Keywords
Formulation of mathematical model, Existence of equilibrium points, Local stability analysis, Globally analysis, Numerically simulation.
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Formulation of mathematical model, Existence of equilibrium points, Local stability analysis, Globally analysis, Numerically simulation.
Environmental sustainability is an important concept that aims to preserve the Earth’s natural and environmental resources to ensure the quality of life today and in the future, as it depends on achieving a balance between human needs, preserving the environment, and eliminating negative impacts on the ecosystem. Diverse activities and tasks across disciplines can be represented in a mathematical model that encompasses the diversity of studies across disciplines in the environmental sustainability system. This principle (sustainability) has become widespread and can be applied to all aspects of life on Earth, at different times, and in different places, such as forests, green areas, or wetlands. Healthy forests are among the most important examples of healthy and vital systems for sustainability.1–3 To provide healthy and vital forests to ensure environmental sustainability, the balance in the ecosystem must be studied through a mathematical model that examines organisms and their relationship with each other, especially the relationship of coexistence between living organisms, as well as reducing the severity of predation by predators on organisms that are considered easy prey. This was the basis of our study. Many researchers in the field of ecosystems and the study of cases of environmental stability and equilibrium have expressed the system using a mathematical model that studies the relationship between living organisms. These systems have become widespread in all sciences and disciplines, and include diverse sciences in the biological sciences, infectious diseases, physiology, environmental sciences, immunology, economics, chemical reactions, genetics, and pharmacodynamics.4,5 In addition to these systems, there are universally agreed upon principles that the relationship between living organisms (humans, animals, and plants) constitutes a key link in the chain of universal life. This relationship is represented by an interaction represented by a mathematical model, which is an important application of applied mathematics in the animal world; weak animals (prey) can become a major food source for other animals (predators). In this case, the role of mathematicians is to choose the model that achieves the principles of safety and environmental sustainability.6,7 This interaction is called a predator-prey relationship.8–11 Two different types of animals and organisms can also interact in a mutually beneficial relationship, known as symbiosis.12 An example is the nectar of flowering plants and bees that feed on it. Nectar is the food of bees, whereas bees pollinate flowers.13,14 The other interaction, which is one of the most important, can be represented by a single food source shared between two or more species of organisms competing for it or organisms competing for survival.15 This type can be described by a set of mostly nonlinear equations (perhaps algebraic or differential equations). Therefore, the science of dynamic behavior in biological systems has become dominant in modeling science over the last few years. One of the important issues that must be highlighted when studying the predator-prey model is that logistic growth reaches the highest carrying capacity in the prey community if the predator is absent, and gives complete equilibrium and an ascending growth curve for the prey in the absence of predators. For this reason, most modelers and biologists have devoted their attention and time to this species, studying it closely and among them.16–21 Hence, biologists have begun to use these models to help them understand and study the biological aspects of organisms under their natural conditions.22–25 Recently, many researchers have been interested in a specific issue, which is the fear of the prey and the effect of its fear of the predator, and discovered through mathematical results that this effect is very negative.26,27 In addition, a study was conducted in28,29 on the impact of direct killing and hunting on birds and invertebrates during their breeding seasons to clarify how predators are affected. A predator-prey model with non-local competition can produce complex dynamics, such as spatiotemporal patterns and spatially heterogeneous periodic solutions. See30 Ghanem, H. and Majeed studied the dynamics of a predator-prey model including two types of epidemics with harvest. The basic structure of a mathematical model with a biological meaning and character that can be studied accurately and carefully, based on the most important members of the population, is the prey and predator, which is described as the density of the prey consumed per unit of time by the predators.31–33 Latest I have come up with Ghanem, H. and Khaleefah, K in,34 a special study on the prey and predator patterns of Lake Victoria tilapia, snook, and coptodon, and recommendations for their conservation from overfishing. The study in this research is based on proposing a mathematical environmental (biological) model for living organisms, achieving a large part of the principles of environmental sustainability by creating the necessary conditions for coexistence and exchange of benefits between organisms that were considered prey for the purpose of controlling their extinction, which causes the death of a large part of the environment and an effective resource in the environmental balance, which consists of a system of differential equations. The local and global stability were analyzed and discussed, and the results were analyzed using numerical simulations related to the conditions of equilibrium and environmental sustainability. The results were positive for the purpose of providing a sustainable ecosystem.
Consider the proposed cooperative ecological mathematical model of three organisms, which involves a predator feeding on two prey species in a symbiotic relationship and exchanging food and resources for survival. Lotka-Volterra considered the response function (Holling function of the first kind) as a response dynamic process. Therefore, the population density hypothesis represented by the symbols X(T), Y(T), and Z(T) is that the first, second, and predator feed on them at any time (T) respectively. The growth density of the two prey groups represented a positive natural growth rate for both species (logistic growth). The relationship between organisms was described according to their effect on each other, where coexistence was considered a close relationship between the first and second prey, and the third organism was considered a predator of the first two species, in addition to the intense competition within each community, which may have been for food or for females during the mating season. Therefore, we describe how population density changes over time for the proposed species by partitioning the system into a set of three independent ordinary differential equations. Based on this model, see the following system:
With initial values of variables (X(0) ⩾ 0, Y(0) ⩾ 0, Z(0) ⩾ 0)
The Table 1 describes the basic parameters of the model (1).
