Keywords
Lavender, induction callus, biostimulants, hormonal, physical, biological.
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Lavender, induction callus, biostimulants, hormonal, physical, biological.
This revised version incorporates substantial improvements in response to the reviewers' comments. The Introduction has been expanded to provide a clearer scientific background and a more comprehensive discussion of the roles of plant growth regulators, physical factors, culture media, and chitosan in callus induction and growth. The Discussion has been strengthened by incorporating additional recent references and providing deeper interpretation and comparison with previous studies. The manuscript has also undergone extensive language editing to improve clarity, coherence, and scientific style. Minor revisions were made throughout the text to enhance readability and consistency. These revisions improve the overall quality of the manuscript without altering the experimental design, results, or conclusions.
See the authors' detailed response to the review by Tassanai Jaruwattanaphan
See the authors' detailed response to the review by Huda Enaya Mahood
See the authors' detailed response to the review by Duha Mysire Majeed
Lavender (Lavandula angustifolia Mill.) is a member of the Lamiaceae family of flowering plants and is the most widely cultivated of the family, which also has over 39 other genera (Vârban et al., 2022; Kumar et al., 2025). The varied secondary metabolites in lavender provide a broad range of industrial uses in the pharmaceutical, cosmetic, and food industries (Crisan et al., 2023). The medicinal value of lavender has been well established over the centuries, considering the wide range of advanced herbal diseases and its other numerous pharmacological attributes, including antimicrobial, antifungal, antispasmodic and other active antioxidant (Hassiotis and Vlachonasios, 2025) as well as digestive function regulating (Firoozeei et al., 2021) properties. It is also used to treat gastrointestinal disorders, cardiovascular diseases, and respiratory and urinary disorders, anxiety, depression, and sleep disorders (Kumar et al., 2025), mood swings (Mehrabian et al., 2022), chronic pain (Pehlivan and Karadakovan, 2019), restless legs syndrome (Ghasemi et al., 2021), migraines (Yuan et al., 2021), and behavioural disorders in dementia patients. (Scuteri et al., 2019) as well as improving physical symptoms such as blood pressure and respiratory rate (Contrada et al., 2021).
There are difficulties in the plant based pharmaceutical industry related to the sourcing of raw materials attributable to the isolation of active compounds being arduous due to plants being low in active compounds or active compounds being affected by climatic conditions. Under these circumstances, the cultivation of plant tissues offers exciting possibilities, as it promotes the incessant active compounds synthesis irrespective of the season. This facilitates the overcoming of climatic and geographical limitations, hence aiding in the supply of growing the demand for medicinal and aromatic plants. This is one of the refined and improved strategies in the biotechnological domain that aims or is directed towards the amplified generation of secondary metabolites relevant for the industry (Ozyigit et al., 2023). This necessitates the adjustment and calibration of every single component in every single process of culture tissue namely, nutrient medium, the different and varied growth hormones, and the different types of stimulants to maximise biomass and secondary metabolites (Adil et al., 2025). The technology of callus culture is one of the sustainable methods for the valuable plant compounds and for the preservation of the habitat of the medicinal plants (Abed et al., 2020; Babich et al., 2020; Alrawi et al., 2022). Compounds can be derived from callus tissue in a lab setting without having to use whole plants, and this method can be commercially scaled by employing bioreactors for compound bioprocessing. In vitro callus tissue development depends on a number of factors; for example, a plant’s genetics, the plant organ type, and the composition of the nutrient medium and the growth regulators (Miri and Roughani, 2018). Furthermore, callus culture technology depends on the growth and sustained production and the qualitative and quantitative production of active compounds. The optimum environment must also be available. The importance of growth regulators during callus formation is significant because, in some cases, a tissue may be unresponsive or may even die if the medium’s hormone levels are set higher or lower than the tissue’s internal level (Yegorova et al., 2020). Previous research indicates that the callus formation is more pronounced when the nutrient medium is enhanced with growth regulators, specifically auxins and cytokinins (Jabr and Mohsen, 2024). The optimization of the hormonal milieu is only attained with the moderators acting synergistically within set ranges. Compared to using each of them separately (Tilkat and Onay, 2009). The balance between auxins and cytokinins in the nutrient medium is a key determinant that stimulates cell division and initiates callus formation.
