Keywords
Pomegranate peel, Leuconostoc, Pediococcus, Ellagic acid, Synergistic fermentation, Antioxidant, probiotic.
This article is included in the Fallujah Multidisciplinary Science and Innovation gateway.
Pomegranate peel, Leuconostoc, Pediococcus, Ellagic acid, Synergistic fermentation, Antioxidant, probiotic.
Fermentation is a traditional bioprocess involving microbial transformation of substrates and has emerged as a promising tool not only for preservation but also for enhancing the nutritional and functional qualities of food materials.
As functional foods, plant foods provide not only the essential nutrients needed to sustain life but also low-energy foods for health promotion and disease prevention (Hadi et al., 2022).
Consumers’ expectations for minimal processing, nutritional value of food, and healthy food are increasing daily (Alabdali et al., 2020). Therefore, as an alternative to traditional pasteurization processes, nonthermal treatment options have recently been evaluated (Alabdali et al., 2020; Hadi et al., 2020).
In this respect, using two or more species of microorganisms is called synergistic fermentation, and it is a system that has recently attracted attention; when compared to single-species fermentation, its biotransformation results are far better (Motghare et al., 2025; Cao et al., 2025).
Pomegranate peel is rich in ellagitannins, natural bioactive phytochemicals. Ellagic acid (EA) is also known as a very good antioxidant and has been shown to possess anti-inflammatory and anti-proliferative activities, as well as cardioprotective effects. These complex polyphenols can be hydrolyzed to produce EA, as demonstrated in previous studies (Jahn et al., 2023; Soares et al., 2025). However, the complexity of continuation to this end and the existing low solubility hamper the natural bioavailability of ellagic acid in pomegranate peel (Ma et al., 2025). It was found that production of Ellagic acid derivative may be obstructed, even under lactic fermentation (Cai et al., 2024; Han and Du, 2023). In contrast, free forms of ellagitannins are formed from complex compounds and new traits arise; thus, lactic fermented pomegranate peel can improve its health and nutritional benefits (Hadi et al., 2021). Lactic acid bacteria (LAB) are very helpful guests and can assist the host with fertilizer, adjusting digestive system function, reducing cholesterol in serum level, developing substances that will prevent cancer cells causing tumors, etc.--so one can give examples such as yeast flocculation or food preservation with sugar (Kaleda et al., 2025). They have been used as natural laboratory test materials in pickles, salted vegetables/fruit products for many years, and these groups make good experimental models with their different regulatory and physiological properties as well as coordinating sensory data (Behera et al., 2020; Huang et al., 2024). These experiment results showed that L. plantarum JGS49 fermented asparagus juice had significantly high antioxidant activity and special flavors, such as increase of the content in flavonoid and total phenol (Hou et al., 2025). LAB fermented food containing live cells has probiotic effects in the human body, so it is suitable for LAB coupled with their bioactive compounds as food supplements (Diez-Ozaeta and Astiazaran, 2022; Saleem et al., 2024).
Pediococcus pentosaceus has characteristic complementary enzyme systems to Leuconostoc mesenteroides. These two species produce large amounts of β-glucosidase and tannase-like enzymes. As a result, when mixed culture fermentation is performed using these strains together in combination, they interact synergistically during their co-operation, such that the rate at which substrates are used accelerates, their efficiency of fermentation increases, and phenolic compound release is further heightened. In addition, these two species are particularly valued for their probiotic properties and can serve to enhance the amount and variety of gut microbiome, an important biological activity that hosts do not possess (Saud et al., 2024). Although polyphenol biotransformation is gaining acceptance, few research have looked at how synergistic fermentation can increase ellagic acid concentration and probiotic enrichment at the same time. Industrial wastes such as pomegranate peel could gain significant value from such a dual-purpose strategy, which is consistent with the circular bio-economy and sustainability principles (Sevindik et al., 2022; Shivani and Sathiavelu, 2024).
The effect of special treatments, such as Ultraviolet and Ultrasound treatments, on the physicochemical and microbial characteristics of pomegranate juice was studied and found to be significant (Alabdali et al., 2020).
The objective of this study was to evaluate the synergistic fermentation of pomegranate peel extract (PPE) using Leuconostoc mesenteroides and Pediococcus pentosaceus for dual purposes: enhancing ellagic acid content through microbial bioconversion and enriching the extract with viable probiotic biomass. It has possible uses in the creation of sustainable functional meals or nutraceutical goods. This study also examined how co-culturing these lactic acid bacteria enhances the nutritional and functional value of PPE.
Fresh pomegranate (Punica granatum L.) peels had been collected from Al-Karkh vegetable market/Baghdad/Iraq. The species was identified taxonomically based on macroscopic morphological traits using standard references on the botany of the respective regions. No voucher specimens were deposited as the material constituted commercially available food-grade produce. Leuconostoc mesenteroides (LM) ATCC 8293 and Pediococcus pentosaceus (PP) ATCC 25745 strains were purchased from the microbial culture collection and each strain has its ATCC no. Gallic acid, methanol, sodium carbonate, and Folin-Ciocalteu reagent were obtained from Sigma–Aldrich.
After being cleaned with distilled water, pomegranate peels were dried for 24 h at 50°C in a hot air oven. A laboratory grinder was used to grind the dried peel into a fine powder. The extract was prepared by mixing 10 g peel powder with 100 mL distilled water (1:10 w/v). The mixture was heated at 80°C for 1 h with continuous stirring, then filtered through Whatman No.1 filter paper. The filtrate was autoclaved at 120°C for 15 min and allowed to cool to room temperature before use.
