG, et al.:
Genome-wide association study reveals two new risk loci for bipolar disorder.
Nat. Commun.
2014; 5: 3339. PubMed Abstract
| Publisher Full Text
56.
Guo X, Zhang Y, Du J, et al.:
Association analysis of ANK3 gene variants with schizophrenia in a northern Chinese Han population.
Oncotarget.
2016; 7(52): 85888–85894. PubMed Abstract
| Publisher Full Text
| Free Full Text
57.
Zhong X, Zhang L, Han S, et al.:
Case control study of association between the ANK3 rs10761482 polymorphism and schizophrenia in persons of Uyghur nationality living in Xinjiang China.
Shanghai Arch. Psychiatry.
2014; 26(5): 288–293. PubMed Abstract
| Publisher Full Text
58.
Cheng Y, Xu J, Dong C, et al.:
Age-related atrophy of cortical thickness and genetic effect of ANK3 gene in first episode MDD patients.
Neuroimage Clin.
2020; 28: 102384. PubMed Abstract
| Publisher Full Text
| Free Full Text
1
Biological Sciences, Faculty of Environmental and Life Sciences, University of Southampton, Southampton, SO17 1BJ, UK 2
University of Bremen, Cell Biology, Bremen, 28359, Germany 3
Institute for Life Sciences, University of Southampton, Southampton, SO17 1BJ, UK 4
NIHR Southampton Biomedical Research Centre, University Hospital Southampton, Southampton, England, SO16 6YD, UK
Tao Guo
Roles:
Data Curation,
Formal Analysis,
Investigation,
Methodology,
Software,
Validation,
Visualization,
Writing – Original Draft Preparation,
Writing – Review & Editing
Guo T, Deinhardt K and Wang Y. Single Nucleotide Polymorphisms in ANK3 and Psychiatric Risk: A Meta-Analysis, Systematic Review, and Quantitative Trait Locus Insights [version 1; peer review: 1 approved with reservations]. F1000Research 2025, 14:1020 (https://doi.org/10.12688/f1000research.169014.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Sengupta D. Reviewer Report For: Single Nucleotide Polymorphisms in ANK3 and Psychiatric Risk: A Meta-Analysis, Systematic Review, and Quantitative Trait Locus Insights [version 1; peer review: 1 approved with reservations]. F1000Research 2025, 14:1020 (https://doi.org/10.5256/f1000research.186294.r450176)
The title is comprehensive and accurately reflects the study scope, incorporating three key methodological components (meta-analysis, systematic review, QTL). The abstract is well-structured with distinct Background, Objectives, Methods,
... Continue reading
TITLE & ABSTRACT
Summary Assessment
The title is comprehensive and accurately reflects the study scope, incorporating three key methodological components (meta-analysis, systematic review, QTL). The abstract is well-structured with distinct Background, Objectives, Methods, Results, and Conclusion sections; however, specific methodological detail is limited, and key statistical parameters lack precision in reporting.
Major Issues
Issue 1: Abstract reporting lacks methodological specificity
- Description: The abstract states "A meta-analysis and systematic literature review were conducted" but omits critical methodological details essential for evaluating reproducibility: databases searched, search date, number of studies included in meta-analysis, inclusion/exclusion criteria summary, and specific genetic models analyzed (allelic, dominant, recessive, additive).
- Location: Abstract, Methods section (lines omitted in pagination)
- Impact on Validity: Readers cannot assess completeness of evidence synthesis or replicate the search strategy; may underestimate heterogeneity or methodological rigor.
- Required Author Action: Expand abstract Methods to include: (1) databases and date range; (2) number of studies analyzed in meta-analysis vs. synthetic review; (3) study design types; (4) genetic models assessed (mandatory for genetic association meta-analyses per PRISMA 2020 item S4); (5) population stratification approach.
Issue 2: Abstract Results section lacks effect estimate precision and comparative context
- Description: Multiple SNP-disease associations are reported with ORs and P-values, but the abstract conflates highly significant associations (rs1938526: P = 7.26×10⁻¹²) with nominally significant findings (three PTSD SNPs: P = 0.045). No mention of which findings met the FDR correction threshold vs. nominal significance only, creating ambiguity about robust vs. suggestive signals.
- Location: Abstract, Results section
- Impact on Validity: Readers may overinterpret nominal findings as definitively associated; the abstract does not clearly distinguish between FDR-significant (sQTL findings) and nominal-only associations (eQTL, apaQTL).
- Required Author Action: Clarify abstract Results to distinguish effect tiers: (1) FDR-significant meta-analyses; (2) FDR-significant QTL findings; (3) nominal-only findings. Specify that PTSD SNPs are based on single studies or small sample meta-analyses.
Issue 3: Abstract Keywords underrepresent study scope
- Description: Keywords list "ANK3, psychiatric disorders, bipolar disorder, schizophrenia, major depression, post-traumatic stress disorder, single nucleotide polymorphism, QTL analysis" but omits critical descriptors: "meta-analysis," "systematic review," "gene expression," "alternative splicing," "population stratification," "Hardy-Weinberg equilibrium."
- Location: Keywords section
- Impact on Validity: Reduced discoverability and indexing specificity in PubMed/Web of Science; readers searching for meta-analysis methodology or gene expression QTL studies may miss this work.
- Required Author Action: Expand keywords to 8–12 terms: Add "meta-analysis," "systematic review," "eQTL," "splicing QTL," "population stratification," "Hardy-Weinberg equilibrium." Consider adding "psychiatric genetics" or "neuropsychiatric risk."
Minor Issues & Editorial Suggestions
1. Title hyphenation: Title reads "Single Nucleotide Polymorphisms in ANK3 and Psychiatric Risk: A Meta-Analysis, Systematic Review, and Quantitative Trait Locus Insights." For enhanced clarity and brevity, consider: "ANK3 Genetic Variants and Psychiatric Disorder Risk: Meta-Analysis, Systematic Review, and Functional QTL Characterization."
2. Abstract structure clarity: Separate "Objectives" as a distinct paragraph; currently merged with Background, reducing clarity.
Suggested Additional References
For strengthening abstract methodology reporting, consider citing:
- Moher D, Shamseer L, Clarke M, et al. Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols (PRISMA-P) 2015 statement. Syst Rev. 2015;4:1. [PRISMA 2020 extension for transparent reporting of genetic association meta-analyses]
- Stroup DF, Berlin JA, Moher D, et al. Meta-analysis of Observational Studies in Epidemiology (MOOSE) guidelines. JAMA. 2000;283(15):2008–2012. [Standard for observational genetic studies]
INTRODUCTION
Summary Assessment
The Introduction is well-motivated and clearly establishes the biological significance of ANK3 in neuronal signaling and its implicated role in multiple psychiatric disorders. The rationale for systematic integration of meta-analysis with QTL analysis is compelling. However, the section lacks an explicit statement of study objectives, and the transition between motivating background and study-specific aims could be more precise. PRISMA 2020 item 5 (PICO specification) is incomplete.
Major Issues
Issue 1: Study Objectives and PICO framework are not explicitly defined
- Description: The Introduction states "To address this gap, the present study integrates meta-analysis with QTL-based transcriptomic analysis..." but does not formally articulate the study's specific objectives in structured PICO format (Population, Intervention/Exposure, Comparison, Outcome). PRISMA 2020 mandates explicit PICO specification in the Introduction or Methods.
- Location: Introduction, final paragraphs (page 3)
- Impact on Validity: Without an explicit PICO definition, readers cannot determine whether specific population strata, diagnostic criteria, or outcome measures were predefined or post-hoc. Critical for assessing risk of selective outcome reporting.
- Required Author Action: Add explicit PICO statement before the Methods section or at the end of Introduction:
- Population: Individuals (case and control) with psychiatric disorder diagnoses (bipolar disorder, schizophrenia, major depressive disorder, post-traumatic stress disorder); across diverse populations (European, North American, Asian).
- Exposure: ANK3 single-nucleotide polymorphisms (SNPs); genetic variants in intronic or exonic regions.
- Comparison: Case vs. control groups matched by population.
- Outcomes: Primary: odds ratio (OR) of psychiatric disorder risk per ANK3 SNP; Secondary: tissue-specific regulatory effects (eQTL, apaQTL, sQTL) in brain tissues.
Issue 2: Insufficient detail on prior inconsistencies in GWAS findings
- Description: The Introduction notes "considerable inconsistencies remain regarding which ANK3 SNPs are most robustly associated with psychiatric disorders and whether these associations are consistent across diverse populations," but does not cite specific conflicting findings or quantify the extent of replication failure.
- Location: Introduction, page 2, paragraph 4
- Impact on Validity: Readers cannot assess whether prior conflicting results reflect genuine population stratification, genotyping artifact, study quality variation, or statistical chance. Limits the understanding of study significance.
- Required Author Action: Insert a table (or extended text) summarizing 3–5 prior ANK3 GWAS findings showing conflicting results, e.g., "rs10994336 was significant for bipolar disorder in European cohorts (Ferreira et al., 2008; Scott et al., 2009) but non-significant in East Asian populations (Takata et al., 2011; Lee et al., 2011), suggesting possible population-specific genetic architecture." This grounds the need for population-stratified meta-analysis.
Issue 3: Alternative splicing and ANK3 isoforms are not discussed in the Introduction
- Description: The Introduction briefly mentions ANK3 isoforms ("expressed as multiple isoforms arising from alternative splicing," page 2) but does not elaborate on isoform-specific subcellular localization or functional differences, limiting reader understanding of why splicing QTL findings are biologically critical.
- Location: Introduction, pages 2–3
- Impact on Validity: Readers unfamiliar with ANK3 biology will not appreciate the mechanistic importance of the sQTL results presented in the Results section (particularly regarding exon 37 inclusion affecting AIS localization). This weakens the Discussion's functional interpretation.
- Required Author Action: Expand the isoform description: "ANK3 encodes multiple protein isoforms ranging from ~190 kDa (postsynaptic form) to 480 kDa (AIS-localized form), differing in their N-terminal and C-terminal regions. The larger isoforms are critical for axon initial segment (AIS) organization, clustering voltage-gated sodium channels (Nav), and cell adhesion molecules (βIV-spectrin, neurofascin) to stabilize neuronal polarity and action potential generation. Loss-of-function mutations or splice variants affecting AIS-localized isoforms compromise neuronal excitability and have been implicated in neurodevelopmental and psychiatric phenotypes."
Minor Issues & Editorial Suggestions
1. Reference formatting inconsistency: References are cited numerically (e.g., "ANK3 variants have been implicated in several neuropsychiatric disorders.⁴") but several lack complete bibliographic details in the main text. Verify that all numbered citations in the Introduction correspond to entries in the References section (cross-check with Ref. 1–7).
2. Terminology precision: The term "noncoding regulatory elements" (page 3) is broad and could refer to enhancers, silencers, miRNA binding sites, or splicing regulatory elements. Consider specifying: "intronic enhancer elements or splicing regulatory motifs" based on QTL findings.
3. QTL methodology preview: The Introduction briefly mentions "expression quantitative trait locus (eQTL)," "alternative polyadenylation QTL (apaQTL)," and "splice QTL (sQTL)" analyses, but does not explain what these measure or why they are complementary. A 2–3 sentence explanation would strengthen reader comprehension, e.g., "eQTL studies quantify the effect of genetic variants on steady-state gene expression levels; sQTL analyses assess effects on splicing patterns; apaQTL studies measure effects on alternative transcript 3′-end polyadenylation. Together, these approaches characterize both quantitative and qualitative transcriptomic impacts of disease-associated variants."
MATERIALS & METHODS
Summary Assessment
The Methods section provides a reasonable structural overview of the meta-analysis workflow and QTL analysis approach; however, critical methodological details are incomplete or imprecisely reported. Search strategy is inadequately documented, risk-of-bias assessment lacks detail, heterogeneity assumptions are not justified, and statistical model choices (fixed vs. random effects) are not explained. PRISMA 2020 items 6–11 are partially compliant.
Major Issues
Issue 1: Literature search strategy is incompletely reported (PRISMA 2020 item 7 non-compliant)
- Description: The Methods states "two independent researchers (TG and YW) systematically searched PubMed, Web of Science, and MEDLINE using combinations of the terms 'genome-wide association study' with 'ANK3', as well as 'nucleotide polymorphisms' with 'ankyrin 3' or 'Ankyrin-G'." No Boolean operators specified, no search date provided, no search strategy documentation (e.g., MeSH terms, truncation, field tags), and no justification for the specific database selection or exclusion of other major databases (e.g., Embase, Cochrane Central Register of Controlled Trials).
- Location: Methods, "Literature retrieval and selection" section, page 3
- Impact on Validity: Readers cannot reproduce the search, assess whether important studies were missed, or determine if publication bias is likely (missing studies in non-indexed sources). Transparency is compromised; reproducibility cannot be independently verified.
- Required Author Action: Provide a detailed search strategy meeting PRISMA 2020 item 7:
- Specify exact date range of searches (e.g., "searches conducted from 1 January 2000 through 30 September 2025")
- Report complete search strings for each database:
- PubMed: `("ANK3" OR "ankyrin 3" OR "ankyrin-3" OR "ankyrin-G" OR "ANKRD4") AND ("single nucleotide polymorphism" OR "SNP" OR "rs[0-9]{7}" OR "polymorphism" OR "genotype" OR "allele") AND ("bipolar disorder" OR "schizophrenia" OR "depression" OR "PTSD" OR "psychiatric disorder")`
- Web of Science: [with appropriate field tags: TS= (topic search)]
- EMBASE (if not already searched): [with EMTREE keywords and field codes]
- Justify database selection (e.g., "PubMed for broad coverage of life sciences, Web of Science for citation tracking and multidisciplinary coverage, MEDLINE (NLM) indexing")
- Document supplementary searches (reference harvesting, contact with authors, trial registry searches)
- Provide PRISMA-S (Search) reporting form documenting search filters applied (peer-review status, publication language, study design filters)
Issue 2: Newcastle-Ottawa Scale assessment is inadequately reported; itemized scoring is unavailable
- Description: Methods mention "The studies with controlled trials were evaluated by the Newcastle-Ottawa Scale (NOS)⁸'⁹ by two different researchers independently. NOS includes 1. Selection... [partial list]... The total score is 9, and if a study can get over 5, it can be assessed as good quality. Those articles that did not meet the quality requirement were excluded." However, no itemized NOS scores are reported for each included study, precluding verification of quality assessments and identification of potential quality-based bias. The threshold of ≥5/9 is not justified.
