Keywords
plasticity, learning, neural model, vertical integration, internal parameters, neuroplasticity, memory, synaptic plasticity
plasticity, learning, neural model, vertical integration, internal parameters, neuroplasticity, memory, synaptic plasticity
The primary focus of the revisions concerns terminology. The original phrasing was overly soft and allowed for potential misinterpretation. Given that the document introduces a new paradigm, precise and consistent terminology is important. In particular, we have strengthened the distinction between synapse-centric and neuron-centric frameworks, which now serve as the central concepts of the paper. Terminology is applied consistently throughout the text, resulting in substantial revisions across many sections. Despite these changes in expression, the underlying intent and conceptual contribution of the document remain unchanged. We have also clarified the scope of the work by emphasizing that this is a position paper. No specific implementations are proposed, as multiple pathways may exist for translating the conceptual framework into concrete neuron-centric models. Finally, the quality of the figures have been improved.
See the author's detailed response to the review by Srikanth Ramaswamy
See the author's detailed response to the review by Ivan Raikov
Experimental research on neurons, and as a consequence theoretical analysis, is often divided into electrophysiology and molecular biology. The first deals with membrane potentials, spikes and activations in networks of neurons linked via synapses, the second deals with intracellular signaling, genetic and epigenetic expression, regulation of receptors, ion channels, transporters etc. In both cases, signal-induced processes of plasticity are investigated. But data from both types of experiments are difficult to integrate. In the first case, we are looking mainly at induction of synaptic plasticity, with its variants such as Hebbian plasticity, STDP, and other forms of synaptic weight changes based on transmission between neurons. In the second case the data are much more varied, but changes in protein abundance, membrane protein expression and gene regulation are observed under a variety of stimuli.
The master framework for neural plasticity that focuses on associative synaptic plasticity, is usually expressed in the paradigm of long-term potentiation or depression (Malenka and Bear 2004; Citri and Malenka 2008; Kullmann and Lamsa 2007; Remy and Spruston 2007; Lei et al. 2003; Zhong et al. 2006). Many interesting and relevant criticisms and alternative suggestions have been raised (Arshavsky 2021, 2022; Langille and Gallistel 2020; Trettenbrein 2016; Abraham et al. 2019; Gallistel and Matzel 2013; Han et al. 2022). Also, complex dynamic models of intracellular signaling (Baudot et al. 2008; Haney et al. 2010; Crampin et al. 2004) and genetic read-out (Ciliberti et al. 2007; Lee et al. 2017; Baudry et al. 2015) exist, as well as elaborate simulation models (Scheler 2013). Attempts have been made to integrate kinetic modeling of protein signaling into neuron simulators (Bhalla 2002; Vayttaden and Bhalla 2004). These simulators were based on compartment-based dendritic simulations, and did not challenge the dominant paradigm of synapse-centric plasticity. They did not gain much traction. From our perspective, such simulation models failed to capture relevant abstractions, such as the storage of information in internal parameters, the strong selectivity of the internal systems for memory of events, or the time-course of storage implied by the tiered internal system.
To further illustrate this, Figure 1 contrasts (A) a conventional AMPA/NMDA based synaptic plasticity model with (B) a fuller view of external membrane and cell-internal events in response to signals. We observe that a theory of use-dependent synaptic plasticity requires direct transmission from synaptic AMPA/NMDA receptor activation to the nucleus and back. The basis for this direct transmission (“synaptic tagging”, s. below) has never been fully described or experimentally established. The neuron’s neuromodulatory (NM) input and the plasticity of its dendritic ion channels are not integrated into the model - often they are seen as static or serving only calibration purposes (Desai 2003). In contrast, in a neuron-centric view, the neuron undergoes plasticity events - mostly mediated by synaptic input events - which affect receptor binding and (de-)sensitization, protein expression, and genetic read-out. This implies plasticity by different time courses, integration of synaptic events by the respective neuronal cell, and the relevance of internally stored parameters. Because of the complexity of the molecular events to establish plasticity, a strong selection of signals, plus accumulation over time, or similar compounding mechanism is to be expected. In a synapse-centric model, the “two-cell”, or even “three-cell” synapse is the only or the main locus of observation. In a neuron-centric model, the interaction of pre- and post-synaptic events is considered a separate and important step.
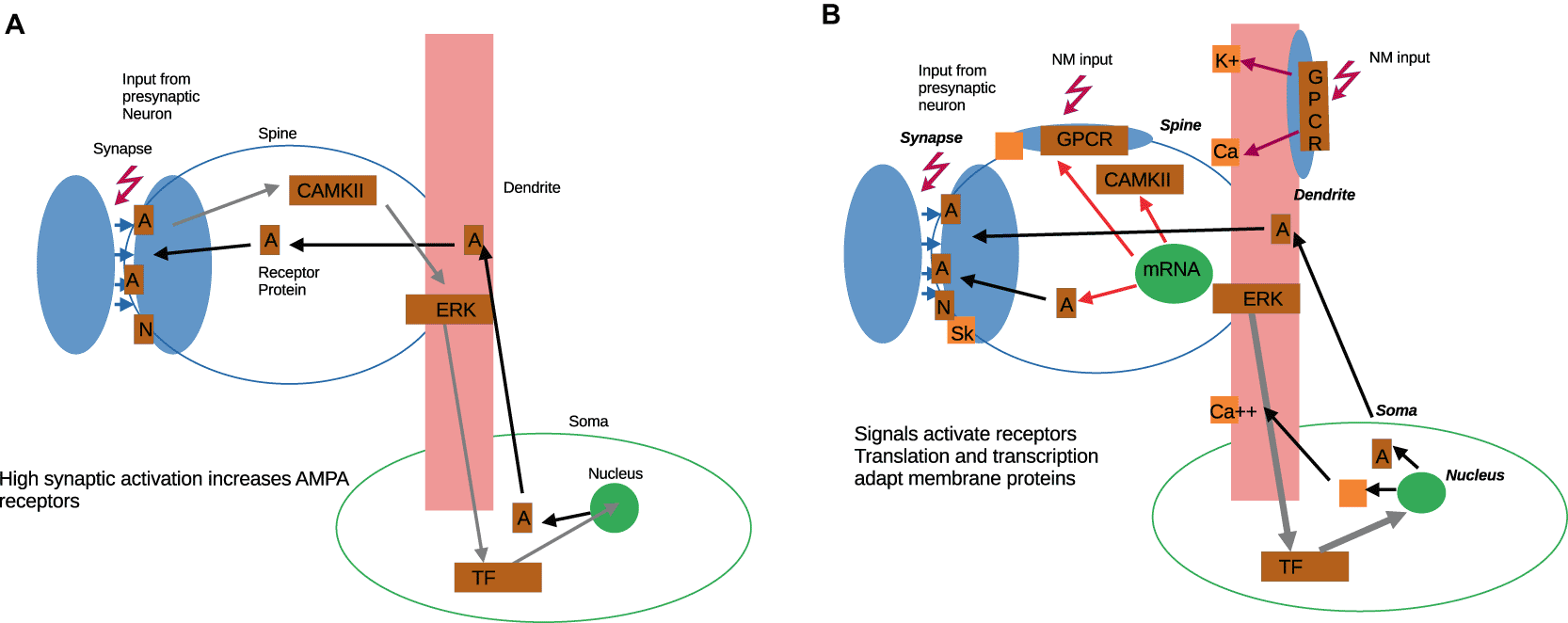
The neuronal cell performs a number of cellular tasks and maintenance mechanisms. Our goal must be to isolate information processing from other complex cellular computations. Therefore, the systems of sub-membrane and cytoplasmic intracellular signaling (Bhalla and Iyengar 1999; Jarvis 2021) may be replaced in a simplified way by amorphous internal parameters, to be elaborated at a later time.
In the experimental realm, membrane receptors respond to signals from the outside, but their localization and sensitivity is guided and controlled by submembrane processes (Scheler 2013, 2004a). This is true for ligand-gated ion channels like AMPA/NMDA and GABA-A, as well as NM-gated G-protein coupled receptors (GPCRs). We suggest to regard submembrane or cytoplasmic proteins and small molecules (‘second messengers’) together as internal parameters.
Membrane parameters – receptors and ion channels – influence neural transmission properties, in particular the activation function, which are visible to other neurons. Internal parameters only have indirect effects on neural transmission and remain hidden from other neurons. External and internal parameters together form the ‘processing layer’ of the neural plasticity system. In contrast to that, the nucleus contains ‘core’ parameters, both epigenetic (histones) and genetic (DNA). This provides another layer of depth for processing information and long-term memory. Core parameters receive and send information to the internal parameters via the nuclear membrane. In this paper, we will not analyze core parameters and their function in detail, we will focus on the actual processing layer instead. This is a position paper which names principles and ideas that will have to be further developed.