Two lactic acid bacteria, Leuconostoc mesenteroides ATCC 8293, and Pediococcus pentosaceus ATTC 25745 was used to ferment PPE. These strains were maintained at -80.0°C in 20% (v/v) glycerol stock until use. To prepare LAB for fermentation, strains were first cultivated on De Man, Rogosa, and Sharpe (MRS) agar, incubated at 30°C for 24 h, and then cultured in MRS broth. After incubation at 30°C for 24 h, supernatants of bacteria were harvested by centrifugation at 4000 rpm for 10 min, followed by a double washing procedure in saline buffer (0.85%), and resuspended in saline to achieve a final optical density (Pramudito et al., 2024).
Four fermentation treatments were prepared, the first was the control (uninoculated) one which represents PPE without microbial inoculation, the second was LM which represents PPE incubated with Leuconostoc mesenteroides (5% v/v), the third was PP which represents PEE incubated with Pediococcus pentosaceus (5% v/v) and the fourth was LM+PP (co-culture) which represents PPE inoculated with both strains (2.5% v/v each). Samples were incubated at 30°C for 72 h under anaerobic conditions and collected at 0, 24, 48, and 72 h for analysis.
Ellagic acid and TPC were measured using a High-Performance Liquid Chromatography (HPLC) apparatus equipped with a UV detector (Agilent 1200 series). The stationary phase was a reversed-phase C18 column (250 mm × 4.6 mm) and the mobile phase was diluted with methanol. with 0.1% formic acid at a flow rate of 1.0 mL/min and the injection volume of all the samples was 20 μL. The samples were filtered before injection, the concentration was expressed in ppm, and the detection of phenolic compounds was performed at a wavelength of 280 nm. The total phenolic content was calculated as the sum of the qualified individual phenolic compounds and expressed as milligram gallic acid equivalents per L of extract. All samples were run in triplicate.
Calibration curves were constructed using pure ellagic acid and gallic acid standards. For ellagic acid quantification, a series of standard solutions (10–200 ppm) was prepared and injected under identical chromatographic conditions. The peak areas were plotted against known concentrations to generate linear regression equations (y = ax + b) with correlation coefficients (R2 ≥ 0.99). Gallic acid was used as the reference compound for TPC determination, and the results were expressed as milligrams of gallic acid equivalents (GAE) per liter of extract.
An HPLC device equipped with a Xtimate Sugar-H column (Welch Materials, China) was used to measure the levels of lactic acid, glucose, and fructose. Twenty microliters was The injection volume, (Sulfuric acid at 0.003 mol/L) served as the mobile phase, the temperature of the column was 60°C, and the flow rate was 0.6 ml/min. A refractive index detector (RID) was used to identify sugars, and a UV detector was used to track lactic acid at 210 nm (Han and Du 2023).
The total antioxidant capacity of the pomegranate peel extract (PPE), before and after fermentation, was measured by the Ferric Reducing Antioxidant Power (FRAP) assay, according to Sachett et al. However, the material ideas in this are) of (Sachett et al., 2021; Vijayalakshmi and Ruckmani, 2016) with minor changes made.
2.7.1 Chemicals and reagents
All the chemicals which were used were of analytical grade. The following chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA): 2,4,6-tripyridyl-s-triazine (TPTZ) (93285-1G), Iron (II) sulfate heptahydrate (FeSO4·7H2O) (F7002-250G), Iron (III) chloride hexahydrate (FeCl3·6H2O) (100 g, F2877), and 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox) (238813-1G), glacial acetic acid (A6283-100ML), and hydrochloric acid (HCl) (H1758-100ML).
2.7.2 Preparation of FRAP reagent
FRAP reagent was freshly prepared at a rate of 10:1:1 (v/v/v) for 300mM acetate buffer (pH 3.6), 10mM TPTZ solution in 40mM HCl, and 20mM FeCl3·6H2O, respectively. Prior to use, the reagent was equilibrated at 37∘C.
2.7.3 Assay procedure
In short, in a test tube 3.0mL of the FRAP reagent were combined with 200μL of the PPE sample. The blend was vortexed and incubated at 37∘C for 30 minutes away from light. Following incubation, the absorbance was read at 593nm using a Shimadzu UV-1800 spectrophotometer. Reproducibility was ensured by completing all measurements in triplicate.
2.7.4 Calibration and quantification
Standard calibration curves for FeSO4 ·7H2 O (100–1000μM) and Trolox (0.1–1.0mM) were constructed. Standards were processed in the same way as the samples. The antioxidant activity was determined based on the interpolation of the absorbance values of the samples compared to the linear regression equations of the standard curves. Correlation coefficients (R2≥0·99) confirmed the precision of the assay.
The viable cell counts of Pediococcus pentosaceus and Leuconostoc mesenteroides during the fermentation of PPE extract were determined using the conventional plate count method. In this method, 1 ml of the fermented sample was removed at time intervals of 0, 24, 48, and 72 h and diluted ten times in sterile 0.85% (w/v) saline solution. Aliquots (200 μL) were spread and plated in duplicate on MRS agar plates using appropriate dilutions. The plates were incubated for 48 h at 30°C under anaerobic conditions in an anaerobic jar with a gaspack. Colonies were counted following incubation, and the results were expressed as log colony-forming units per milliliter (log CFU/mL).