- Location: Methods, "Quality assessment of included studies" section, page 3
- Impact on Validity: High risk of quality bias; readers cannot assess whether lower-quality studies were differentially included/excluded or whether risk-of-bias patterns differ between psychiatric disorders. Inability to conduct sensitivity analysis by study quality.
- Required Author Action:
- Create supplementary table (Appendix or Extended data) listing all 27 included studies with complete NOS itemized scores across all 8 categories (Selection: 4 items; Comparability: 1 item; Outcome: 3 items) and total score
- Justify ≥5/9 threshold with reference to NLM/Cochrane guidance (note: NOS maximum is typically 9 points; confirm scoring system used)
- Conduct sensitivity analysis excluding low-quality studies (NOS <5) and report whether meta-analysis effect estimates are stable
- Report inter-rater agreement (Cohen's κ) between TG and YW for NOS assessments
- Disclose whether specific NOS domains (e.g., Selection of cases, Selection of controls) correlated with effect size heterogeneity
Issue 3: Meta-analysis model selection is not justified; fixed vs. random effects are not discussed
- Description: Methods state: "Utilizing the 'meta (version 4.18-0)' package within R 4.0.1 and RStudio, we conducted a meta-analysis on the specified SNPs. Odds ratios (OR) and 95% confidence intervals (95%CI) were input to yield synthesized outcomes." No specification of whether fixed-effect or random-effects models were used, no justification for model choice, and no reporting of I² heterogeneity statistics or τ² (tau-squared) estimates in the Methods (though these appear in Results).
- Location: Methods, "Synthesis and analysis of data and assessment of publication bias" section, page 4
- Impact on Validity: Model choice affects confidence intervals and P-values critically; random-effects models produce wider CIs and lower P-values than fixed-effect models. Absence of pre-specification suggests possible post-hoc model selection driven by desired results. Standard guidance (Cochrane Handbook 2021) recommends random-effects for anticipated heterogeneity.
- Required Author Action: Add an explicit statement to Methods:
- "We employed random-effects meta-analysis models using restricted maximum likelihood (REML) estimation as default, as we anticipated heterogeneity across studies due to differences in population ancestry, study design, diagnostic criteria, and genotyping platforms. For each SNP-disorder association with fewer than 5 studies (e.g., rs139972937 for bipolar disorder), we reported individual study estimates and effect directions but did not conduct quantitative meta-analysis due to limited power to estimate τ². Heterogeneity was quantified using I² (percentage of variance due to heterogeneity) and τ² (between-study variance). We report 95% prediction intervals (PI) to characterize the range of effects expected in future populations. If I² ≤50%, we also report fixed-effect estimates for comparison."
- Specify software defaults: If using R meta package, document whether tau.common=FALSE (default REML)
- Clarify whether genetic models (allelic, dominant, recessive) were specified a priori or determined post-hoc based on data fit
Issue 4: Handling of genetic models, allele coding, and population stratification is not transparent
- Description: Methods state "Risk estimates were extracted in the form of odds ratios (OR), while SNP relationships were assessed using either minor allele frequency (MAF) or specific genotype distributions. For cohorts stratified by region, meta-analysis was performed only when a consistent direction of effect was observed across all subgroups." However, the Methods does not specify: (a) which genetic models were extracted/analyzed for each SNP (allelic, dominant, recessive, additive/genotype contrast), (b) how studies reporting different genetic models were harmonized, (c) how studies reporting MAF-based analysis were converted to genotype-based OR, and (d) what constitutes "consistent direction of effect" (e.g., all subgroups protective, all subgroups risk-increasing).
- Location: Methods, "Criteria for inclusion" and "Synthesis and analysis of data" sections, pages 3–4
- Impact on Validity: Standard genetic association best practices (including PLINK, GWAS-MA guidelines) mandate transparency in genetic model specification to avoid selective reporting of significant models. Pooling different genetic models without documentation risks bias. Population stratification heterogeneity may reflect genuine allelic heterogeneity vs. linkage disequilibrium differences; undocumented handling obscures this.
- Required Author Action:
- Create supplementary table extracting for each SNP-disorder pair: (1) genetic models reported (allelic, dominant, recessive, additive), (2) per-study effect estimates for each model if available, (3) decision criteria for selecting primary model for meta-analysis
- Explicitly define "consistent direction of effect": e.g., "All subgroups with P<0.05 showed protective effects (OR<1)" vs. "≥75% of subgroups showed the same direction, even if nominal significance varied."
- For studies reporting MAF frequencies and case/control counts, show the conversion formula: e.g., "OR = [case_MAF/(1-case_MAF)] / [control_MAF/(1-control_MAF)]."
- Report whether the Hardy-Weinberg equilibrium (HWE) was assessed for each study and population separately. In Methods, clarify HWE threshold used (p<0.05, <0.01, <0.001) and whether studies with HWE violation were excluded or separately analyzed (currently only mentioned in Results for rs10761482)
Issue 5: Publication bias assessment rationale inadequately justified
- Description: Methods state: "We combined the funnel plot and the Egger test¹¹ to assess the publication bias of the meta-analysis. As none of the meta-analyses included 10 or more studies, publication bias was not assessed, in accordance with Cochrane guidance. This is internally contradictory: the statement says publication bias "was not assessed," but earlier claims funnel plot and Egger test "were combined." Additionally, the Cochrane Handbook 2021 states that publication bias assessment is unreliable with <10 studies, but does not recommend abandoning it; instead, contour-enhanced funnel plots and interpretation with caution are advised.
- Location: Methods, "Synthesis and analysis of data and assessment of publication bias" section, page 4
- Impact on Validity: Undermines assessment of potential bias in effect estimates; small study effects (which may reflect publication bias, selective outcome reporting, or true effect heterogeneity) are not characterized. Readers cannot determine whether reported associations may be inflated due to the selective publication of positive findings.
- Required Author Action:
- Clarify that funnel plots were generated for all meta-analyses (even if <10 studies) to visually assess asymmetry
- Report Egger regression test results for all SNP-disorder combinations (provide slope, intercept, P-value, I²)
- Justify the post-hoc decision not to assess publication bias: cite Cochrane Handbook p. XXX or alternative guidance; alternatively, report results with statement of statistical limitations
- Consider alternative publication bias approaches suitable for small samples: trim-and-fill method, or Begg's rank correlation test (less sensitive to heterogeneity than Egger test)
- Create visual funnel plots for Figures/Supplementary Materials
Issue 6: QTL analysis methodology lacks critical details on multiple testing correction and interpretation thresholds
- Description: Methods state: "To correct for multiple testing across tissues and regulatory layers, we applied Benjamini–Hochberg false discovery rate (FDR) correction to the nominal p-values within each QTL modality (eQTL, apaQTL, and sQTL). Associations with FDR < 0.05 were considered statistically significant." However, several important details are missing: (1) How many tissues and cell types were tested per SNP? (2) What constitutes a "tissue" for FDR correction—are brain regions (neocortex, cerebellum, hippocampus, etc.) and cell types (neurons, astrocytes, microglia) counted separately? (3) Was the FDR correction applied within each QTL type separately (as stated) or across all three modalities combined? (4) Nominal P<0.05 is mentioned as a threshold for interpretation of suggestive findings, but no justification is provided for this threshold.
- Location: Methods, "QTL analysis" section, page 4
- Impact on Validity: Transparency of multiple-testing correction directly impacts the reliability of findings. If FDR correction was lenient (applied only within each tissue, not across tissues), more associations would appear significant; if stringent (applied across all tissues and all three modalities), fewer would. The Discussion acknowledges that eQTL and apaQTL signals "did not remain significant following false discovery rate (FDR) correction," suggesting correction was stringent. However, readers cannot assess whether the sQTL FDR=0.010 results are robust.
- Required Author Action:
- State total number of tests: "We tested 10 SNPs × N tissues × 3 regulatory modalities = XXX tests total. FDR correction was applied independently within each regulatory modality (eQTL, apaQTL, sQTL), yielding XXX independent testing families. For eQTL, we tested across XXX tissues/cell types; for apaQTL across XXX tissues; for sQTL across XXX tissues."
- Define tissues explicitly: list all included tissues/cell types (e.g., "neocortex, neurons, microglia, cerebellum, putamen, hippocampus, prefrontal cortex, amygdala" or cite Supplementary Table)
- Justify FDR threshold: "FDR<0.05 was chosen to control the expected proportion of false discoveries at 5%, balancing sensitivity and specificity given the focused, hypothesis-driven design of our analysis (testing 10 pre-selected SNPs identified through meta-analysis, not genome-wide scan)."
- For nominally significant but FDR-nonsignificant findings (eQTL, apaQTL), report effect sizes and consistency across tissues to supportthe biological plausibility claim in the Discussion
Issue 7: Python and Bash script details are insufficient for reproducibility
- Description: Methods state: "All datasets were parsed and harmonised by rsID using custom Python 3.11 and Bash scripts." No mention of script availability, algorithm details, or validation procedures. This is critical for QTL data curation.
- Location: Methods, "QTL analysis" section, page 4
- Impact on Validity: Others cannot verify data parsing accuracy, assess whether variants were correctly matched across databases, or reproduce the analysis. Potential for systematic data misalignment errors.
- Required Author Action:
- Provide GitHub repository or Figshare code deposit with annotated scripts and documentation
- Describe parsing algorithm: e.g., "SNPs were matched across eQTL Catalogue and GTEx using exact rsID match (e.g., rs10761482). For multi-allelic SNPs, we retained the major/minor allele designation from GTEx v10 and verified consistency with eQTL Catalogue. X in 10, XXX SNPs could not be uniquely matched and were excluded with details in Supplementary Table X."
- Report validation: e.g., "Randomly selected 50 SNPs (~0.5%) were manually verified against source databases; 100% concordance achieved"
Minor Issues & Editorial Suggestions
1. Criteria for exclusion clarity: Exclusion criteria state "Studies employing hazard ratios (HR) for SNP risk estimation were excluded" without justification. HRs are less common for SNP-disease associations but may appear in prospective cohort studies. Consider clarifying whether prospective study designs were excluded or only HR estimates; if the latter, should HR studies be converted to OR for meta-analysis?
2. Diagnostic criteria specificity: Methods do not specify which diagnostic criteria were required (DSM-5, ICD-10, expert-assessed, self-reported). This affects phenotype harmonization; psychiatric disorders have evolved diagnostic definitions. Recommend: "Included studies were required to use standardized diagnostic criteria (DSM-III-R, DSM-IV, DSM-5, or ICD-10) and to provide data allowing assessment of case definition clarity."
3. Control group definition: Methods state "For each patient cohort, a matched control group comprising healthy individuals from the same population and study was selected," but do not specify: Were control groups screened for absence of psychiatric disorders (negative screening) or merely reported as healthy/normal? This affects the interpretation of effect estimates.
4. Extended Dataset sourcing: Methods reference "Extended dataset 1 (Figshare, https://doi.org/10.6084/m9.figshare.29886620)" but Figshare URLs are not stable; recommend archiving on an institutional repository with a persistent identifier. Verify accessibility at submission time.
5. Gene structure sourcing: Methods cite Ensembl database for exon/intron mapping: "Intron and exon details for human ANK3 were sourced from the Ensembl protein database." Specify Ensembl release version (e.g., Ensembl release 114, GRCh38.p14) for reproducibility and to account for future genome builds.
6. Statistical software citation: Citation of "meta (version 4.18-0)" package is appropriate; however, cite the R package publication: Schwarzer G. meta: An R package for meta-analysis. R News. 2007;7(3):40–45. Similarly, specify GraphPad Prism version and matplotlib version.
RESULTS
Summary Assessment
The Results section is well-organized, presenting study selection flow, quality assessment, meta-analysis findings by psychiatric disorder, SNP localization, and QTL analyses. However, critical numerical details are missing: forest plots lack heterogeneity statistics (I², τ²), effect estimates for some SNPs/populations are incomplete, and QTL tissue selection is not justified. Data tables are presented, but summaries lack precision.
Major Issues
Issue 1: Heterogeneity statistics (I², τ²) not reported for meta-analysis results
- Description: Results section reports meta-analysis point estimates and P-values (e.g., "rs10994336... emerged as a significant risk locus (OR = 1.28, 95% CI: 1.13–1.45, P = 1.04×10⁻⁴)") but does not report I² (proportion of variance due to heterogeneity) or τ² (between-study variance) for any SNP-disorder combination. Figures 2–5 show forest plots but lack visual or tabular heterogeneity reporting. This is a standard requirement of meta-analysis reporting (PRISMA 2020 item 20).
- Location: Results section, pages 5–10 (Meta-analysis sections for each disorder)
- Impact on Validity: Readers cannot assess the degree of heterogeneity, identify whether the fixed-effect model assumption (homogeneity) is justified, or determine whether effect estimates are generalizable across populations. Large heterogeneity suggests true effect variation due to population genetics, diagnostic criteria, or genotyping methods; small heterogeneity suggests consistent effects. This distinction is critical for interpretation.