We study the initial conditions of model (1) in space and show that the functions of model (1) are continuous and differentiable on , thus, the Lipschitz condition is satisfied, which is proof of the existence and uniqueness of model (1). In addition, all solutions of model (1) with its nonnegative initial condition are completely determined, as shown in the following theorem:
The solutions to Model (1) are uniformly bounded.
Let be any solution to model (1), and define the function.
We differentiate the function with respect to time and then the time derivative of over all of the form (1) yields:
Where and
Now, based on the Gronwell theory, gets that:
This gives: Therefore, the proof is complete, because it is independent of the initial conditions.
The study in this part is concerned with searching for the existence of possible and expected (fixed) equilibrium points for Model (1), where it was noted that no more than seven points are the possible equilibrium points.
1- The point that always exists is the simple equilibrium point, = (0, 0, 0).
2- The point called the equilibrium point of the axial = ( , 0, 0) where exists if the following condition is met:
This point is present if we adopt conditions and in addition to the following condition:
is found based on condition (3) add the following condition:
is found based on condition (2) add the following condition:
is found based on conditions (2), (3) and (4) add the following condition:
The stability analysis of model (1) is dealt with in this section for all stability points, and by calculating the Jacobian matrix and studying all cases of convergence and stability depending on the resulting eigenvalues to compensate for the points of model (1) that were obtained, where the matrix can be written as follows:
System (1) contains a distinct equation for matrix close to in the following form:
Consider Equation (9) based on condition (2) we get by (3) we have that and unstable point.
The Jacobian matrix for equilibrium point in model (1) is written as follows:
The following equation is the characteristic equation of and is defined as follows:
Where , ,
Consider Equation (10) based on condition (3) we get and
Without the need for an eigenvalue indication , (Saddle) unstable point.
The Jacobian matrix for equilibrium point in model (1) is written as follows:
The following equation is the characteristic equation of and is defined as follows:
Consider Equation (11) based on condition (2) we get and (Saddle) unstable point.
The Jacobian matrix for equilibrium point in model (1) is written as follows:
The following equation is the characteristic equation of and is defined as follows:
According to the Routh-Hurwitz criterion, there are roots with a negative real part of Equation (12) if and only if
and based on the condition (4) add the following conditions:
We have that and with the above conditions.
And accordingly,
So, it is locally asymptotical stable, otherwise unstable.
The Jacobian matrix for equilibrium point in model (1) is written as follows:
The following equation is the characteristic equation of and is defined as follows:
Where , ,
According to the Routh-Hurwitz criterion, there are roots with a negative real part of Equation (15) if and only if
and if the following conditions hold:
We have that and with the above conditions.
And accordingly,
So, it is locally asymptotical stable, otherwise unstable.
The Jacobian matrix for equilibrium point in model (1) is written as follows:
The following equation is the characteristic equation of and is defined as follows:
Where , ,
According to the Routh-Hurwitz criterion, there are roots with a negative real part of Equation (18) if and only if
and if the following conditions hold:
Now,
We have that and with the above conditions.
And accordingly,
So, it is locally asymptotical stable, otherwise unstable.
The Jacobian matrix for equilibrium point in Model (1) is expressed as follows:
The following equation is the characteristic equation of and is defined as follows:
Where , , .
According to the Routh-Hurwitz criterion, there are roots with a negative real part of Equation (21) if and only if
and based on the condition (17) add the following conditions:
Now,
We have that and with the above conditions.
And accordingly,
So, it is locally asymptotical stable, otherwise unstable.
The comprehensive study and analysis in particular is studied in this section for the balanced points found in the previous section, called the local stability of model (1), and that is by using the Lyapunov function that satisfies the conditions of global equilibrium, as shown in the following theorems:
(Lyapunov stability theorem):
Suppose that be an equilibrium point of system (1.12) and be an C1 real-valued function defined on some neighborhood of with such that
⋄ and for all and (i.e., mean V is a positive definite function).
⋄ If in , then is stable.
⋄ If in , then is asymptotically stable.
Note that the function given above is known as the Lyapunov function (weak Lyapunov function) if and only if the first and second conditions hold, whereas it is known as a strict Lyapunov function (strong Lyapunov function) if and only if the first and third conditions hold. In addition, if can be chosen to be all of , then is said to be globally asymptotically stable if the first and third conditions hold.