The Statistical Package for Social Sciences (SPSS, 2019) program was used to detect the effects of different factors on the study parameters. The least significant difference (LSD) was used to compare significant means in this study.
The surface features and cellular configurations of Leuconostoc mesenteroides and Pediococcus pentosaceus were analyzed using scanning electron microscopy (SEM). The morphology of Leuconostoc mesenteroides is depicted in Figure 1A which appeared as short rods and oval with smooth and intact cells, indicating healthy cell membranes with no structural damage. It is also clear that, they are frequently found as single, in pairs, or in short chains. Meanwhile, Figure 1B represents the morphology of Pediococcus pentosaceus, which appears as small, round cocci with smooth and uniform surfaces, well-defined boundaries, and no signs of cellular deformation. These results are consistent with the characteristics of these bacteria.

TIFF File.
The morphology of Leuconostoc mesenteroides is depicted in Figure 1A which appeared as short rods and oval with smooth and intact cells, indicating healthy cell membranes with no structural damage. It is also clear that, they are frequently found as single, in pairs, or in short chains. Meanwhile, Figure 1B represents the morphology of Pediococcus pentosaceus, which appears as small, round cocci with smooth and uniform surfaces, well-defined boundaries, and no signs of cellular deformation. These results are consistent with the characteristics of these bacteria.
The pH change during various fermentation durations is depicted in Figure 2. It is evident that all inoculated samples showed a notable drop in pH during fermentation, indicating active microbial metabolism and the generation of organic acids such as lactic acid. The results showed that a notable drop in pH during the fermentation process for all samples was significant (P < 0.05). For the entire fermentation period, the pH value of the control sample was not significantly altered compared with the initiation value. However, this difference was not statistically significant.
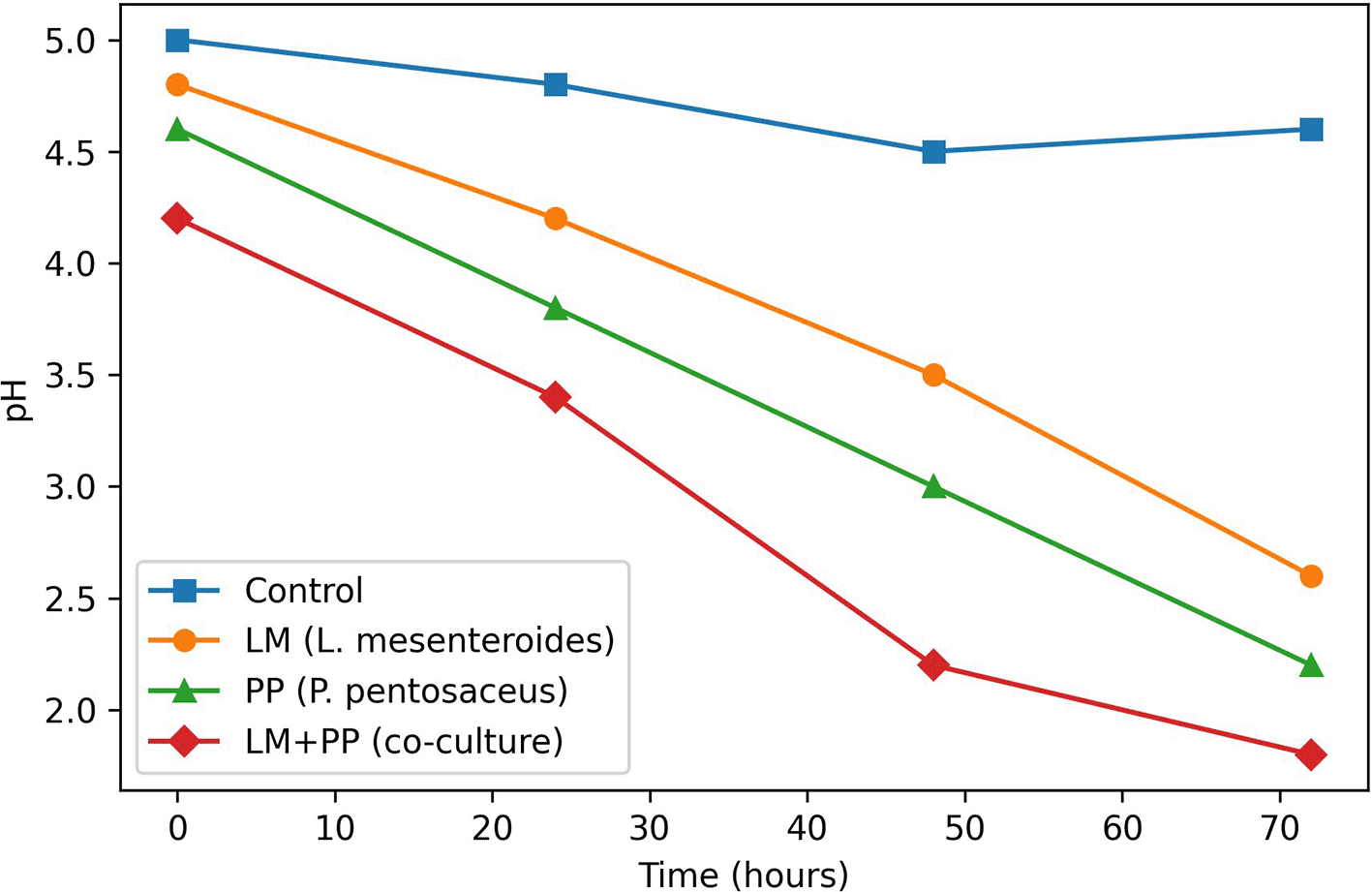
JPG File.