- Required Author Action:
- For each SNP-disorder combination analyzed quantitatively, report: I², 95% CI for I² (if available from meta package), τ², Q-test P-value
- Add column to Results tables (if created) or supplementary table with heterogeneity statistics
- For forest plots (Figures 2–5, 6), add text overlay showing I² and P(heterogeneity), or create a companion table
- Example revision for rs10994336 in bipolar disorder: "Rs10994336, included in nine studies from European and North American populations, emerged as a significant risk locus (OR = 1.28, 95% CI: 1.13–1.45, P = 1.04×10⁻⁴, I² = XX%, τ² = XX). Heterogeneity was [low/moderate/high], suggesting [consistent/variable] effects across populations."
Issue 2: Study selection Flow diagram (Figure 1) lacks detail; numbers do not reconcile
- Description: Figure 1 shows the literature screening process: "1,140 articles retrieved → [screening] → Full-text review resulted in exclusion of 214 studies → 27 studies included." However, the flow diagram does not show intermediate steps: How many articles were excluded during title/abstract screening? How many underwent full-text review? The caption refers to "For exclusion, please refer to section Criteria for Exclusion," but does not quantify exclusion reasons, which is a PRISMA 2020 item 13 requirement.
- Location: Results, page 5, Figure 1
- Impact on Validity: Readers cannot assess whether screening was systematic or selective. For example, if 900 of 1,140 articles were excluded during abstract screening (likely given search breadth), were they appropriately excluded, or did reviewers prematurely filter? Without exclusion reason tallies, the risk of selection bias cannot be evaluated. PRISMA 2020 mandates a flow diagram showing: (1) Total records identified through searching, (2) duplicate records excluded, (3) records screened, (4) records excluded, (5) full-text articles assessed, (6) reasons for exclusion at full-text stage, (7) studies included in review.
- Required Author Action:
- Revise Figure 1 to include:
- n= records identified through database searching (e.g., PubMed: n=XXX; Web of Science: n=XXX; MEDLINE: n=XXX)
- n= duplicates excluded
- n= records screened (after duplicate removal)
- n= records excluded during title/abstract screening [with top 3–5 exclusion reasons and counts]
- n= full-text articles assessed for eligibility
- n= studies excluded at full-text stage [itemized: e.g., "n= without ANK3 genotype data, n= non-psychiatric outcomes, n= case reports/reviews, etc."]
- n= studies included in meta-analysis
- n= studies included in qualitative synthesis
- Create Supplementary Table detailing exclusion reasons for all 214 full-text rejections (at minimum, top reasons and counts)
Issue 3: Subgroup analysis rationale for rs10761482 HWE stratification is not clearly justified in the Results
- Description: Results state: "To further explore potential population-specific effects, subgroup analysis was conducted for rs10761482 within the Asian population, which was stratified into three subgroups based on Hardy–Weinberg equilibrium (HWE) status and ethnicity. The first subgroup comprised Chinese cohorts not in HWE..., the second subgroup included Iranian participants, and the third subgroup consisted of Chinese cohorts with significant HWE conformity. However, the Results do not explain: (a) What was the predetermined HWE P-value threshold used to classify "in HWE" vs. "not in HWE"? (b) Why were Chinese studies stratified by HWE while Iranian studies were not? (c) How many studies fell into each subgroup (n=?), and were cell counts adequate for meta-analysis? This subgroup decision appears post-hoc, driven by observing HWE violations in data, increasing the risk of cherry-picking.
- Location: Results, page 6, Schizophrenia section
- Impact on Validity: If HWE stratification was not pre-specified, this subgroup analysis inflates Type I error and may reflect data-driven hypothesis generation. The Discussion mentions HWE issues (pages 11–12) but does not clearly state whether the HWE analysis was predefined.
- Required Author Action:
- Clarify in Results: "HWE status was assessed a priori for each included study using the standard χ² goodness-of-fit test (threshold P < 0.05 for HWE deviation). Studies significantly deviating from HWE were stratified separately in subgroup meta-analyses, as recommended by [citation], to investigate potential sources of heterogeneity (e.g., population substructure, genotyping error, selection bias)."
- Report complete stratification: "For rs10761482 in schizophrenia, n=X Chinese studies with HWE conformity (P≥0.05) were meta-analyzed separately; n=Y Chinese studies with HWE deviation (P<0.05) were analyzed separately; n=Z Iranian studies were analyzed separately (HWE status: [report]). Overall meta-analysis combining all Asian studies: [results]. Subgroup-by-ethnicity interaction: [report I² and τ² for between-subgroup heterogeneity]."
Issue 4: PTSD SNP associations based on inadequate sample sizes (single studies or n=3 total)
- Description: Results state: "For PTSD, all three investigated variants—rs28932171, rs11599164, and rs17208576—were found to be significantly associated with reduced risk, each exhibiting the same odds ratio (OR = 0.48) and a nominally significant P-value (P = 0.045)." However, Extended Dataset 3 (not fully visible in the manuscript) likely shows that these three SNPs were reported in only 1–2 studies; meta-analysis with 1 study is meaningless (it is simply the study estimate), and meta-analysis with 2–3 small studies has minimal power. The identical OR=0.48 and P=0.045 for all three variants is also suspicious and suggests these may be from a single study or linkage disequilibrium. This finding is misrepresented in the Abstract as a major result.
- Location: Results, page 7, section "Meta-analysis and synthetic review... PTSD"
- Impact on Validity: Readers may over-interpret these PTSD associations as robust findings when they are actually nominal signals from single studies or small samples. No sensitivity analysis or replication evidence is presented. This weakens the overall conclusions.
- Required Author Action:
- In Results, explicitly report sample sizes per SNP-disorder pair: "For PTSD, meta-analysis of rs28932171 included n=X independent studies (total cases=XXX, controls=XXX); rs11599164 included n=Y studies; rs17208576 included n=Z studies. [Note: If n<3 studies, these are not true meta-analyses but rather synthetic reviews of single or paired studies.]"
- Explain identical OR and P-values across three SNPs: "These three SNPs were derived from [1 study / 2 studies] (authors: [citation]). The identical effect estimates reflect their origin in a single study population and should be interpreted with caution; replication in independent cohorts is required."
- In Discussion, downgrade PTSD findings to "preliminary" or "suggestive" pending replication
Issue 5: QTL analysis, tissue selection, and justification are absent
- Description: QTL analyses state: "eQTL slopes and p values were retrieved from the eQTL Catalogue... apaQTL (alternative polyadenylation) and sQTL (splicing) summary statistics were retrieved from the GTEx v10 resource... For every brain tissue, we extracted the slope (β), its standard error, and the nominal p value." However, which specific tissues and cell types were included is not stated in Methods or Results. For example, did the analysis include all available eQTL Catalogue tissues, or was a priori selection (e.g., psychiatric-relevant tissues: prefrontal cortex, amygdala, nucleus accumbens, hippocampus, anterior cingulate cortex) applied? The eQTL Catalogue contains diverse tissues; arbitrary inclusion may inflate the multiple testing burden and reduce power.
- Location: Results, pages 8–11, QTL analysis sections; Methods page 4
- Impact on Validity: If all available tissues were analyzed without selection, the FDR correction may have been too stringent, and true signals in relevant tissues may be missed. Conversely, if tissue selection was post-hoc (motivated by observing significant results), this increases false discovery risk.
- Required Author Action:
- Provide supplementary table listing: (1) All tissues analyzed for eQTL; (2) All tissues analyzed for apaQTL; (3) All tissues analyzed for sQTL
- State selection criteria: "We extracted data for all available tissues in the eQTL Catalogue and GTEx v10 to comprehensively assess regulatory effects. No a priori tissue selection was performed. FDR correction was applied within each regulatory modality across all tested tissues, accounting for multiple testing."
- OR if tissue selection was applied: "We prioritized brain tissues implicated in psychiatric disorder etiology: prefrontal cortex, amygdala, anterior cingulate cortex, nucleus accumbens, hippocampus, and neuron-enriched cell types. Additional tissues (putamen, cerebellum, etc.) were included if data were available."
Issue 6: Missing data on effect sizes, confidence intervals, and sample sizes for forest plots
- Description: Figures 2–5 show forest plots but do not report: (a) Number of studies per SNP; (b) Total sample size (cases + controls); (c) 95% CI ranges for each included study; (d) Study weights; (e) Heterogeneity statistics. While forest plot visualization is useful, tabular supplementary data are required per PRISMA 2020 for verification and meta-analysis reproducibility.
- Location: Figures 2, 3, 4, 5 (pages 6–7, 10)
- Impact on Validity: Readers cannot verify reported effect estimates, assess which studies contributed most weight, or perform sensitivity analyses.
- Required Author Action:
- Create Supplementary Tables with columns: Author (year), Population, N_cases, N_controls, Genotype/MAF, Effect measure (OR/RR), 95% CI, P-value, Weight (%) in meta-analysis, Study design, Quality (NOS score)
- Ensure forest plot axes and legends clearly label: Effect measure (OR), null value (OR=1.0), CI ranges, study names, and subgroup labels
- Cite table in figure caption: "Forest plot of meta-analysis results; detailed study-level data and heterogeneity statistics provided in Supplementary Table X."
Issue 7: QTL results reporting inconsistency—nominal vs. FDR-significant unclear in text
- Description: QTL results are reported with nominal P-values first, then FDR correction mentioned in parentheses, creating ambiguity. Example: "Rs10761482 demonstrated a notable association with ANK3 expression in both neuron-enriched and neocortical cell populations. In neurons, the variant was associated with increased expression (β = 0.12, SE = 0.06, P = 0.038), although the signal did not remain significant following false discovery rate (FDR) correction (FDR = 0.999)." Readers may misinterpret P=0.038 as significant without reading the parenthetical FDR note. Additionally, FDR=0.999 is extremely conservative and raises questions about the number of tests or the correction method.
- Location: Results, pages 8–11, QTL sections; Tables 2–4
- Impact on Validity: Misinterpretation of statistical significance; readers may cite nominal P-values without recognizing FDR inflation. The extremely high FDR values (0.999, 0.965) suggest liberal multiple testing or lenient FDR control, which should be discussed transparently.
- Required Author Action:
- In Results and Tables 2–4, clearly label findings: "rs10761482 was nominally significant for ANK3 expression in neocortex (P=0.001) but did not meet FDR-adjusted significance threshold (FDR=0.162, n_tissues_tested=XX). In neurons, the association was nominally significant (P=0.038, FDR=0.999), indicating marginal evidence requiring replication."
- Explain high FDR values: "The high FDR-adjusted P-values reflect correction for testing across [XX] tissues and cell types within the eQTL modality. Although nominally significant, these associations should be interpreted as suggestive pending validation in independent cohorts."
- Consider reporting Bonferroni-corrected P-values as a comparison threshold, or provide both for transparency
Minor Issues & Editorial Suggestions
1. Study characteristics table incomplete: Results reference "Extended dataset 2 (https://doi.org/10.6084/m9.figshare.30090913)" containing study characteristics, but recommend including a summary table in the main Results showing: Author (year), Sample size, Population, Psychiatric disorder, Diagnostic criteria, Genotyping platform, Key findings. This aids reader comprehension without requiring external data access.
2. SNP localization table (Table 1) clarity: Table 1 shows exon/intron positions but does not clearly indicate which SNPs are intronic (regulatory) vs. exonic (potentially protein-coding). Recommend adding column: "Location Type: Intronic (regulatory) / Exonic (coding region)." Currently, all appear intronic per the Results text, but the table formatting is ambiguous.
3. Figure 6 gene map resolution: Figure 6 shows the ANK3 gene structure and SNP positions, but lacks a scale bar and precise bp coordinates on axes. For readers to assess SNP clustering, provide: "Map scale: 1 cm = [X] bp" or "Genomic coordinates (GRCh38.p14, NC_000010.11) on x-axis showing ANK3 locus (60.0–60.7 Mb)." The current visualization does not permit readers to discern SNP clustering in the two introns mentioned in the Discussion.
4. Extended Datasets accessibility: Results reference six Extended datasets hosted on Figshare; verify that all DOI links are active and data are publicly accessible (not under embargo). Recommend also depositing on an institutional repository (e.g., University of Southampton Research Portal) with redundant backup.
5. Missing prediction intervals: Random-effects meta-analyses should report 95% prediction intervals (PI) to characterize the range of effects expected in future populations. PRISMA 2020 item 20 recommends PI reporting. Add to Results tables.
DISCUSSION
Summary Assessment
The Discussion provides thoa ughtful interpretation of meta-analysis findings, appropriately contextualizes population-specific heterogeneity, and offers plausible biological mechanisms for intronic SNP effects. However, the Discussion overstates findings regarding mechanistic causality, does not adequately address limitations (only briefly mentioned), and does not discuss implications for clinical practice or future research directions. Limitations acknowledgment is essential for balanced interpretation.
Major Issues
Issue 1: Mechanistic claims overstate functional evidence available from QTL data
- Description: Discussion states: "These findings suggest that the risk variants may influence alternative splicing decisions that determine exon 37 inclusion. Such splice isoform shifts could lead to changes in the intracellular distribution of Ankyrin-G, potentially altering its ability to cluster ion channels at the AIS and thereby affecting neuronal polarity, excitability, or signaling, leading to cellular and circuit-level changes in neuronal signalling, and to disease in the long term." While plausible, this mechanism is speculative. QTL data show association between SNP genotype and splicing patterns (sQTL), not causality. The Discussion does not acknowledge: (a) sQTL associations are correlative; (b) splicing QTL may tag regulatory variants in linkage disequilibrium (not the SNPs themselves); (c) experimental validation (CRISPR perturbation, reporter assays) is required to establish causality.
- Location: Discussion, page 12
- Impact on Validity: Readers may over-interpret QTL associations as causal proof of mechanism, leading to unwarranted biological claims or premature drug development investment targeting these variants. This is especially problematic for intronic variants, which are rarely directly causal but instead mark regulatory regions.
