Keywords
Wind speed, blooming plants-honeybees model, mutualistic relationship, Beddington-DeAngelis functional response, dynamical systems, stability analysis, bifurcation.
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Wind speed, blooming plants-honeybees model, mutualistic relationship, Beddington-DeAngelis functional response, dynamical systems, stability analysis, bifurcation.
The mutualistic relationship between blooming plants and honeybee populations is a crucial ecological interaction for the sustainability of both local ecosystems and global agricultural systems.1 Animal pollinators, including honeybees, provide pollination services through symbiosis, which is essential for the successful reproduction of approximately 300,000 plant species worldwide. In symbiosis, pollinators are vital for promoting plant cross-pollination, genetic diversity, and crop quality and yield, substantially contributing to agricultural productivity.2 Consequently, this is a critical research area in conservation biology and ecology. A significant number of studies have been undertaken regarding the symbiotic relationships between plants and pollinator systems.3 Hadani4 suggested that the symbiosis can be characterized by the Beddington-DeAngelis response function, which incorporates competition for resource exploitation among pollinators and the obligatory relationship between the plant and the pollinator. This relationship has been observed to maintain a steady state, provided that the initial population level is sufficiently substantial.5 Biswas et al. examined a plant–pollinator model to investigate the impact of predation on pollinator species. They conclude that the pollinator is at risk of extinction if the predation rate cannot be controlled. Moreover, this hypothesis has prompted the advancement of extensive studies examining the impacts of nectar theft and ants on the plant-pollinator system.6 The reduction of biodiversity is a widespread issue, although the decrease of pollinating insects is especially alarming due to its possible effects on plant reproduction.7 A recent report on the global reduction of honeybee and bumblebee populations has highlighted pollination’s ecological and economic significance.8 The reduction of pollinator species can be ascribed to various ecological and environmental reasons, including habitat loss, illnesses, climate change, pesticides, and predation.9–12 Invasive predators can significantly affect pollinators by diminishing their quantity, altering plant reproductive success, and undermining the plant-pollinator relationship.13
One factor that negatively affects the mutualism in plant-pollinator systems is high-speed wind.5,14–17 High winds disrupt flower fragrance messages, reducing honeybee attraction. Strong winds quickly distribute flowers’ aromatic chemicals in different directions, reducing their “scent signal”. As aroma dispersion reduces, honeybees’ ability to discover food sources decreases, resulting in fewer trips to flowers.18 In addition, flying in severe winds demands more energy for balance and advancement. Honeybees may stay in the hive or fly less to preserve energy. Pollination opportunities decrease as fewer flowers are visited. Strong winds can break flowers’ petals or open their parts abnormally, decreasing honeybees’ access to their reproductive organs. This temporal shift may diminish pollination when flowers are most pollinating. High winds can cause honeybees to crash, fall, or be blown off course, destroying the colony. Lost workers or persistent stress can harm the colony and its capacity to provide enough workers for visits.19
In this study, we discuss the effect of high wind speed on a three-dimensional blooming plant–honeybee–honey production mathematical model ( model) that takes into account saturated mutualism between blooming plants and honeybees by the Beddington–DeAngelis response function. The primary focus is to investigate the dynamics of the plant–pollinator system while considering the impacts of wind speed on the mutualism between blooming plants and honeybees. The study aims to understand the interactions between mutualism and wind speed, and how these interactions affect the overall ecological balance and sustainability of the blooming plant–honeybee–honey production system.
In this section, a model is formulated to describe the interaction among blooming plants , honey honeybees and the production of honey at time . Then the blooming plant, the honey honeybees, and the production of honey system can be depicted by
1. The blooming plants are assumed to grow in the absence of honeybees at the intrinsic growth rate , depletion rate and carrying capacity . Since blooming plants provide nutrients for honeybees, honeybees offer pollination services to blooming plants; hence, their relationship is mutualistic. Beddington-DeAngelis functional response can be used to express the mutualistic relationship, where denotes the positive effect of honeybees (a kind of pollinator) on plants, refers to the undepleted equilibrium rate for the blooming plant–honeybee interaction, which incorporates travel and unloading durations at a central location along with individual-level blooming plant–honeybee interactions, and indicates the intensity of competition among honeybees for floral resources.
2. High winds break plants’ stems and branches, causing flowers to collapse and damage their petals. This damage reduces the flowers’ attractiveness to pollinating insects. Also, wind speeds reduce the honeybees’ ability to search in a windy environment. Let be the efficiency of wind, which satisfied the following
3. The honeybee population are expected to grow from external resources at the intrinsic growth rate , depletion rate and carrying capacity . represents the nutrients that blooming plants provide to honeybees, where stands for the corresponding value of honeybee nutrients from blooming plants.
4. The term represents the honey production in the colonies where is the rate of production that is contingent upon the quantity of honeybees, and is the half-saturation rate. Wind speed negatively affects the amount of honey produced since it diminishes the honeybees’ capacity to search for nutrition.
5. During winter or drought, honeybees rely on their stored honey to survive. This represents a natural consumption process but diminishes the quantity available for harvest. Therefore, signifies the rate at which honeybees consume honey to survive.
6. The amount of honey produced decreases due to many factors, such as absorbing moisture from the atmosphere. In humid environments, honey absorbs moisture, which reduces its concentration and makes it susceptible to fermentation and spoilage. High temperatures cause the honey content to evaporate. Therefore, denotes the rate of natural causes of honey loss.
Further, the schematic sketch of system is illustrated in Figure 1.

Before analyzing our model, it is pertinent to invoke the following lemmas, the demonstration of which is available in Refs. 20-22.
(Comparison lemma) Suppose that with . Then for , then , and if , then .
Uniqueness.
Since the right side of the model , so they satisfy the Lipschitz condition. Therefore, the solution to model that starts in exists and is unique.
All solutions of system are uniformly bounded.
From the blooming plants equation of the system, we obtain
After using the honeybees’ carrying capacity, we get
Using Lemma 1, we deduced that.
Similarly, for the honey honeybee equation of the system, we find
Finally, from the amount of honey produced equation, we attain
Then, by Lemma 2, we get
Therefore, any solutions of system will be attracted to .
The system is said to be persistent if all its components survive in future times.
First, to prove that If it is persistent, we have to show that i.e. will not decay to zero.
From the blooming plants equation, we get
By Lemma 2, we have , where provided . Thus, for a small , a positive number such that
Applying the same strategy for the honeybee equation, we get
From the honey production equation, we attain
Since
Then, by Lemma 2, , where
Thus, for a small , a positive number such that
Therefore, system is uniformly persistent.
The possible equilibria of system are
1) The extinction point .