Assume that = ( , , 0) is locally asymptotically stable. Then, point is globally asymptotically stable in space under the following conditions:
Choose
We differentiate the function directly with respect to time t and get:
Depending on the biological environmental facts, which are ( ) and ( with condition we obtain the following:
Under condition is accepted, and it turns out that is definitely negative depending on the local condition; thus, is a Lyapunov function with respect to , and is close to global asymptotical stability.
Assume that , ) is locally asymptotically stable. Then, point is Globally asymptotically stable in space under the following conditions:
Choose
We differentiate the function directly with respect to time t and get:
Depending on the biological environmental facts, which are ( ) and ( with condition we obtain the following:
Under condition is accepted, and it turns out that is definitely negative, depending on the local condition; thus, is a Lyapunov function with respect to , and is close to global asymptotical stability.
Assume that is locally asymptotically stable. Then, point is globally asymptotically stable in space under the following conditions:
Choose
We differentiate the function directly with respect to time t and get:
Depending on the biological environmental facts, which are ( ) and ( with condition we obtain the following:
By condition is accepted, and it turns out that is definitely negative depending on the local condition; thus, is a Lyapunov function with respect to , and is close to the global asymptotical stability.
Assume that is locally asymptotically stable. Then, point is globally asymptotically stable in space under condition (24) with the following conditions:
Where
Choose
We differentiate the function directly with respect to time t and get:
Now, depending on the biological environmental facts, which are ( ) and ( under conditions we obtain the following:
Under condition is accepted, and it turns out that is definitely negative depending on the local condition. Thus, is a Lyapunov function with respect to , and is close to the global asymptotical stability.
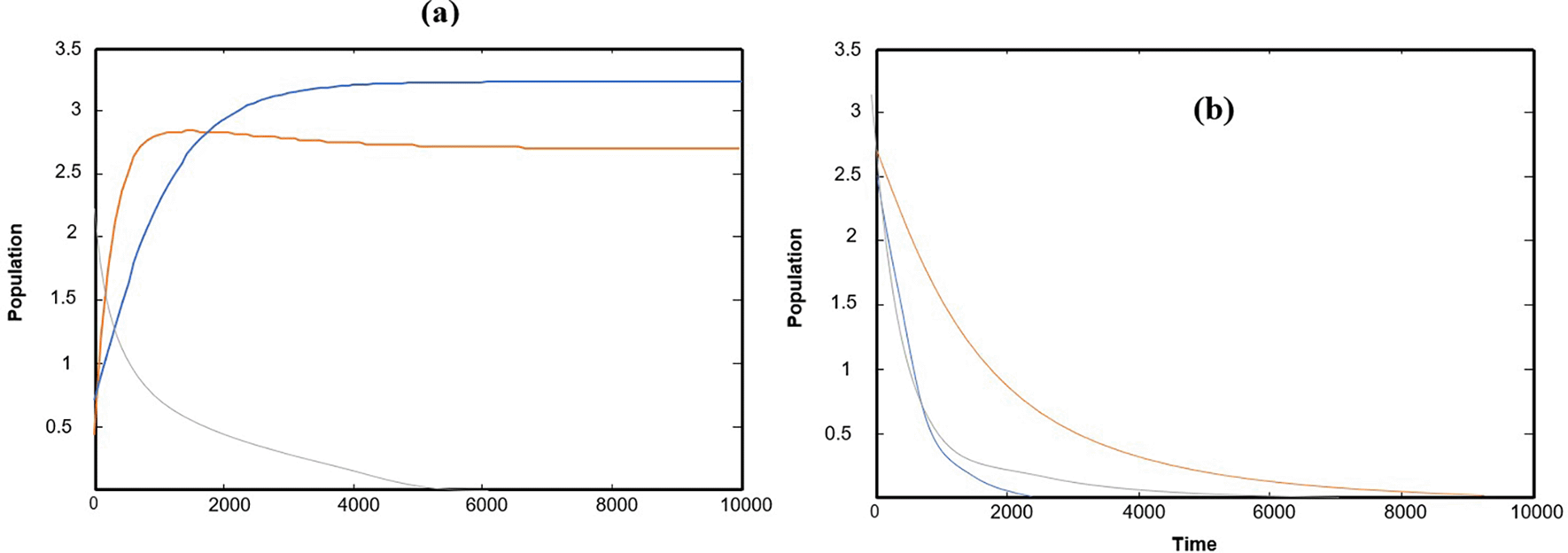
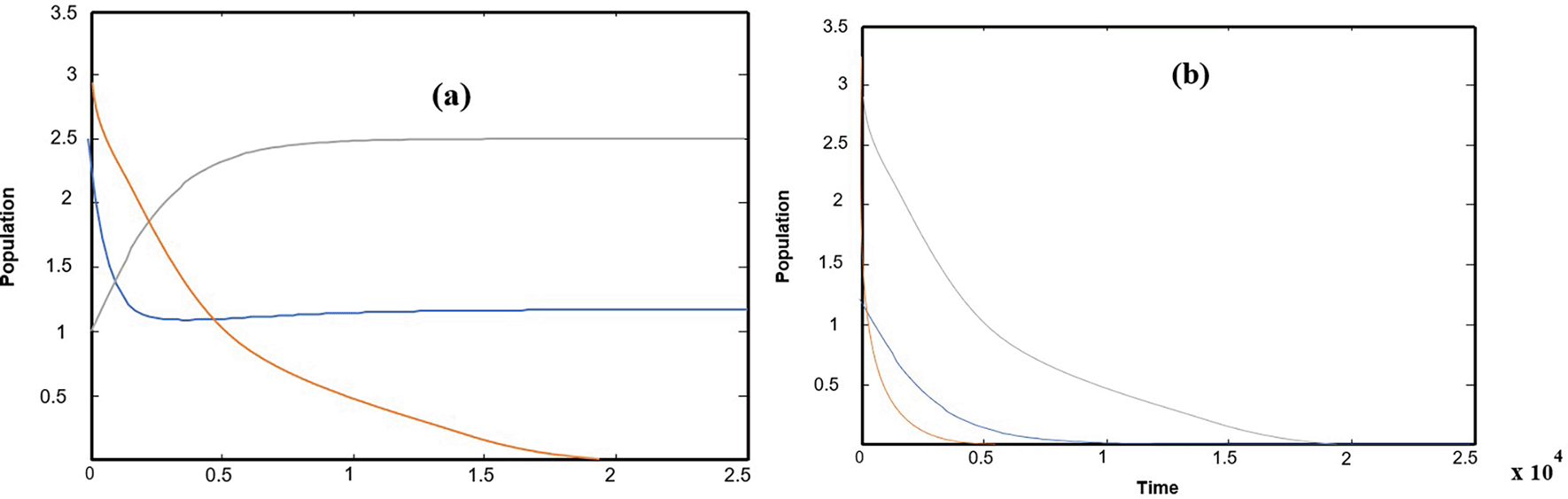
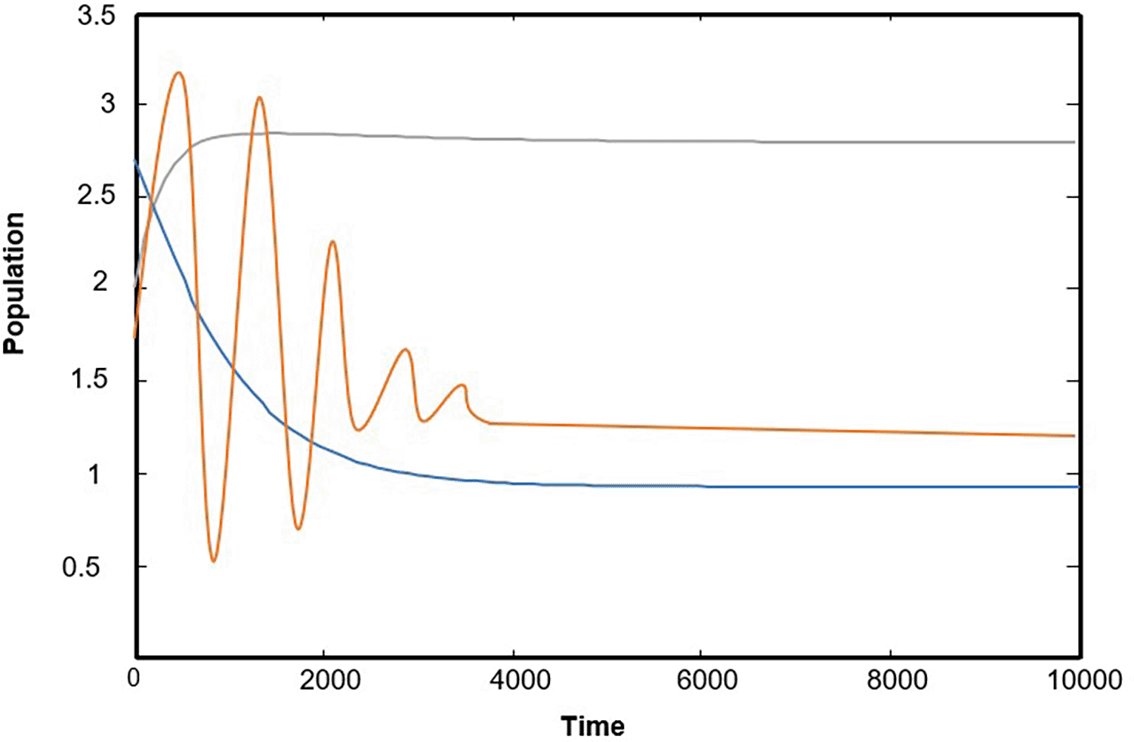
Numerical solutions using numerical software programs confirm the solution and provide an important interpretation for each point along the solution path (the equilibrium points of the system). We relied on the numerical solution using plotting programs in (matlab) and (C++), and the results were completed after providing approximate values for the key parameters in the system of differential equations. In the numerical interpretation of the paths and changes that occur in dynamics and stability, we show the stages of change that occur in the ecological community over time. As shown in Figure 1 that the population growth of the community (whether predator or prey) exhibits a state of global stability with more than one equilibrium point, and the results are convergent. This explains why predation by the predator is natural, and the population density in the prey community has caused it to grow naturally and steadily ( Figures (a), (b), and (c)), which clearly represents