- Required Author Action:
- Temper mechanistic claims: "These findings provide suggestive evidence that the identified SNPs may regulate ANK3 splicing in a brain region–specific manner. The sQTL signal at rs28932171, rs11599164, and rs17208576 in cerebellum suggests that these intronic variants may affect alternative exon 37 inclusion, potentially altering isoform distribution. However, these are correlative associations; functional validation through CRISPR perturbation, reporter assays, or allele-specific expression studies in neural cells would be required to confirm mechanistic causality. Additionally, the identified SNPs may not be the causal variants but rather proxy markers in linkage disequilibrium with unmapped regulatory elements."
- Clarify mechanistic pathway: "Although we cannot infer causality from QTL associations alone, the biological plausibility of our findings is supported by: (1) the documented role of exon 37 in determining AIS-targeted vs. postsynaptic ANK3 isoforms; (2) the functional relevance of AIS ion channel clustering to neuronal excitability; (3) the involvement of AIS dysfunction in neuropsychiatric phenotypes (autism spectrum disorder, epilepsy, ataxia)."
Issue 2: The limitations section is inadequate; key methodological limitations are not discussed
- Description: Discussion briefly states: "Nonetheless, our study has limitations. We did not perform genotype-stratified analyses, which could refine population-specific effects, nor did we incorporate epigenetic QTLs—such as methylation QTLs—that may modulate ANK3 expression." However, several major limitations are not mentioned:
- (a) Meta-analysis sample sizes are small for some SNP-disorder combinations (e.g., PTSD variants from single studies), limiting statistical power and generalizability
- (b) Psychiatric phenotypes may differ across studies in diagnostic criteria, severity, age of onset, subtypes (bipolar I vs. II, schizophrenia subtypes), reducing phenotype homogeneity
- (c) QTL samples are predominantly of European ancestry; regulatory effects may not generalize to non-European populations
- (d) No causal inference framework employed (e.g., Mendelian randomization); associations may be confounded by linkage disequilibrium or population stratification
- (e) Candidate SNP selection was driven by meta-analysis findings, potentially limiting the discovery of additional regulatory variants
- (f) Publication bias was not formally assessed due to the small sample size of the meta-analyses
- Location: Discussion, pages 12–13
- Impact on Validity: Overstated generalizability; readers may not appreciate limitations of effect estimates or applicability to diverse populations. Transparent limitation discussion is essential for balanced interpretation per PRISMA 2020 item 24.
- Required Author Action:
- Expand Limitations section to include:
Our study has several limitations that warrant acknowledgment. First, meta-analysis sample sizes vary considerably across SNP-disorder combinations, from single studies (PTSD variants) to nine studies (rs10994336 in bipolar disorder), limiting statistical power for smaller subgroups and precision of effect estimates. Second, psychiatric phenotypes differed across studies in diagnostic criteria (DSM versions), assessment methods (clinical interview vs. medical record), and sample composition (age, sex, severity), introducing phenotypic heterogeneity that may attenuate effect estimates. Third, the majority of included studies were conducted in European or East Asian populations; regulatory QTL effects (derived predominantly from GTEx v10, which has limited ancestry diversity) may not generalize to African, Latin American, or South Asian populations, limiting the diversity of our sample and the applicability of findings to underrepresented populations. Fourth, we did not employ causal inference methods (e.g., Mendelian randomization, Bayesian colocalization) to distinguish between true causal variants and proxy markers in linkage disequilibrium, limiting mechanistic inference. Fifth, publication bias was not formally assessed using standard methods due to small sample meta-analyses (n<10 studies per SNP-disorder pair); small-study effects or selective reporting of significant findings may bias effect estimates upward. Sixth, the selection of ten SNPs for QTL analysis was based on meta-analysis findings, potentially limiting the discovery of additional regulatory variants not tested in sufficient studies to meet meta-analysis criteria.
Issue 3: Clinical implications and future research directions were not discussed
- Description: Discussion does not address: (a) How these findings might inform psychiatric risk assessment or precision medicine; (b) Whether identified variants might be therapeutic targets; (c) What follow-up experiments are needed to validate findings; (d) Whether findings have implications for psychiatric nosology or endophenotype definition. PRISMA 2020 and editorial best practices recommend discussing clinical/research implications.
- Location: Discussion, pages 12–13
- Impact on Validity: Readers cannot assess practical significance or next steps for the field. Leaves open questions about translational relevance.
- Required Author Action:
- Add paragraph on Clinical/Research Implications: "Clinical implications of these findings remain limited at present. While ANK3 variants are associated with psychiatric disorder risk in the populations studied, effect sizes are modest (ORs~1.1–1.3), and no individual genetic variants are sufficiently predictive to warrant clinical genetic testing in current practice. However, these findings contribute to the polygenic risk architecture of psychiatric disorders and may inform larger risk score models. From a research perspective, the identification of tissue-specific regulatory effects (particularly the robust cerebellar sQTL signals for PTSD variants) suggests that ANK3-mediated splicing alterations in specific brain regions may contribute to psychiatric vulnerability. Future studies should: (1) Validate sQTL and eQTL findings in independent neural cell models (iPSC-derived neurons, primary neural cultures) using CRISPR perturbation or reporter assays; (2) Assess whether ANK3 isoform shifts (particularly affecting AIS-localized vs. postsynaptic forms) are detectable in postmortem psychiatric tissue or neuroimaging-derived neural populations; (3) Determine whether ANK3 expression or isoform usage correlates with psychiatric symptoms or cognitive endophenotypes; (4) Conduct cross-ancestry validation in African, Latin American, and South Asian cohorts to assess effect heterogeneity and generalizability."
Issue 4: Prior ANK3 meta-analyses were not comprehensively compared
- Description: Discussion notes: "Although there is no previous comprehensive meta-analysis and systematic review specifically addressing the relationship between ANK3 variants and multiple psychiatric disorders, prior studies have performed meta-analyses focused on individual SNPs—such as rs10994336 in bipolar disorder." However, no detailed comparison of current findings with the prior rs10994336 meta-analysis (Roby, 2017) is provided. For example, do effect estimates differ, and if so, why? Are confidence intervals narrower (due to inclusion of additional studies)? This comparison is important for assessing consistency and meta-meta-analysis trends.
- Location: Discussion, page 13
- Impact on Validity: Readers cannot assess whether findings are novel or consistent with prior work, which affects the interpretation of the contribution's significance.
- Required Author Action:
- Compare current rs10994336 meta-analysis (OR=1.28, 95% CI: 1.13–1.45, P=1.04×10⁻⁴, n=9 studies) with Roby (2017): "The prior meta-analysis by Roby (2017) reported rs10994336 in bipolar disorder with effect estimate [cite results]. Our expanded meta-analysis, including [X additional studies since 2017], yields a slightly [larger/smaller] effect estimate (OR 1.28 vs. 1. XX in Roby), with [narrower/wider] confidence intervals, reflecting increased sample size and improved precision. The [consistency/divergence] of these findings suggests [stable effect / population-specific variation]."
Issue 5: The population stratification discussion lacks clarity on genetic background mechanisms
- Description: Discussion addresses population-specific heterogeneity (e.g., rs10761482 showing opposite effects in Chinese vs. Iranian populations) and mentions "different populations and genetic backgrounds," but does not clearly explain mechanisms for the opposite direction of effects. This could reflect: (a) genuine allelic heterogeneity (different causal variants in LD with the index SNP in different populations); (b) population-specific linkage disequilibrium structure; (c) gene-by-environment interactions (e.g., different environmental prevalences of risk factors); (d) publication bias or selection bias in study sampling. Discussion is vague and would benefit from explicit hypothesis testing or LD mapping.
- Location: Discussion, pages 12–13
- Impact on Validity: Readers may attribute heterogeneity to vague "population differences" without understanding genetic mechanisms, limiting insight into the true sources of discrepancy.
- Required Author Action:
- Expand population stratification discussion: "The opposite direction of effect for rs10761482 in Chinese vs. Iranian schizophrenia cohorts (protective in Chinese: OR=0.74; risk-increasing in Iranian: OR=1.57) could reflect several mechanisms: First, linkage disequilibrium structure varies across populations; rs10761482 may be in LD with different causal variants in East Asian vs. West Asian/Middle Eastern haplotypes, leading to apparent direction reversal despite association with the same causal locus. Second, population-specific allele frequencies may alter statistical power and effect estimate precision; rare variants in one population may be moderately common in another. Third, true population-specific effects could arise from gene-by-environment interactions; for example, if environmental stressors associated with schizophrenia risk differ between Chinese and Iranian populations, ANK3 variants may modulate risk differentially depending on environmental context. To distinguish between these mechanisms, future studies could: (1) perform fine-mapping and regional LD analysis in both populations to identify potential causal variants; (2) assess replication in independent samples separately in Chinese and Iranian cohorts; (3) investigate environmental risk factor distributions and potential interaction with ANK3 genotypes."
Minor Issues & Editorial Suggestions
1. Biological plausibility framing: Discussion appropriately contextualizes the role of the neocortex and cerebellum in psychiatric disorders; however, consider strengthening with recent literature. For example, cerebellar involvement in psychiatric disorders (schizophrenia, autism, depression) is increasingly recognized; cite recent reviews such as Schmahmann JD. Proc Natl Acad Sci USA. 2019;116(19):9375–9384 (cerebellar contribution to psychiatric disorders).
2. Comparison with meQTL findings: Discussion mentions "a previous methylation QTL (meQTL) study, which reported that rs10994336 was significantly associated with ANK3 methylation levels in the prefrontal cortex." Expanding this comparison would strengthen functional annotation: "Integrating our findings with prior meQTL results suggests multi-layered regulatory control of ANK3: methylation-mediated transcriptional regulation (meQTL) and post-transcriptional regulation (eQTL, sQTL). This multi-layered regulation may reflect the critical role of ANK3 in neuronal development and circuit formation, processes requiring precise dosage control."
3. Intronic SNP concentration discussion: Discussion notes "SNPs in both introns could mean that these regions are evolutionarily conserved and therefore may have some functionality." This is reasonable, but could be strengthened by referencing conservation metrics (PhyloCSF, GERP, etc.) or checking whether intronic sequences show evidence of constraint in gnomAD or ClinVar databases.
4. Missing discussion of psychiatric nosology implications: Given that ANK3 variants are associated with multiple psychiatric disorders (bipolar, schizophrenia, depression, PTSD), discuss whether this supports a "transdiagnostic" genetic architecture: "The association of ANK3 variants with multiple psychiatric conditions raises the possibility that ANK3-mediated neuronal dysfunction contributes to shared biological pathways underlying diverse psychiatric phenotypes, supporting a transdiagnostic genetic model of psychiatric illness. This is consistent with recent evidence that psychiatric disorders share common genetic risk factors (Lee et al., 2013; Cross-Disorder Group of the Psychiatric Genomics Consortium)."
CONCLUSIONS
Summary Assessment
The Conclusion succinctly summarizes key findings (ANK3 intronic variants associated with psychiatric disorders, tissue-specific regulatory effects) and notes the functional relevance of QTL integration. However, the Conclusion is brief and lacks concrete next steps or implications. Recommendations for future research are absent, and the scope of generalizability is not stated.
Major Issues
Issue 1: The conclusion does not specify the scope of evidence or generalizability
- Description: Conclusion states: "Multiple intronic ANK3 SNPs are significantly associated with psychiatric disorders and display tissue-specific regulatory effects." This statement combines FDR-significant findings (sQTL in cerebellum) with nominal-only findings (eQTL, apaQTL) without distinction. Readers unfamiliar with statistical thresholds may assume all findings are robustly supported; however, effect sizes vary, and some are not validated across populations.
- Location: Conclusion, page 14
- Impact on Validity: Over-interpretation of evidence; readers may attribute equal confidence to FDR-significant sQTL results and nominal eQTL findings.
- Required Author Action:
- Revise to specify evidence tiers: "Multiple intronic ANK3 SNPs are associated with psychiatric disorders: bipolar disorder and schizophrenia meta-analyses included up to nine studies for individual SNPs, while PTSD variants are based on single or small-sample analyses requiring replication. ANK3 variants display tissue-specific regulatory effects, with the most robust evidence for cerebellar splicing effects (FDR=0.010), modest evidence for neocortical expression effects (FDR=0.162), and nominal evidence for alternative polyadenylation effects (FDR=0.965) that require independent validation."
Issue 2: Conclusion lacks actionable recommendations or research directions
- Description: Conclusion ends: "Integrating QTL data provides insights into their potential functional roles in neuropsychiatric pathogenesis." This is a very general statement offering no specific next steps, no recommendations for clinical translation, no discussion of required validation studies, and no timeline for follow-up research.
- Location: Conclusion, page 14
- Impact on Validity: Readers do not understand what future work is needed to advance the field or translate findings into clinical practice.
- Required Author Action:
- Expand Conclusion with actionable recommendations:
Integrating QTL data with genetic association findings provides insights into the tissue-specific regulatory mechanisms by which ANK3 genetic variants may contribute to psychiatric disorder risk. However, several steps are necessary to translate these findings toward clinical or therapeutic application: (1) Experimental validation of sQTL and eQTL associations using CRISPR perturbation in primary neural cells or iPSC-derived neurons to establish causality; (2) Cross-ancestry replication of meta-analysis findings in non-European populations to assess effect heterogeneity and generalizability; (3) Integration of ANK3 variants into polygenic risk score models for psychiatric disorders to assess predictive accuracy; (4) Investigation of potential gene-by-environment or gene-by-sex interactions that may modify ANK3 variant effects; (5) Functional characterization of ANK3 isoform distributions in postmortem psychiatric tissue to link genetic associations with pathological phenotypes. Until such validation is complete, ANK3 variants should be considered part of the polygenic architecture underlying psychiatric disease risk rather than individual targets for clinical stratification or drug development.