External parameters roughly correspond to membrane proteins, such as receptors and ion channels. They undergo plasticity on at least three time-scales: fast (milliseconds, (de)sensitization), intermediate (seconds-minutes, endocytosis, insertion), and slow (hours, new proteins and morphological features like spines). Only the fast responses, which consist of protein conformational changes, do not need the participation of the internal processing system. The endocytosis/insertion of membrane proteins requires a system of internal interactions. The long-term ubiquitination of proteins and the generation of new proteins uses core parameters.
The responses to signals by neuronal cells in the immediate domain has been investigated as short-term (ST) synaptic plasticity. ST synaptic plasticity is known to be highly differentiated, uses all 4 logically possible types, is specific to neuron type and sometimes even synapse, and also highly variable, sometimes changing response type even while processing (Zucker and Regehr 2002; Larsen and Sjöström 2015; McFarlan et al. 2022). Presumably, there are underlying mechanisms not apparent at the surface, which guide responsivity and explain the variability, such as abundance and availability of internal signaling proteins. The set of internal parameters is the first line in orchestrating the plasticity of external parameters. Both dendritic spines and axonal boutons contain subsets of cellular proteins and RNA for local production and regulation, at least about hundred protein species, which regulate membrane protein abundance in the absence of signaling to the core (Donlin-Asp et al. 2021; Kasai et al. 2021). This allows limited compartmental (not synaptic, but integrated), plasticity at the neurons which carry spines, which are usually projection neurons with the strongest adult plasticity.
Two examples of internal parameter activity are offered here:
1. calcium buffers. Under high signaling activity, calcium influx into the cell is high, which activates a cellular braking mechanism via Sk/Bk-channels lowering the firing rate. However, with many available calcium buffers in the cell, the cell can absorb calcium influx for considerable time, store it with the available proteins and prolong high firing rates. Such effects have been used in neural models (Rodrigues et al. 2023), which relied exclusively on synaptic plasticity rules based on calcium regulation.
2. cAMP production. The cAMP production system has fluctuating abundances of the production enzymes (AC) or the degrading enzymes (PDE). This means that GPCR activation during low cAMP availability will lead to a limited amount of downstream activity. Only very strong signals will have any effect. The concept of fluctuations without lasting plasticity applies to this domain. When PDE is weakly expressed (or AC strongly), cAMP accumulates under GPCR signaling events and leads to a broad range of effects, including activating the ERK/MAPK signaling pathways and the nucleus. The neuron may then undergo transforming events such as reading out new proteins, changes in morphology, and insertion of new receptor proteins at the synapse.
We hypothesize that abundance of internal parameters may code for an internal model of signal response. While the neuron receives a variety of signals at its membrane, they are not processed in a uniform way. Rather, the processor itself has state-dependence. When a main signaling pathway is highly active, signals are transferred to actuators – such as ion channels – and the system responds externally, for instance by altering its firing rate, its responsivity to fast signals, its receptivity to AMPA vs. NMDA etc. If a signaling pathway is down-regulated, the same external signals have little effect. This also affects storage. Highly active responses initiate new protein expression via mRNA translation consistent with information storage, while down-regulated pathways will block storage and prevent memorization. This view is well justified from the available literature (Park et al. 2020), but it is rarely taken into account even when plasticity is explained as a set of molecular events in the brain. A static view of a predictable, fixed chain of events prevails (Lisman et al. 2002; Wu et al. 2006; Hayashi and Huganir 2004). In reality, synaptic and membrane plasticity is not only and not directly dependent on the signals between neurons. Instead, the activity of the internal system is necessary for selecting and responding to signals (Park et al. 2020; Alejandre-García et al. 2022). The response of the neuron cannot be determined exclusively from the signals it receives. Its internal state matters.
One task of the processing layer to induce lasting plasticity is to take distributed, time-structured input signals and produce a spatially centralized, temporally integrated signal to the core, i.e. transcription factors which move to the nucleus and cause DNA read-out of individual genes or genetic programs. This is not a simple task.
The internal system contains localized elements in distinct positions (e.g. at dendritic spines, axonal boutons, at a synapse). It also has a central ‘workspace’ in the form of the cytoplasm with its organelles and protein complexes. The change in protein expression which is caused by RNA translation and especially DNA transcription can be described as a “write-once” memory event. Once it has been executed, the system retains the new structure of protein abundances, essentially indefinitely. This is long-term memory of a type that synaptic weight models lack. While new strong signaling may overwrite the structure, such events may be kept very rare by the internal parameter settings.
Signals are selected at the membrane. They are then shaped and re-structured in time. They need to be sorted or transferred to protein complexes in the cytoplasm and ultimately activate the proteins which enter into the nucleus. In the nucleus, epigenetic adaptation regulates access to DNA, transforming signals by enhancement and suppression (Josselyn and Tonegawa 2020). It is clear then that signals from the periphery have to be processed on several levels, and with several types of control structures, before they can actually be used to control or regulate DNA read-out, or be stored in the epigenetic layer around it. In this way, whole genetic programs can be started, such as morphological growth of spines, axons, or dendrites. Individual genes (e.g. AMPA receptors, GPCRs) may also be transcribed during periods of neural plasticity.
The other task of the internal parameter system is to enact short-term plasticity. Signals, fast glutamatergic/GABA synaptic or slower GPCR activation, arrive at the membrane, where they are being transformed through internal parameters by feedback cycles and similar control structures (cf. Figure 2). These can suppress, but also lengthen, or augment a signal by engaging the intracellular signaling network. A well-known example is concurrent activation of calcium- and cAMP-pathways in some cases as a requirement for synaptic weight increase (Manninen et al. 2010). The result is new external membrane expression resulting from a combination of internal parameters and incoming signals.
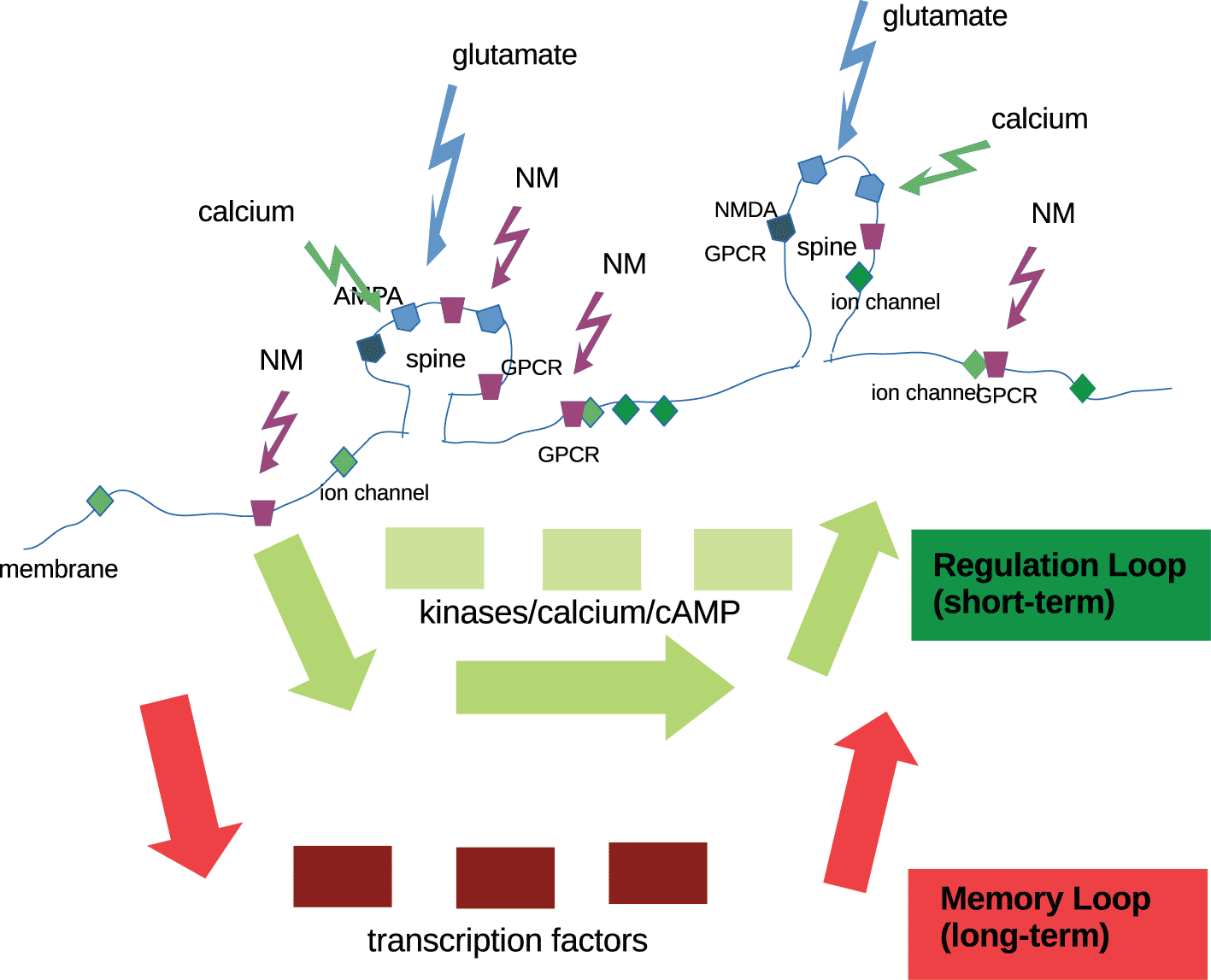
The membrane with its many inserted proteins defines the neuron’s spike response, i.e. its activation function. This controls neural transmission, i.e. the horizontal interactions between neurons. The membrane compartmentalizes into dendritic branches, spines and synapses, for incoming signals. Signals are therefore received by a highly structured system. They can be located by the system, and processed in a spatially distributed way. Synaptic glutamatergic signals, which are brief (milliseconds), produce calcium transients mostly from AMPA and NMDA receptors and voltage-gated calcium channels (VGCCs). Calcium buffering proteins and calcium release from internal stores (Humeau and Lüthi 2007; Gulledge et al. 2005) contribute to the signal. Calcium is a major integrated signal for induction of plasticity where different shapes of the signal result in different outcomes (Ramaswamy and Markram 2015). The selection and filtering of membrane signals is performed by the internal system with its parameters (e.g. availability of calcium-buffering proteins, release from internal stores) which integrates information from internal and external sources.