The Figure 2 depicts pH change during 72-h of incubation for control (no inoculum), Leuconostoc mesenteroides (LM), Pediococcus pentosaceus (PP) and the synergistic co-culture (LM+PP). All inoculated treatments experienced a marked (P<0.05) decrease in pH, suggesting active microbial metabolism and organic acid production. The combined synergistic co-culture (LM+PP) performed the best with the fastest acidification and a final pH (approximately 2.0) after 72 h that was significantly lower than the final pH of the single-strain fermentations. Data points are the average of triplicate readings (n = 3).
Similar acidification tendencies were observed in the LM and PP single-strain fermentations, with the fastest pH decrease in the first 30 h; therefore, it was highly significant (P ≤ 0.01). Additionally, it is evident that the co-culture of LM and PP produced the lowest pH (approximately 2), indicating improved fermentation efficiency and metabolic synergy in the formation of acid.
Statistical analysis using two-way ANOVA demonstrated significant effects of treatment (F = 0.683, P < 0.0001), time (F = 0.815, P < 0.0001), and their interaction (F = 1.037, P < 0.0001) on the pH values. The co-culture (LM+PP) achieved the greatest acidification, reducing the pH to ~2.0, which was significantly lower than that of the LM or PP monocultures. These findings confirm that microbial fermentation drives acidification, with co-culture strategies enhance efficiency through metabolic complementarity.
The TPC generated from the fermentation of pomegranate peel extract (PPE) in all treatments is shown in Figure 3. It is obvious that, over the period of 72 hours, the control sample (non-inoculated PPE) demonstrated very little change in TPC with no significant (P > 0.05) change, suggesting that natural enzymatic or chemical changes were restrained in the absence of microbial action.
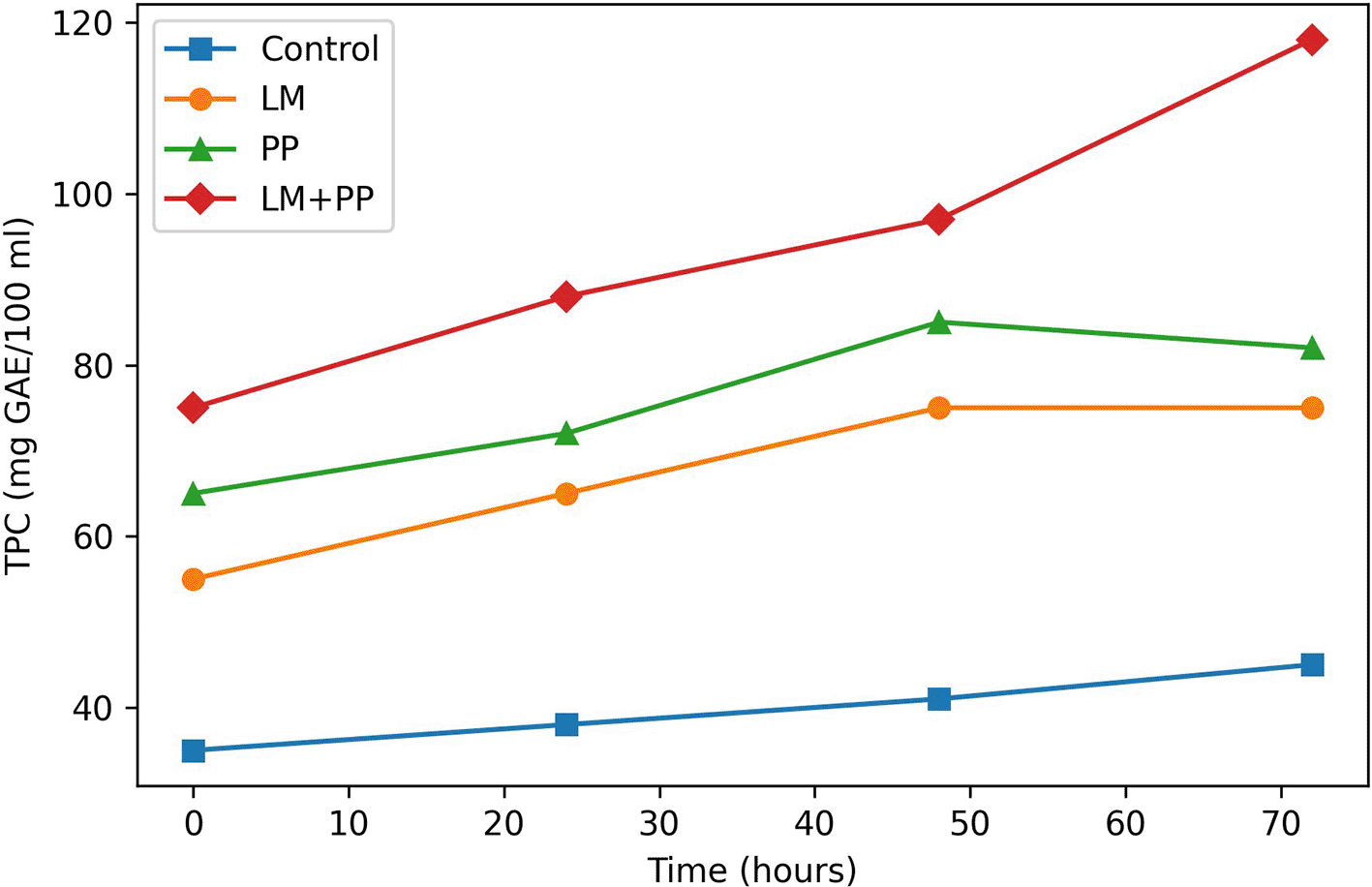
JPG File.