Minor Issues & Editorial Suggestions
1. Plural vs. singular reference: Conclusion says "Multiple intronic ANK3 SNPs," but Results identify 10 SNPs total. Consider specifying: "At least 10 intronic ANK3 variants, identified across four psychiatric disorders, display significant genetic associations..."
2. "Insights" vagueness: The phrase "provides insights into...functional roles" is passive and vague. Consider: "elucidate potential mechanisms by which ANK3 genetic variants contribute to psychiatric disease" or "enable hypothesis generation regarding ANK3-mediated neuronal dysfunction in psychiatric pathogenesis."
FIGURES & TABLES
Summary Assessment
Figures 1–7 are generally clear and appropriately support the manuscript text; however, resolution, labeling, and data transparency issues reduce their utility. Tables 1–4 are well-formatted but lack supplementary detail (cell counts, weights, heterogeneity statistics) required for full reproducibility and meta-analysis verification.
Major Issues
Issue 1: Forest plots (Figures 2–5) lack essential statistical annotations
- Description: Figures 2–5 show forest plots with individual study effect estimates and pooled OR, but lack: (a) heterogeneity statistics (I², τ²) on the plot or in caption; (b) study weights (proportional to bubble size); (c) number of studies and combined sample size; (d) confidence intervals for individual studies (confidence intervals should be shown on plot); (e) funnel plot estimates or publication bias commentary.
- Location: Figures 2, 3, 4, 5 (pages 6–7, 10)
- Impact on Validity: Readers cannot assess statistical rigor or identify studies with outlier effects. Standard forest plot reporting per Cochrane/PRISMA requires confidence intervals and heterogeneity statistics.
- Required Author Action:
- Add to each forest plot caption: "Heterogeneity statistics: I² = XX%, τ² = XX, Q-test p-value = X.XXX. Pooled effect estimated using random-effects meta-analysis. Individual study squares represent point estimates (OR); horizontal lines span 95% confidence intervals; the diamond represents the pooled estimate and 95% CI. Study labels show author (year); sample sizes (cases/controls) are indicated in parentheses."
- Provide forest plot data tables in Supplementary Materials showing: Study, N_cases, N_controls, OR, 95% CI, Weight (%)
- If publication bias is suspected, add funnel plot and Egger test results to supplement forest plots
Issue 2: Figure 1 (literature screening flow diagram) is missing data
- Description: Already detailed in the Results section, Issue 2 above. Figure 1 is overly simplified and lacks exclusion reason tallies.
- Location: Figure 1, page 5
- Impact on Validity: [See Results Issue 2 for full discussion and required action]
- Required Author Action: [As specified in Results Issue 2]
Issue 3: Figure 6 (SNP localization on ANK3 gene) lacks scale and clarity
- Description: Figure 6 displays the ANK3 gene structure with SNP positions; however, the figure lacks: (a) a genomic scale bar (bp or Mb); (b) axis labels with coordinates (GRCh38.p14); (c) exon numbering clarity; (d) visual distinction between major and minor SNP clusters. Currently, SNP positions are not easily readable without high-resolution magnification.
- Location: Figure 6, page 8
- Impact on Validity: Readers cannot discern the scale of SNP clustering or the gene's size relative to SNPs. The Discussion emphasizes "concentration of SNPs in both introns," but Figure 6 does not make this apparent.
- Required Author Action:
- Redraw Figure 6 with: (1) Genomic coordinates on x-axis (e.g., "Chr10: 60.2 Mb – 60.7 Mb"); (2) Scale bar (e.g., "1 Mb"); (3) Exon numbering above gene structure; (4) Intron numbers or names; (5) SNP labels with rsID and associated disorder color-coding (e.g., red for bipolar, blue for schizophrenia, green for PTSD); (6) clear visual distinction between the two main introns harboring clusters (Intron 2–3 and Intron 36–37)
- High-resolution version suitable for publication (at least 300 dpi)
Issue 4: Table 1 (SNP localization summary) lacks phenotype specificity
- Description: Table 1 summarizes SNP positions in exons/introns and associates them with psychiatric disorder columns (Bipolar disorder, Schizophrenia, Major depression, Posttraumatic stress disorder). However, the table does not indicate which SNPs are significant in each disorder (italics or bold formatting would help), and the distinction between meta-analysis significant and single-study reported variants is not clear.
- Location: Table 1, page 9
- Impact on Validity: Readers cannot quickly identify which SNPs are robustly associated with which disorders. Table readability is reduced.
- Required Author Action:
- Reformat Table 1 as follows:
```
| SNP | Location | Chr:bp (GRCh38) | Bipolar Disorder | Schizophrenia | Major Depression | PTSD |
|--|-|--||-|||
| rs10994336 | Intron 2–3 | 10:60,XXX | OR=1.28 (n=9) | ns (n=X) | ns (n=X) | — |
| rs9804190 | Intron 36–37 | 10:60,XXX | OR=0.88 (Eur) (n=3) | ns | — | nominal |
| rs10994359 | Intron 2–3 | 10:60,XXX | — | — | OR=0.69 (Asian) (n=X) | — |
Significance levels: FDR<0.001; FDR<0.01; FDR<0.05; ns, not significant in meta-analysis; —, not tested. Abbreviations: OR, odds ratio; Eur, European populations; n, number of independent studies in meta-analysis.
- Add footnote clarifying which results are from meta-analysis (n≥2 studies) vs. synthetic review (n=1 study)
Issue 5: Tables 2–4 (QTL results) do not show sufficient context (number of tissues tested, n values)
- Description: Tables 2–4 report QTL effect estimates (β, SE, P-value, FDR) for individual SNP-tissue pairs, but lack context: How many tissues were tested in total for eQTL? How many SNPs were tested? Tables appear to show only significant or selected findings; complete results are referenced to the Extended datasets, but summary tables in the main text would improve interpretation.
- Location: Tables 2, 3, 4 (pages 10–11)
- Impact on Validity: Readers cannot assess the total multiple testing burden or whether selective reporting of results has occurred. For example, if 10 SNPs × 50 tissues = 500 eQTL tests were performed, FDR correction should reflect this, and the single significant eQTL for rs10761482 should be interpreted cautiously.
- Required Author Action:
- Add table header or caption: "eQTL analysis: 10 SNPs tested across 47 brain tissues/cell types in the eQTL Catalogue = 470 SNP-tissue pairs tested. FDR correction applied within eQTL modality; FDR<0.05 considered significant. Results shown for SNPs with nominal P<0.05; complete results in Extended dataset 4."
- Create supplementary summary table showing: (1) Total number of SNP-tissue pairs tested per QTL modality; (2) Number of significant associations (FDR<0.05); (3) Number of nominally significant but FDR-nonsignificant findings; (4) Range of effect sizes observed
- Provide complete results tables for eQTL, apaQTL, sQTL in Supplementary Materials or Extended datasets
Minor Issues & Editorial Suggestions
1. Figure legibility: Ensure all figure text (axis labels, legends, SNP labels) is legible at publication size (typically 8–10pt font for figures). Current Figures 2–5 forest plot study labels may be too small.
2. Color coding consistency: If different colors are used for different disorders across figures (e.g., red for bipolar, blue for schizophrenia in Figure 6), ensure the color scheme is consistent and colorblind-friendly (avoid red-green combinations).
3. Figure panel labels: Ensure all multi-panel figures (e.g., Figure 7 with panels A–D, Figure 2 with panels A–D) are clearly labeled with capital letters and referenced in the caption.
4. Supplementary materials reference: Ensure all references to "Extended dataset" include active DOI links and verify Figshare accessibility at submission.
ETHICAL CONSIDERATIONS, FUNDING, DATA AVAILABILITY
Summary Assessment
The manuscript appropriately states that the study used only publicly available summary-level data and did not involve human participants, no ethics approval was required. Funding and competing interests disclosures are provided. However, the data availability statement could be more explicit regarding which raw data are available vs. which are restricted, and the PROSPERO registration status is not mentioned.
Major Issues
Issue 1: PROSPERO registration status not reported
- Description: The manuscript does not state whether this systematic review was registered with PROSPERO (International Prospective Register of Systematic Reviews) before publication. PRISMA 2020 item 4 requires reporting of PROSPERO registration number and registration date. Pre-registration is essential for reducing bias and demonstrating that analyses were not post-hoc selected based on observed data.
- Location: Methods section, or missing entirely
- Impact on Validity: Inability to verify whether eligibility criteria, quality assessment, and statistical analysis plans were pre-registered vs. decided after observing data. Increases risk of selective reporting.
- Required Author Action:
- If PROSPERO registration was completed: Include PROSPERO registration number (e.g., "PROSPERO registration: CRD42024XXXXXXX") in Methods and cite in Results (e.g., "This systematic review was registered with PROSPERO before the start of study selection (PROSPERO registration: CRDXXXXXXX, registered [date]).")
- If PROSPERO registration was not completed (post-hoc registration of completed review is possible): State in Methods: "This systematic review was completed before PROSPERO registration but has been registered retroactively (PROSPERO registration: CRDXXXXXXX). The predetermined protocol [if available as supplementary document] specified the eligibility criteria, search strategy, quality assessment tool (Newcastle-Ottawa Scale), and planned analyses."
- If PROSPERO registration was not completed and the pre-registration document is not available: Acknowledge as a limitation and state that future reviews will be pre-registered
Issue 2: Data availability statement incomplete regarding access restrictions
- Description: Data Availability section states: "The data used in this meta-analysis and QTLs analysis, including the eQTL Catalogue and GTEx v10, were obtained from published articles and public databases... All datasets are released under the terms of the Creative Commons Attribution 4.0 International licence (CC BY 4.0)." However, it does not specify: (a) Are the extracted datasets (study-level ORs, 95% CIs) provided as supplementary files or only on Figshare? (b) Can the raw individual-participant data from included studies be accessed, and if not, why? (c) Are the R scripts and data parsing code available for reproducibility? (d) Are there any data use restrictions from the eQTL Catalogue or GTEx v10?
- Location: Data Availability section, pages 13–14
- Impact on Validity: Readers cannot easily access or verify the extracted data, limiting reproducibility and independent verification of reported findings.
- Required Author Action:
- Expand Data Availability statement to:
Data Availability:
This study is based entirely on publicly available summary-level data from published literature and public genomic databases. No new primary data were generated. The extracted meta-analysis datasets, including study-level effect estimates (odds ratios, 95% confidence intervals, P-values) for all included studies and SNP-disorder combinations, are openly available on Figshare under Creative Commons Attribution 4.0 International (CC BY 4.0) license:
All R and Python scripts used for data parsing, harmonization, and QTL analysis are available on GitHub at [repository URL] and archived on Figshare [DOI] under a CC BY 4.0 license.
The eQTL Catalogue and GTEx v10 summary statistics are publicly available through their respective portals (https://www.ebi.ac.uk/eqtl/ and https://gtexportal.org/) under their respective data use agreements. No restrictions apply to their use for secondary analysis and research purposes.
Individual-level genotype and phenotype data from the included genome-wide association studies are typically not publicly available due to privacy regulations (e.g., GDPR); however, summary-level statistics have been extracted and are provided in our datasets. Access to individual-level GWAS data may be available through the original study authors or through the Psychiatric Genomics Consortium (https://www.med.unc.edu/pgc/) with appropriate data use agreements.
Computed outputs (forest plots, QTL heatmaps, supplementary tables) are provided as Supplementary Materials and Extended datasets.
Issue 3: Funding statement adequacy
- Description: Grant information states: "The author(s) declared that no grants were involved in supporting this work." However, the acknowledgments section mentions: "TG gratefully acknowledges the invaluable guidance and support provided by his supervisors and research team throughout the course of his PhD studies. TG also extends his sincere appreciation to the China Scholarship Council for their financial support." This is contradictory: if the China Scholarship Council provided financial support, it should be listed under funding, not only in acknowledgments.
- Location: Grant information section (page 2) and Acknowledgments (page 15)
- Impact on Validity: Incomplete funding disclosure; readers may not recognize the source of financial support, affecting the assessment of potential bias. ICMJE guidelines require complete disclosure of all funding sources.
- Required Author Action:
- Revise Grant information to: "Grant information: TG was supported by the China Scholarship Council (CSC) PhD Scholarship (2019–2023). YW acknowledges support from [list any research grants or institutional funding]. KD acknowledges support from [list any research grants or institutional funding]. No additional grants were involved in supporting this work."
- Streamline Acknowledgments to: "TG thanks his supervisors for mentorship and research team guidance. Figshare hosting and extended data management were supported by [institutional funding or F1000Research open access initiative]."
Minor Issues & Editorial Suggestions
1. Open Peer Review Option: F1000Research is known for open peer review, where reviewer reports are published with the article. The manuscript does not mention whether the authors have opted into open peer review. Consider: "Authors consent to open peer review; reviewer reports will be published alongside this article" (or alternative statement from F1000Research policy).
2. Ethics approval statement: Current ethics statement appropriately notes: "This study was based entirely on publicly available summary-level genomic data... Therefore, no ethical approval or informed consent was required." Consider adding a reference to the ICMJE guidelines: "In accordance with ICMJE guidelines (http://www.icmje.org/recommendations/browse/roles-responsibilities/author-responsibilities-conflicts-of-interest.html), this meta-analysis did not require ethics approval as no primary data were collected."
LINE-BY-LINE EDITORIAL SUGGESTIONS
Abstract
Current Text (Methods Section):
"A meta-analysis and systematic literature review were conducted to identify disease-associated SNPs in ANK3. QTL analyses were performed using datasets from the eQTL Catalogue and GTEx v10 to evaluate regulatory effects in brain tissues."