We assume that the internal system itself cannot regulate its own plasticity (Deng et al. 2011; Shinar and Feinberg 2010; von Bertalanffy 1969). Plasticity occurs via regulation of the abundances of protein species, via mRNA translation or DNA read-out. In this way, the properties of the intracellular system can be re-set and re-calculated. This observation strengthens the view of the internal parameter system as a processing layer, programmable by signals from the outside and updated from inside via parameter re-sets such as local mRNA translation and the core DNA transcription system (cf. Figure 2).
To summarize, the membrane layer is adaptive on several time scales, and in the long term by genetic read-out. Under specified conditions, protein abundances in the processing layer or at the membrane change, adapting the neuron to a new environment. Long-term potentiation/depression (LTP/LTD) can be seen as a special case of signal-induced long-term membrane plasticity, after strong ongoing or temporally structured stimulation, temporal integration for membrane signals, and extended intracellular responses, such as engaging the kinase/phosphatase system (Hunter 1995). In general, the neuron regulates its responses, keeping itself intact as a functioning cell, and adjusting to its position in a network of other neurons.
In our conceptual model, parameters stand for the expression levels of proteins/molecules, sometimes for groups of them, or variants of the same molecule. A precise matching of parameters to biochemistry is not intended at this point. These parameters may sit externally, in synaptic or non-synaptic positions at the membrane, internally near the membrane, or localized somewhere in the cytosol, or as core parameters in the nucleus. On a short time scale, external parameters mediate signal response and internal parameters adapt the neuron’s external parameters. On a slow scale, the values for these parameters are plastic. Neurons receive and respond to signals which are first sensed by external parameters. These occur mostly at synapses as the entry points for signals. There are also receptors in non-synaptic dendritic positions, where neuromodulatory (NM) receptors may act in a’bulk’ fashion (“volume transmission”, (Özçete et al. 2024)). Another variant is GABAergic input from chandelier cells (Contreras 2004) at the axon initial segment, which act as gatekeepers, and input to NM receptors controlling axonal release. While synaptic signals are driving the spiking activity of the neuron, they are also being processed with the help of internal parameters, i.e. filtered or selected for further computation and storage. Immediate and reversible plasticity of receptors and ligand-gated ion channels (AMPA, NMDA, GABA-A) is the first response. When receiving signals, these parameters adapt on a short-term basis, e.g. by conformational changes in receptor proteins (receptor sensitization and de-sensitization). In the absence of strong signal selection, these adaptations are short-term reversible and the effect may be described as fluctuation rather than adaptation. When calcium influx and increase of small molecules, like cAMP, engage the internal parameter system, in particular the kinase/phosphatase balance, control structures and computations by internal parameters decide about the fate of signals (cf. Figure 2).
An important question is how signals are accumulated over space and time. In many situations, external parameters will return to their original values (‘fluctuation’), but they may also retain new adaptive values, when certain circumstances are met (‘plasticity’). To achieve long-term memory, new protein translation by mRNA read-out is necessary. In Figure 2, this is exemplified by transcription factors, which engage the nucleus, in the slower loop, the’memory’ loop. The faster’regulation’ loop may occur within a dendritic spine, which appears as a specialized substructure, almost an organelle for synaptic adaptation. Dendritic spines (and possibly axonal boutons) act as cellular compartments with a reduced but functioning intracellular signaling apparatus (Nimchinsky et al. 2002).
There is a notable difference in plasticity between spiny and aspiny neurons. Spines are localized at projection neurons in areas of strong adult plasticity, such as pyramidal neurons in cortex, hippocampus and amygdala, medium spiny neurons in striatum and in certain dendritic regions of Purkinje cells in cerebellum – areas of high fine-tuned plasticity. For spiny neurons, spine removal and generation is an efficient modification of existing information. In contrast, synapses of aspiny neurons don’t have a dedicated local system of intracellular signaling. Aspiny plasticity relies either on immediate local or on global, rather than compartmental, intracellular parameters. In general, adult plasticity at aspiny neurons is less articulated and less differentiated (Renner and Rasia-Filho 2023).
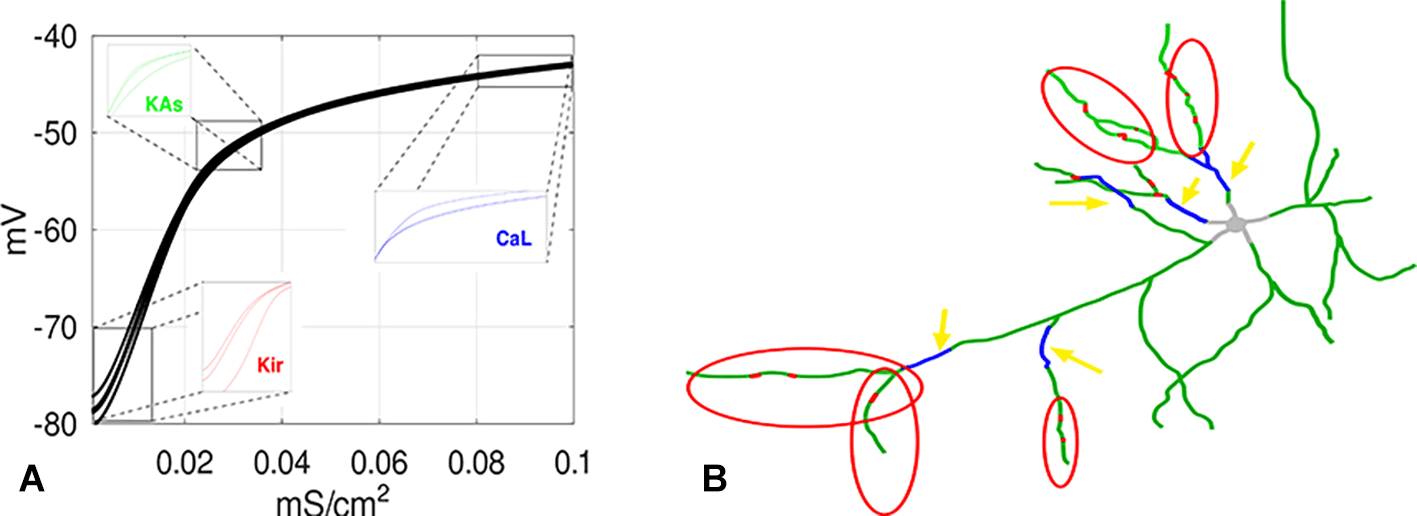
To elucidate when neural transmission causes plasticity is not easy to do in a general manner. Many conflicting data and effects have been experimentally observed (see below). We hypothesize that membrane proteins have a strong tendency towards homeostasis, i.e. return to existing values, after short-term disruption (O’Leary et al. 2014). But in some situations, the accumulation of traces synaptic signaling events is dominant (Abraham et al. 2024). For the synaptic ligand-gated ion channels AMPA and NMDA a number of experimental protocols exist which cause lasting plasticity (up- or down-regulation, (Chen et al. 1999)). It is also known that ion channels integrated in the membrane are variable in their expression levels (Figure 3A,B). The factors influencing this, often summarily referred to as intrinsic plasticity (Debanne 2009; Debanne et al. 2019), are not well-explored (cf. below). Calcium modulation has often been indicated as a major factor in initiating synaptic plasticity (Johenning et al. 2015; Nishiyama et al. 2000; Bloodgood and Sabatini 2007; Sabatini et al. 2002), but there are many other indications for necessary signal types and co-factors (Dolmetsch 2003; Mellström et al. 2008; Fujii and Bito 2022). G proteins are internal proteins linked to neuromodulatory receptors (GPCRs (Liu et al. 2024; Rockman and Lefkowitz 2024)), which have a number of functions. They affect – usually restrict – transmitter release at axonal boutons when placed in presynaptic position (Scheler and Schumann 2003), and transiently regulate the ion channels in the membrane, reducing or enhancing their efficacy. Ion channels determine the excitability of the neuron. This means neuronal activation functions are conditional on which (central) NM signal modulates them. The activation of different NMs highlights the set of neurons that are modulated by them (Scheler and Schumann 2003). NM-ion channel interactions can reversibly remodel dendrites at intermediate time frames (Figure 3B, (Furusawa and Emoto 2021; Kanamori et al. 2015)).
