Keywords
Apocynin · Selenite · Cataract · Oxidative Stress
This article is included in the Eye Health gateway.
Apocynin · Selenite · Cataract · Oxidative Stress
In response to the reviewers’ comments, the manuscript has been revised to improve clarity, methodological transparency, and data presentation. Statistical annotations in tables and figures were clarified to ensure that all results are fully self-explanatory, and the rationale for the experimental design, including the DMSO control group, was explicitly defined. In addition, the presentation of cataract grading data was refined to reflect its ordinal nature, and relevant sections were edited to improve language and readability.
See the authors' detailed response to the review by Syska Widyawati
See the authors' detailed response to the review by Rajesh Choudhary
Cataract, defined as the opacification of the lens, is the leading cause of reversible blindness, accounting for approximately 51% of all cases worldwide.1 Although modern cataract surgery is generally regarded as highly safe and remains the only effective treatment for cataracts, it is not always accessible or affordable. Moreover, potential intraoperative and postoperative complications further highlight the need for non-surgical alternatives.2 Oxidative stress is a major contributor to cataractogenesis, primarily through the imbalance between reactive oxygen species (ROS) and endogenous antioxidant defence systems such as CAT, superoxide dismutase (SOD), and glutathione peroxidase (GPx).3 The accumulation of ROS promotes lipid peroxidation, protein aggregation, DNA damage, and apoptosis of lens epithelial cells, all of which contribute to lens opacification.3–5 Recent metabolomic studies also indicate that cataract formation is accompanied by disruptions in glutathione-related pathways and other antioxidant-linked metabolites, further supporting the central role of oxidative imbalance in cataractogenesis.6 Given its central role in cataract development, antioxidant therapy has been widely proposed as a promising non-surgical approach for delaying or preventing disease progression.7 Various compounds, including sildenafil, melatonin, ascorbic acid, resveratrol, ellagic acid, N-acetylcysteine and rosmarinic acid have demonstrated protective effects in experimental cataract models.2,4,8–12
Of these models used in previous research, the selenite-induced cataract model in rats is one of the most extensively utilized due to its reproducibility and close resemblance to age-related human cataracts.2,4 Selenite administration disrupts calcium homeostasis, accelerates proteolysis, and depletes glutathione levels, thereby mimicking the molecular and structural alterations observed in human cataractogenesis.2,13
Apocynin (4-hydroxy-3-methoxyacetophenone) is a natural NADPH oxidase inhibitor derived from Picrorhiza kurroa and Apocynum cannabinum. It prevents the assembly of the NADPH oxidase complex and consequently reduces reactive oxygen species (ROS) production. Apocynin has been reported to exhibit antioxidant, anti-inflammatory, and cytoprotective effects in several systemic disease models.14 Previous studies have demonstrated its beneficial roles in cardiovascular disease, renal injury, neuroinflammation, and metabolic disorders, largely attributed to its capacity to suppress oxidative stress and modulate redox-sensitive signaling pathways.15 Furthermore, apocynin has been reported to enhance endogenous antioxidant defenses, preserve mitochondrial function, and attenuate inflammatory cytokine production in multiple in vivo models.16 Despite these documented systemic benefits, its potential therapeutic effects in ocular tissues, particularly in cataract formation, remain insufficiently explored, highlighting the need for further investigation.
The aim of this study was to investigate the potential protective effect of apocynin against selenite-induced cataract formation. In addition to assessing cataract severity, the impact on key oxidative stress biomarkers in the lens tissue was evaluated, including malondialdehyde (MDA), catalase (CAT), and glutathione reductase (GR). MDA is a well-established marker of lipid peroxidation and reflects structural damage caused by ROS,17 whereas CAT and GR are critical enzymatic components of the endogenous antioxidant defence system.18 These parameters were selected as they have been shown to be sensitive and relevant for the evaluation of redox imbalance in experimental cataract models.19 Based on this background, we hypothesized that apocynin may exert a protective effect against selenite-induced cataract formation through modulation of oxidative stress in lens tissue. An earlier version of this study was posted as a preprint.20
The establishment of the cataract rat model and other experimental procedures were conducted in the Experimental Animals Application and Research Centre of Abant Izzet Baysal University. Biochemical analyses were performed in the Biochemistry Laboratory of Basaksehir Cam and Sakura City Hospital. Prior to the study, ethical approval was obtained from the Animal Experiments Local Ethics Committee of Abant Izzet Baysal University (Date: 9th June 2021, Approval Number: 2021/19).