Suggested Replacement:
"A systematic literature review and meta-analysis were conducted to identify ANK3 single-nucleotide polymorphisms (SNPs) significantly associated with psychiatric disorders across published studies (PubMed, Web of Science, MEDLINE; search period: January 2000–September 2025; n=27 included studies). Population-stratified meta-analyses were performed using odds ratios from included studies and random-effects models. Secondary expression quantitative trait locus (eQTL), alternative polyadenylation QTL (apaQTL), and splicing QTL (sQTL) analyses were conducted using public datasets (eQTL Catalogue, GTEx v10) to characterize tissue-specific regulatory effects of disease-associated variants in the human brain."
Current Text (Results Section):
"In bipolar disorder, rs10994336 (OR = 1.28, P = 1.04×10−4) was significant in European+North American populations; rs9804190 was protective in Europeans (OR = 0.88, P = 0.020)..."
Suggested Replacement (with specificity of significance):
"Meta-analysis identified ten ANK3 variants significantly associated with psychiatric disorders. In bipolar disorder, rs10994336 (OR = 1.28, 95% CI: 1.13–1.45, P = 1.04×10⁻⁴) emerged as a significant risk variant in European and North American populations (n=9 studies); this association was not replicated in Asian cohorts (P = 0.600, n=X studies). Rs9804190 showed significant protective effects in European studies (OR = 0.88, 95% CI: 0.79–0.98, P = 0.020, n=3 studies); however, this association did not extend to combined European and North American analysis (P = 0.173). Rs1938526 demonstrated robust cross-population associations in both European/North American (OR = 1.32, 95% CI: 1.22–1.43, P = 7.26×10⁻¹²) and Asian populations (OR = 1.08, 95% CI: 1.04–1.12, P = 4.68×10⁻¹²), with pooled effect estimate OR = 1.17 (95% CI: 1.05–1.29, P = 0.003)..."
Introduction
Current Text:
"Over the past decade, genome-wide association studies have identified significant associations between ANK3 single nucleotide polymorphisms (SNPs) and a range of psychiatric disorders, including bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD). These findings suggest that genetic variation within the ANK3 locus may contribute to shared pathophysiological mechanisms underlying multiple neuropsychiatric phenotypes. However, despite accumulating evidence, considerable inconsistencies remain regarding which ANK3 SNPs are most robustly associated with psychiatric disorders and whether these associations are consistent across diverse populations."
Suggested Replacement (with specific examples):
"Over the past decade, genome-wide association studies (GWAS) have identified multiple significant associations between ANK3 single nucleotide polymorphisms (SNPs) and a range of psychiatric disorders, including bipolar disorder (BD), schizophrenia (SZ), major depressive disorder (MD), and post-traumatic stress disorder (PTSD). These findings suggest that genetic variation within the ANK3 locus may contribute to shared pathophysiological mechanisms underlying multiple neuropsychiatric phenotypes. However, considerable inconsistencies remain in the literature: for example, rs10994336 was significantly associated with bipolar disorder in European cohorts (Ferreira et al., 2008; Scott et al., 2009) but not in East Asian populations (Takata et al., 2011; Lee et al., 2011), and rs10761482 showed opposite associations in Chinese (protective) versus Iranian (risk-increasing) schizophrenia cohorts (Dashtban et al., 2022; Karimian et al., 2020). Such population-specific effect heterogeneity suggests that ANK3 genetic architecture may vary across ancestries, highlighting the need for systematic synthesis of findings with explicit population stratification and comprehensive functional characterization of disease-associated variants."
Methods
Current Text (Search Strategy):
"In accordance with the predefined inclusion criteria, two independent researchers (TG and YW) systematically searched PubMed, Web of Science, and MEDLINE using combinations of the terms 'genome-wide association study' with 'ANK3', as well as 'nucleotide polymorphisms' with 'ankyrin 3' or 'Ankyrin-G'. Relevant studies were screened, and data were extracted independently by both reviewers using a standardized data extraction form."
Suggested Replacement:
"In accordance with the predefined inclusion criteria and PRISMA 2020 guidelines, two independent researchers (TG and YW) systematically searched PubMed, Web of Science, and MEDLINE from 1 January 2000 through 30 September 2025. The search strategy combined controlled vocabulary and text terms as follows:
PubMed search string:
`(ANK3 OR "ankyrin 3" OR "ankyrin-3" OR "ankyrin G" OR "ankyrin-G" OR ANKRD4 OR "ankyrin 3 gene") AND (polymorphism OR SNP OR genotype OR allele OR "rs[0-9]" OR association OR GWAS) AND (psychiatric OR schizophrenia OR bipolar OR depression OR PTSD OR "post traumatic" OR "mental disorder")`
Web of Science topic search:
`TS=(ANK3 OR "ankyrin 3" OR "ankyrin-G") AND TS=(SNP OR polymorphism OR genotype) AND TS=(schizophrenia OR bipolar OR depression OR psychiatric OR PTSD)`
Searches were limited to peer-reviewed publications in English. Retrieved references were imported into a reference management system (Mendeley or Zotero), duplicates removed, and remaining articles (n=1,140) underwent two-stage screening: (1) title and abstract review by both researchers independently, and (2) full-text assessment of potentially eligible articles. Disagreements were resolved through discussion with a third researcher (KD) until consensus was achieved. A Preferred Reporting Items for Systematic Review and Meta-Analysis (PRISMA) 2020 flow diagram documenting the study selection process is provided in Figure 1."
Results
Current Text (Study Selection):
"Using the predefined search strategy, a total of 1,140 articles were retrieved from the selected databases. Following title and abstract screening, duplicate entries were removed. Full-text review resulted in the exclusion of 214 studies, primarily due to methodological insufficiencies or unclear study protocols. Ultimately, 27 English-language studies met the inclusion criteria and were retained for analysis."
Suggested Replacement:
"Using the predefined search strategy, a total of 1,140 unique articles were retrieved from PubMed (n=XXX), Web of Science (n=XXX), and MEDLINE (n=XXX) [insert actual numbers from original search results]. After removal of XXX duplicate records, 1,XXX articles underwent title and abstract screening, resulting in XXX articles advanced to full-text review. Full-text assessment resulted in exclusion of 214 studies for the following reasons: [n=XX] did not specifically test ANK3 SNP associations; [n=XX] focused on non-psychiatric outcomes; [n=XX] were reviews, case reports, or expert commentaries; [n=XX] lacked clear information on participant geography; [n=XX] reported hazard ratios rather than odds ratios; [n=XX] other (specify). Ultimately, 27 studies met all inclusion criteria and were retained for systematic review and meta-analysis. All included studies achieved quality scores ≥5/9 on the Newcastle-Ottawa Scale, indicating adequate study quality (Supplementary Table X). The study selection process is documented in the PRISMA 2020 flow diagram (Figure 1)."
Discussion
Current Text (Mechanistic Claims):
"Interestingly, three of the SNPs significantly associated with psychiatric disorders—rs28932171, rs11599164, and rs17208576—that are located in the intronic region between exon 36 and exon 37 exhibit strong splicing QTL (sQTL) signals specifically in cerebellum tissue (Table 4). These findings suggest that the risk variants may influence alternative splicing decisions that determine exon 37 inclusion. Such splice isoform shifts could lead to changes in the intracellular distribution of Ankyrin-G, potentially altering its ability to cluster ion channels at the AIS and thereby affecting neuronal polarity, excitability, or signaling, leading to cellular and circuit-level changes in neuronal signalling, and to disease in the long term."
Suggested Replacement (Tempered Claims with Caution):
"Notably, three ANK3 variants significantly associated with psychiatric disorders—rs28932171, rs11599164, and rs17208576—are located in the intronic region upstream of exon 37 (a brain-specific exon encoding the AIS targeting motif of Ankyrin-G) and show robust evidence of association with alternative splicing in human cerebellum tissue (sQTL FDR=0.010, Table 4). These quantitative trait locus associations provide suggestive evidence that these intronic variants may regulate ANK3 exon 37 inclusion, potentially affecting isoform composition in cerebellum. Given the documented role of exon 37 in determining AIS-localized vs. postsynaptic ANK3 isoforms, and the established importance of AIS ion channel clustering for neuronal excitability, one plausible mechanism is that splice isoform shifts driven by these variants could alter the relative abundance of AIS-targeted and postsynaptic Ankyrin-G pools, thereby modulating neuronal polarity, action potential generation, or circuit connectivity in cerebellum. However, these sQTL associations are correlative rather than causal; the identified SNPs may serve as proxy markers in linkage disequilibrium with unmapped regulatory elements or non-coding RNA genes, rather than being directly functional. Experimental validation through CRISPR perturbation of these intronic elements, allele-specific expression assays, and functional characterization of ANK3 isoform shifts in neural cells would be necessary to establish mechanistic causality. Furthermore, the relevance of cerebellar ANK3 splicing alterations to psychiatric symptomatology remains unknown; future studies directly assessing ANK3 isoform distributions in postmortem psychiatric brain tissue or identifying correlations between ANK3 splicing variants and clinical phenotypes would clarify the functional impact on disease pathogenesis."
Conclusion
Current Text:
"Multiple intronic ANK3 SNPs are significantly associated with psychiatric disorders and display tissue-specific regulatory effects. Integrating QTL data provides insights into their potential functional roles in neuropsychiatric pathogenesis."
Suggested Replacement:
"Multiple intronic ANK3 variants demonstrate significant associations with psychiatric disorders, particularly bipolar disorder and schizophrenia, with effect sizes varying across populations. These variants display tissue-specific regulatory effects on ANK3 gene expression and splicing, with the most robust evidence for cerebellar alternative splicing (sQTL FDR=0.010) and modest evidence for neocortical expression regulation (eQTL FDR=0.162). However, several important limitations warrant acknowledgment: (1) meta-analysis sample sizes are modest for some SNP-disorder combinations, limiting statistical power and generalizability, particularly for PTSD variants derived from single studies; (2) psychiatric phenotypes differ across studies in diagnostic criteria and assessment methods, introducing phenotypic heterogeneity; (3) regulatory QTL effects are predominantly derived from European ancestry samples and may not generalize to underrepresented populations. Integrating genetics with multi-layered functional genomics (eQTL, sQTL, apaQTL) provides important insights into potential tissue-specific mechanisms by which ANK3 intronic variants may contribute to psychiatric disorder vulnerability. However, these associations are correlative; future experimental validation in neural cell models, cross-ancestry replication, and investigation of gene-by-environment interactions are necessary to establish causality and clarify the functional role of ANK3 in psychiatric pathogenesis. Integration of ANK3 variants into larger polygenic risk models may improve psychiatric disease prediction, but individual variants are not yet suitable for clinical genetic testing given modest effect sizes. Further research integrating genetics, molecular biology, and psychiatric endophenotyping is essential to translate these findings toward clinical or therapeutic application."
OVERALL RECOMMENDATION
Summary of Key Findings
Strengths:
1. Timely and comprehensive systematic review addressing an important psychiatric genetics question
2. Well-motivated integration of meta-analysis with functional genomics (QTL analyses)
3. Appropriate acknowledgment of population stratification and heterogeneity
4. Transparent reporting of FDR-corrected vs. nominal findings
5. Open data and transparent science practices (Figshare data deposition, CC-BY license)
6. Clear acknowledgment of ethical practices (no human subjects, appropriate data sources)
Critical Weaknesses:
1. Search strategy insufficiently reported: Databases, dates, Boolean operators, MeSH terms, completeness filters not transparent. Risk of selection bias.
2. Heterogeneity statistics omitted from main Results: I² and τ² not reported for meta-analyses, limiting assessment of effect generalizability.
3. PRISMA 2020 non-compliant: PROSPERO registration status not reported; PRISMA checklist not included in submission; explicit PICO statement absent.
4. Mechanistic claims overstated: QTL findings presented as causal evidence; requires tempering with acknowledgment of correlative nature of associations.
5. Limitations inadequately discussed: Small sample sizes (PTSD variants single-study), phenotype heterogeneity, QTL ancestry bias, lack of causal inference, publication bias assessment not conducted.
6. Data transparency gaps: Heterogeneity statistics, individual study data, NOS item-level assessments not in main text; reliance on Extended datasets may limit reproducibility.
7. Conflicting funding disclosures: Grant information states "no grants" while Acknowledgments mention China Scholarship Council support.
Moderate Weaknesses:
1. Clinical implications and future research directions minimally discussed
2. Comparison with prior ANK3 rs10994336 meta-analysis incomplete
3. QTL tissue selection and justification not documented
4. Figures (forest plots, gene map) lack essential statistical annotations and legend detail
5. PTSD findings (three SNPs with identical OR=0.48, P=0.045) require careful interpretation due to small sample sizes
MAJOR REVISIONS REQUIRED
Justification:
The manuscript presents a solid evidence synthesis of ANK3 genetic associations with psychiatric disorders, integrates functional genomics meaningfully, and demonstrates good scientific rigor in several areas (multi-reader screening, quality assessment, QTL multiple-testing correction). However, critical gaps in methodological transparency, compliance with reporting standards (PRISMA 2020, ICMJE), and statistical reporting significantly limit publication readiness. The work is scientifically sound but requires substantial revision for publication in a rigorous, open-access venue such as F1000Research.