It seems questionable that abstractions of synaptic and neural plasticity could take on the general shape of a ‘learning rule’.
Many factors influencing outcomes have been experimentally observed: neuromodulation, subsequent or concurrent; spacing of synaptic signals; reinforcement by recurrence in a network (‘re-entrant’); capacity of internal neural plasticity pathways like CaMKII, PKA, or MAPK; amount or strength of neuromodulation; the NMDA Mg++ switch, a non-linear effect; concurrent GABA signaling; dendritic processing such as synaptic positioning on dendritic branches; backpropagation of calcium signals to distant synapses; dependence on ion channels gating backpropagation, synaptic spacing (AHP); additional hyperpolarization (GIRK) or depolarization (Musc, K + A- channels), cross-talk among internal pathways; globality of the G protein pathway;and possibly others). Intracellular calcium buffers and mechanisms for release from intracellular stores influence calcium transients and are an example for the integration of outside signals and internal parameters, with strong, high calcium transients vs. lower ongoing signals. Sk-channels act as plasticity blockers at a synapse. Neuromodulation (NM) via G-protein coupled receptors (GPCRs) (Shenoy and Lefkowitz 2011) also influences signal selection using presynaptic NM receptors for gating.
However, it may be possible to postulate principles for a number of learning rules which interact to produce an outcome in specific well-described situations. Synaptic signals to a neuron have the potential to activate transcription factors as immediate early genes (e.g. Arc, CREB) within minutes (5–20 minutes for Arc in hippocampal neurons (Guzowski et al. 1999, 2001)). These transfer to the nucleus (Rienecker et al. 2022; Minatohara et al. 2016), and start new transcription. But they have to pass the intracellular signaling system. This has a large number of control structures such as thresholds, feedback loops, feed-forward loops, antagonistic signaling (Scheler 2014c). For instance, negative feedback loops serve to broaden a signal and extend its duration. They may also dampen a signal below threshold (Scheler 2012). Signals may be modified, such that later signals in a sequence are being suppressed. A sequence of signals may be enhanced to reach a threshold (Maus et al. 2020). These control structures are activated by signals but they depend on internal parameters. Control structures may use existing neighboring structures, such as the neighboring relations on the dendrite, to regulate signals (Gidon et al. 2020; Winnubst and Lohmann 2012; Makino and Malinow 2011), either potentiating them, or canceling them out against each other. In this sense, internal control structures serve to adapt a signal and influence whether it is passed on to the next layer.
The synaptic model of plasticity assumes that an activated synapse signals to the core, and the core signals back to the relevant synapse. This “synaptic tag” hypothesis has been much discussed (Rogerson et al. 2014; Redondo and Morris 2011; Li et al. 2014). There are many problems with this simple assumption. One of them is how to disentangle several synapses in different positions, which all signal through similar pathways. Another one is graded plasticity, when only single bits of information can be sent in this way. A third one is the temporal depth of synaptic adaptation, and the question of ongoing processing of synaptic weight information over periods of minutes and hours while long-term plasticity occurs. Finally, this system places plasticity on the postsynaptic side, while forms of presynaptic (axonal) plasticity (modification of release probability) also exist. As explained above, the first question is when a synapses undergoes fluctuation, and when it is marked for update and this first question is already influenced by the existing state of the internal signaling system. This leads to an integration of strength or shape of transmission events with internal state parameters.
Under conditions to be specified, local marker signals are translated into signals to the intracellular network. The number of marked synapses is low at any time (strong selection of transmission events which leave a trace), and update events are centrally regulated in an integrated way and are essentially sparse. There needs to be a strong selection of signals before updating is initiated, if only because adaptation is metabolically costly.
Criteria for signal selection could be strength (high amplitude, co-occurring at various sites), or repetition (at the same sites), or long duration (even at a lower level of signaling). Even fairly weak signals can be retained, if signals combine within a specified time frame, and if the neuron is highly responsive. Strong signals may fail to register, if the signal is isolated and the neuron has low responsivity.
Several models are possible:
1. We may assume that external parameters fluctuate in response to signals, they follow a random walk and need reinforcement by internal parameters to store a value. Such a model could emulate modern feature selection methods (Guyon and Elisseeff 2003) very effectively.
2. External signals are selected by their magnitude and pattern for plasticity. For instance, an individual spikes may not cause plasticity, but a number of neuronal spikes which occur consecutively may cause plasticity, especially if the pattern of such spiking recurs with breaks over time (Wu and Maass 2025). This would be suitable to store events which occur within short time frames and are being replayed internally. Synaptic events which do not cause the neuron to spike would be ignored.
3. External signals are stored if the internal system is highly responsive with main plasticity pathways (e.g. kinases) all ready to be turned on. After a neuronal or synaptic plasticity event, the system turns unresponsive (e.g. high phosphatase activation) and for periods of time the probability of further storage is low and may not even return to full naive responsivity. Experimentally, the effect of responsivity has been studied along these lines by (Zhou et al. 2009; Park et al. 2020; Dehorter et al. 2015).
The common theme is that signals are ignored or passed on depending on the responsivity of the neuron, as well as the strength of the signal itself. In spiny projection neurons, the spine may act independently of the main cell body within the regulation loop.
Within the paradigm of a neuron-centric view of plasticity, each neuron represents a processor which will process input information according to its internal functions and current state. It is important to realize that neural computations are performed by heterogeneous neurons with different genotypes and cellular identity.
Recent work on neuronal genotypes has often confirmed that neuronal connectivity is highly correlated to its genetic make-up (Planert et al. 2025). This emphasizes a significant restriction on adult plasticity, since a large part of connectivity is not set up by data or patterns, instead it is genetically determined. Perceptual input may be necessary to trigger waves of activity to establish the synaptic connectivity in each case.
Evolution has done the ‘main work’ in establishing the neuronal types. For instance, pyramidal neurons, as a type of excitatory projection neuron, have a variant in most mammals, the stellate cell, which appears during development as truncated in its dendrites and axons. There are cortical, hippocampal, amygdalar and other locations for pyramidal neurons. Within cortical neurons, there are dozens of genotypes which can be clearly distinguished (Chang and Luebke 2007; Kim et al. 2015; Mao and Staiger 2024; BRAIN Initiative Cell Census Network (BICCN) 2021; Gouwens et al. 2019). To model neuronal types and their connectivity, the evolutionary processes do not need to be re-created, even though evolutionary modeling could also be very interesting. Instead, we may learn from biology and set up neuronal types according to type and regard them as type-specific programmable units. This greatly reduces space-time complexity for adult learning. Nonetheless, our understanding of the existing genotyped structures for cortical (or hippocampal, striatal, etc.) functions is still developing.
We hypothesize that neurons have generic programs for signal induced plasticity. A neuronal cell has a complex control structure for protein signaling which may undergo adaptation to acquire a specific internal model.
When undergoing plasticity, they may acquire an adapted program (Alejandre-García et al. 2022; Paik et al. 2020; Traunmüller et al. 2025) which then becomes part of their activation properties. E.g. in Bulovaite et al. (2022), the lifetime distribution of PSD-95, a postsynaptic protein, was tracked on neurons in cortex and hippocampus. They found short-lifetime PSD-95 at young or innate (‘naive’) neurons, and longer PSD-95 lifetimes at aged (‘mature’) neurons. This points to a loss of flexibility in synaptic learning during maturation and aging, or vice versa, an increased stability for learned information.