The study sample comprised a total of 35 healthy, male Sprague-Dawley rats, each weighing approximately 50 g on postnatal day 10, obtained from the Experimental Animals Research and Application Centre of Bolu Abant Izzet Baysal University. The experimental animals were kept in a laboratory room at a temperature of 20-24 °C and 40-60% humidity, with a 12 hr light-dark cycle. The rats had ad libitum access to standard feed and water, refreshed daily. All the rats were confirmed as having healthy eyes before starting the study.
A selenite-induced cataract model was established in each rat on postnatal day 10 with a single dose (30 nmol/gr) subcutaneous injection of sodium selenite (Na2SeO3). In the literature, this model has been reported to usually manifest with the development of bilateral cataracts in rats within 16 days after eye opening.2
The experimental timeline consisted of selenite administration on postnatal day 10, followed by daily treatment for 7 consecutive days, with morphological cataract assessment and tissue collection performed on day 14.
The rats were separated into 5 groups of 7 animals. Group allocation was performed using a simple randomization method based on a computer-generated random number sequence. The Control group (n = 7) received only saline solution subcutaneously. Group 1 (n = 7) received 30 nmol/gr Na2SeO3 (Sigma Aldrich, Merck Group, USA, Cat. No: 214485-100G) subcutaneously. Group 2 (n = 7) received 30 nmol/gr Na2SeO3 subcutaneously and dimethyl sulphoxide (DMSO) (Sigma Aldrich, Merck Group, USA, Cat. No: 472301-500ML) intraperitoneally. This group was included as a vehicle control to distinguish the potential effects of DMSO from those of apocynin treatment. Group 3 (n = 7) received 30 nmol/gr Na2SeO3 subcutaneously and 10 mg/kg apocynin (Sigma Aldrich, Merck Group, USA, Cat. No: 178385-1GM) intraperitoneally. Group 4 (n = 7) received 30 nmol/gr Na2SeO3 subcutaneously and 20 mg/kg apocynin intraperitoneally. The treatments (low- and high-dose apocynin) were administered daily for 7 consecutive days. Sodium selenite was dissolved in saline, and apocynin was dissolved in DMSO. No anaesthesia was administered to the rats during injections. The selected apocynin doses were based on previous in vivo studies reporting antioxidant and cytoprotective effects without overt toxicity, and were chosen to explore potential dose-dependent responses rather than to provide pharmacokinetic characterization.
The control group established baseline lens morphology and biochemical parameters. The selenite-only group confirmed cataract induction. The DMSO group controlled for solvent effects, while the low- and high-dose apocynin groups were included to assess dose-dependent effects on cataract severity and oxidative stress.14–16
At the end of 14 days, the eyes of all the rats were dilated with 1% tropicamide (Tropamid Forte®, Bilim Pharmaceuticals, Turkey). Ketalar® (50 mg/kg) (Ketamin hydrochloride, Pfizer Pfe Medicines, Germany) and xylazinebio® (10 mg/kg) (Xylazine, Bioveta, Czech Republic) were administered intramuscularly to induce anesthesia, and evisceration was performed. Euthanasia was then carried out by exsanguination after the surgery, under deep anesthesia, in accordance with institutional and international guidelines for the care and use of laboratory animals. The formation of cataract in each eye was examined under a biomicroscope (Topcon DC3 Japan) and the cataract was graded by coaxial illumination of an operating microscope (Zeiss Microsystem, Germany) using a seven-level grading system (0–6) ( Table 1) based on the extent and distribution of lens opacity:
Grade 0: Normal transparent lens
Grade 1: First signs of nuclear opacity with small scatterings
Grade 2: Mild nuclear opacity
Grade 3: Diffuse nuclear opacity accompanied by cortical scattering
Grade 4: Partial nuclear opacity
Grade 5: Distinct nuclear opacity
Grade 6: Mature cataract involving the entire lens
Magnification and illumination settings were kept constant for all examinations, and both direct and retroillumination views were obtained. Each lens was evaluated twice; in case of disagreement >1 grade, a third evaluation would be performed. To minimize bias, the order of animal assessment was randomized, and all observations were conducted by a single investigator blinded to group allocations. Although cataract grading involves an element of subjectivity, several measures were implemented to minimize observer-related bias, including standardized illumination and magnification, predefined grading criteria, repeated assessments, and masking of the examiner to group allocation.