Primary Issues Requiring Resolution (Critical):
1. Complete search strategy documentation (databases, dates, Boolean operators, counts per stage)
2. Explicit PICO statement in Introduction or Methods
3. Report heterogeneity statistics (I², τ², Q-test P-values) for all meta-analyses in Results and Tables
4. PROSPERO registration documentation or acknowledgment of post-hoc registration
5. Funding disclosure correction (resolve Grant information vs. Acknowledgments discrepancy)
6. Expanded Limitations section addressing: small sample sizes, phenotype heterogeneity, ancestry bias in QTL, lack of causal inference, publication bias, small-study effects
7. Tempering of mechanistic claims with explicit caveat that QTL associations are correlative, not causal
8. Addition of clinical implications and future research directions discussion
Secondary Issues Requiring Resolution (Major):
1. Comprehensive statistical tables with individual study data, weights, confidence intervals, and heterogeneity statistics for forest plots
2. Forest plots with heterogeneity statistics in captions or overlaid on plots
3. Complete QTL methods detail: tissues/cell types tested, total number of tests, FDR correction rationale, interpretation of nominal vs. FDR-significant findings
4. Risk-of-bias assessment transparency: NOS scores for all 27 studies in supplementary table
5. Clarification of PTSD variant data source and sample size justification
DIRECT QUESTIONS FOR AUTHORS (REVISION RESPONSE)
During revision, authors should address the following numbered questions in a point-by-point response document:
1. PROSPERO Registration: Was this systematic review pre-registered with PROSPERO prior to study selection? If yes, provide registration number (CRD4XXXXXXX). If no, explain why pre-registration was not conducted, and indicate whether post-hoc registration is feasible before publication.
2. Search Strategy Reproducibility: Can authors provide the complete, exact search strings used in PubMed, Web of Science, and MEDLINE, including MeSH terms, field tags, date ranges, and any filters applied (e.g., humans, English language, peer review status)? Have the search strategies been validated against known relevant studies (e.g., Ferreira et al. 2008, Schulze et al. 2009 on ANK3 bipolar disorder)?
3. Heterogeneity Statistics: Authors state that meta-analysis was conducted using the R meta package version 4.18-0. Can authors provide the I² (and 95% CI for I²), τ², Q-test statistic, and Q-test P-value for EACH SNP-disorder meta-analysis (not just those reported in Extended datasets)? Were prediction intervals calculated and reported?
4. HWE Stratification Pre-Specification: In the schizophrenia rs10761482 analysis, was the stratification by Hardy-Weinberg equilibrium (HWE) status predetermined before data extraction, or decided post-hoc after observing HWE violations in Chinese studies? If post-hoc, can authors justify this decision and acknowledge it as an exploratory subgroup analysis?
5. PTSD Sample Sizes: The three PTSD SNPs (rs28932171, rs11599164, rs17208576) are reported with identical OR=0.48 and P=0.045. How many independent studies contributed to each PTSD SNP meta-analysis? Were these derived from a single study or linked studies? If from a single study, this should be clearly labeled as a "synthetic review" finding, not a meta-analysis.
6. QTL Tissue Selection: What was the total number of tissues and cell types tested in the eQTL analysis? For example, was every available tissue in the eQTL Catalogue included, or were specific tissues (e.g., psychiatric-relevant tissues) selected a priori? How many SNP-tissue pairs were tested in total for each QTL modality (eQTL, apaQTL, sQTL)? This number is needed to verify the FDR correction magnitude.