ecological sustainability; however, if we observe Figures 2, 3, and 4, we will see in each Figure that there is a complete absence and decline of one of the main ecological communities in the model, and this situation indicates that the increase in the predator community is the reason for the complete absence and predation of all organisms present in one of the other two types of prey communities. Likewise, Figures 5 and 6 show that in the community, there is only one organism that is still present in its ecological community: the prey organism. The complete absence of the predator in this case can be explained by the scarcity of its food and the intensity of competition within its ecological community. As for the last drawing, which is drawing Figure 7 it represents environmental evidence of the presence of the three organisms in the environment, complete stability in society, balance, and global convergence resulting from the availability of conditions for environmental coexistence, food sources for prey, and the abundance of sufficient food for predatory animals. All Figures are drawn to provide values for the parameters, and these values are approximate, as shown in the following sequence:
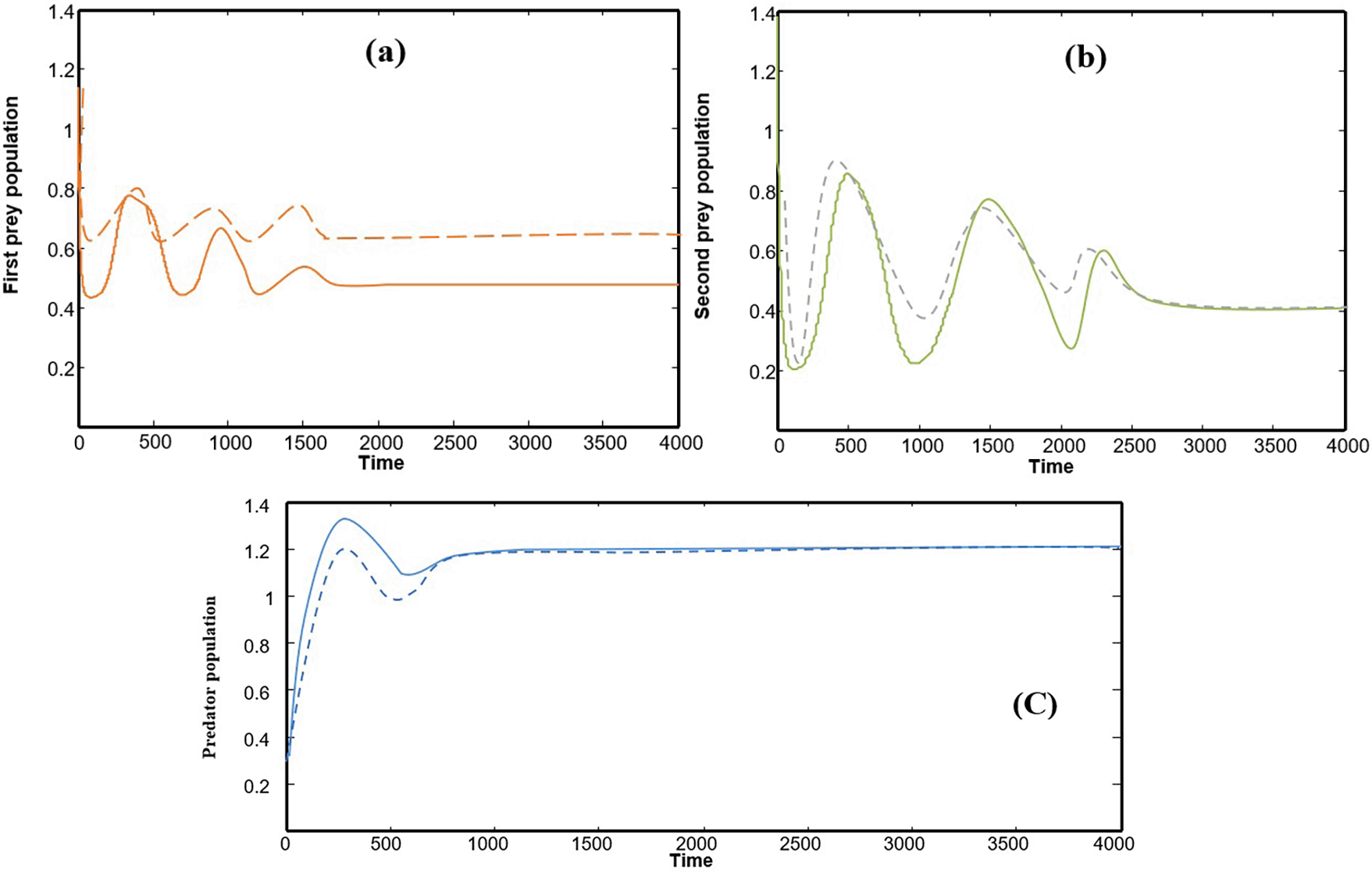
(a) Function of the time in which the path of the first prey is X, (b) Function of the time in which the path of the second prey is Y, (c) Function of the time in which the path of the predator is Z. Figure 1 express model (1) has an asymptotically of globally stable and the solution oncoming asymptotically point asymptotically from starting tow initial of different points.