2) The blooming plants point , where . For to be positive, condition (2) must be satisfied.
3) The honeybee point , where . Clearly if condition (3) is satisfied.
4) The blooming plants free point , where and . Clearly if the condition (3) is satisfied.
5) The coexistence point , here , where,
and is the root of the following equation
By Descartes’s rule of signs, Equation (4) has a unique positive root , if one of the following conditions holds:
Further, if one of the following conditions holds:
To investigate the local stability, one needs to determine the Jacobian matrix at any point. Thus, the Jacobian matrix at any point is
Then, the eigenvalues of are , and .
Therefore, is asymptotically stable under conditions 5 and 6.
The biological interpretation of conditions 5 and 6 indicates system reaches asymptotically the extinction point when the depletion rates of the blooming plants and honeybees exceed their intrinsic growth rates.
So, the eigenvalues of are
, under the existence condition of the blooming plants point.
and .
Thus, is asymptotically stable if condition 8 is satisfied
So, the eigenvalues of are , and . Thus, is asymptotically stable if the conditions 10 and 11 are satisfied.
The eigenvalues of are , , and . So, is asymptotically stable if conditions 13 and 14 are satisfied.
In this section, the Lyapunov method is used to illustrate the global stability of the previous points, as shown in the following theorems.
The extinction point is a global asymptotic stability (GAS) if the following conditions are met.
Let , where , which satisfies and for all with . Then
Then, by using the upper bound of the honeybees’ population, we get