7. Newcastle-Ottawa Scale Scores: Can authors provide a supplementary table with complete NOS itemized scores for all 27 included studies? Specifically, for each study, report scores for: (a) case definition adequacy; (b) case representativeness; (c) control selection; (d) control definition; (e) comparability on design; (f) comparability on analysis; (g) exposure ascertainment; (h) same ascertainment method; (i) non-response rate. Total score (/9) should also be provided.
8. Publication Bias Assessment: Authors state "publication bias was not assessed" because fewer than 10 studies per meta-analysis. However, do funnel plots exist for the larger meta-analyses (e.g., rs10994336 BD with n=9 studies)? Can authors provide visual funnel plots and Egger test results (even with caution regarding small sample limitations) in supplementary materials?
9. Funding Clarification: The Grant information section states "no grants were involved," yet the Acknowledgments state "TG also extends his sincere appreciation to the China Scholarship Council for their financial support." Was the China Scholarship Council PhD scholarship considered "grant funding" by the authors? Please clarify the funding status and revise the Grant information section to accurately reflect all funding sources.
10. Effect Direction Consistency: In schizophrenia rs10761482, Chinese cohorts show a protective effect (OR=0.74) while Iranian cohorts show a risk effect (OR=1.57). Did authors assess whether these could reflect the same causal variant (with differential LD across populations) vs. genuinely opposite effects? Can authors provide linkage disequilibrium data or fine-mapping evidence for this locus across populations?
11. Generalizability of QTL Findings: The eQTL Catalogue and GTEx v10 datasets have limited ancestry diversity, predominantly European. Can authors assess or mention whether eQTL, sQTL, and apaQTL effect directions and magnitudes are likely conserved across ancestry groups? Alternatively, is there published evidence on trans-ethnic generalizability of these specific QTL associations?
12. Causal Inference Frameworks: Did authors consider employing Mendelian randomization or colocalization analyses to distinguish between (a) direct functional variants, (b) regulatory variants affecting ANK3 expression, and (c) proxy markers in linkage disequilibrium? If not, is this a planned direction for future work?
Are the rationale for, and objectives of, the Systematic Review clearly stated?
Partly
Are sufficient details of the methods and analysis provided to allow replication by others?
Partly
Is the statistical analysis and its interpretation appropriate?
Partly
Are the conclusions drawn adequately supported by the results presented in the review?
Partly
If this is a Living Systematic Review, is the ‘living’ method appropriate and is the search schedule clearly defined and justified? (‘Living Systematic Review’ or a variation of this term should be included in the title.)
No
Competing Interests: No competing interests were disclosed.
Reviewer Expertise: Oncology; Cancer genetics
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Sengupta D. Reviewer Report For: Single Nucleotide Polymorphisms in ANK3 and Psychiatric Risk: A Meta-Analysis, Systematic Review, and Quantitative Trait Locus Insights [version 1; peer review: 1 approved with reservations]. F1000Research 2025, 14:1020 (https://doi.org/10.5256/f1000research.186294.r450176)
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
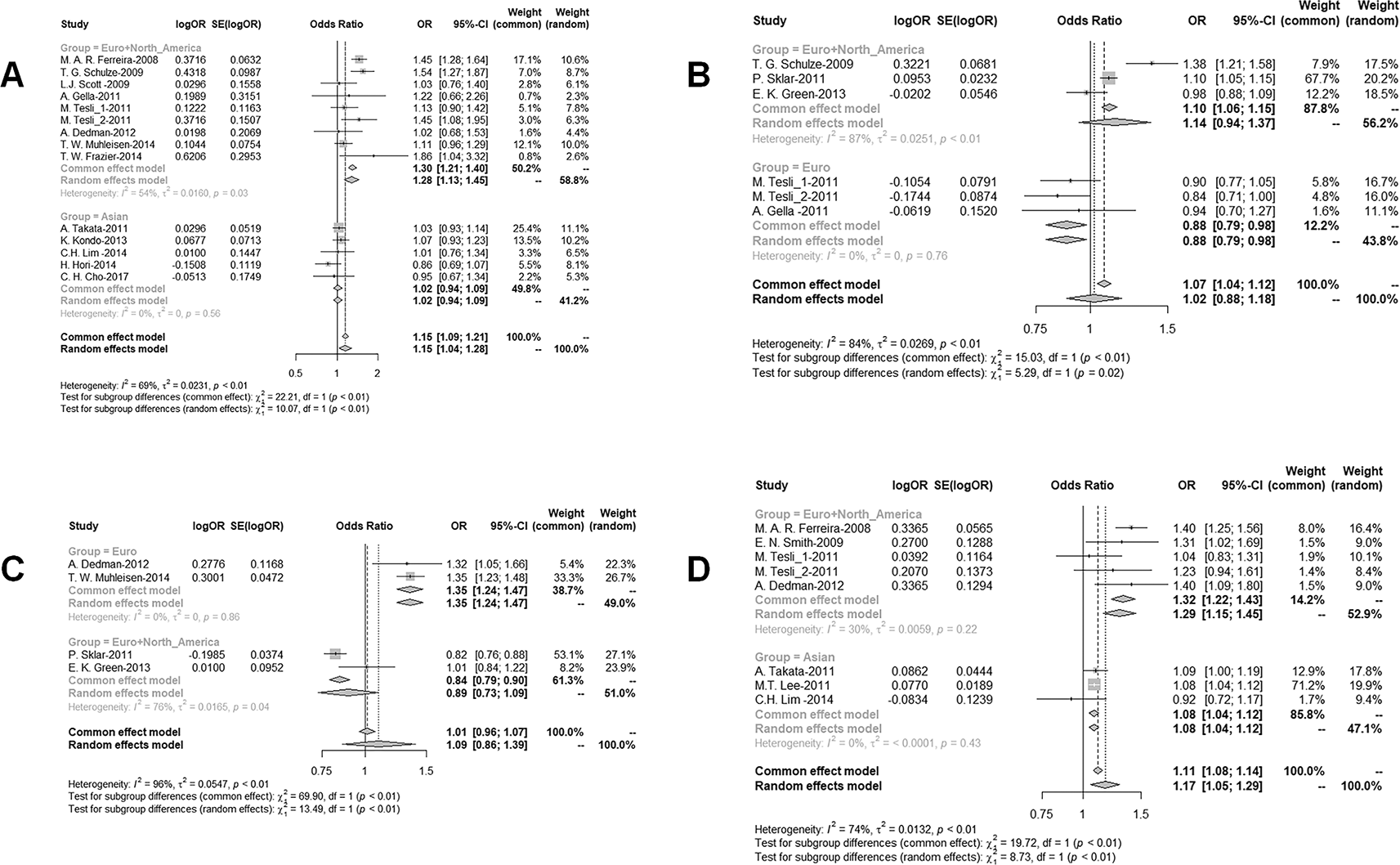
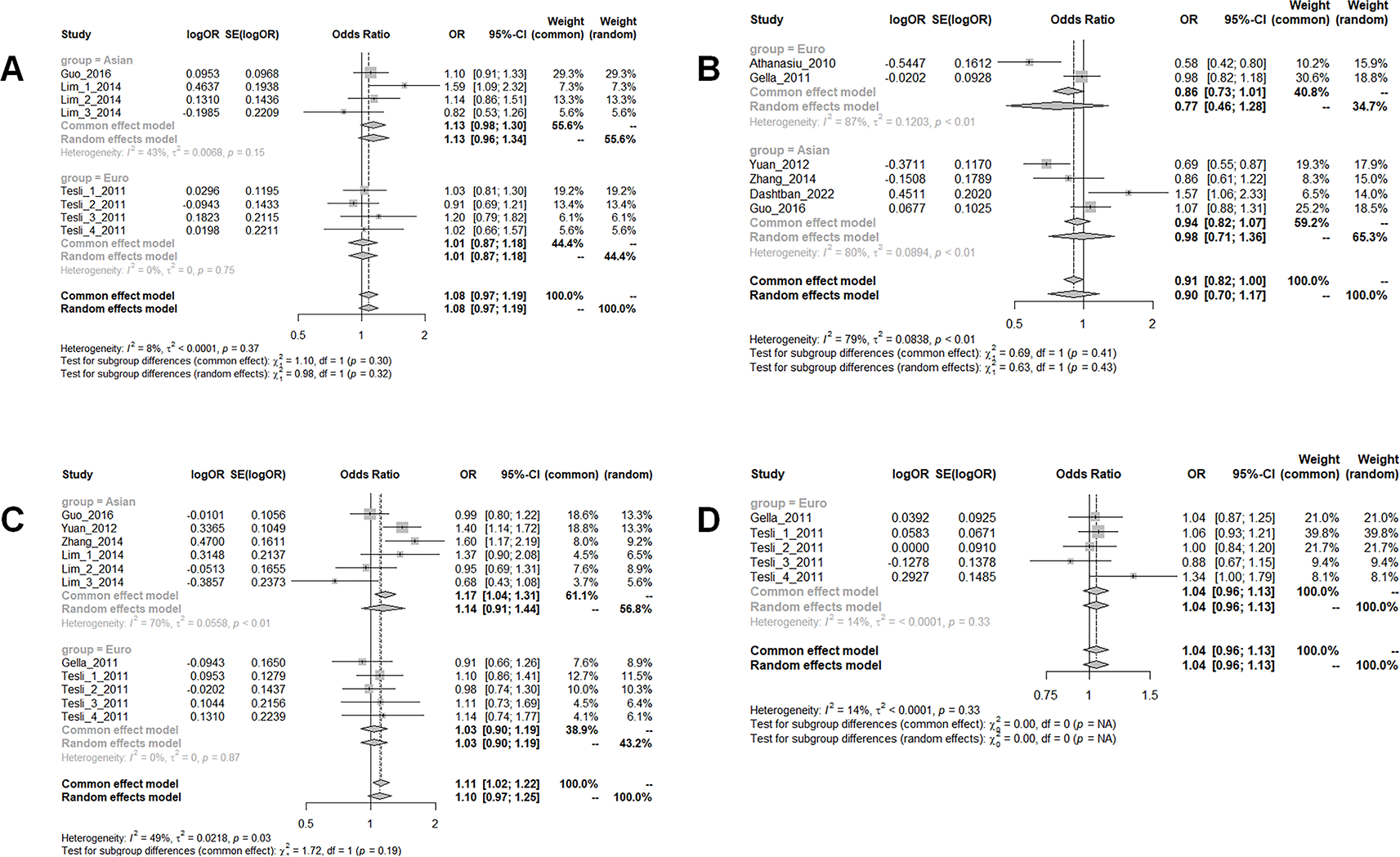
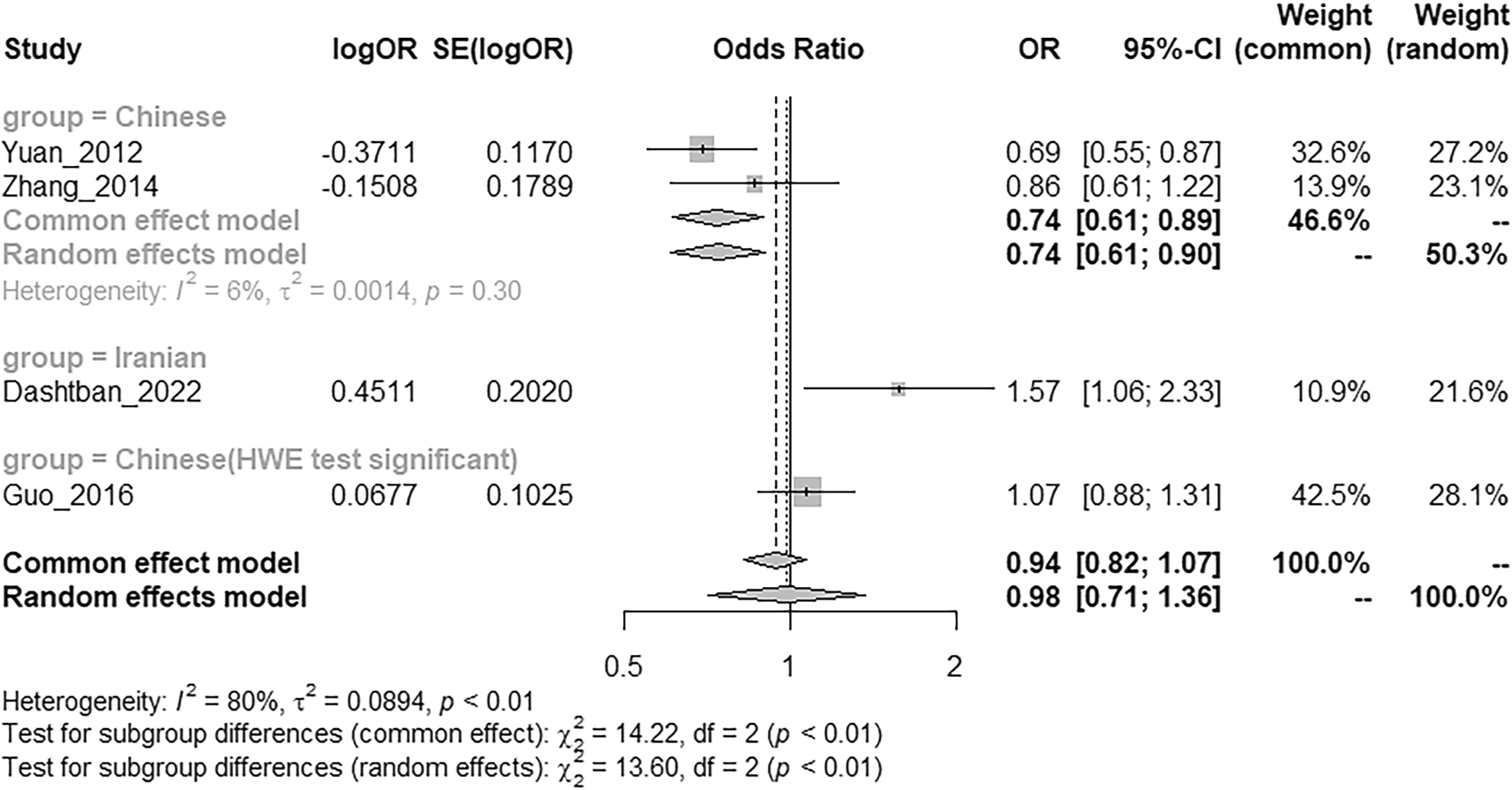
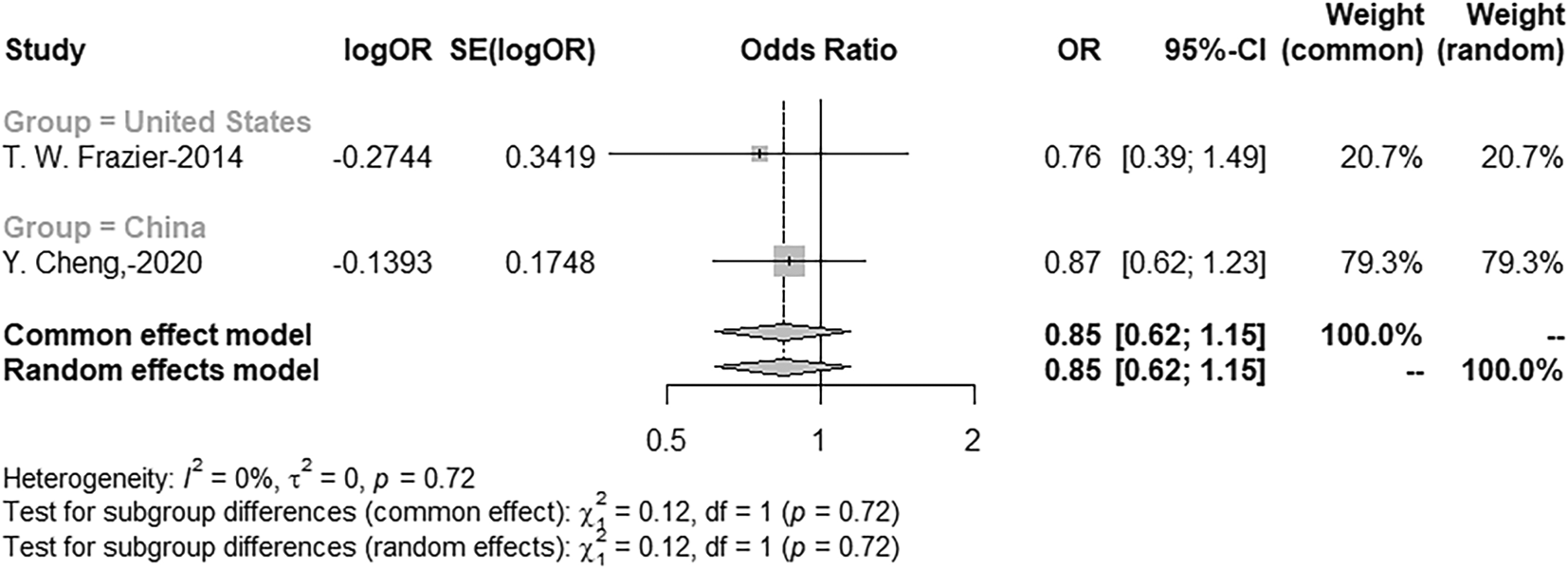
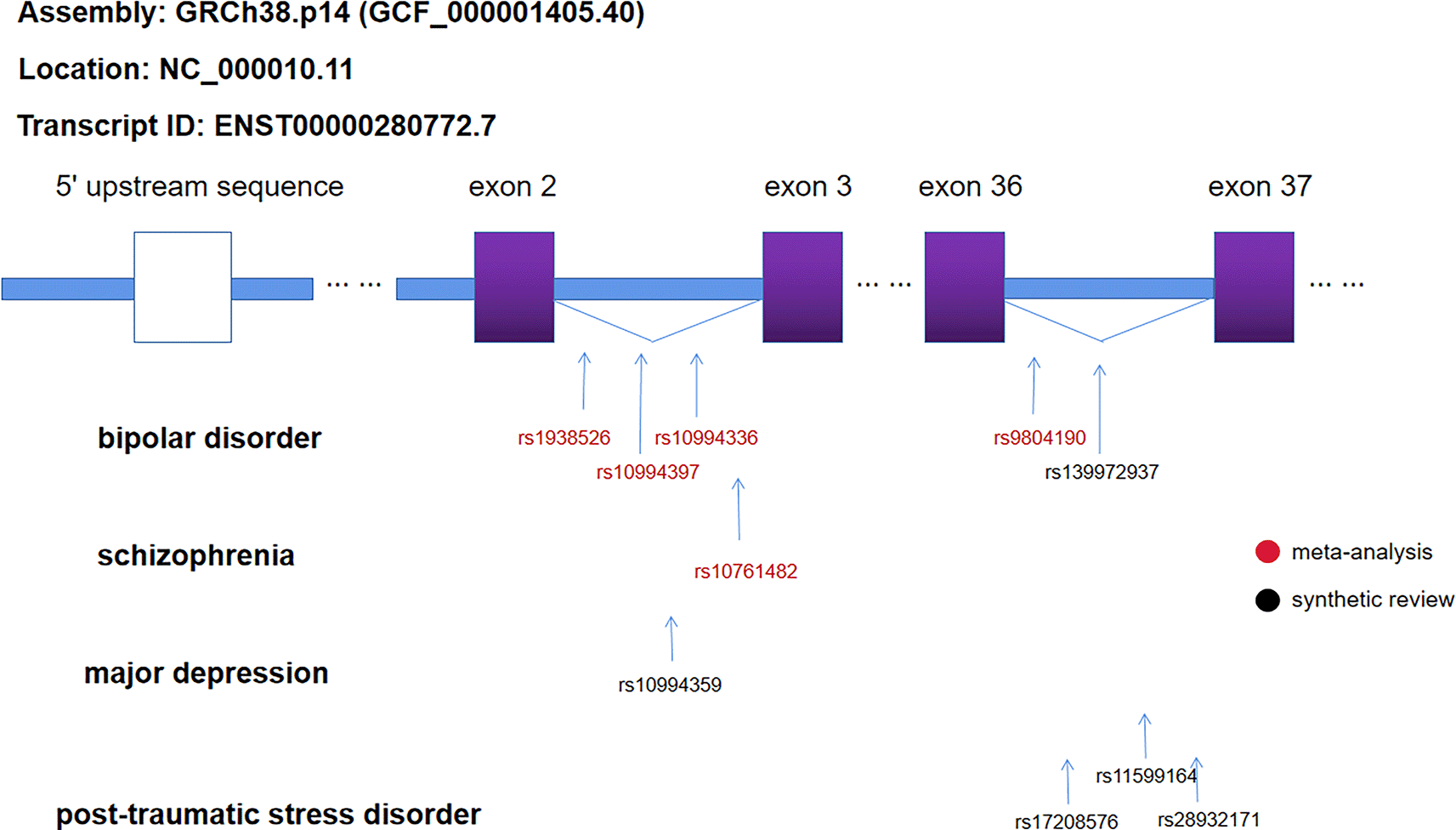
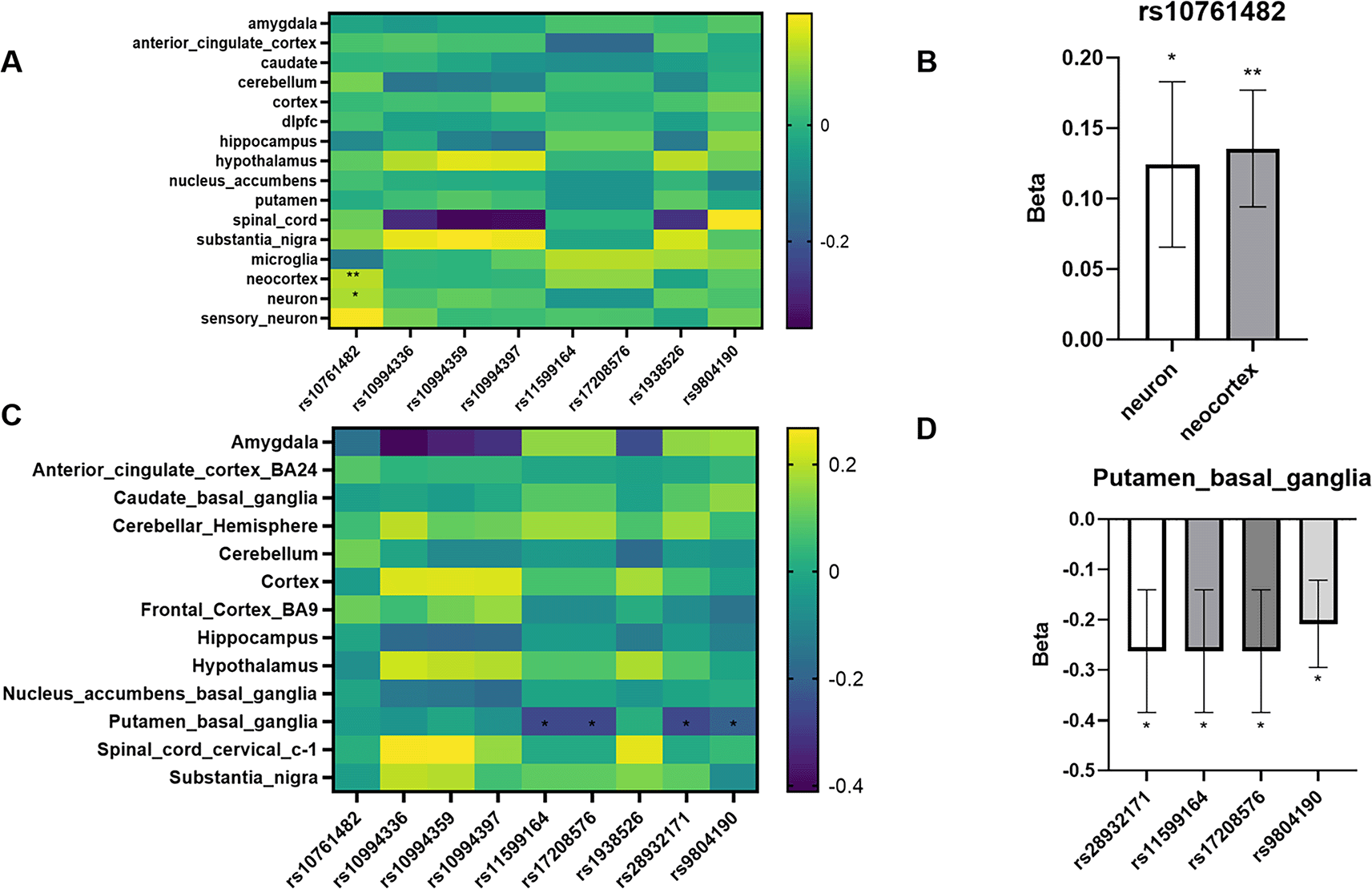
Comments on this article Comments (0)