Default or generic plasticity configurations possibly contain random elements. Exposure to patterned signals lets them acquire a specific generative model which reflects the neuron’s experience and expectation. This is a’mature’ neuron, or’adapted’ neuron, which we can probe for its behavioral response pattern (Quiroga 2012; Sharma et al. 2024; Waidmann et al. 2022). A mature neuron may still re-learn, and possibly even re-set to the naive state under exceptional circumstances. In the naive neuron there is no individualized control structure to filter the signal. Patterned signals have the effect of imprinting a neuron with a specific control structure which keeps the state of the neuron and its synapses in a certain configuration. Many behavioral learning events may fulfill the criteria of such an imprinting effect on naive neurons - there is also evidence that the brain, or at least the cortex, keeps many naive neurons available even in adults for new learning of situations and events (Ovsepian 2019). Imprinting or instantiation of a neuron involves setting-up a copy of the external parameter set as the basis of a generative model or as an existing model for later processing (Scheler and Schumann 2019). Once a neuron has acquired a specific external structure (synaptic strengths and distribution of NM receptors), new signals are then matched to the existing parameter set. Close matches may continue to refine the external parameter set, while non-matches will route the signal to other places. A neuron may contain both naive and mature sites (especially at spines), it could be ‘chimeric’ with respect to maturity. A naive site will store or read in a value, a mature site already contains a value, compares a new value with the existing one, and it can be protected against overwrite (Sk-channels). It is however important to realize that for many cognitive operations the unit of processing will be neither the individual neuron nor its spines, but groups of neurons which together form a concept, or unit of computation at a higher level. The individual properties of the neurons help to explain how these larger units can develop and function.
We showed that neuronal cells have internal signaling mechanisms which are activated by external signals. The main known pathways for neural plasticity are the calcium-related, the cAMP-related, and the MAPK/ERK pathways. Other pathways, such as BDNF-Trk interact with them or signal through other means (such as NF-KB (Schirò et al. 2022)). The interesting observation is that these pathways fulfill the idea of a programmable system. By changing the abundance of protein species there is long-term plasticity in the system, and by activating/de-activating proteins (e.g. by kinases/phosphatases) the system is plastic on the seconds-to-minutes time scales. The system consists of interactions with characteristic time-scales and strengths, expressed by kinetic parameters. Connections may be strengthened or disappear beyond a lower threshold based on the available numbers of protein molecules.
An example for this is’transactivation’, or an overflow switch, as a control motif in computation cf. Figure 4A. In Scheler (2006), we showed how a strong signal at a receptor results in activation of a target protein, which is not involved in signaling by this receptor at baseline. This is facilitated by high protein expression of the target pathway. In this way a pathway is activated conditionally, i.e. only when internal conditions are met, and when the signal at the receptor is sufficiently strong. It is not fully understood how both conditions interact. A suitable model could be fully linear, by summation of strength of signal and protein expression, but it could contain a non-linear element, such as a threshold, e.g. for the protein expression. This is what we assumed in Scheler (2006), when we talk of a switch that becomes activated.
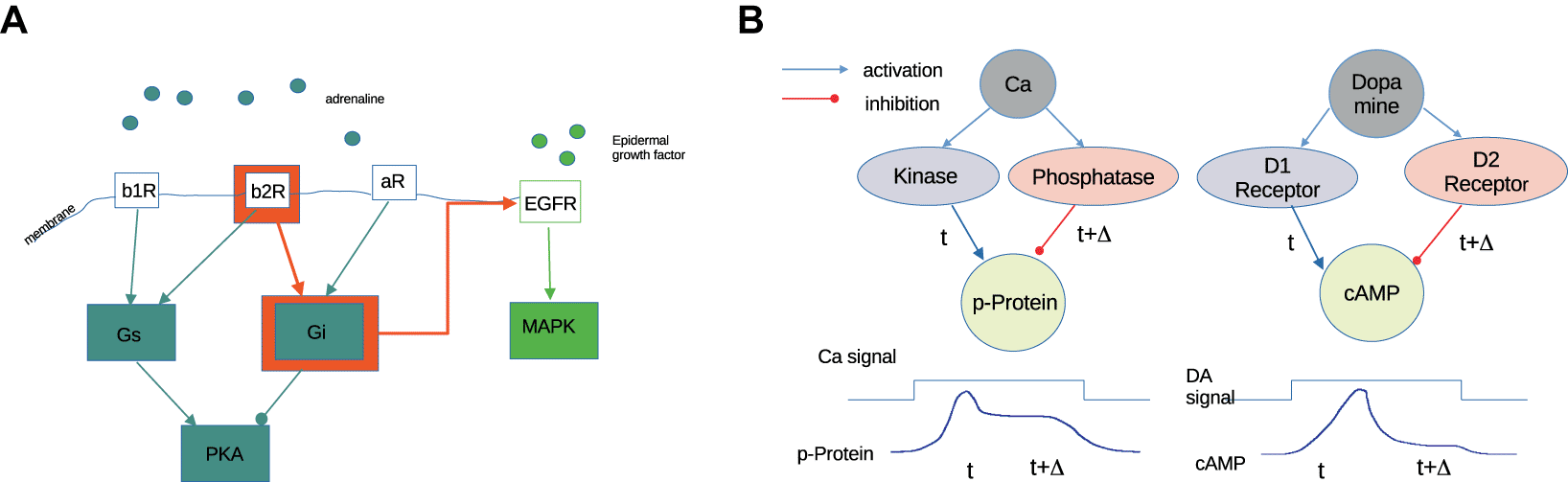
Another common, ubiquitous motif in biology is antagonistic signaling. Within protein interaction, there are several examples for this. For receptors such as D1/D2-type dopaminergic signaling, or beta/alpha-adrenergic signaling, this occurs because the signal input is linked to its output by two antagonistic pathways (positive and negative) (Scheler 2014c). The antagonistic motif allows re-scaling of inputs which vary over a large dynamic range, compressing multiple orders of magnitude into a single-scale response. Also, antagonistic signaling can generate peak transients when one pathway is slightly faster than the other (Figure 4B). We can modify the transient peak, or the amplitude of the outcome of both pathways by protein abundance, i.e. long-term plasticity.
Internal signaling can also reset presynaptic vesicle pools (e.g. by cAMP-dependence), and it controls spine maturation and spine decay. It performs spatial integration on the spine (e.g. when AMPA receptors reservoirs on the spine shaft move to the spine synapse (Araki et al. 2010)). There are local membrane-near calcium buffers (e.g. CaMKII), which can store synaptic signals for minutes by autophosphorylation of the kinase in the synaptic positions where they are deployed. In the cytoplasm, there are timed delays, queues, and feedback cycles, which suppress a signal or enhance it and the position and significance of which can be learned. There is spatial integration over several dendritic compartments (Shenoy and Lefkowitz 2011) which can cause activation or de-activation of a dendritic branch.
A significant specific property of internal signaling is the’shared workspace’ of the cytoplasm which provides an access system for dendritic and axo-somatic compartments. Inputs from postsynaptic signaling are routed into the shared workspace, from where they can access the nucleus and induce new DNA read-out. At spinal synapses, internal signaling may remain localized (Steward and Schuman 2003) or spread within minutes along the dendrite. The’workspace’ is therefore not evenly distributed, but has distinct probabilities for molecules to interact. Signals may be fed back to nearby locations on the membrane, such as ion channels on specific dendritic branches. However, when signals are received at multiple locations, there is then signaling via the shared workspace, and a system-wide response. Again, it is not entirely clear at this point, how these signals are combined, but multiple control structures have been identified.
Molecular abundances are important for establishing (by high expression) or removing (by low expression) connections between pathways via their kinetic interactions. They also dictate the temporal execution of the biochemical reactions underlying intracellular signaling (Scheler 2014b). High expression means that reactions are fast and homeostatic endpoints are reached in short time. When protein expression is low, internal reactions are slowed down and many signals fail to be transmitted. In terms of a neuronal response, the system is not processing and storing internal information, while still transmitting information by activation functions within the neuronal network.
An example for the power of the ‘shared workspace’ is shown in Figure 5A. The system is versatile enough with its many control structures to perform interesting functions. Using different types of buffers available, we can even achieve a re-sorting of signals in time. For instance, CaMKII which buffers calcium by autocatalytic activity, releases calcium depending on the activation status of each buffer molecule. This allows to create a reverse sequence of calcium transients, which may be supported by internal calcium release patterns. Individual synapse/spine input order may be reversed according to internal properties, providing a basis for queued access to nuclear transcription. A sequence of synaptic inputs, which are sorted and combined from their spatial distribution into larger components (e.g. like a bit code which is translated into ‘words’), could provide a ‘canonical’ shape of signal that can be easier read.