Despite the central role of plant growth regulators in controlling cellular differentiation and morphogenesis, callus formation is influenced by multiple interacting factors, including hormonal signals (Thiry et al., 2026), physical conditions, culture medium composition, and biostimulants (Pasternak et al., 2024). Among physical factors, light plays a key role in regulating cellular reprogramming through its effects on auxin stability, hormonal balance, and primary metabolism, thereby influencing dedifferentiation efficiency (Cioć et al., 2022; Miguel, 2025). However, these responses vary considerably among plant species (Liu et al., 2025), which limits the generalization of results. Culture medium composition is another critical factor governing morphogenic responses. Even minor changes in ionic balance, organic additives, or nitrogen sources can lead to noticeable shifts in growth patterns (Okao et al., 2025). Therefore, culture media should be considered dynamic systems whose effectiveness depends on species-specific physiological requirements rather than fixed formulations. Biostimulants have recently gained attention as supplementary regulators of plant growth responses. Chitosan, a deacety-lated form of chitin, has been widely studied due to its ability to trigger defense-related pathways and influence secondary metabolism (Javed et al., 2025; Iber et al., 2022). However, its mode of action in relation to hormonal signaling and environmental interactions is still not fully understood, particularly under in vitro conditions. Overall, callus formation results from the interaction of several factors rather than a single regulatory pathway (Liu et al., 2025). Most studies still examine these factors separately, which limits understanding of their combined effects. Although Lavandula spp. are economically and pharmacologically important as sources of essential oils and bioactive compounds, the regulatory mechanisms controlling callus induction and stability in vitro remain poorly defined. This study therefore investigates the combined effects of plant growth regulators, light conditions, culture medium composition, and chitosan on callus formation and growth in lavender.
Lavender seeds were obtained from a specialised seed production company Al-Wadi seed Company located south of the capital Baghdad, Iraq. The seeds were stored in a laboratory at a temperature of 15°±2 C until use.
Medium preparation
The culture medium used in the present study consisted of half MS salts (Murashige and Skoog, 1962), medium supplemented with 30 g. L−1 sucrose as a carbon source. Gibberellic acid was added to break seed dormancy at a concentration of 0.5 mg L−1. The pH of the medium was adjusted to 5.7–5.8, and agar at 7 g. L−1 was used as a solidifying agent. The prepared medium was dispensed into culture vials, with each vial containing 10 mL of medium. All culture vessels were autoclaved at 121°C and 15 psi for 15 min for sterilization. The medium was then allowed to solidify in the vials prior to inoculation.
The medium was distributed into glass culture vessels 10 ml vessel. The containers were sealed with tight fitting lids and sterilised in an autoclave Model ABC at a temperature of 121°C and a pressure of 1.04 kg cm for 15 minutes. The sterilised medium was stored at 25°C until use.
Seed sterilization
The seeds were sterilised and thoroughly washed under running tap water for 10 minutes to remove surface debris. The healthy seeds were transferred to a laminar air cabinet, then a 3% sodium hypochlorite solution was added for surface sterilization for 15 minutes with continuous stirring. The seeds were rinsed with distilled water for 5 minutes, and the washing process was repeated three times to ensure complete removal of the sterilizing agent (Al-Alwani and Mohammed, 2022).