Figure 3 Changes in TPC throughout a 72-h fermentation period (expressed as mg gallic acid equivalents/100 ml -mg GAE/100 ml). A comparison of the noninoculated control, Leuconostoc mesenteroides (LM), Pediococcus pentosaceus (PP), and their synergistic co-culture (LM+PP). Compared to control, a significant increase (P<0.05) in TPC was observed across all fermented treatments. The phenolic enrichment was significantly higher in the synergistic co-culture (LM+PP), indicating its efficiency in the biotransformation and release of bound phenolic compounds from pomegranate peel matrix. Data are mean (n=3) of triplicate measurements.
However, all inoculation samples showed a significant (P < 0.05) increase in total phenolic content, with the co-culture (LM + PP) showing the most pronounced increase (highly significant, P ≤ 0.01). This revealed a synergistic metabolic interaction between Leuconostoc mesenteroides and Pediococcus pentosaceus in digesting complex polyphenols and generating helpful compounds such as ellagic acid.
TPC content increased gradually during LM administration, reaching its peak value at 58 h, which is in line with other studies showing that phenolic-transforming enzymes are present in L. mesenteroides. Additionally, PP treatment increased the phenolic content, but the increase was somewhat smaller than that in LM; this could be due to variations in fermentation dynamics and enzyme profiles. The largest increase in TPC release, however, was observed in the (LM + PP) co-culture, which peaked at 72 h. This suggests that both bacteria may offer complementary enzymatic functions such as phenolic acid decarboxylase.
Two-way ANOVA revealed significant effects of treatment (F = 15.88, P < 0.0001), time (F = 19.63, P < 0.0001), and their interaction (F = 27.04, P < 0.0001) on total phenolic content (TPC). Among all treatments, the co-culture (LM + PP) exhibited the greatest increase in TPC, reaching its maximum at 72 h, followed by the PP and LM monocultures, whereas the control showed no significant change over time. These results indicate that microbial fermentation markedly enhances phenolic release, with co-culture fermentation offering the strongest effect, likely due to the metabolic complementarity between the strains.
The antioxidant capacity of pomegranate peel extract (PPE) was evaluated using the ferric reducing antioxidant power (FRAP) assay before and after fermentation with lactic acid bacteria and results obtained are illustrated at Table 1. The table shows that the fermentation process significantly (P < 0.05) increased PPE's reducing power of PPE. Since the baseline antioxidant activity of the unfermented control extract was primarily derived from native phenolic constituents, such as flavonoids and ellagitannins, it displayed comparatively low FRAP values. Following fermentation, a noticeable increase in the absorbance was observed. This result was significant (P < 0.05), which suggests a better antioxidant potential due to the increased ferric ion reduction ability. The repeatability of the trend was confirmed by the fact that this improvement was true among the biological duplicates.
| Sample | Absorbance (AU) |
|---|---|
| Control (PPE) | 0.373 ±0.02 b |
| LM | 0.522 ±0.04 ab |
| PP | 0.641 ±0.04 a |
| (LM+PP) | 0.719 ±0.05 a |
| L.S.D. (P-value) | 0.207 ** (0.0054) |
The observed increase in the antioxidant capacity after fermentation can be attributed to several factors. First, free ellagic acid and smaller phenolic derivatives with higher reducing abilities were probably released during the microbial enzymatic hydrolysis of ellagitannins and other bound polyphenols. Second, new bioactive metabolites with antioxidant potential, such as phenolic acids and short-chain organic acids, may have been produced as a result of the metabolic activity of lactic acid bacteria. These procedures enhance the bioavailability and reactivity of antioxidants, in addition to increasing the total phenolic content. The magnitude of the improvement follows the trend.
Statistical analysis using LSD revealed significant differences among treatments (LSD = 0.207, P = 0.0054). The co-culture (LM + PP) exhibited the highest antioxidant activity (0.719 ± 0.05 AU), followed by PP (0.641 ± 0.04 AU), LM (0.522 ± 0.04 AU), and control (0.373 ± 0.02 AU). These results confirm that fermentation with lactic acid bacteria significantly enhances the reducing power of PPE, with the co-culture strategy providing the greatest improvement owing to metabolic complementarity and increased release of bioactive phenolic compounds.