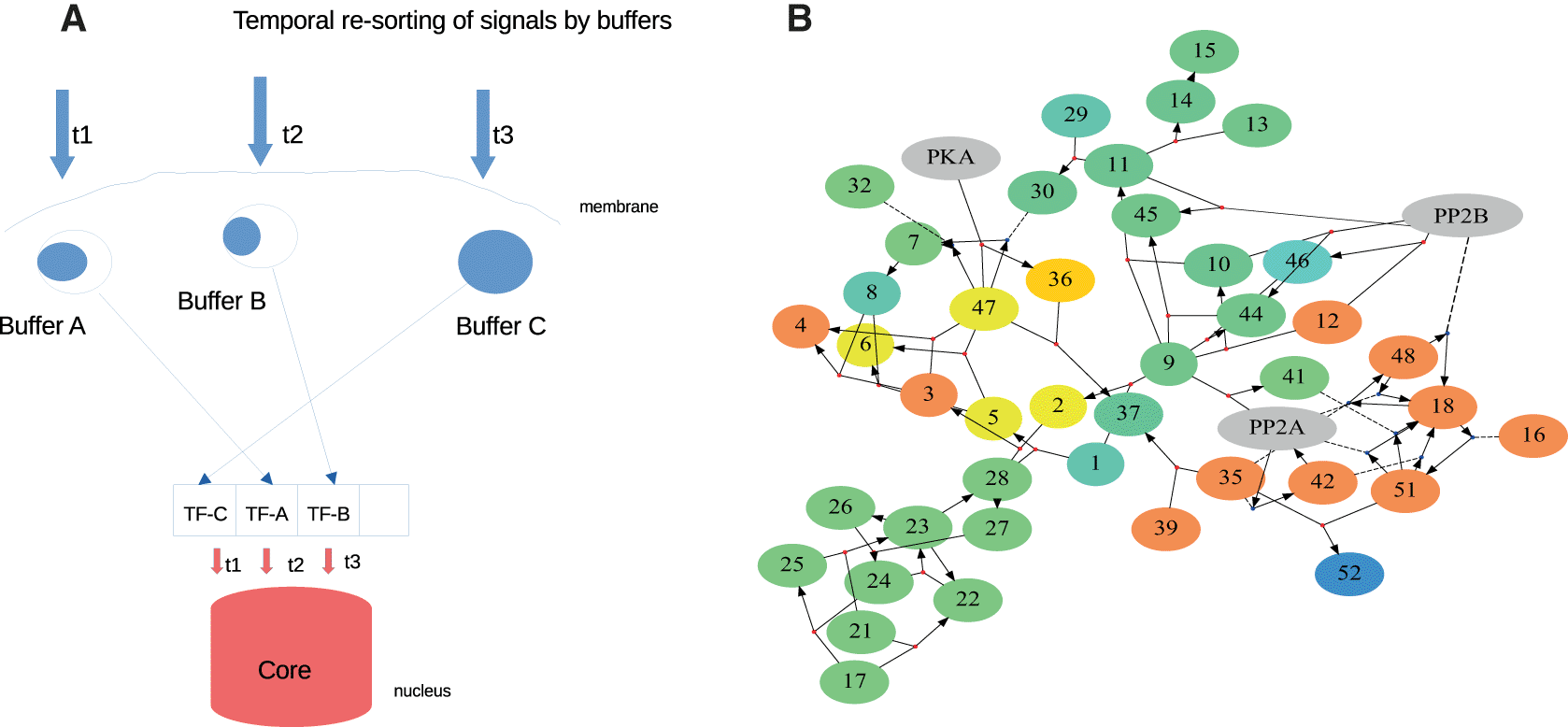
A: Temporal re-sorting shows how buffers can re-order the timing of signals for activating transcription factors (TFs). Buffer C reacts first because it is saturated. B: Changing the concentration of proteins (shown by numbers) in a biochemical reaction network (by 100%, details in Scheler (2014b)). Speed changes are shown in yellow/red (faster) and blue (slower) hues (green unchanged). Increasing both kinase (PKA) and phosphatase (PP2A/PP2B) leads to a strong speedup of a large number of reactions.
To summarize, outside signals are received by external parameters and activate a program in the internal system which determines the neural plasticity response (Scheler 2005). The internal computational system that filters and temporally re-sorts signals, that combines signals, and stores them, is a programmable and re-programmable system. The system contains parameters which program its functionality. In Scheler (2013), we employed a transfer function approximation for internal signaling. Transfer functions allow to generate (context-dependent) look-up tables for endpoints of internal parameters in response to outside signals. On this basis, internal parameters can be acquired and adapted by setting outcomes. To re-program the system requires new transcription or mRNA translation. It can be reset by the core, a genetic read-out system. To investigate the power of such systems, finite state machines, constraint satisfaction, optimization, as well as machine learning techniques could be suitable.
The formation of neuronal groups (ensembles) is prominent in various areas of the brain, e.g. in mammalian cortex, where neurons are arranged in layers and microcircuits. These ensembles are almost always comprised of projections neurons interspersed with inhibitory interneurons, no matter whether the projection neurons are excitatory or inhibitory. In many neural areas, e.g. hippocampus, cortex, we see widespread electrophysiological activation of 30–50% of neurons within a behaviorally relevant brain area in response to a significant event, like a fear experience. We also often see activation of immediate early genes (IEGs, e.g. Arc) at a similar density, however, transient activation of IEGs is not a reliable indicator of lasting neural plasticity (Robbins et al. 2008; Bruins Slot et al. 2009). Everyday memorization of facts and data may rely on a much lower proportion of neurons being activated, and use highly restricted localized plasticity (Scheler et al. 2025).
Especially in cortex, input patterns face the task of finding the best match among the existing structure of cortical microcircuits. There are experimental data (Sherman and Usrey 2024a, 2024b; Usrey and Sherman 2019) that thalamic input activates many cortical ensembles in parallel and selects best matches by activations which are recurrently potentiated, and probably connected via synchronization (Scheler 2018), while weaker matches are suppressed and fail to become activated. A network of activated cortical ensembles gives rise to representational functional states which are measurable by brain imaging.
Sensory or input patterns in general undergo transformation via the signals they produce at individual neurons and their integration into an ensemble. At those sites, a pattern may be matched to an existing stored template and become adapted to existing knowledge. Strong matching signals may generate internal signals and re-program a neuron. For a non-matching signal, internal inaction will stop the outside signal from passing into the processing layer. In this sense, a whole ensemble is selected for memorization of a pattern, not just a single neuron (Josselyn and Tonegawa 2020).
In an earlier paper (Scheler 2018), we argued that neurons may function either in a synchronized transmission mode or in an irregular-asynchronous mode, where they can read out and adapt membrane parameters. Pattern matching and selection would result in high activation of an ensemble and synchronization across ensembles. Such a regime could be described as high’strain’. (‘Strain’ could be defined and measured by both synchronization and amplitude of a neuronal group.) Only under high strain may we assume internalization of information to be initiated. As’strain’ builds up, it gets resolved by adjusting parameters. Parameter adjustment (internalization or externalization) would occur only under specific regimes. High strain may cause a network to internalize information from the external membrane, or vice versa to read out parameters so as to reduce activations. This would allow a return to a well-adjusted regime of low strain over an ensemble and an end to widespread synchronization.
The interesting thing is that we may use ‘high strain’ states to solve the question of when to retain information and when to discard it for a functional group. When strain is high, information is retained and features are delegated to each neuron. Under low strain or relaxed strain the group interacts and solves internal constraints. The overall function of the group is used as the learning goal, objective function or output target.
Neuromodulation has an interesting role in that it codes for neuronal identity in a combinatorial fashion (Scheler 2004a). Each neuronal type is differentiated by the NM receptors it carries, and neurons may even be unique on an individual level in terms of the localization of the NM receptors. NM receptors can thus serve for labeling, both for neurons and connections (nodes and edges of a graph structure). The label states what NM receptor, if any, is present (graph’coloring’). Such labeling is extremely useful because it allows for constrained inferences (Scheler and Schumann 2003). For instance, all neurons labeled for dopamine D2 receptors would hyperpolarize (via GIRK channels) at a dopamine signal, while D1 neurons would abolish afterhyperpolarization (AHP) and fire at higher rates.
In the short term, control structures using internal parameters in the membrane-near intracellular layer influence external membrane properties directly and immediately. Spatial resolution is kept. This is fast adaptation of neural membrane properties. Additionally, membrane-near internal control structures select strong signals, which are internalized and maintained (e.g. with the help of calcium buffers, widespread protein phosphorylation, increase of cAMP, inositol etc.) When the signal reaches cytoplasmic protein complexes and transfers into the nucleus, spatial resolution is lost and global neural remodeling occurs. In general, lasting plasticity requires this step.
Overall, we anticipate a network of processors with their own individual memory, connected by graded signals, participating in low-dimensional representational configurations, to offer a highly flexible and sophisticated framework suitable for complex computations (Gallistel and Matzel 2013; Langille and Gallistel 2020).
Neurons typically communicate by action potentials (spikes), transmitter release and variable spike rates over time. These range from sparse spiking neurons at 0.1–1 Hz, to medium rates at 10–20 Hz, to fast-spiking neurons usually in the interneuron domain (40 Hz+). While neurons vary their baseline rate under excitation or inhibition, the variance is typically low, about 50–100% (Scheler 2017). Neurons also communicate in a slower fashion by release of neuromodulators or postsynaptic slow receptors for glutamate and GABA (mGLU receptors, GABA-B).