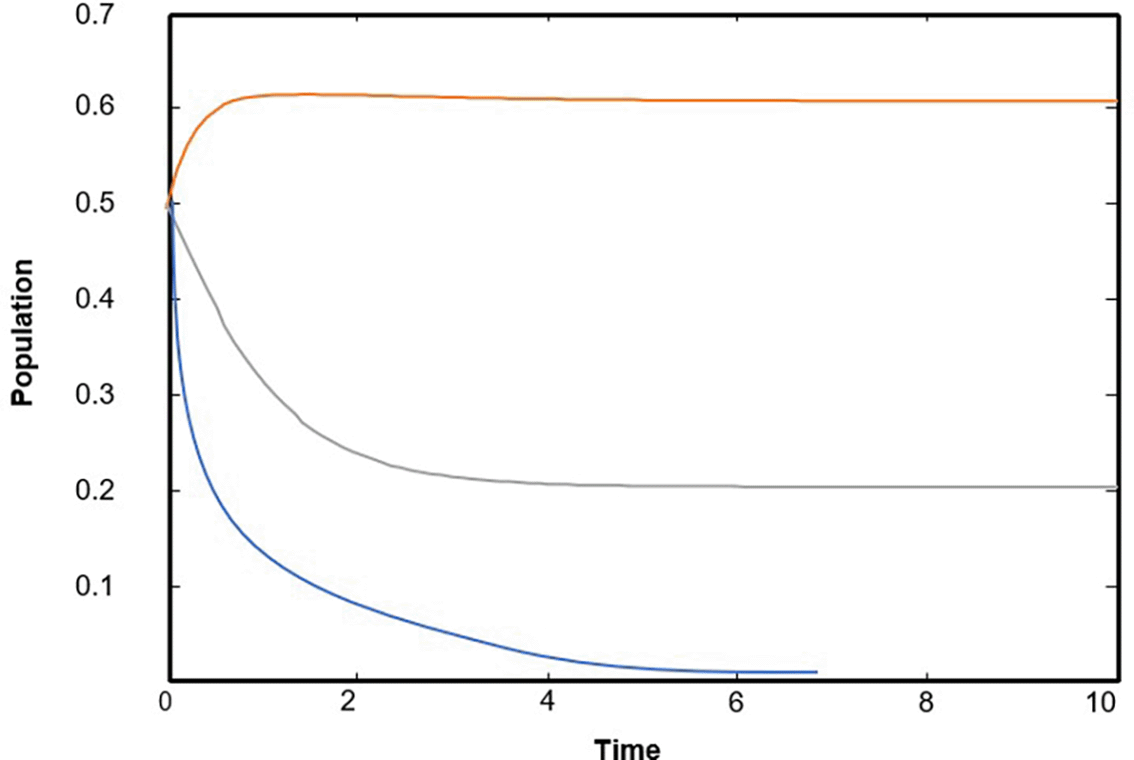
The varying of commensal relationship between the first prey X and the second prey Y in the intervals and ) the rest of parameters are keeping as data in (32) observed that the model (1) has a solution which is approaches as asymptotically to , as shown in Figure 2, for typical value ( 0.5 and 0.6).