3.5.1 Glucose consumption
The glucose concentration was measured over 72 h in the four samples (control, LM, PP, and (LM+PP)). The data collected are presented in Figure 4A from which it is clear that Leuconostoc mesenteroides (LM), Pediococcus pentosaceus (PP), and their co-culture (LM + PP) exhibited distinct glucose utilization characteristics during the fermentation of pomegranate peel extract (PPE). The most extensive glucose consumption was observed in LM + PP, followed by PP culture. (P ≤ 0.01), whereas the LM monoculture displayed the slowest rate. Since it steadily dropped from 180 mg/L at zero time to 58 mg/L at 72 h, or a 68% reduction, it is also evident that the (LM+PP) co-culture exhibited the highest glucose consumption rate and it was highly significant (P ≤ 0.01). Two-way ANOVA showed a significant effect of time-dependent coefficient variation on glucose consumption (F = 32.59, P < 0.0001) with a highly significant interaction (F = 49.66, P < 0.0001).
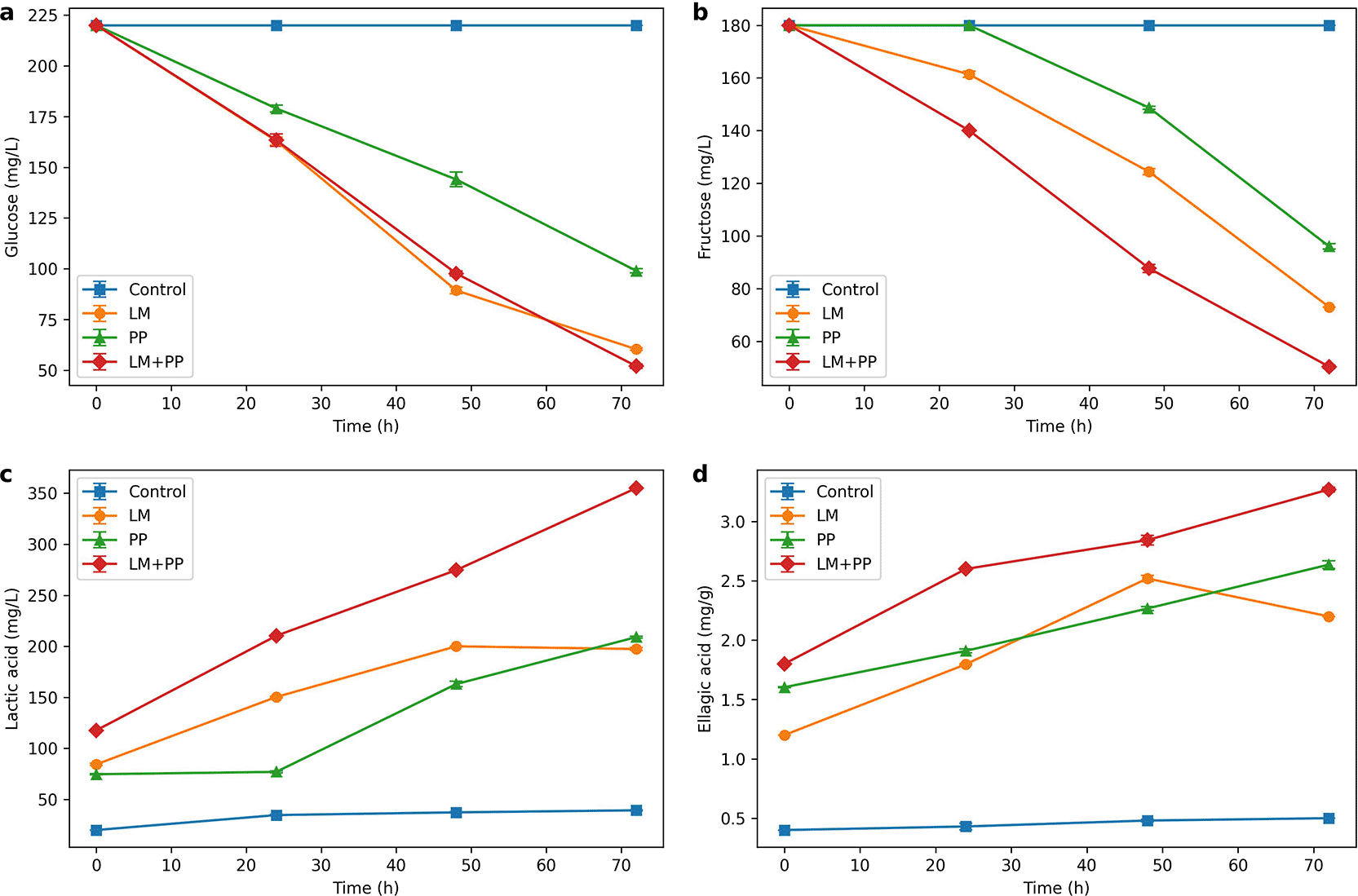
JPG File.
Figure shows the performance of the non-inoculated control, Leuconostoc mesenteroides (LM) Pediococcus pentosaceus (PP) and their synergistic co-culture (LM+PP). (A) Glucose usage (mg/L): All fermented samples show rapid glucose depletion with the highest glucose consumption in the LM+PP co-culture. (B) Fructose uptake (mg/L): Fructose was used as a second carbon source with the co-culture significantly decreased compared to both strains separately. (C) Lactic acid production (mg/L): Continuous production of lactic acid was continuously observed, with the complete synergistic interaction of LM and PP yielding the highest-final concentration (340 mg/L). (D) Ellagic acid concentration (mg/g): As a result of the biotransformation of complex ellagitannins, free ellagic acid concentrations were increased relative to the monoculture, reaching a maximum for the co-culture at 3.4 mg/g.