The first two terms are negative definite if conditions 20 and 21 are satisfied. Hence, is a negative definite. Therefore, the extinction point is GAS.
Let , where , which satisfies and for all with , then
Then, by using the upper bound of the blooming plants population, we get
The first two terms are negative definite if conditions 22 and 23 are satisfied. Hence, is a negative definite. Therefore, the blooming plants point is GAS.
Let , where , which satisfies and for all with , then
Then, by using the upper bound of the honeybees’ population, we get
The first three terms are negative definite if conditions 24-26 are satisfied. Hence, is a negative definite. Therefore, the blooming plants point is GAS.
The blooming plants free point
Let , where , which satisfies and for all with , then
Then, by using the upper bound of the honeybees’ population, we get
The first and the third terms are negative definite if conditions 27 and 28 are satisfied, while the second term is negative under condition 24. Hence, is a negative definite. Therefore, the blooming plants free point is GAS.
This section explores the probability of occurrence of transcritical (TB) and Hopf bifurcation (HB) around the non-hyperbolic equilibrium points. For more details, see Refs. 23-26.
For , the model faces TB at the extinction point .
According to given by Eq. (7), the system at has a zero eigenvalue , at , and at becomes
Now, suppose that , and be eigenvectors to of , and , respectively. The calculation gives ), and by solving , and for and .
Therefore, there is a TB around with the parameter .
According to given by Eq. (9), the system at has a zero eigenvalue , at , and at becomes
Now, suppose that and be eigenvectors with respect to of and respectively. Solving , and for , and gives and , where .