Seed germination
The sterilised seeds were planted on the prepared nutrient medium. The planted seeds were then transferred to the growth chamber at a temperature of 25°C and a light intensity of 1000 lux for 16 hours of light and 8 hours of darkness (Sekhi et al., 2025). After four weeks, the seedlings were transferred to MS medium (hormone-free) and incubated under the same conditions for four weeks, seedlings ready for use were obtained for callus induction experiments. The chemicals listed in Table 1 were used in this study.
The experiment was conducted in three successive phases, each designed to achieve a specific objective. Each subsequent phase built upon the results of the previous phase ( Figure 1), to evaluate callus production efficiency and the growth and enhancement of biomass production. The following experiments were conducted.
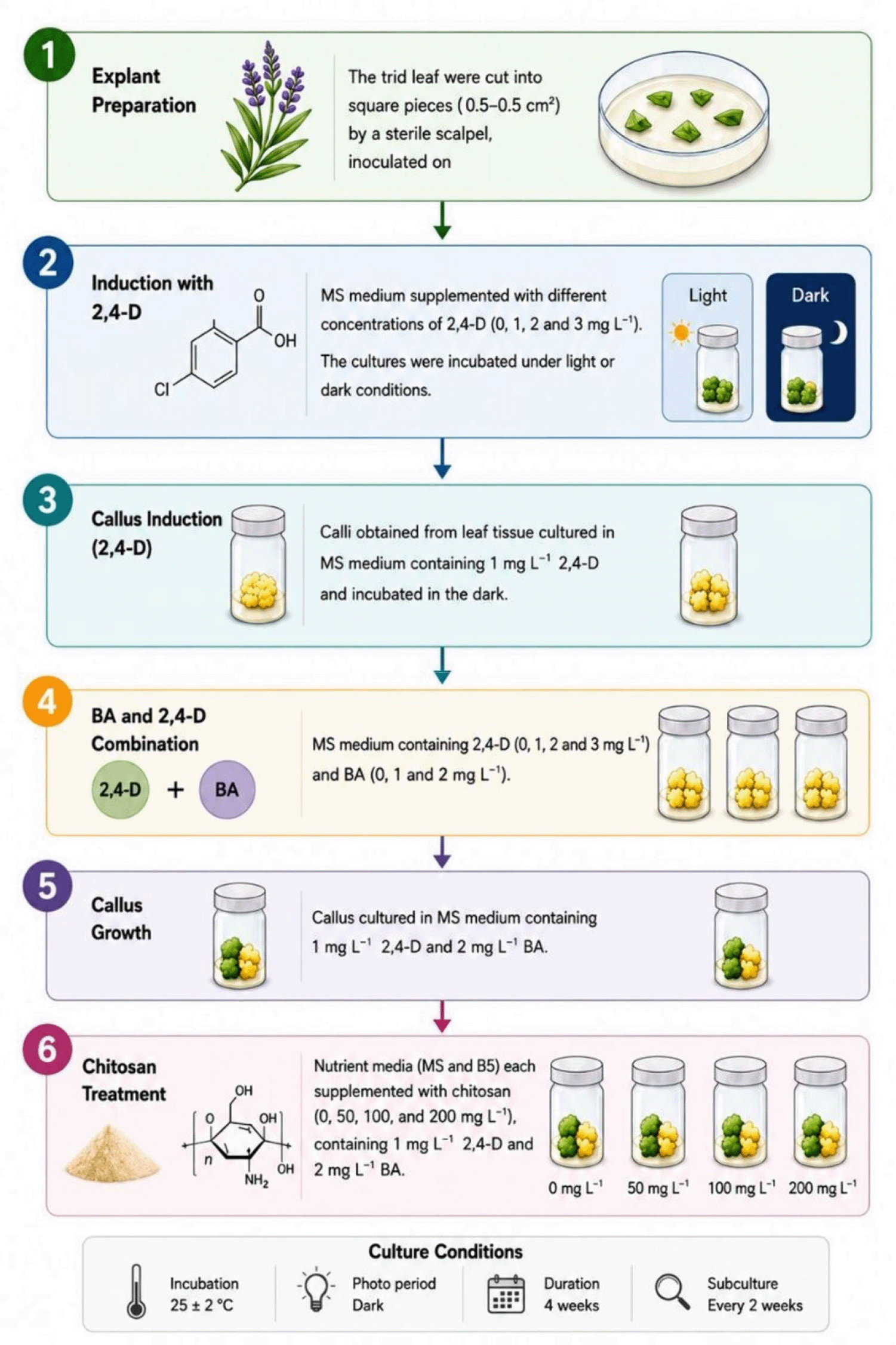
(1) Callus induction
A nutrient medium based on full MS salts strength was prepared, with varying concentrations of the growth regulator dichlorophenoxyacetic acid (2,4-D) (0, 1, 2 and 3) mg L−1. Then 30 g L−1 of sucrose was added to the medium as a carbon source and 7 g L−1 of agar as a stabilising agent. The nutrient medium was then sterilised according to the standard protocol. After preparing the media The tird leaf from the growing tip (the four weeks-old healthy tissue culture lavender plants) have used as explants. The leaves tissue were cut into a square pieces measuring 0.5*0.5 cm2 by a sterile scalpel on the clean bench and afterword inoculated on the MS medium the specified concentrations of the growth regulator 2,4-D. The cultures were incubated in a growth chamber with controlled conditions at a constant temperature of 25 C. To investigate the effect of light on callus induction, the cultures were exposed to two different lighting regimes complete darkness and 1000 lux light intensity under a cyclic light regime 16 hours of light 8 hours of darkness. At the end of the four-week incubation period, data were collected by measuring Induction rate the percentage of tissue sections in which callus was formed, Fresh weight and Dry weight.
(2) Callus Proliferation
Calli (150 mg in fresh weight), induced from leaves tissue on the MS medium containing 1 mg.L−1 2,4-D for 4 weeks, were used. These calli were transferred to the MS medium supplemented with 30 g.L−1 sucrose, 7g.L−1 agar and 2,4-D at 0, 1, 2 and 3 mg.L−1 in combination with 0, 1 and 2 mg.L−1 BA. The cultures were incubated in complete darkness at a constant temperature of 25°C for four weeks. At the end of the incubation period, data were collected by measuring the fresh weight and then the dry weight of the callus growth.
(3) The role of nutrient components and chitosan in promoting callus growth
Based on the best combination was reported in the previous experiment, 150 g of 4-week-old propagated calli (cultured on MS medium supplemented with a combination of 1 mg of 2,4-D and 2 mg of BA) were transferred to two media (B5 and MS). Each media was supplied with four concentrations of chitosan (0, 50, 100, and 150) mg, with the addition of 1 mg.L−1 of 2,4-D and 2 mg of BA, 30 g sugar and 7 Agar. After a four-week incubation period, growth rates were determined by measuring the fresh and dry weights of the callus formed.
The data were statistically analysed using GenStat statistical software (Gen 12 ed) (Payne et al., 2009) according to a completely randomised design (C.R.D.), with ten replicates for each treatment. The treatments were arranged in a factorial arrangement. The means were compared on LSD test after partitioning the variance components via fisher’s test by using probability ≤ 0.05 whereas, the callus percentage was analysed using originPro 2025b (origin 2025).