The removed lens tissues were placed in liquid nitrogen and stored at -80°C for one week until the analysis. The rat lens tissues were sent to the Biochemistry Laboratory of the Department of Medical Microbiology, Basaksehir Cam and Sakura City Hospital, and the levels of malondialdehyde (MDA), glutathione reductase (GR), and catalase (CAT) were measured using the enzyme-linked immunosorbent assay (ELISA) method. Tissues were weighed and then homogenized in phosphate buffer solution (PBS; pH 7.4) [tissue weight (g): PBS (ml) volume = 1:9] on ice to prevent overheating. After homogenization, the samples were centrifuged at 5000 g for 5 min. MDA levels were measured using a Rat MDA ELISA kit (BT Lab, China), catalase levels were measured with a Rat CAT ELISA kit (BT Lab, China) and glutathione reductase levels with a Rat GR ELISA kit (BT Lab, China) using the colorimetric sandwich solid-phase enzyme-immunoassay method.
The sample size (n = 7 per group) was determined based on previous experimental cataract studies using the selenite-induced rat model, in which similar group sizes were sufficient to detect statistically significant differences, while minimizing animal use in accordance with ethical principles.2,4
SPSS version 25.0 software (IBM Corporation, Armonk, NY, USA) was used to analyze the data collected from this experimental study. The descriptive data of cataract grades and biochemical parameters measured in lens tissues were presented as mean and standard deviation (minimum and maximum) values. The distribution of the data was tested using the Kolmogorov-Smirnov test. Comparisons of variables between groups were performed using One-way ANOVA. The Tukey Kramer Multiple Comparison test was applied in post-hoc analyses. The Mann-Whitney U test was used for pairwise comparisons. A value of p < 0.05 was accepted as statistically significant.
The microscopic images obtained from the rat lenses are presented shown in Figure 1, and the clinical assessment results regarding cataract grading are summarized shown in Table 2. Cataract formation was observed in all rats in Group 1, confirming the success of the selenite-induced cataract model used in this experimental study.

Comparison of cataract grades among control, selenite-induced cataract, and treatment groups in the experimental model
| NUMBER of LENSES | CONTROL (Saline) | GROUP 1 (Selenite) | GROUP 2 (Selenite+Dmso) | GROUP 3 (Selenite + 10 mg/kg apocynin) | GROUP 4 (Selenite + 20 mg/kg apocynin) |
|---|---|---|---|---|---|
| 1 | 0 | 6 | 5 | 1 | 2 |
| 2 | 0 | 5 | 5 | 1 | 2 |
| 3 | 0 | 5 | 4 | 2 | 3 |
| 4 | 0 | 6 | 5 | 1 | 3 |
| 5 | 0 | 6 | 4 | 1 | 2 |
| 6 | 0 | 5 | 4 | 2 | 4 |
| 7 | 0 | 6 | 4 | 2 | 2 |
| 8 | 0 | 5 | 4 | 2 | 4 |
| 9 | 0 | 5 | 5 | 1 | 2 |
| 10 | 0 | 5 | 4 | 1 | 2 |
| X ± SD Min-Max | 0 ± 0 0 – 0 | 5.40 ± 0.52*** 5 – 6 | 4.40 ± 0.52*** 4 – 5 | 1.4 ± 0.52 ## , † 1 – 2 | 2.60 ± 0.84*, # 2 – 4 |
Slit-lamp biomicroscopic evaluation revealed that sodium selenite administration resulted in marked lens opacities in Groups 1 and 2, whereas apocynin treatment significantly reduced cataract severity (p < 0.001, p < 0.05). Specifically, cataract grades were significantly reduced in the apocynin-treated groups compared to the selenite-induced cataract group (Group 1), as also reflected in Table 2. The low-dose apocynin group (Group 3) showed numerically lower cataract grades compared to the high-dose group (Group 4); however, this difference did not reach statistical significance (p > 0.05). The detailed cataract grading of all the groups is shown in Table 2. Table 2 displays individual lens-level cataract grades to ensure transparency of morphological findings; however, all statistical comparisons were conducted at the animal level. Although cataract grades are ordinal by nature, mean values are presented for descriptive comparison across groups, while non-parametric statistical tests were applied for group comparisons, and individual lens-level data are also provided to allow direct interpretation of the ordinal distribution.