under condition 31, here, , , , ,
Therefore, there is a TB around with the parameter .
According to given by Eq. (11), the system at has a zero eigenvalue , at , and at becomes
Now, Suppose that and be eigenvectors to of and respectively. Solving , and for and gives and , where .
Therefore, there is a TB around with the parameter .
According to given by Eq. (15), the system at has a zero eigenvalue , at , and at becomes
Now, Suppose that , and be an eigenvector of to of and , respectively, which gives and .
Therefore, there is a TB around with the parameter .
The undergoes a Hopf bifurcation at the coexistence point to the bifurcation parameter if
The Jacobian matrix at with is given by
where , and .
The Hopf bifurcation occurred if the following conditions are satisfied.
where and are defined in the characteristic equation given by (19). Now, we set to find , which gives
Solving the above equation for , gives
The discriminant of the above equation is
That means condition (1) is satisfied at , and the characteristic equation given by (19) can be rewritten as
Solving the above equation yields , . Clearly and are complex conjugates if condition 34 is satisfied. In addition, the general roots of Eq. (19) in the neighborhood of as , then
So, system undergoes a Hopf bifurcation at with the bifurcation parameter .
A numerical confirmation is carried out to complete the analytical results for system using MATLAB. The simulations are conducted by using the following set of parameters.
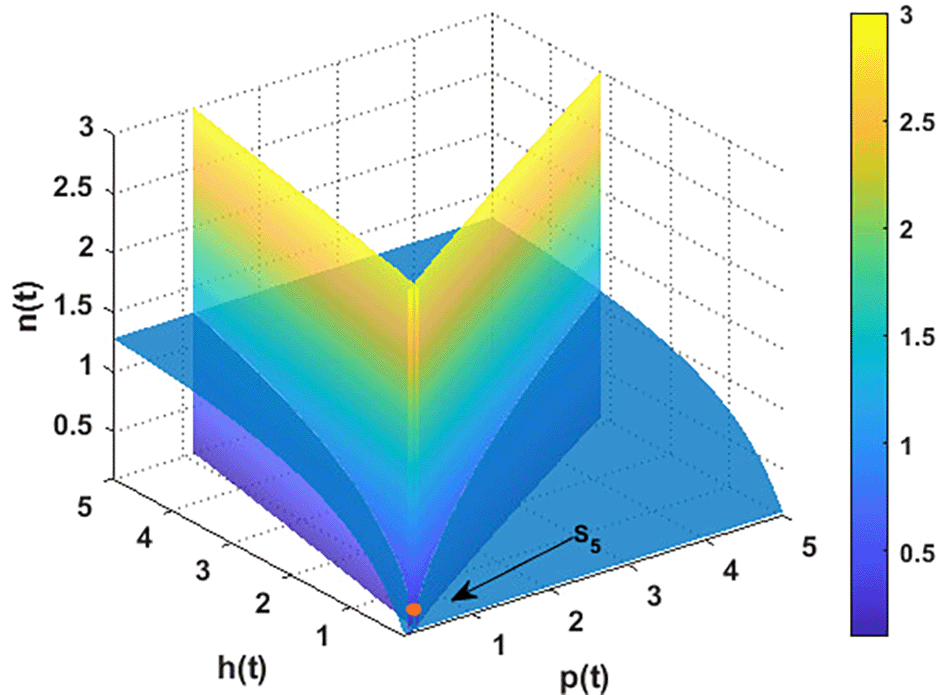
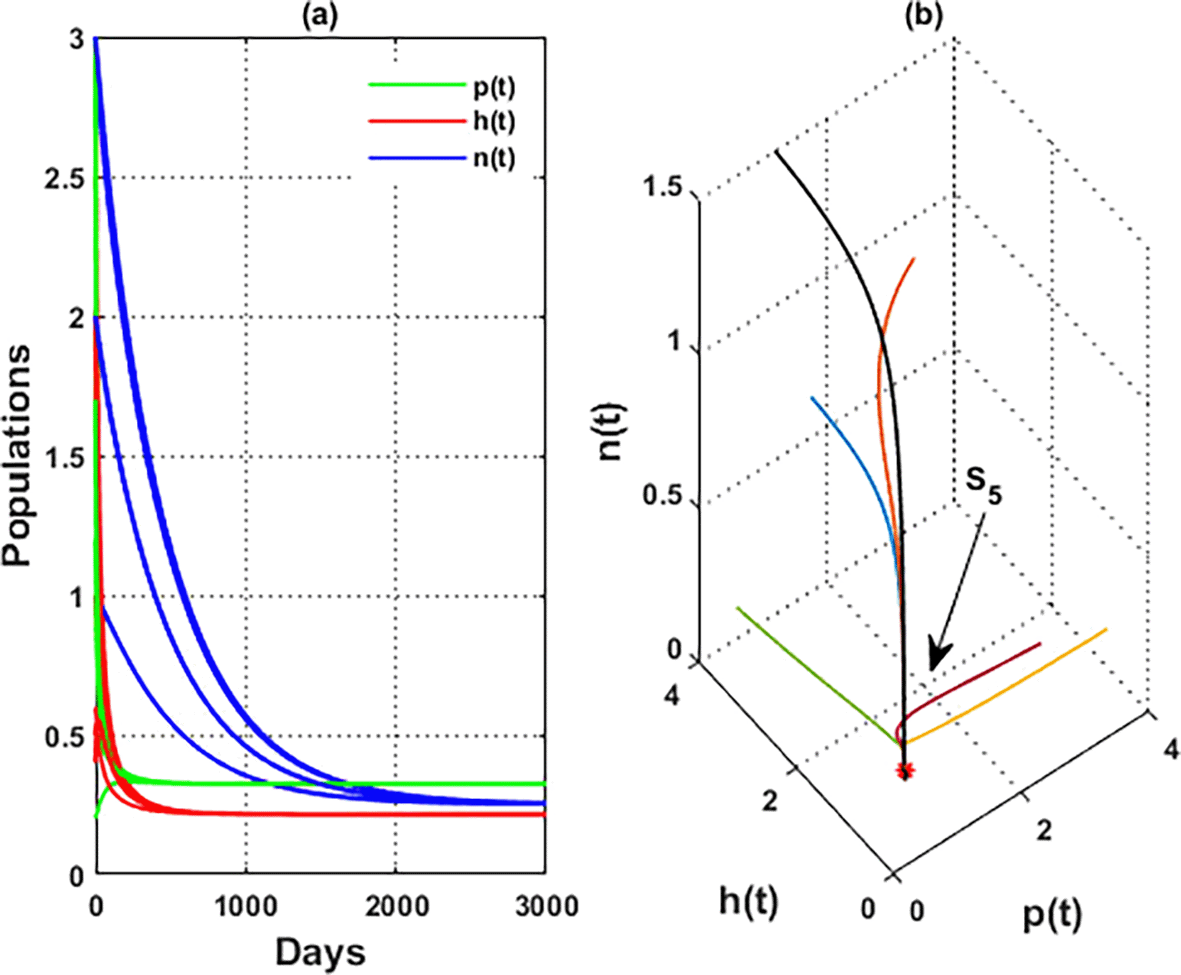