Another form of neuronal transmission has been explored as a result of biophysical investigations into membrane excitability (the “soliton” theory, (Heimburg 2022)). This would implement strong and ultrafast direct transmission of excitation across membranes of neighboring neurons. The transmission is strong enough to cross the synaptic cleft. This would lead to chains of neurons which can be activated in this fundamentally different way. Building chains of neurons (or paths through a network) is in general a good strategy to ensure fast transmission in spite of recurrence. The soliton type of transmission may be of particular interest, because, in contrast to transmitter release/receptor activation, it may not undergo plasticity. A significant component of neural transmission may be thus devoid of plasticity.
Current theories of synaptic plasticity implement a model of memory where information processing always leaves a trace, and is performed in a uniform way, building knowledge by accumulation of facts or data, or simply patterns. It is also fully reversible, i.e. can be obliterated by new traces, That is not very realistic. From a cognitive and behavioral perspective, memory does not appear as a reversible operation where a blank slate is inscribed and wiped again, but as an operation where permanent changes occur when memories are stored. (An important observation in this respect is “extinction” (Myers and Davis 2007) as opposed to erasure of fear events.) In general, only a fraction of events is stored with chance of retrieval. Furthermore, a developmental trajectory exists from neonate to juvenile to adult to aged individual, with effects on ease of memorization and retrieval. At any age, much information is deemed not worth keeping, it is lost within minutes, hours or overnight. Information that is sufficiently represented by existing knowledge is possibly used for adaptation or strengthening, or it is simply discarded, i.e. processed without leaving a memory trace. In general, much information that is processed on an everyday basis undergoes abstraction and may become part of a schema complex that it adds to in small ways.
We outlined a model with external parameters at the membrane, which are relevant for the transmission of information to other neurons. Internal parameters, hidden from interactions, store and filter information and are capable of adapting external values. The core system programs and re-programs the processing system. The difference between novel signals and existing information drives adaptation. It forces a system, such as the brain, to remain an open system with respect to its environment. Such a system lacks the properties of a formally’closed’ system with fixed properties. It is necessary to control such a system, to prevent it from arbitrary adaptations. Memory management is central for that. It ensures consistency, promotes signal loss where appropriate (Scheler 2001), and prevents’catastrophic forgetting’ (Grossberg 2020; Paik et al. 2020), i.e. adaptation without control.
A very general perspective of what drives neuronal plasticity is the difference between stored information and new, incoming signals. Also, there is a role for internal states in selecting new information for storage. The mechanisms that guide the interaction of internal state parameters and incoming information can be defined along the following lines:
Any neural system must contain elements with high stability
a) which provide the backbone of the knowledge structure and are protected against change
b) and elements which flexibly adjust to a current situation.
Neuronal types, defined by genetics and as the product of evolutionary learning, are essentially stable, and they form part of the background structure. Axonal/dendritic morphology is also a stable component. It is defined during development and requires core-mediated programs for alteration. The stability-flexibility trade-off is probably at the center of the capacity of brains to build structured representations (Grossberg 2020).
Neurons as processors operate with graded stability from outside to inside (Figure 6). This principle seems very intuitive and sound, but it is unusual in computational theory. Memorization involves transforming transient information (signals) into stable information elements. Concentric abstraction of temporal depth provides short-term adaptation, long-term stability and the capacity to store information into increasingly stable formats (Figure 6).
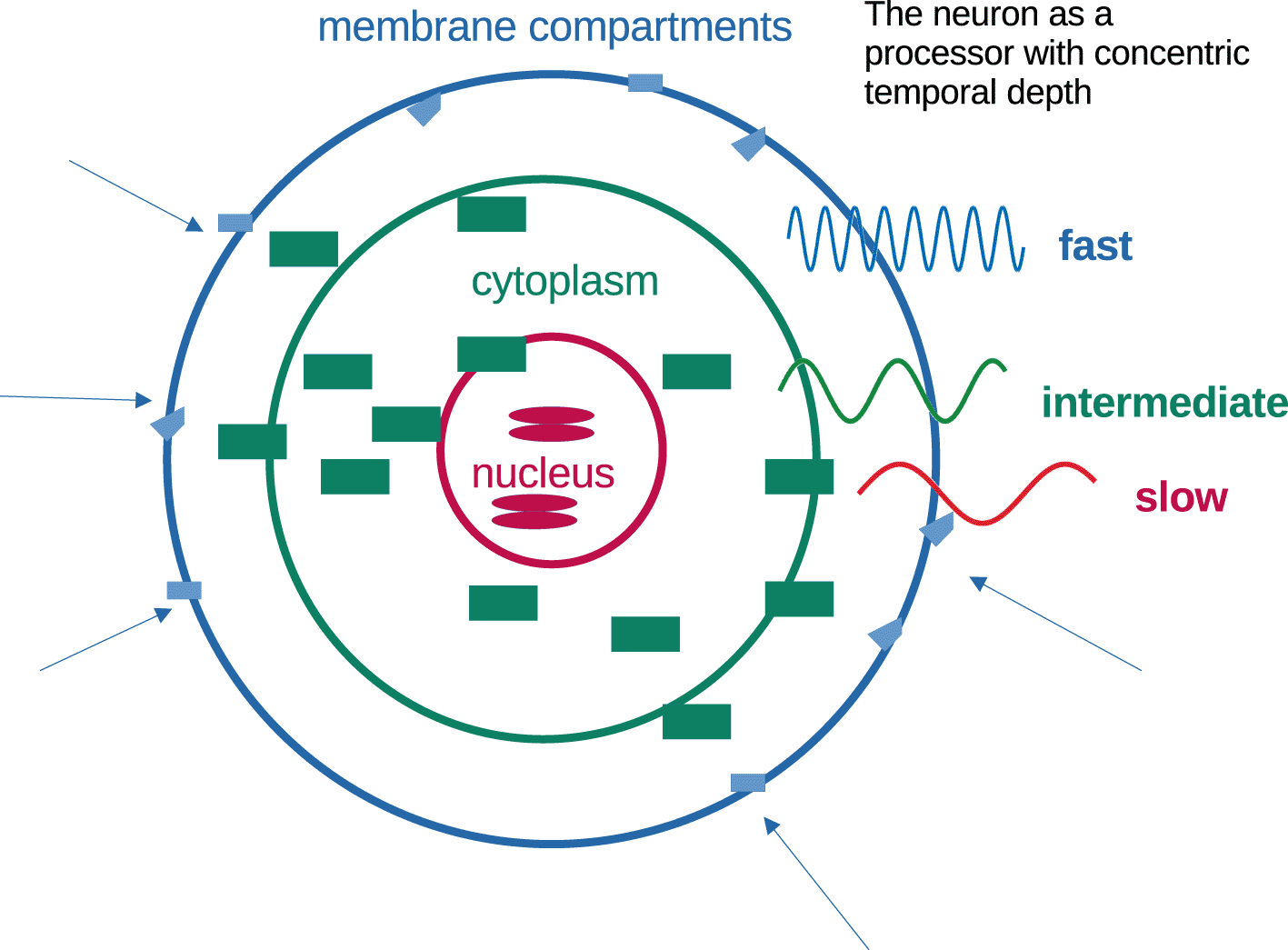
A synapse-centric theory of memory has a number of unrealistic consequences:
• synapses are adapted independently of each other. They are also adapted independently of the neuronal cell state,
• there is no higher unit of adaptation beyond the synapse itself
• adaptation in ion channel expression or NM receptor placement are not part of the theory
• since synapses have high turnover (Mongillo et al. 2017; Fauth et al. 2015), information is periodically lost or overwritten, unless it is transferred to other synapses in a pattern completion process (Fauth et al. 2015). Memory has to be moved just to make up for the biological limitation of the synapse.
Another major problem with the synaptic hypothesis is that adjusting weights in graphs has weak expressive power. Treating neurons as adjustable activation functions improves efficiency of implementation, but does not significantly extend the expressive power of the system (Scheler 2004b; Kang and Palmer-Brown 2006; Soltiz et al. 2013). When such systems store whole knowledge bases, they acquire and store curated data sequentially, and they require enormously large graphs because of a lack of compressibility. They overwrite storage by recency. This leads to problems for more complex hierarchicaln knowledge and action plans (Hosseini-Asl et al. 2020).