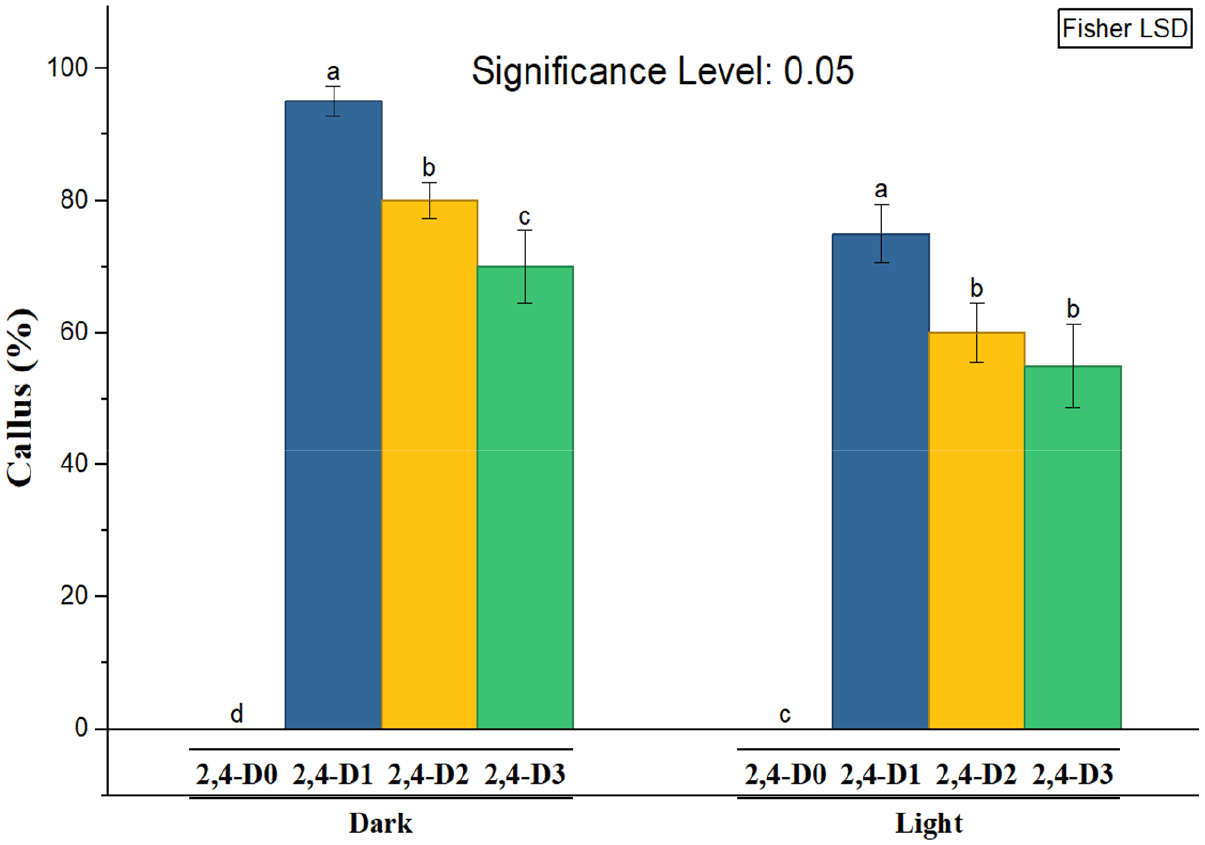
Statistical analysis, as shown in Figure 1 revealed that the growth regulator 2,4-D had a significant effect on callus induction. It was found that the optimal concentration for callus formation from leaf tissue was 1 mg L−1 of 2,4-D added to the nutrient medium, which reached 95%, while an increase in concentration above this level led to a significant decrease in the rate of callus formation.
On the other hand, lighting had a relatively limited effect on callus induction compared to the effect of 2,4-D growth regulator concentration. The highest callus production values were recorded under complete darkness, and all auxin containing treatments outperformed their counterparts grown under lighting conditions ( Figure 1).
The results of statistical analysis according to the data shown in Table 2 showed a statistically significant difference in the fresh weight values of callus induced from lavender leaf tissue, where the fresh weight of callus differed according to concentration (0, 1, 2 and 3) mg L−1 of 2,4-D added to the MS nutrient medium. The auxin concentration of 1 mg L−1 showed the highest significant value for callus fresh weight, which reached 0.796 g. An increase in auxin concentration above this level led to a gradual decrease in the rate of callus formation, indicating that high auxin concentrations have an inhibitory effect on the callus formation process. It should be noted that the auxin free treatment 0 concentration did not show any visible or statistical response in callus formation, confirming the pivotal role of auxin in initiating induction and stimulating cell division in plant leaf tissues. As for the effect of lighting, the results showed a clear superiority of callus growth under conditions of complete darkness for all concentrations studied, with the average fresh weight values under these conditions showing a remarkable superiority of 0.557 g compared to their counterparts grown under lighting conditions, which reached 0.527 g. The effect of the light factor can be observed in Figure 3, where the callus appeared dark greenish to gray (friable, granular, comact), while in the dark, the callus appeared Pale yellowish to green (compact dense less granular). Callus grown in darkness was also larger and more developed, making it suitable for further experiments on callus cultures.