For the parameters listed above in Eq. (36), the nullclines of system are indicated in Figure 2. The figure depicts the coexistence point , while Figure 3 illustrates the global behavior of . That means the parameters listed in Eq. (36) corroborate the findings of Theorem 12, which shows that all other points act as saddle points, except for . These findings also confirm the uniform persistence for system, which confirms the output of Theorem 3.
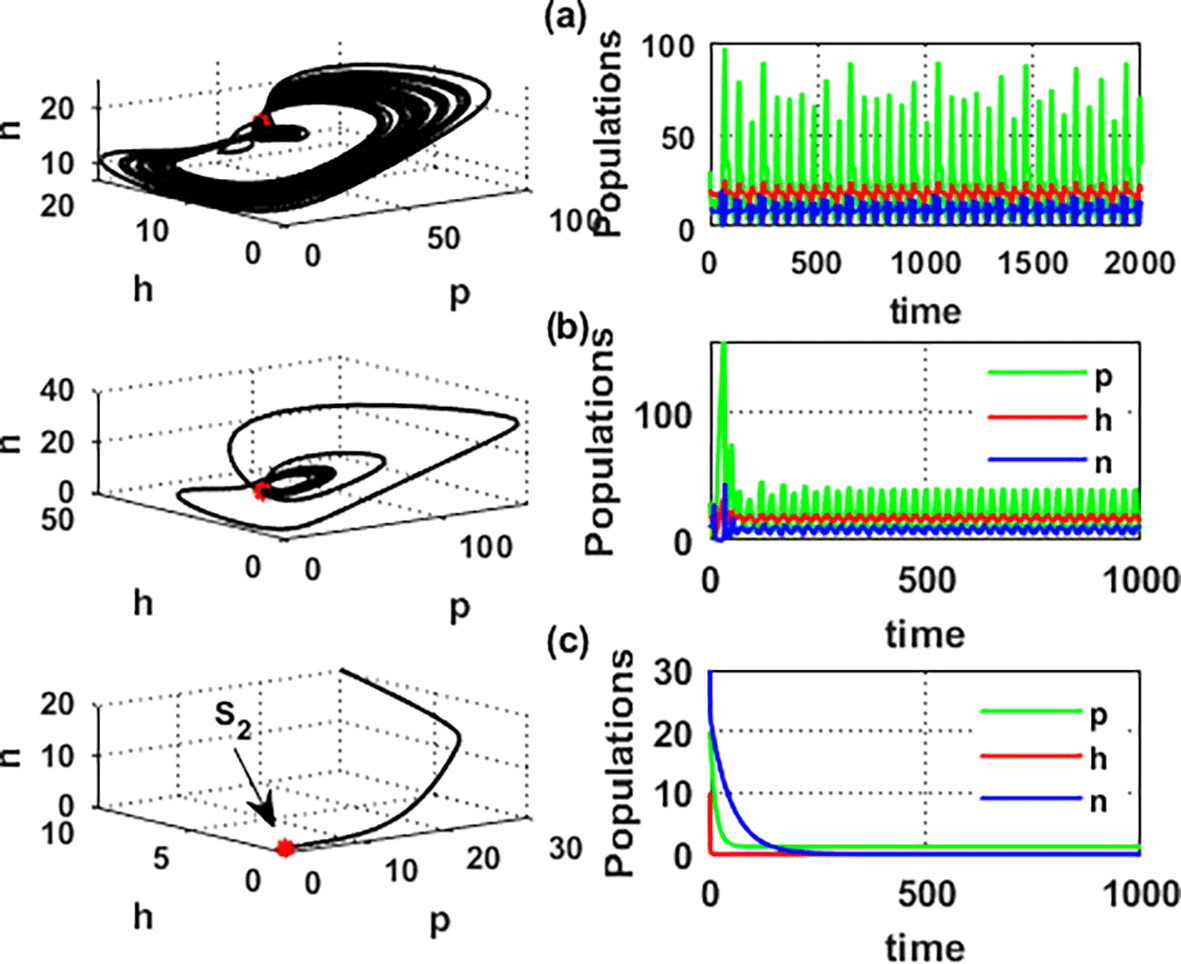
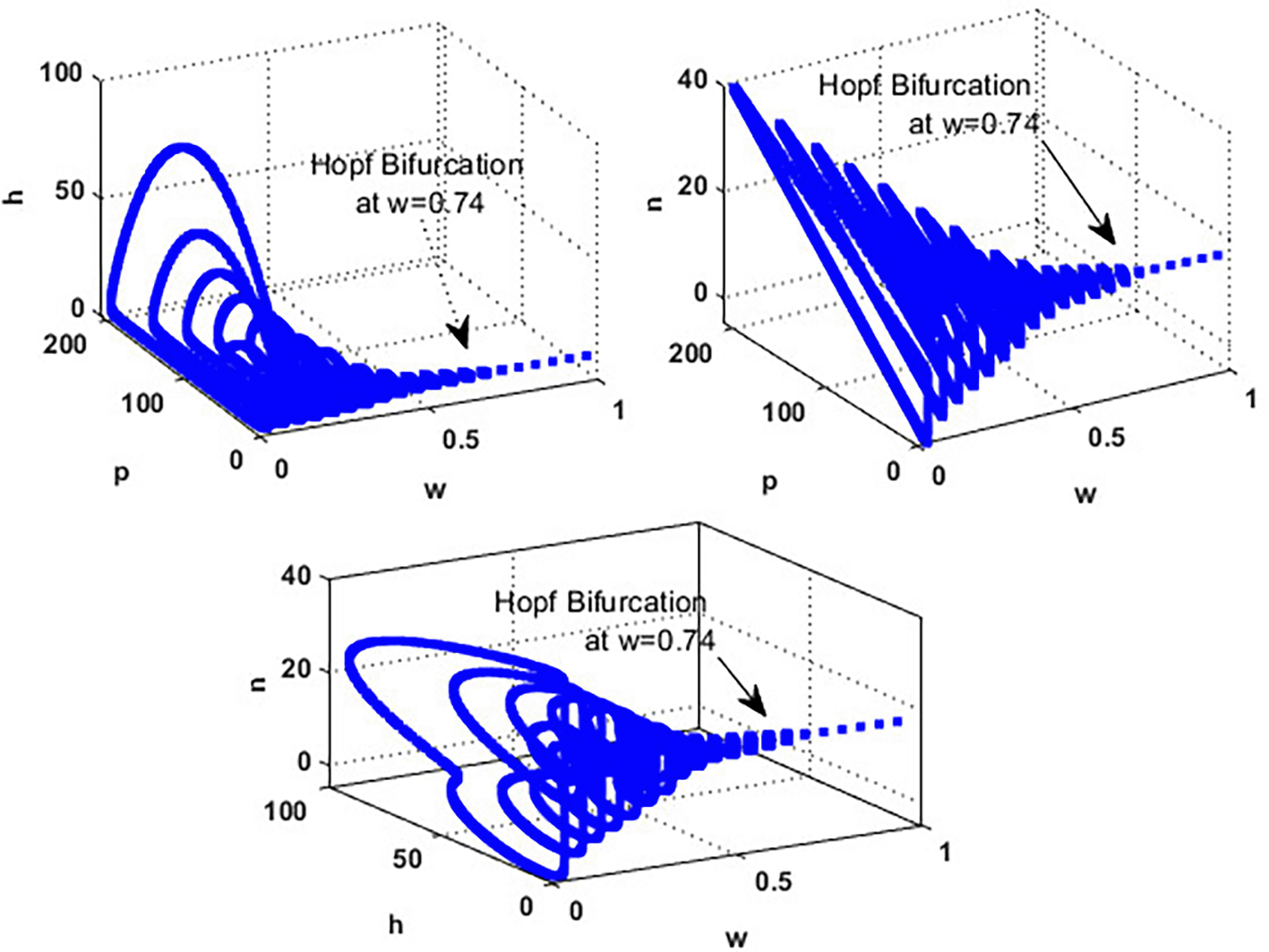
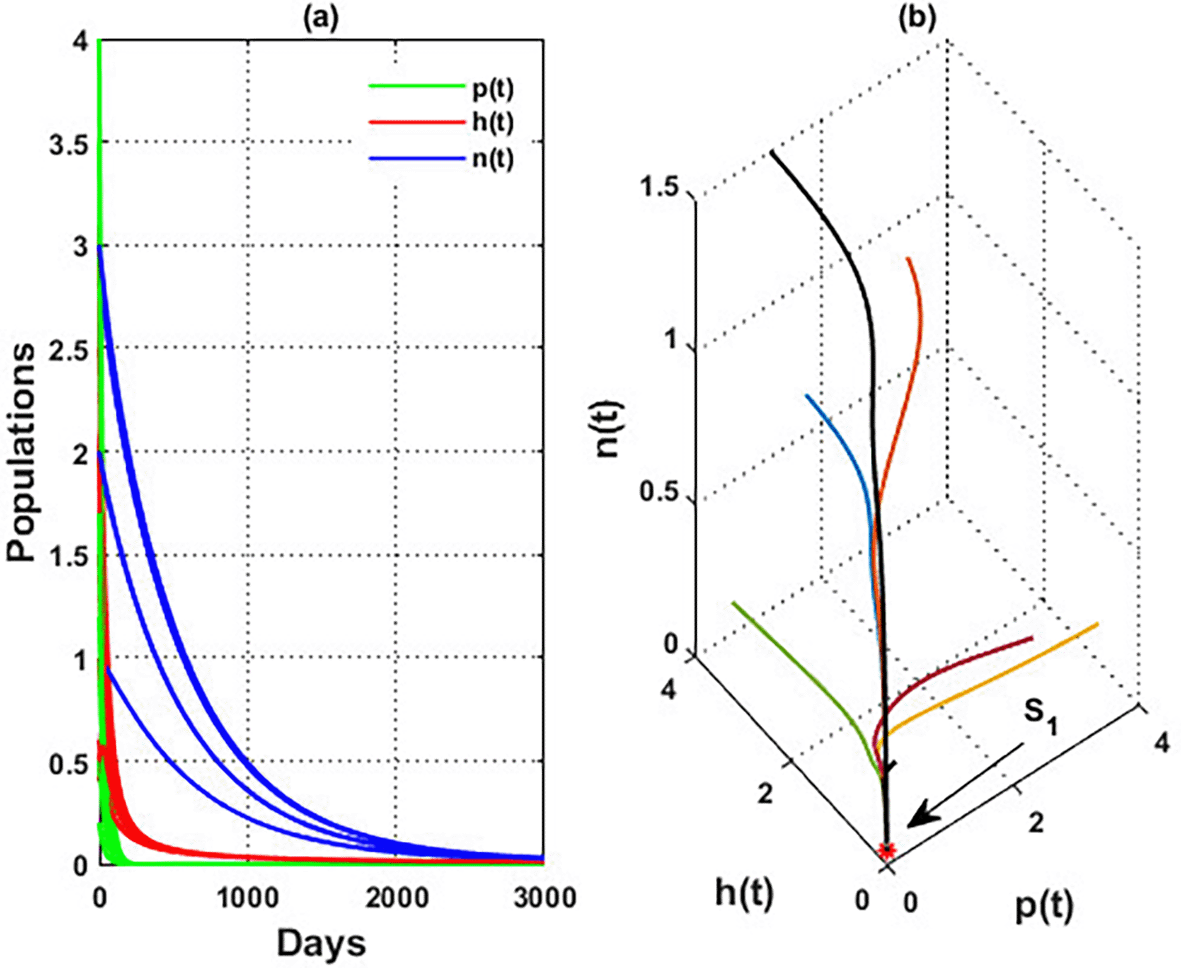
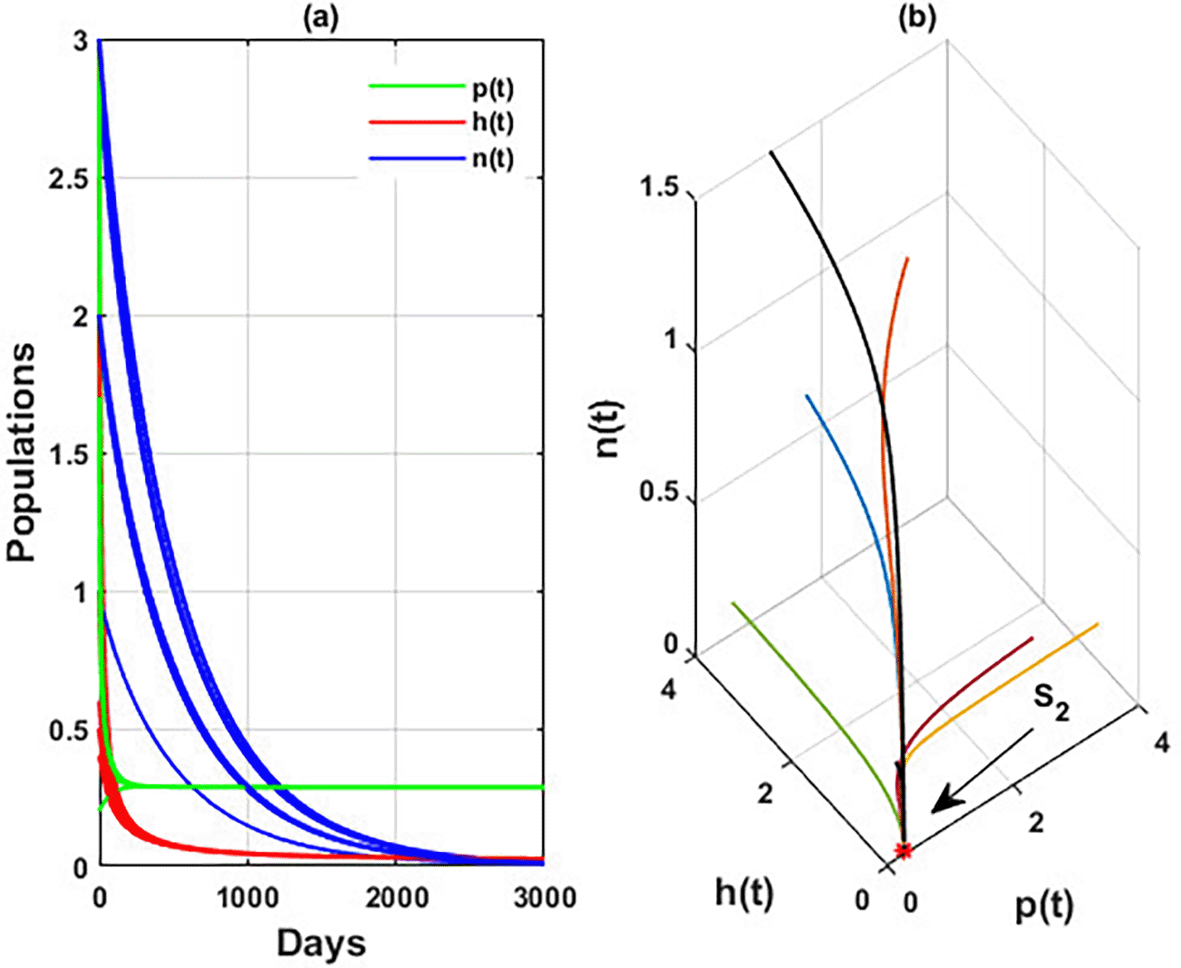
An important parameter to investigate is the effect of the wind level on the interaction of blooming plants , honey honeybees and honey production . We address two aspects of : first, the extent to which it influences the species densities in the inner equilibrium, and second, how it can alter system’s stability. For a low level of wind flow, i.e., the solution of the system approaches a chaotic attractor, see Figure 4(a). While in the interval , the solution of the system converges to a limit cycle, see Figure 4(b) and Figure 5. Consequently, for the region , the solution stabilized at the coexistence point , see Figure 2, which was plotted when . Finally, for a high level of wind speed, i.e., for , system loses two of its components, and the solution in this case settles down to the blooming plants’ point , see Figure 4(c). This indicates that when the wind speed is light, the wind helps pollinate or carries the seeds of flowering plants from one place to another, aiding their reproduction. Therefore, the presence of flowering plants contributes to the system’s persistence. In contrast, for a high wind flow, we see that the populations of honeybees and the production of honey face extinction.