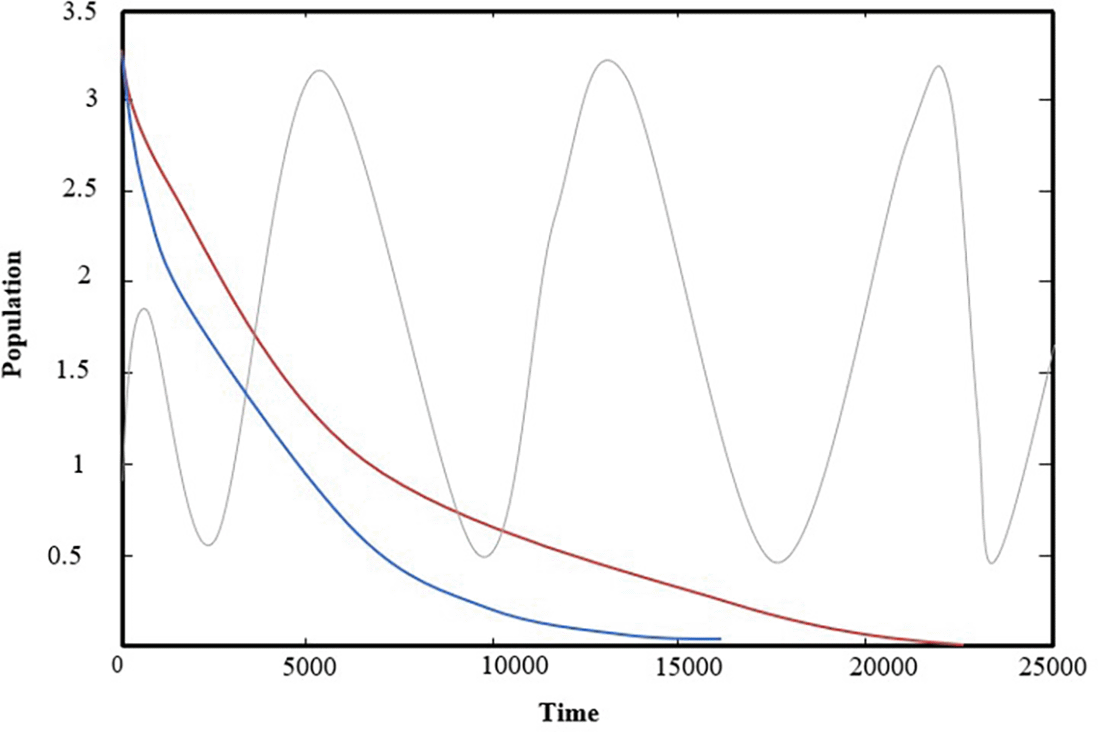
Now, varying the predator’s attack rate on the second prey and the break of factors are kept as data take in (32). it is notice for 0.08 ≤ the solution of (1) approaches , as shown in Figure 3(a), for typical1value = 0.095, if we increase the parameter to 0.651 ≤ , it is observed that system (1) asymptotically approaches So = 0.651 is a bifurcation point, as shown in Figure 3(b), for typical1value = 0.7.
The change parameter , represent the predator’s attack rate on the first prey, and the break of Factors are kept as data take in (32), it is noticed that for 0.003 ≤ the system (1) has a solution approaching as shown in Figure 4(a) for typical1value if we increasing the parameter for ≤ , it is observed that system (1) asymptotically approaches So = is a bifurcation point, as shown in Figure 4(b), for typical1value = 0.527.
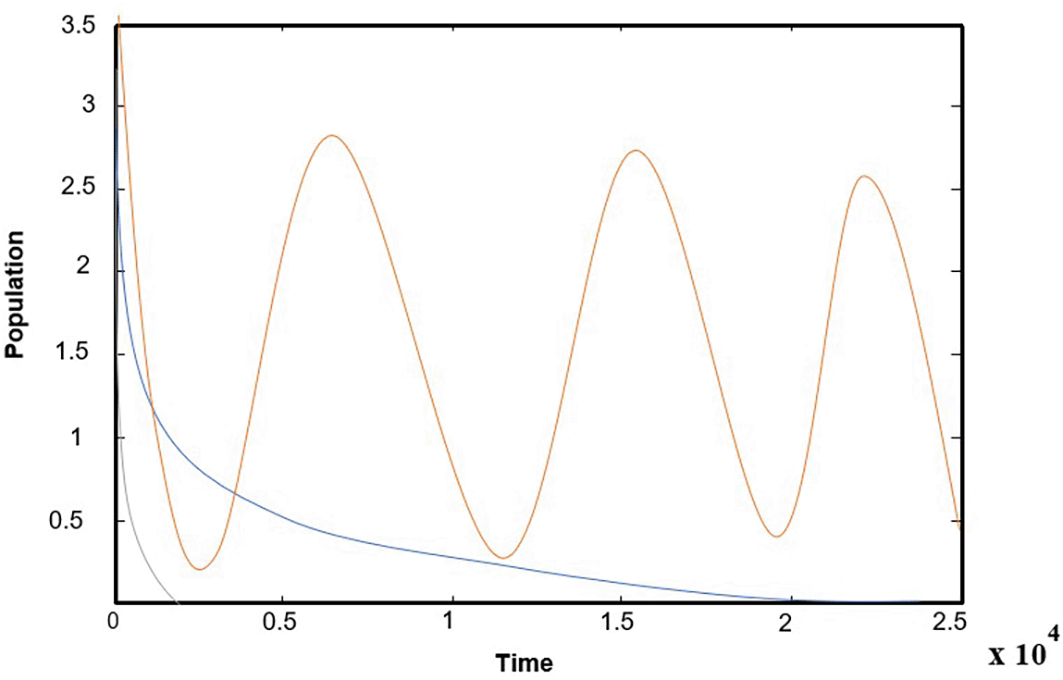
The effect of the natural growth rate of the second prey in the interval and the rest of the parameters are kept as data in (32), system (1) has a solution that asymptotically approaches as asymptotically to , as shown in Figure 5, for a typical value .