| Light conditions | 2,4-D (mg.L−1) | Mean | |||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | ||
| Dark | 0.000 d | 0.830 a | 0.752 b | 0.647 c | 0.557 a |
| Light | 0.000 d | 0.762 b | 0.723 bc | 0.624 c | 0.527 b |
| Mean | 0.000 d | 0.796 a | 0.737 b | 0.635 c | |
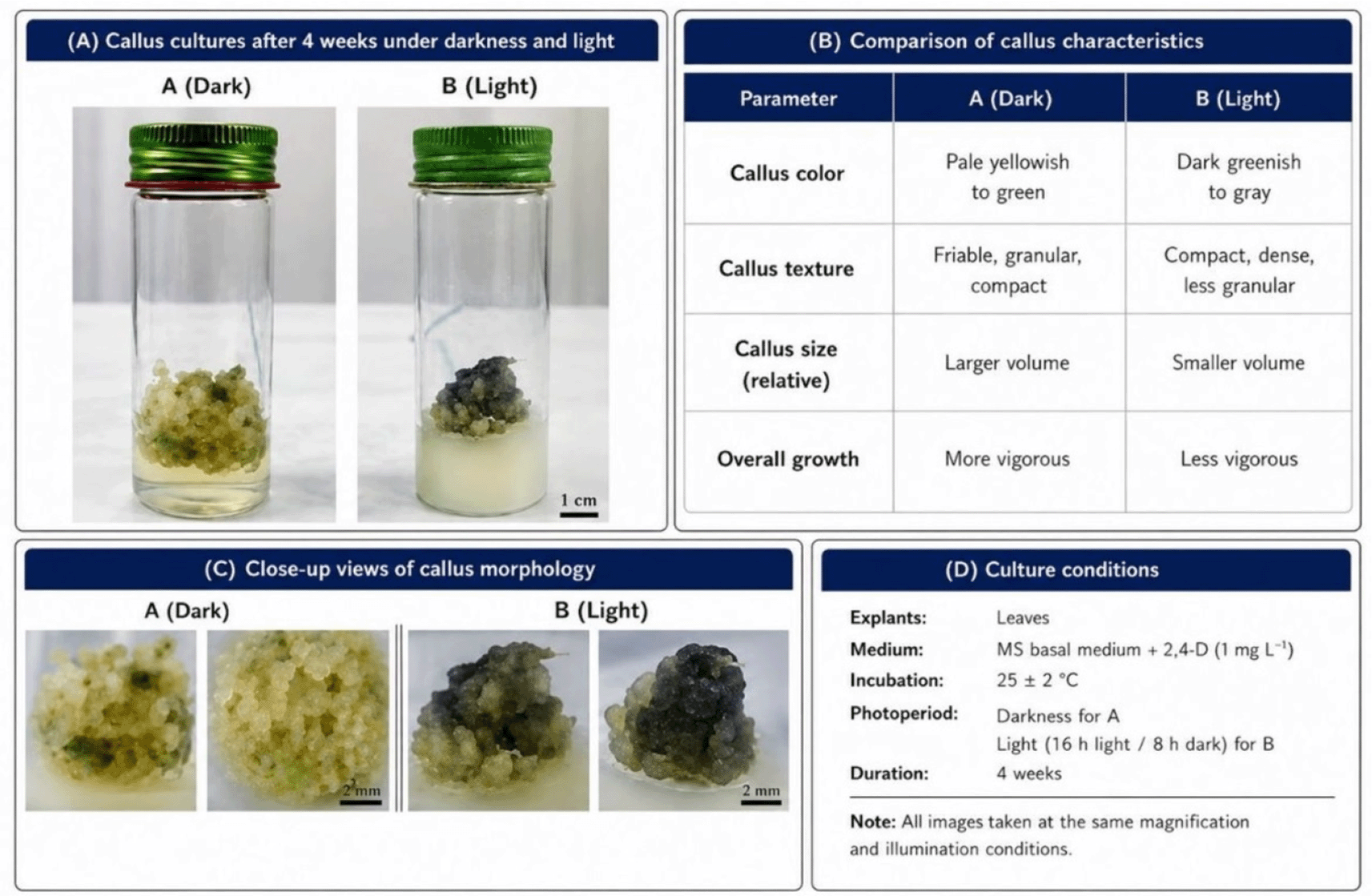
(A): Callus cultures after 4 weeks under darkness and light. (B): Comparision of callus charcteristics. (C): Close-up views of callus morphology. (D) Culture conditions.
It can be concluded that the process of callus induction from lavender leaf tissue requires a delicate balance between the optimal concentration of auxin and suitable environmental conditions, emphasising the complex interaction between hormonal and environmental factors in regulating growth and differentiation processes in plant cells. The results also show that the highest fresh weight of callus was achieved at a concentration of 1 mg L−1 of regulator 2,4-D when grown under dark conditions, reaching 0.830 g. While no value was recorded for fresh weight of callus (0 g) in treatments free of auxin under dark or light conditions. This confirms the importance of auxin as a basic factor in callus formation.