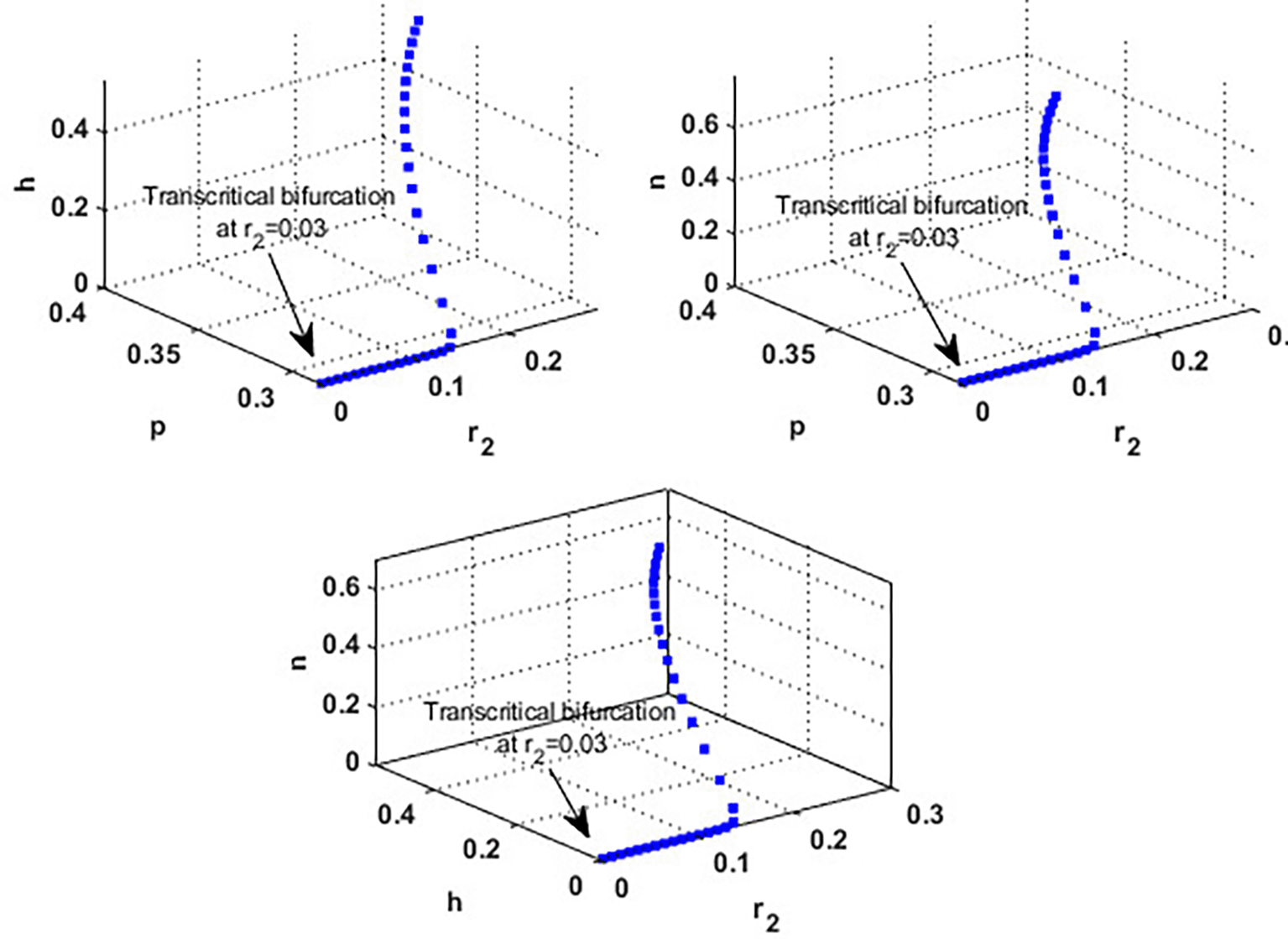
Now, we have the intrinsic growth rate of the honeybee population ( , which is an essential quantity to discuss since it can potentially influence the population’s densities of blooming plants, honeybees, and honey production. It is clear from Figure 6 that the solution of system settles down to the coexistence point for , while it stabilized at the extinction point for . This result confirms the occurrence of a transcritical bifurcation at , which confirms the output of Theorem 13, see Figure 7. Further, Figure 8 indicates the global behavior of the extinction point . This result confirms the global stability condition of which has honeybee stated in Theorem 8. Further, this result shows that is a critical parameter that impacts the continuity of the whole system’s coexistence.
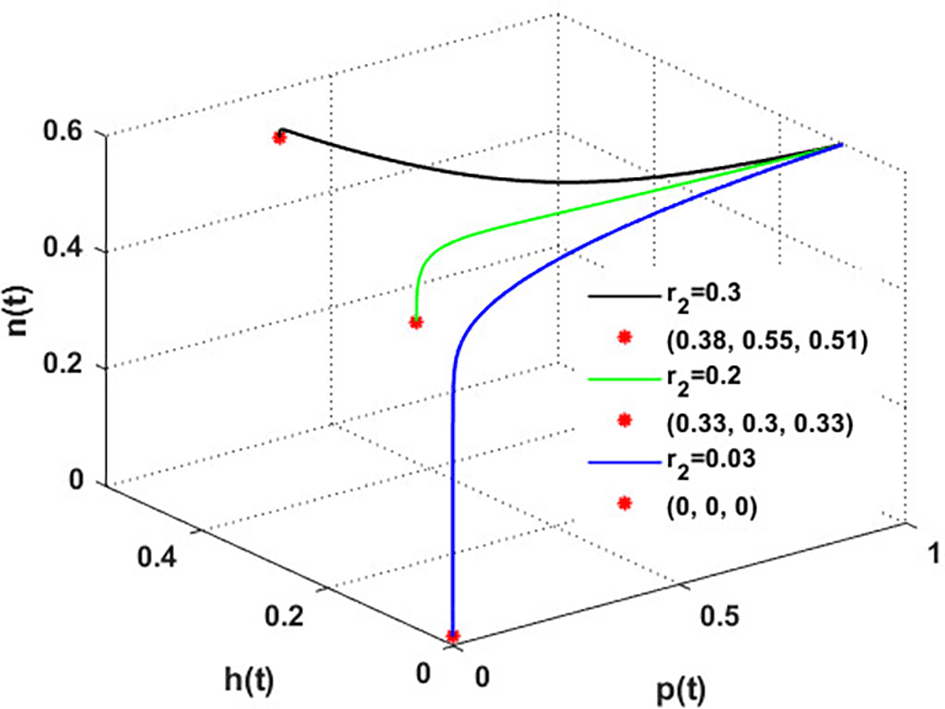
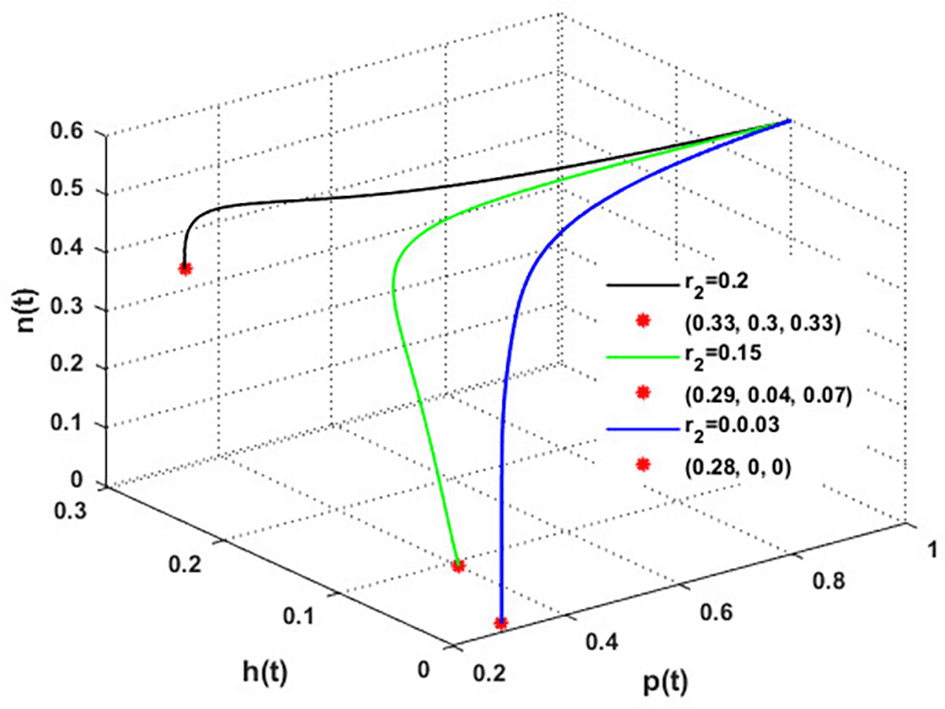