The effect of the natural growth rate of the first prey in the interval and the rest of the parameters are kept as data in (32), system (1) has a solution that asymptotically approaches as asymptotically to , as shown in Figure 6, for a typical value .
The change of the parameter , the rate of Food conversion from first prey to predator, and the break of Factors are keeping as data take in (32) it is notice for 0.05 ≤ < 0.2 the system (1) have a solution approaches if we increase the parameter for ≤ , it is observed that the system (1) asymptotically as approaches to which has already been drawn above So = is a bifurcation point as shown in Figure 7, for typical1value = 0.06.
The change of the parameter , the rate of Food conversion from second prey to predator, and the break of Factors are keeping as data take in (32) it is notice for 0.03 ≤ < 0.15 the system (1) have a solution approaches if we increase the parameter for 0.15 ≤ , it is observed that the system (1) asymptotically as approaches to so = is a bifurcation point.
The effect of the parameters the natural rate of death for the three organisms, which are the first prey, the second prey, their predator, and the competition within the predator community, respectively, at all times, and the rest of the parameter keeping as data in (32), there is no effect on the dynamics of the behavior of model (1) and model (1) have a solution approach to
This study aimed to draw conclusions from an equilibrium model of an ecosystem consisting of three organisms. A predator that feeds on the other two organisms, which have a positive relationship with survival and growth, is the primary food source for the predator. Predation (attack) by a predator on both prey depends on the first type of Holling functional response as a function of predation. In addition to what has been presented, and using numerical, analytical, and simulation methods, it has become clear that the natural growth or self-growth of both prey species plays an important role in ecological balance. Their mutual support also leads to a positive exchange of benefits, and reduces the risk of extinction in the face of continuous predation. In other words, the ecosystem will not collapse if the coexistence relationship continues. Therefore, the percentage of natural death and competition that occurs within the predator community must be on a curve parallel to the percentage that occurs within the community of the two prey, as the two prey are assumed to have competition and natural death within the same community, and these percentages make the system have a balanced natural growth rate that does not harm the prey as long as the curve between them and the predators is parallel. The effects of all parameters are studied numerically on the dynamic behavior of system (1), and the trajectories in the typical figures that represent the solutions and according to the solutions of system (1) for the data given by (32), the conclusions are obtained.
1- Model (1) does not have periodic dynamics for the data set, as given in (32).
2- The variations in the values of the set parameters in Equation (32), model (1) asymptotically approaches the globally stable point, .
3- The effect of the parameters the natural rate of death for the three organisms, which are the first prey, the second prey, their predator, and the competition within the predator community, respectively, at all times, and the rest of the parameter keeping as data in (32), there is no effect on the dynamics of the behavior of model (1) and model (1) have a solution approach to
4- The effect of the natural growth rate of the first prey in the interval and the rest of the parameters are kept as data in (32) observed that system (1) has a solution that asymptotically approaches as asymptotically to .
5- The effect of the natural growth rate of the second prey in the interval and the rest of the parameters are kept as data in (32) observed that system (1) has a solution that asymptotically approaches as asymptotically to .
6- The change parameter represents the predator’s attack rate on the first prey, and the break of factors is kept as data take in (32); for 0.003 ≤ the system (1) has a solution approaching to if we increase the parameter for ≤ , it is observed that system (1) asymptotically approaches So = is a bifurcation.
7- Varying the predator’s attack rate on the second prey and the break of factors are kept as data are taken in (32), it is notice for 0.08 ≤ the solution of (1) approaches to , if we increasing the parameter for 0.651 ≤ , it is observed that the system (1) asymptotically as approaches to So = 0.651 is a bifurcation point.
8- The effect of the commensal relationship between the first prey X and the second prey Y in the intervals and ), the rest of the parameters are kept as data in (32) observed that model (1) has a solution that approaches asymptotically to .
9- The change of the parameter , the rate of Food conversion from first prey to predator, and the break of Factors are keeping as data take in (32) it is notice for 0.05 ≤ < 0.2 the system (1) have a solution approaches if we increase the parameter for ≤ , it is observed that the system (1) asymptotically as approaches to So = is a bifurcation point.
10- Finally, the change in parameter , the rate of food conversion from second prey to predator, and the break of Factors are kept as data taken in (32). It is noticed that for 0.03 ≤ < 0.15, system (1) has a solution approach if we increase the parameter for 0.15 ≤ , it is observed that system (1) asymptotically approaches so = is a bifurcation point.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)