Effect of auxin on dry weight of callus the results of statistical analysis shown in Table 3. Indicate a clear variation in biological response when treating lavender (Lavandula angustifolia Mill.) leaf tissues with different concentrations of the growth regulator 2,4-D. A concentration of 1 mg L−1 of 2, 4-D recorded the highest significant dry weight of callus 0.064 g. An increase in concentration to 2 mg L−1 resulted in a significant decrease in dry weight 0.058 g, while a concentration of 3 mg L−1 recorded a decrease of 0.046 g. The auxin free treatment did not show any visible or statistical response in callus formation, confirming the pivotal role of auxin in the process of induction and initiation of cell division.
| Light conditions | 2,4-D (mg L−1) | Mean | |||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | ||
| Dark | 0.000 e | 0.068 a | 0.059 b | 0.048 c | 0.044 a |
| Light | 0.000 e | 0.061 b | 0.057 bc | 0.044 d | 0.040 b |
| Mean | 0.000 d | 0.064 a | 0.058 b | 0.046 c | |
Statistical analysis showed a significant interaction between auxin concentration and lighting conditions. Treatments under lighting conditions recorded an average dry weight of 0.040 g, while treatments in complete darkness significantly exceeded this with an average dry weight of 0.044 g.
The results in Table 4, Showed that adding synthetic auxin of type 2,4-D at different concentrations to the nutrient medium had a significant statistically significant effect on the fresh weight of the callus formed. A clear response to the concentrations added to the nutrient medium was observed, with the low concentration 1 mg L−1 outperforming all other treatments, recording the highest significant value for fresh weight of 0.920 g. With an increase in hormone concentration to 2 and then 3 mg L−1, a gradual and significant decline in growth efficiency was observed, with the fresh weight of callus decreasing to 0.811 g and 0.632 g, respectively. This decrease indicates the possibility of an inhibitory effect of high concentrations of auxin on the process of development and growth.
| BA (mg L−1) | 2,4-D (mg L−1) | Mean | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| 0 | 0.838 c | 0.728 d | 0.628 f | 0.731 c |
| 1 | 0.880 b | 0.843 c | 0.636 ef | 0.786 b |
| 2 | 1.042 a | 0.863 bc | 0.634 ef | 0.846 a |
| Mean | 0.920 a | 0.811 b | 0.632 c | |
On the other hand, the results also showed statistically significant differences between the different concentrations of cytokinin benzyladenine (BA) added to the nutrient medium. The effect of BA was characterised by a positive linear relationship, where the growth rate of callus measured by fresh weight increased significantly with increasing hormone concentration ( Figure 4). Growth peaked at a concentration of 2 mg L−1, achieving the highest fresh weight value of 0.846 g. In contrast, the treatment containing a concentration of 0 mg L−1 free of cytokinin recorded the lowest fresh weight value of 0.731 g, highlighting the vital importance of cytokinin in supporting cell division and callus formation. The most significant scientific importance of this study lies in evaluating the complementary and reciprocal effects of these two hormones when used together. The results showed a clear superiority of some combinations over others. The addition of a combination of auxin 2,4-D at a concentration of 1 mg L−1 in conjunction with cytokinin BA at a concentration of 2 mg L−1 resulted in the highest total value of 1.041 g. This result reflects a synergistic effect between the two hormones, each enhancing the effect of the other, leading to optimal stimulation of growth and development processes. In contrast, the combination of auxin 2,4-D at a concentration of 3 mg.L−1 with cytokinin at a concentration of 0 mg.L−1 recorded the lowest fresh weight value, which was 0.628 g. This poor performance confirms the negative effects of high auxin concentrations in the absence of a balancing factor such as cytokinin, supporting the hypothesis that the balance between these two hormones, rather than the presence of only one, is the decisive factor in directing plant cell growth and differentiation in tissue cultures.

The results in Table 5 showed a clear response when auxin was added at different concentrations to the nutrient medium. The concentration of 1 mg L−1 significantly outperformed all other treatments, recording the highest dry weight of callus at 0.064 g. This result indicates that this concentration is optimal for stimulating cell division and hypertrophy, which positively affects biomass accumulation. However, a gradual and significant decline in growth efficiency was observed with increasing auxin concentration. Dry weight decreased to 0.060 g at a concentration of 2 mg L−1 and to 0.044 g at a concentration of 3 mg L−1. This decrease indicates that higher concentrations of auxin may have an inhibitory or toxic effect on cells, hindering their growth and limiting callus formation, which is common in plant responses to growth regulators when the optimum limit is exceeded.
| BA (mg L−1) | 2,4-D (mg L−1) | Mean | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| 0 | 0.065 ab | 0.055 cd | 0.041 f | 0.054 c |
| 1 | 0.062 bc | 0.060 bc | 0.042 ef | 0.055 bc |
| 2 | 0.070 a | 0.065 ab | 0.050 de | 0.062 a |
| Mean | 0.065 a | 0.060 b | 0.044 c | |
The results showed statistically significant differences in the dry weight of callus when different concentrations of cytokinin were added. The effect of cytokinin was characterised by a positive linear relationship, with the dry weight of induced callus increasing significantly with increasing hormone concentration. This effect peaked at a concentration of 2 mg L−1, achieving the highest dry weight value of 0.062 g. This confirms the vital role of cytokinin in regulating cell division and differentiation, which is a complementary and important factor in the callus induction process.