Boxplots were included to visually illustrate the distribution and variability of biochemical parameters across groups and to complement the tabulated summary statistics.
In the selenite (Group 1) and DMSO (Group 2) groups, MDA levels were significantly elevated compared to the control group (p < 0.001). MDA levels in the apocynin-treated groups (Groups 3 and 4) were evaluated relative to the selenite-induced cataract group (Group 1). No significant change in MDA levels was observed in Group 3 (p = 0.99), and Group 4 showed a significant increase in MDA levels (p = 0.04). This finding suggests a potential pro-oxidant effect at the higher dose of apocynin (shown in Figure 2 and Table 3).
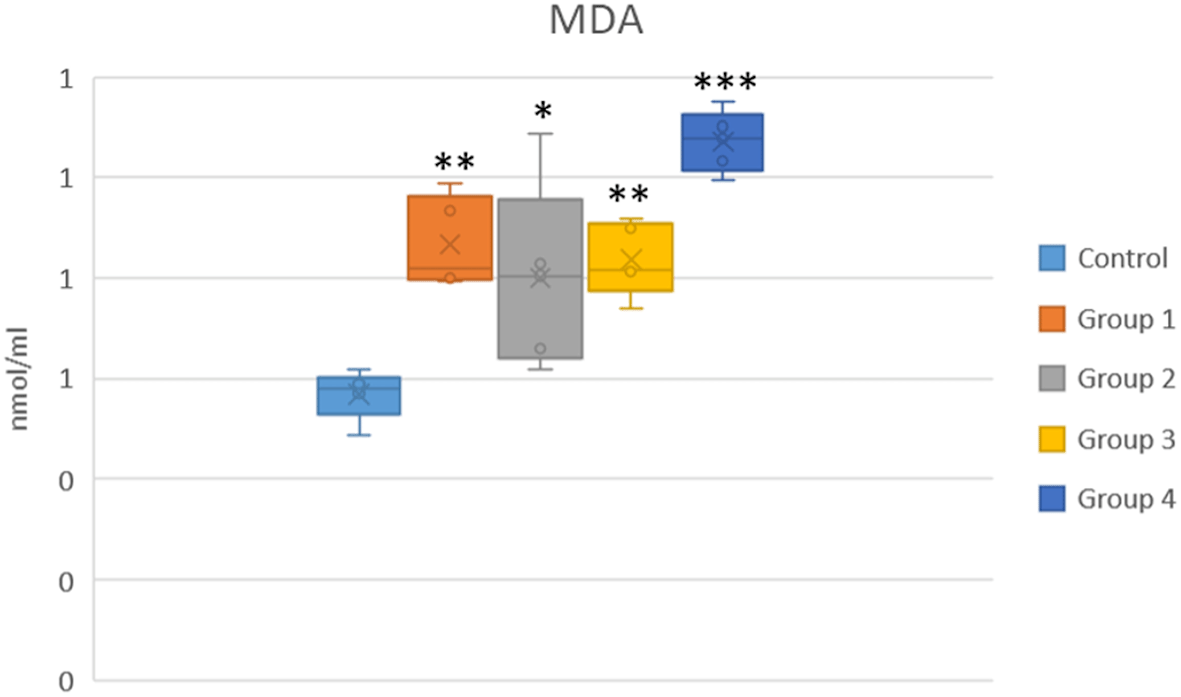
*p < 0.05, **p < 0.01 and ***p < 0.001 vs the control group.
Comparison of biochemical parameters among control, selenite-induced cataract, and treatment groups
All values are given as mean ± standard deviation, minimum-maximum values. MDA: Malondialdehyde, GR: Glutathione reductase
a-h: Different superscript letters indicate statistically significant differences between groups (p < 0.05, one-way ANOVA followed by Tukey post hoc test). In addition, comparisons relative to the control group and the selenite-induced cataract group (Group 1) were specifically evaluated to improve interpretability of treatment effects.