The increased glucose consumption in the co-culture was most likely due to mutualistic interactions and complementary metabolic traits between the two species. Additionally, LM reduced glucose less efficiently than PP, achieving 76 mg/L reduction at 72 h (58% reduction). Compared to LM, the combined treatment (LM+PP) exhibited a moderate decrease in glucose levels, dropping from 180 mg/L to 102 mg/L at 72 h (43% reduction). PPE contains numerous polyphenolic compounds with antibacterial properties. Because PP has a higher phenolic tolerance than LM, it is likely able to initiate rapid glucose absorption, which lowers the pH and alters the redox balance, thereby reducing phenolic stress in LM. This interaction appears to shorten the lag phase for LM in the co-culture by enabling more efficient sequential and parallel substrate consumption. These findings are consistent with earlier research that demonstrated that mixed lactic acid bacterial cultures outperformed monocultures in the fermentation of phenolic-rich plant substrates due to cross-protection and metabolic complementarity. According to the bioprocess theory, the co-culture method may be able to more efficiently bio-convert PPE into products with added value, such as lactic acid and bioactive metabolites, while maximizing substrate depletion and reducing residual sugar content.
3.5.2 Fructose consumption
All cultures quickly consumed glucose, but fructose was consumed more slowly than glucose. This is consistent with the preferred glucose metabolism shown by lactic acid bacteria (LAB) as a result of carbon catabolite suppression. Figure 4B makes it evident that at the start of fermentation (24 hours), the remaining fructose levels were relatively high across treatments, with (LM+PP) retaining roughly 80% of the initial fructose, LM 88%, and PP 92%. As fermentation progressed, differences in fructose utilization became increasingly apparent. By 48 h, co-cultivation reduced residual fructose to 55%, LM to 65%, and PP to 78%. At the end stage (72 h), LM and PP retained 50% and 65% of the initial fructose, respectively, while the co-culture retained only 30%. This is a distinctive feature of LAB with a hetero-fermentative metabolism. The more efficient fructose consumption in LM than in PP in monoculture settings may be explained by the most efficient L. mesenteroides metabolism (P < 0.05). The bacterium LM may use the phosphor-ketolase pathway to utilize fructose as a carbon source. Even in the presence of glucose, LM can use fructose, albeit more slowly owing to its metabolic adaptability. Conversely, P. pentosaceus is primarily responsible for the production of low-sugar functional beverages, and its improved ability to reduce the glucose and fructose contents of PPE from an application perspective results in increased bioconversion efficiency. The complementary carbohydrate metabolism of the LM+PP combination not only enhances the removal of total sugar but also promotes the synthesis of mannitol and other beneficial metabolites, potentially increasing the health benefits of the final product.
A two-way ANOVA showed that time and treatment variation had a significant effect on fructose uptake (F = 26.07, P < 0.0001), with a highly significant interaction effect (F = 37.57, P < 0.0001). Meanwhile, co-fermentation using two bacterial strains (LM + PP) demonstrated the highest fructose uptake efficiency, retaining 30% after 72 h, compared to 50% in LM and 65% in PP.
These results indicate metabolic complementarity between the two strains used in the fermentation process, L. mesenteroides and P. pentosaceus, which not only leads to the biotransformation of sugars but also to an increase in functional metabolites, such as mannitol, thus improving the nutritional and health value of the fermented product.
3.5.3 Lactic acid production
The concentration of lactic acid increased over time in all fermentation modes while the pomegranate peel extract was fermented; the results are shown in Figure 4C. Lactic acid production in the LM monoculture began rapidly within the first 12 h and peaked after 48 h. This initial acceleration is caused by the hetero-fermentative metabolism of L. mesenteroides, which converts available sugars into lactic acid and byproducts such as acetic acid, ethanol, and CO2 using the pentose–phosphate and phosphor-ketolase pathways. However, the PP monoculture accumulated acid steadily and linearly throughout the fermentation process, despite exhibiting a slower start of acid generation in the first 12 h. Because of P. pentosaceus, the final concentration of lactic acid (214 mg/L) was greater than that of the LM monoculture, which was 198 mg/L. This could be because the homofermentative lactic acid bacteria used the Embden–Meyerhof pathway to transfer the majority of the carbon from the substrate directly into lactic acid. Conversely, the non-inoculated control sample accumulated very little lactic acid (22 mg/L), suggesting that spontaneous lactic acid fermentation was not present in the experimental setup. On the other hand, the (LM + PP) co-culture had a discernible synergistic effect, since LM successfully lowered the pH and created an environment that was favorable for PP metabolism and growth by initiating rapid production during the early fermentation phase (0–12 h). PP's homofermentative activity of PP subsequently sustained and enhanced the production of lactic acid throughout the mid- to late-stages of fermentation.
The different treatments with the pomegranate peel extract, along with the bacterial strains used, had a significant and statistically significant effect on lactic acid production (LSD = 19.22, P = 0.0001).
The effect of time on lactic acid production was also statistically significant (LSD = 19.22, P = 0.0001), indicating that the amount of lactic acid produced depended on both the bacterial strain used and the fermentation time. In addition, the combined effect of time and treatment was statistically significant (LSD = 19.22, P = 0.0001), with lactic acid production gradually increasing from 0 to 1 h.