The effect of varying the intrinsic growth rate of the population of blooming plants is investigated in Figure 9. The figure indicates that the solution of system converges to the blooming plants point for , which means the system faces an occurrence of transcritical bifurcations at = 0.04, which confirms the output of Theorem 14. So, system loses two of its components for . While for , system converges to the coexistence point . Further, the global stability of is illustrated in Figure 10. We can conclude that the intrinsic growth rate of the blooming plants population is a critical parameter affecting the honeybees’ persistence and honey production.
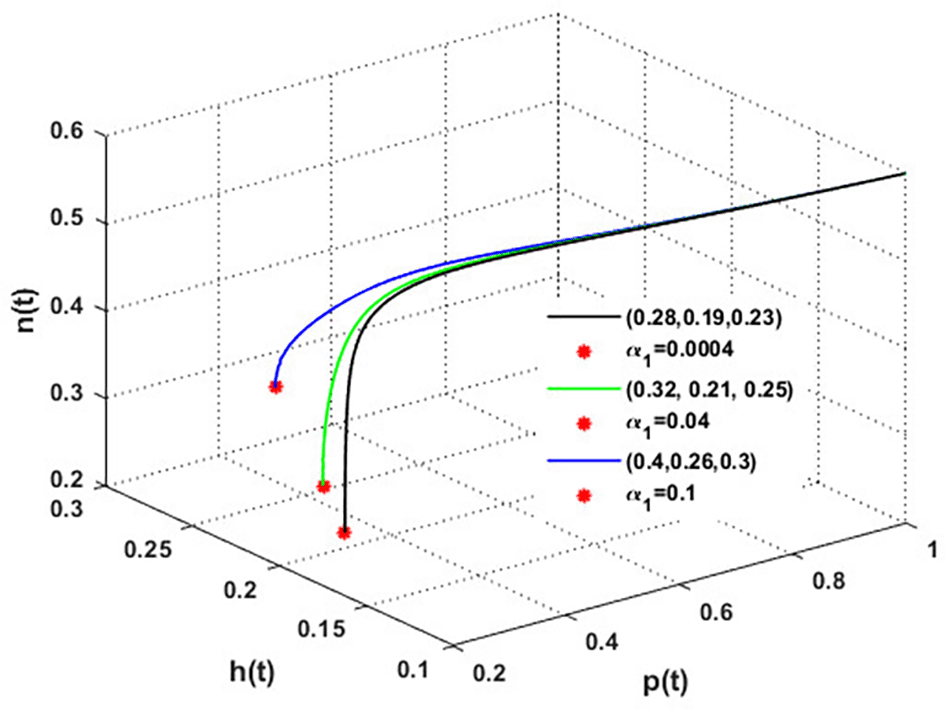
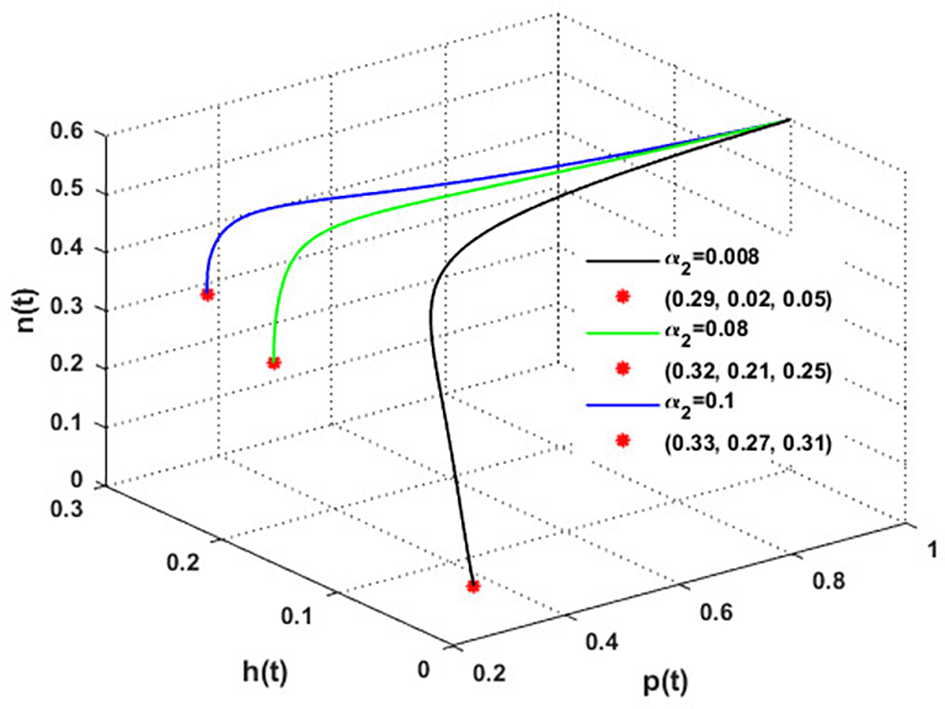
The mutualistic rates between the honeybee population and the blossoming plants, and , are examined in Figures 11 and 12. The density of blossoming plants, the honeybee population, and honey production are all improved as a result of the increase in mutualistic rates.