CAT activity was significantly reduced in the selenite and DMSO groups (Groups 1 and 2) (p < 0.01). In Group 3, CAT activity improved significantly compared to the selenite-induced cataract group (Group 1) (p = 0.003), and a moderate increase was observed in Group 4, not of statistical significance (p > 0.05) (Shown in Figure 3 and Table 3).
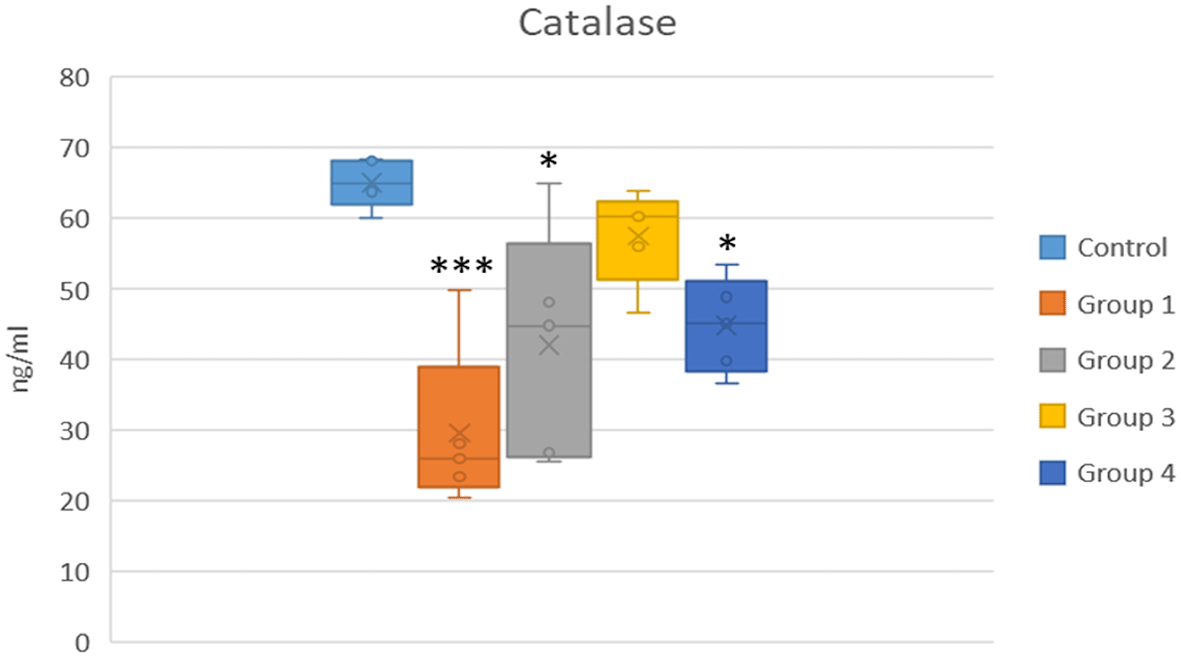
*p < 0.05 and ***p < 0.001 vs the control group.
GR levels were significantly decreased in the selenite group. A statistically significant increase in GR activity was observed in both Group 3 and Group 4 compared to the selenite-induced cataract group (Group 1) (p < 0.05) (shown in Figure 4 and Table 3).