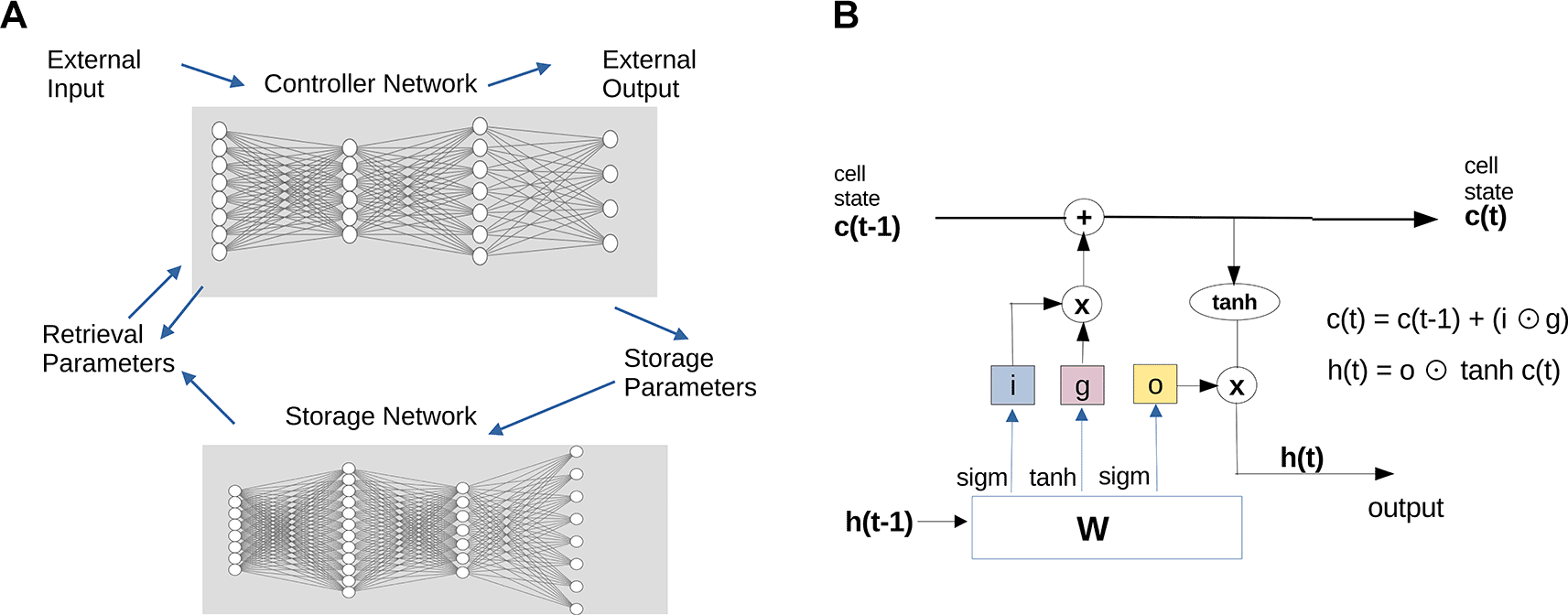
In response to some of the challenges of synaptic adaptation models, numerous improvements and workarounds have been suggested. For instance, separate memory structures were proposed to solve problems of sequentiality and compressibility (Graves et al. 2014; Hochreiter and Schmidhuber 1997), (cf. Figure 7). When memory is conceived as separate from processing, a selection of memory traces becomes possible. Another recent improvement (Graves 2014) involves the insertion of large amounts of residual connections (“attention”) into learned graphs to create giant connected networks. This allows for significantly enhanced pattern completion, even for large and complex patterns (“transformer architectures”). However, the problems of efficient modularization, of hierarchical schemata abstraction and inferencing are not addressed by these models. A system which combines ensemble processing, selection of items and cascaded depth in internal storage might be able to solve some of these problems.
An important advance of the proposed model is that it provides a blueprint for a biologically adequate paradigm with applications to both psychological and medical disease models. There are many extremely detailed, biologically realistic models, both large (Ito et al. 2026; Shichkova et al. 2025; Ali et al. 2026) and small (Scheler 2013, 2014a, 2018), which however lack applicability to even very simple tasks. There remains an unmet need for brain theories and models that demonstrate robust functionality. Such frameworks should incorporate the neuronal cell as a fundamental component, enabling them to adequately capture disruptions of brain systems that arise from disease processes on molecular levels. Many diseases of the brain relate to processes, molecules and interactions that are insufficiently described within the paradigm of synaptic-centric plasticity.
We want to put forward a single example of a pharmacological effect which illustrates the limitations of synaptic plasticity models and expands the view towards neuron-centric models. It was found (Lopez et al. 2022; Shinohara et al. 2021; Kang et al. 2022) that ketamine as a drug drives genetic expression of the KCNQ2 channel, which increases the (K+) current. In pyramidal cells, e.g. in hippocampus, this reduces bursting and increases reset after spike firing, limiting the effects of afterdepolarization (ADP). It also reduces spontaneous glutamate release at the excitatory synapse. The one-time increase of the genetic expression of an ion channel provides effects for many weeks. From this perspective the long-lasting anti-depressant effect of single-dose ketamine (Abdallah et al. 2015) for psychiatric patients can be explained and modeled.
This type of result is easy to incorporate into a neuron-centric model with its integration of neuronal and synaptic properties, and its focus on levels of persistence in the internal part of the cell (cf. “write-once” memory). A synaptic weight adjustment model is too poor to adequately model such effects – if such problems are tackled at all, they are solved by various additions to synaptic weight adjustment as the expression of all forms of memory (Citri and Malenka 2008; Ramirez and Arbuckle 2016; Magee and Grienberger 2020), resulting in “epicycles” (arbitrary amendments) of the main paradigm.
Memorization relies on biochemical events. Neuron-centric theories with internal computation can be considered a substantial way forward, especially compared to synaptic associative weight adjustment models. They derive data and results from biophysics, electrophysiology and molecular biology while the theory itself results in an abstract, and highly functional model. The proposed neuron model will allow to realize self-programming of neurons, support modularization into ensembles or groups, and instantiate complex models of cognition. The brain, of course, has many additional components, such as glia cells (astrocytes, oligodendrocytes), the capillary and glymphatic networks, and the signaling molecules that pass the blood-brain barrier. To reach the goal of even more comprehensive brain theories we need first to build models with the full capacity of the neuronal cell.
Theoretical neuroscience has focused on synaptic weights as the main memory mechanism for a considerable time. There are several reasons for this. The neuron is a highly specialized cell type, strongly compartmentalized with extensive axons and dendrites. The synapse is a highly specialized connective element. Several types of neurons have spiny extrusions of the dendritic membrane with several synapses, spine shaft elements, and internal proteins and RNA which develop and disappear within hours or days. The postsynaptic density protein composition, and the presynaptic vesicle release mechanism are very intricate cellular structures. But the experimental literature on neuronal plasticity shows that synapses are an abstract constructive element, and there are other components of neuronal plasticity (West and Greenberg 2011; Mahon et al. 2003; Debanne et al. 2019; Sehgal et al. 2014; Nataraj et al. 2010; Gill and Hansel 2020; Scheler 2004b, 2014c; Tully et al. 2014). Of particular relevance are plasticity via ion channels and NM receptors. The internal signaling system constitutes a processing layer with access both to long-term information in the nucleus and incoming signals at the membrane from the horizontal network interactions (West and Greenberg 2011). Therefore we focus on the neuron as the relevant unit for plasticity. Glia-induced and other forms of non-neuronal plasticity are not discussed here, even though they add to functioning neural ensembles. The development of a theoretical framework, linking all those aspects of neural plasticity, is a difficult task. We have begun by re-thinking the concept of neural plasticity from the perspective of the individual neuron. We believe that synaptic plasticity is incompletely understood when placed outside of the context of the individual neuron and its cellular functioning.
Plasticity has to deal with the problem of stability vs. flexibility, such as overwriting information over time. Mechanical, direct transmission could provide a backbone of stability. There is also the problem of selecting which information to keep. Since neurons have internal components which set and re-set responsivity, these problems can be solved within a neuron-centric framework. We have also put forward a simple example from disease modeling where the internal dimension of the neuron and plasticity in protein expression have been shown to be of significance.
Synaptic weight adaptation models store the history of the use of the individual synapse, no matter whether by an associative or any other update rule (such as backpropagation (Rumelhart et al. 1986)). That is very restrictive. Huge networks with billions of parameters are needed to solve simple problems by storing millions of patterns. These restrictions are systematic and mathematically explainable, since the expressive power of networks with adjustable weights is weak. A programmable memory at each neuronal site allows for more complex and concise operations. For instance, patterns can get stored into a small set of dedicated neurons (ensembles) for a particular problem. These ensembles can be activated when similar problems have to be solved, and by modularization ever new configurations of ensembles to larger knowledge structures can be constructed. By separating transmission and storage of information (not every transmission event leaves a trace), we can focus on the problems of processing and memorization separately, and achieve a new kind of modular system within the field of large-scale parallel and distributed computing architectures.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)