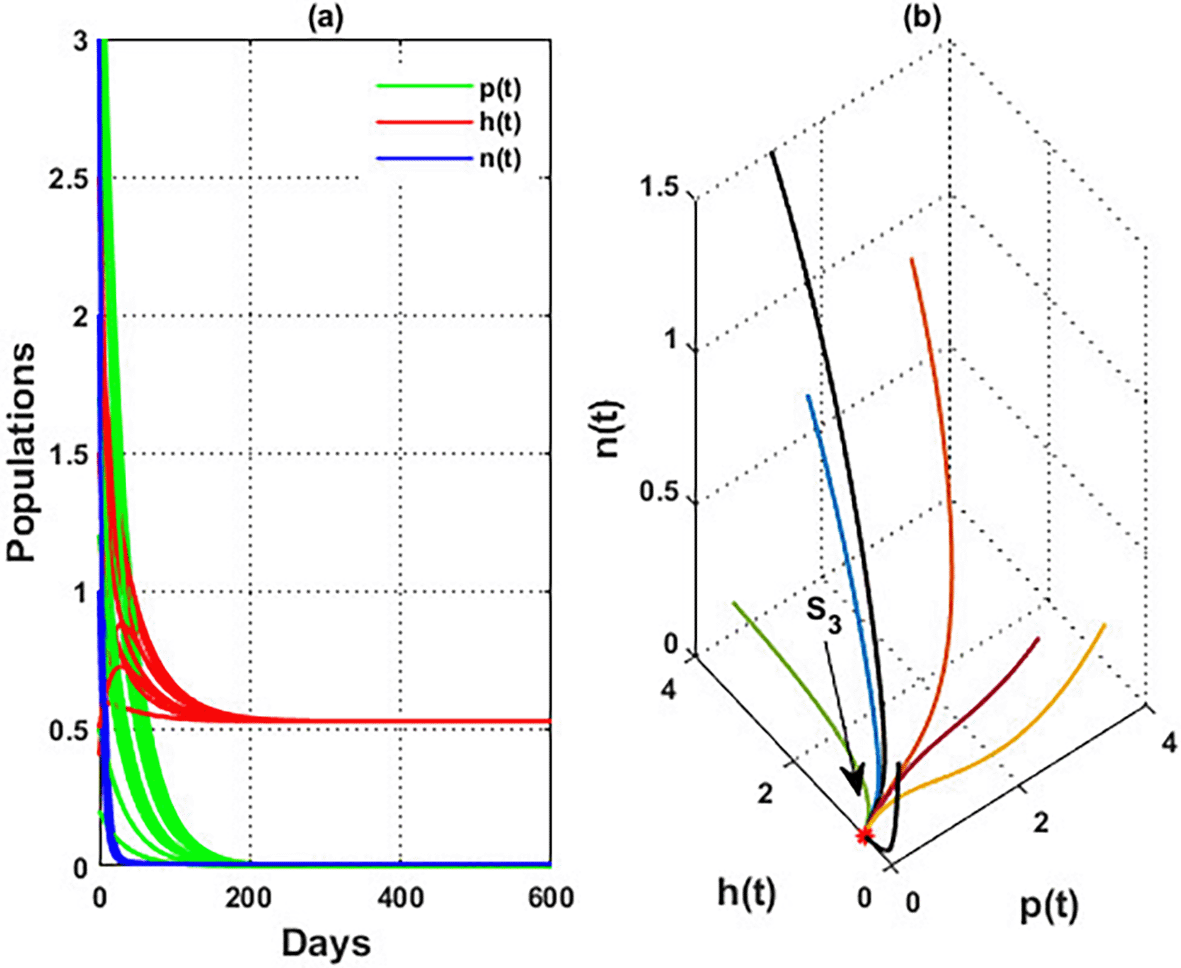
A rise in the depletion rate of the population of blossoming plants population (when leads to losses of the honeybee population and honey in system, and the solution stabilized at the honeybee point from various initial conditions. This result confirms the global stability theorem of which was stated in Theorem 10. As a result, the continued presence of a blossoming plant population significantly impacts the persistence of honey production since the flowering plants are the primary source of sustenance for pollinators. See Figure 13.
To establish the effect of on the dynamics of , Figure 14 has been drawn with three values of the rate at which honeybees consume honey to survive, i.e., . The figure shows the solutions settling down to the coexistence point . The increase in substantially results in a decline in honey output.
In order to investigate the sensitivity of the coexistence point of system, we implement partial rank correlation coefficients (PRCC). The parameters and serve as input parameters, whereas and the output variables. We subsequently generate Figure 15 by utilizing the parameter set in Eq. (36). Blossoming plants and the honeybee population demonstrate heightened sensitivity to honeybees’ carrying capacity and the corresponding value of honeybee nutrients from blooming plants . While the honey production determines heightened sensitivity to . On the other hand, the wind flow, i.e., significantly influences , and . The wind flow significantly reduces the blooming plants, honeybee population, and honey production. It can be inferred that the wind flow is a critical parameter that influences the coexistence of and , see Figure 15.
Wind flow profoundly affects blossoming plants, honeybee populations, and honey production dynamics, and impacts ecosystem stability. An ODE mathematical model has been studied to understand these dynamics. The solution of system has been established to possess the fundamental attributes, such as positivity and persistence, boundedness, local and global stability, and bifurcation. Numerical results indicated that the honeybee species may be extinct due to increased wind velocities within specific parameter ranges. Furthermore, the coexistence equilibrium becomes unstable as a result of a Hopf bifurcation when a low wind flow induces periodic oscillations. Further, the simulations indicated that the threshold values for the transcritical bifurcation have been precisely determined at a decreased honeybee and blooming plant growth rate. However, the system may reach a point where both the blooming plant and the honeybee populations are no longer viable due to an elevated mortality rate of flowering plants. On the other hand, the increase in mutualistic rates between the honeybee population and the blooming plants has a regenerative effect, supporting the sustainability of the honeybee–honey production system.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)