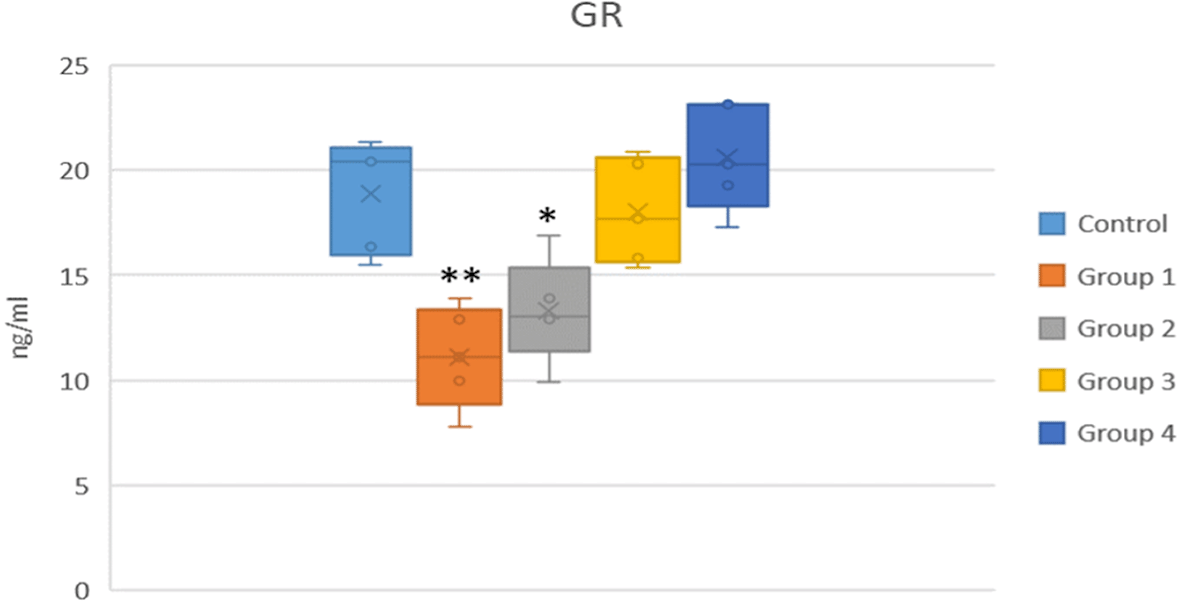
*p < 0.05 and **p < 0.01 vs the control group.
Following statistically significant overall group differences, post-hoc pairwise comparisons were performed as specified in the Methods section, and corresponding p-values are reported for the relevant group comparisons. In these analyses, apocynin-treated groups (Groups 3 and 4) demonstrated significantly more favorable oxidative stress profiles compared with the selenite-only (Group 1) and DMSO (Group 2) groups.
The results of this study demonstrated that apocynin, particularly at a low dose (10 mg/kg), exerted protective effects against selenite-induced cataractogenesis in rats. These effects were supported by reduced lens opacification and restoration of antioxidant enzyme activities. Low-dose apocynin significantly increased catalase (CAT) and glutathione reductase (GR) activities to levels comparable with those of the control group. Moreover, the oxidative stress marker malondialdehyde (MDA) did not increase and remained similar to control levels, indicating effective prevention of lipid peroxidation.
Oxidative stress plays a central role in cataract pathogenesis. The selenite-induced cataract model is widely used due to its rapid onset, simplicity, reproducibility, and strong resemblance to human age-related cataracts, particularly in terms of glutathione depletion, lipid peroxidation, and insoluble lens protein accumulation.2,4,13 In the current study, the model validity was confirmed by the development of dense nuclear opacities in Group 1, together with elevated MDA levels and reduced CAT and GR activities, indicating oxidative stress.
Despite inducing a partial enhancement in antioxidant enzyme activities (demonstrated by selective enhancement of GR activity, with minimal influence on CAT levels), high-dose apocynin (20 mg/kg) was found to increase MDA levels. This paradox suggests that the antioxidant benefits of apocynin may be counteracted by dose-dependent toxicity, resulting in a pro-oxidant response. This interpretation is consistent with literature indicating that excessive suppression of physiological ROS signaling can disrupt cellular homeostasis.21,22
Similar dual effects have been reported for other redox-active compounds such as resveratrol and quercetin, which show cytoprotective effects at low doses but may induce oxidative stress, DNA damage, and apoptosis at higher concentrations.23,24 Similar antioxidant protection has also been demonstrated for curcumin in rabbit cataract models, where curcumin reduced oxidative stress and delayed lens opacification.25 Although curcumin and apocynin act through different upstream pathways, both compounds converge on modulating oxidative damage, supporting the broader relevance of redox-targeted therapeutic strategies in cataract prevention. In addition, the potential contribution of DMSO as a vehicle to these effects should not be overlooked, as it has been shown to exert cytotoxic and pro-oxidant effects under certain conditions.26 The findings of this study highlight the importance of carefully optimizing both the dose and duration when evaluating antioxidant compounds in preclinical studies. Additionally highlight the need for careful dose optimization in human applications, as dose-dependent shifts between antioxidant and pro-oxidant activity may also occur in clinical settings.