The greatest scientific value of this study lies in assessing the combined and complementary effects of auxin and cytokinin hormones, as it is known that the balance between them is the main driver of plant cell growth. In this context, the results showed a significant advantage for the combination of auxin at a concentration of 1 mg.L−1 with cytokinin at a concentration of 2 mg.L−1. This combination achieved the highest dry weight value for callus 0.070 g. This outstanding performance is attributed to the synergistic effect between the two hormones, where auxin at its optimal concentration 1 mg.L−1 stimulates cell division and callus formation, while cytokinin at its optimal concentration 2 mg L−1 promotes cell division which together leads to optimal cell mass enlargement and higher dry matter accumulation.
In contrast, the combination with the highest concentration of auxin 3 mg.L−1 and a complete absence of cytokinin 0 mg L−1 recorded the lowest dry weight value, at 0.041 g. These results confirm that the success of callus induction and its maximum growth measured by dry weight depend critically on the selection of the optimal concentration of each plant regulator separately and more importantly, on the selection of the optimal combination of the two. The interaction between auxin and cytokinin is what directs the final physiological response of plant tissues.
The results of statistical analysis, shown in Table 6, revealed a clear variation in the components of different nutrient media in stimulating callus growth. The comparison showed that the nutrient medium B5 significantly outperformed the other media, recording the highest average fresh weight of callus induced from leaf tissue, with a value of 2.197 g. In contrast, the MS medium showed the lowest fresh weight growth rate for the same type of callus, with a value not exceeding 1.233 g.
| Medium components | Chitosan (mg. L−1) | Mean | |||
|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | ||
| MS | 1.000 f | 1.206 e | 1.904 c | 0.824 g | 1.233 b |
| B5 | 1.892 c | 2.374 a | 2.392 a | 2.131 b | 2.197 a |
| Mean | 1.446 d | 1.790 c | 2.148 a | 1.477 d | |
As for the results related to the analysis of the effect of chitosan concentration, presented in Table 6. They showed statistically significant differences between the four concentrations studied (0, 50, 100 and 200) mg L−1 was observed that all tested concentrations of chitosan significantly outperformed the control group ( Figure 5), confirming the positive effect of this substance in stimulating soft weight gain in callus. This effect followed a clear pattern of proportionality, as the rate of soft weight gain in calves increased significantly with increasing chitosan concentration, reaching its highest value at a concentration of 100 mg L−1, with a peak of 2.148 g. Performance reversed significantly when the highest concentration 200 mg L−1, where the fresh weight of the callus decreased compared to the optimal concentration 100 mg L−1, indicating that the positive effect of chitosan is at its maximum at a specific concentration, and that increasing the concentration beyond this limit may lead to adverse results.

The results of statistical analysis, as shown in Table 7, revealed a clear variation in the performance of nutrient medium components in terms of their effect on the dry weight of callus. The results showed a marked superiority of the B5 diet in achieving the highest average dry weight of callus induced from leaf tissue, recording a value of 0.103 g. In contrast, the nutrient medium MS recorded the lowest dry weight, with a value of 0.076 g. indicating that the composition of the nutrient medium B5 was more suitable and effective in promoting growth and dry matter accumulation in callus compared to the medium MS.
| Medium components | Chitosan (mg. L−1) | Mean | |||
|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | ||
| MS | 0.068 e | 0.071 d | 0.087 c | 0.081 cd | 0.076 b |
| B5 | 0.101 b | 0.103 ab | 0.106 a | 0.102 ab | 0.103 a |
| Mean | 0.084 c | 0.087 b | 0.096 a | 0.091 ab | |
As for the results shown in Table 7, Statistical analysis revealed statistically significant differences between the different concentrations of chitosan included in the study (0, 50, 100 and 200) mg L−1. It was observed that all concentrations treated with chitosan significantly outperformed the control group, to which no concentration of the substance was added 0 mg L−1. This superiority confirms the stimulating and positive effect of chitosan on callus growth.
The results also revealed a positive correlation between increased chitosan concentration and the dry weight of induced callus, with values increasing steadily with higher concentrations, peaking at a concentration of 100 mg L−1. The average dry weight of the callus reached its highest value, amounting to 0.096 g. A decline in this positive effect was also observed when using the highest concentration 200 mg L−1, where the average dry weight decreased to 0.091 g. This decrease indicates that the stimulating effect of chitosan follows an optimum pattern, achieving maximum growth response, and that exceeding this concentration may have an inhibitory or less effective effect on the accumulation of dry matter in callus.
Growth regulators, especially auxins, are key factors in stimulating callus formation. The addition of auxin to the nutrient medium elicited a response in the leaf tissues of lavender plants, stimulating callus formation. The added auxin induces fundamental changes in the plant cells in contact with the medium and stimulates the initiation of cell division. During this stimulation phase, constructive processes such as protein synthesis and DNA replication are activated, followed by a series of changes in cellular activity that culminate in cell division and the formation of a callus mass covering most parts of the plant fragment.