The study showed that using two types of bacteria in the pomegranate peel fermentation process significantly increased lactic acid production, followed by the sample using PP bacteria and then the sample using LM bacteria. These were the percentages of variation in this cooperative interaction that generated the highest final lactic acid concentration (340 mg/L), which increased over any monoculture, by optimizing conversion efficiency and reducing by-product production. These findings indicate that LM is better at early acidification owing to its rapid sugar metabolism, whereas PP is better at producing high-purity, extended lactic acid. Combining the advantages of both, co-culture raises the output of total lactic acid, suppresses undesirable microbial contaminants, and lowers pH more rapidly.
Figure 4D exhibits the changes in ellagic acid concentration during the fermentation of pomegranate peel extract (PPE) under different treatments at different intervals. From this figure it is clear that, throughout duration of experiment, the levels of ellagic acid in the control (unfermented PPE) stayed constant (0.5 mg/g), with only slight variations due to analytical variance. As there was no enzymatic or microbial hydrolysis of ellagitannins in the untreated extract, no discernible release of bound EA was observed. The EA concentration in the LM monoculture increased moderately, particularly after 24–48 h of fermentation. Owing to its β-glucosidase activity, L. mesenteroides can partially hydrolyze ellagitannin-bound moieties into free EA. However, the conversion efficiency was limited because of its hetero-fermentative metabolism and comparatively small repertoire of phenolic-degrading enzymes, which led to a net EA concentration of 2.2 mg/g. Compared to LM, the PP monoculture displayed a more noticeable increase in the EA concentration (2.8 mg/g). P. pentosaceus is known to exhibit increased β-glucosidase and tannase activities, which facilitate the more efficient cleavage of ester bonds and glycosidic linkages in ellagitannins. A synergistic effect between the two species was demonstrated by the highest elevation in EA content in the LM + PP co-culture. While PP maintained a high and sustained release rate of EA through its more powerful hydrolytic enzyme system, LM helped to reduce the pH early and partially break down tannins, which made the substrate more accessible. This combined mechanism led to a 3.4 mg/g ellagic acid concentration. These findings show that by enzymatically hydrolyzing ellagitannins, LAB fermentation of PPE increases EA availability. Although EA release was enhanced by both LM and PP monocultures, co-culture enhanced this effect by combining PP's potent tannase and β-glucosidase activities of PP with early acidification and cell wall-disrupting action. In addition to releasing more EA, this dual action probably increases its stability by lowering oxidative degradation in acidic environments (Sepúlveda et al., 2020).
Statistical analysis using the least difference test confirmed that the treatment factor had a significant effect (LSD = 0.776, P = 0.0001), indicating that different treatment conditions resulted in varying ellagic acid levels and concentrations. Time also had a significant effect (LSD = 0.776, P = 0.0001), demonstrating a consistent increase in the ellagic acid concentration with prolonged fermentation.
Furthermore, the variation in the samples over time had a statistically significant effect (LSD = 1.024, P = 0.0001), indicating that increasing the fermentation time leads to an increase in ellagic acid. The combined LM+PP treatment showed greater and more time-related improvement in ellagic acid concentration than single samples or treatments fermented using only one type of bacteria in the ellagic acid ratio.
Table 2 shows the viable cell counts for the three samples, from which it is clear that for LM, there was a significant (P < 0.05) rapid increase from 5.0 to 8.2 log CFU/ml at 48 h, followed by a slight decline to 7.8 at 72 hr. In contrast, PP showed a moderate rise from 5.0 to 7.5 log CFU/ml at 48 h, maintained around 7.3 at 72 hr. A sharp increase was observed with LM+PP co-culture (9.5 log CFU/ml at 48 h), with a slight drop to 9.0 at 72 h. As a result, the co-culture (LM + PP) supports optimal microbial proliferation, outperforming single cultures. This supports the use of synergistic LAB combinations for enhanced probiotic enrichment, which was significant (P < 0.05).
This study demonstrated that the fermentation of pomegranate peel extract (PPE) by Leuconostoc mesenteroides (LM), Pediococcus pentosaceus (PP), and their co-culture (LM + PP) enhanced the bioactive potential and fermentation efficiency compared to the uninoculated control. All inoculated fermentations exhibited rapid acidification, with the LM + PP co-culture achieving the lowest pH (~2.0). The total phenolic content significantly increased in all treatments, with the co-culture producing the most pronounced enhancement. Sugar utilization patterns revealed distinct strain-specific differences: LM consumed glucose more slowly (43% reduction), PP showed moderate efficiency (58% reduction), and LM+PP co-culture achieved the greatest glucose depletion (68%). Similarly, fructose was more effectively consumed in the co-culture, retaining only 30% at 72 h compared with 50% (LM) and 65% (PP). In terms of ellagic acid (EA release), LM fermentation moderately increased EA (2.2 mg/g), PP achieved a higher yield (2.8 mg/g), and the LM+PP co-culture reached the maximum concentration (3.4 mg/g), reflecting synergistic enzymatic action and complementary metabolic interactions. Overall, the co-culture of LM and PP was superior to monocultures by combining acidification, sugar depletion, and enzymatic hydrolysis, thereby enhancing phenolic release and bioactive metabolite production. These findings highlight the potential of LM+PP co-fermentation as a promising bioprocessing approach to convert PPE into functional, health-promoting products.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)