The production of NADPH oxidase–driven reactive oxygen species (ROS) production plays a critical role in cataractogenesis by promoting protein aggregation, lipid peroxidation, and lens opacification.27 Given that oxidative stress is a key mechanism in selenite-induced cataract formation,4,9 targeting ROS-generating pathways has therapeutic relevance. Therefore, apocynin, which is an inhibitor of NADPH oxidase, was selected for its potential to suppress intracellular ROS production and enhance endogenous antioxidant defence systems, including catalase and glutathione reductase.28,29 Consistent with previous reports of oxidative stress-related disease models the current study results demonstrate that apocynin exerts protective effects on lens tissue, most likely through both antioxidant and anti-inflammatory mechanism.28–30 In line with earlier findings in studies of retinal injury and diabetic retinopathy,31,32 apocynin treatment was found to reduce cataract severity and modulate oxidative biomarkers in the current study model.
Importantly, this study is among the first to have compared different apocynin doses in a cataract model. The findings indicate that low-dose apocynin is more effective than a high dose in reducing oxidative damage, thereby emphasizing the importance of dose optimization to achieve therapeutic benefit while minimizing toxicity.
The biochemical analysis results further support these outcomes. Elevated malondialdehyde (MDA) levels in the selenite group confirmed increased lipid peroxidation, while apocynin treatment, particularly at low doses, reduced MDA levels. Similarly, catalase and glutathione reductase (GR) activities were preserved or enhanced with apocynin, indicating improved redox balance. As disruption of antioxidant enzymes is associated with lens protein denaturation and cataract progression,33,34 the ability of apocynin to stabilize these enzymes may underlie its protective mechanism.
The results of this study also support a strategic approach for the non-surgical prevention of cataract. Although cataract surgery remains the most effective treatment, access to surgical care is limited in many low- and middle-income countries due to resource constraints and economic burden.35,36 Pharmacological interventions based on natural compounds may offer a safe, cost-effective, and preventive alternative for early-stage cataracts. Given its pharmacokinetic profile and pleiotropic effects, apocynin emerges as a strong candidate for further preclinical and clinical investigations.
Unlike other antioxidant agents such as ascorbic acid, resveratrol, melatonin, ellagic acid, N-acetylcysteine, rosmarinic acid, and sildenafil, apocynin directly targets the NADPH oxidase complex, a key enzymatic source of ROS generation, thereby representing a mechanistically distinct approach to modulating oxidative stress at the level of ROS generation.14 This unique mechanism may confer broader and more sustained protective effects in lens tissue.
This study had several limitations that should be acknowledged. First, although the results suggest beneficial effects of apocynin, there was no assessment of the molecular mechanisms underlying its modulation of redox signaling in the lens. Key antioxidant pathways such as Nrf2/ARE, thioredoxin, and glutaredoxin systems, as well as apoptotic markers, were not evaluated.21 A second limitation was that the biochemical analysis was limited to ELISA-based measurements, and more detailed molecular techniques such as Western blotting or RT-PCR were not performed. In addition, apocynin was administered exclusively via the intraperitoneal route. Therefore, the comparative efficacy of alternative delivery methods, such as oral, topical, or intravitreal administration, remains unknown. The treatment duration was short-term, and the effects of long-term administration were not investigated. Finally, the absence of a positive control group using a well-characterized antioxidant agent limits the broader interpretability and comparability of the findings.
The results of this study demonstrated that apocynin, particularly at a low dose (10 mg/kg), exerts a significant protective effect against selenite-induced cataract formation in a rat model. This beneficial outcome is believed to be mediated through the enhancement of endogenous antioxidant defence mechanisms in the lens, as evidenced by increased catalase and glutathione reductase activities and the absence of elevated MDA levels. In contrast, high-dose apocynin (20 mg/kg) was associated with increased lipid peroxidation, suggesting a potential dose-dependent pro-oxidant effect. These findings emphasize the importance of dose optimization in the therapeutic application of antioxidant compounds. Although the results are promising, they remain preliminary and require further validation through comprehensive histopathological and molecular studies. If confirmed by future research, apocynin may represent a promising pharmacological candidate for preventing or delaying cataractogenesis through modulation of redox signaling in the lens.
The procedures applied to all animals involved in the study comply with the standards outlined in the Universal Declaration of Animal Rights, proclaimed at UNESCO in Paris in 1978. Prior to the study, ethical approval was obtained from the Animal Experiments Local Ethics Committee of Abant Izzet Baysal University (Date: 9th June 2021, Approval Number: 2021/19).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)