The response of plant tissue varies depending on the concentration of auxin in the nutrient medium. An increase in concentration promotes callus formation until the optimum concentration of 1 mg L−1 is reached, which achieved the highest callus formation rate 95%, in addition to an increase in fresh and dry weight. Conversely, exceeding this optimal concentration, as observed at concentrations of 2 and 3 mg L−1, leads to adverse effects that inhibit growth. The results also showed that light acted as an inhibitory factor for callus growth induced from lavender leaves, whereas darkness provided more favorable conditions for callus induction and growth (Bojko et al., 2024). This may be attributed to improved auxin stability under dark conditions, reduced photo-oxidative stress, and reduced early cell differentiation, thereby supporting cellular dedifferentiation and enhancing callus proliferation (Bojko et al., 2024). In contrast, light conditions may negatively affect auxin stability, thereby reducing its effectiveness and contributing to the lower level of callus formation observed under light. Light also had a clear effect not only on growth but also on the morphological characteristics of the callus. Callus cultured under light conditions was characterized by a dark green to gray color and a brittle, granular, and compact texture, whereas callus induced under dark conditions appeared pale yellow to green with a compact, dense, and less granular structure ( Figure 3). It was also observed that callus grown in darkness was larger and more developed compared to that grown under light conditions, which gives it an advantage in this study. However, the effect of light on callus growth is complex and may vary depending on the plant species and culture conditions. Similar responses have been reported in several plant species, confirming the regulatory role of darkness in morphogenic responses during in vitro culture (Keykha et al., 2014; Suhartanto et al., 2022; Bojko et al., 2024). It is important to note that continuing to supplement the nutrient medium with auxin at the optimal concentration in order to increase callus production may inhibit growth due to auxin accumulation, necessitating the addition of cytokinin. Cytokinin plays an important role in sustaining callus growth. and previous studies indicate that the interaction between auxins and cytokinins at different concentrations is a decisive factor in achieving optimal hormonal balance (Jabr and Mohsen, 2024; Tilkat and Onay, 2009). In this regard, the treatment consisting of 1 mg L−1 auxin and 2 mg L−1 benzyladenine BA recorded the highest fresh and dry weight values, indicating that it is a suitable combination for sustained callus growth. This can be explained by the ability of cytokinins, especially benzyladenine, to promote nutrient uptake, stimulate cell division and inhibit protein degradation, which supports the division process when the appropriate hormonal balance is achieved. Cytokinins also promote the synthesis of RNA, proteins, and enzymes within the cell. Overall, the increase in soft and dry weight of callus reflects changes in cellular components such as carbohydrates, proteins and amino acids associated with the process of division and growth (Bulya et al., 2023). Therefore, the balance between auxins and cytokinins in the nutrient medium remains a key determinant of continued cell division and callus growth.
In addition to plant growth regulators, the basal culture medium significantly influenced both fresh and dry biomass accumulation, where Gamborg’s B5 medium showed superior performance compared with MS medium. This difference is mainly attributed to variations in the nutritional composition of the two media. Gamborg’s B5 medium provides higher levels of potassium nitrate along with additional nitrogen sources such as ammonium sulfate, which contribute to improved nitrogen assimilation, protein biosynthesis, and enhanced morphogenic potential in plant tissue cultures (Yin et al., 2017; Bari et al., 2022; Riba, 2026). In addition, its relatively higher thiamine concentration may support key enzymatic functions and metabolic pathways required for active cell proliferation and tissue differentiation (Fitzpatrick et al., 2020; Vollmer et al., 2023; Riba, 2026). Overall, plant tissue responses to culture media are highly species-dependent, suggesting that Gamborg’s B5 medium may be more suitable than MS medium for specific plant species (Kurmi et al., 2020; Riba, 2026).
Bioactive chitosan was also used as a callus growth stimulator, and the results were consistent with previous studies on the role of chitosan in stimulating growth (Pirbalouti et al., 2017; Byczyńska, 2018; El-Khateeb et al., 2018; Atteya et al., 2023). chitosan treatments recorded significant increases in growth indicators and a positive correlation was observed between chitosan concentration up to a certain level and callus growth, with the highest fresh and dry weight values recorded at a concentration of 100 mg L−1. This stimulation is attributed to the biological properties of chitosan, which enhances the absorption of key nutrients such as nitrogen and phosphorus, thereby supporting metabolic processes and the accumulation of organic matter. This was reflected in an increase in fresh weight due to higher water and organic matter content and dry weight due to the accumulation of proteins and carbohydrates, resulting in improved biomass of the callus. Stimulation must be limited and sensitive to concentration; at a concentration of 200 mg L−1, growth indicators decreased significantly, and this decrease can be explained physiologically by ionic disturbance and inhibition of nutrient absorption.
Callus formation in Lavandula angustifolia is influenced by several interacting factors, including plant growth regulators, light, culture medium, and chitosan. These factors work together and affect how cells dedifferentiate and develop into callus. Both light conditions and nutrient composition of the medium play an important role in this process, while chitosan contributes to improving overall plant physiological responses. In general, successful callus induction requires considering all these factors together as one system rather than separately. This helps improve the efficiency and stability of tissue culture in lavender and supports its use in micropropagation and secondary metabolite production.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)