Keywords
biosynthetic gene, chloroplast genome, Dipterocarpus, pelahlar, phylogeny
biosynthetic gene, chloroplast genome, Dipterocarpus, pelahlar, phylogeny
This section should include why the data were gathered or produced. Conservation for critically endangered endemic trees is essential to prevent immediate extinction, as these species often have extremely small, restricted, and vulnerable populations. Due to their limited geographic range and specialized, often damaged habitats, they are far more susceptible to extinction than species with broader distributions (Işik 2011). However, they are vital to ecosystem health, supporting biodiversity, regulating the climate, and purifying the air. Protecting them maintains ecological balance and preserves unique genetic resources for resilience. One of the key endemic trees in Indonesia is Dipterocarpus littoralis (Blume) Kurz., or locally known as the pelahlar or plahlar tree (Yulita and Partomihardjo 2011; Dwiyanti et al. 2014; Primajati et al. 2017; Widiastuti et al. 2022), which belongs to the Dipterocarpaceae family and is endemic to Nusakambangan Island, located in Cilacap Regency, Central Java Province, Indonesia, specifically in the West Nusakambangan Nature Reserve (Dwiyanti et al. 2014; Robiansyah and Davy 2015).
The tree grows in lowland tropical forest, reaching 50 cm in diameter and 150 cm in height, sometimes with high buttresses (Hamidi and Robiansyah 2018). The bark tends to peel, especially in old trees, pale grey to grey, resinous (Hamidi and Robiansyah 2018). Furthermore, the species is heavily harvested for local timber needs and is sometimes tapped for resin (dammar) used for fire or to produce glue for boats (Hamidi and Robiansyah 2018). Since 1998, the species has been classified as critically endangered (CR) by the International Union for Conservation of Nature (IUCN) Red List of Threatened Species (Hamidi and Robiansyah 2018) due to illegal logging and is increasingly threatened due to the expansion of the invasive Arenga obtusifolia, which inhibits the regeneration of D. littoralis seedlings (Robiansyah and Davy 2015).
Dwiyanti et al. (2014) reported the presence of 18 adult trees on the island, while Robiansyah and Davy (2015) found 11 adult trees. Furthermore, the genetic diversity of this species, assessed using microsatellite markers, was low, with an expected heterozygosity of 0.476. This suggests that reductions in population size have been occurring for an extended period (Dwiyanti et al. 2014). Therefore, urgent efforts are needed to prevent this species' extinction. Several studies have been conducted to support conservation strategies for D. littoralis, including assessments of population status and habitat preferences (Robiansyah and Davy 2015; Primajati et al. 2017), and of genetic diversity (Yulita and Partomihardjo 2011; Dwiyanti et al. 2014). However, investigations into the chloroplast genome (cp genome), molecular phylogeny, and biosynthetic gene cluster (BGC) have not yet been addressed. Studying the chloroplast genome and molecular phylogeny is crucial, as these provide essential tools for biodiversity conservation and taxonomic resolution, helping to prioritize and implement protective measures. For instance, high-throughput sequencing of the entire chloroplast genome can facilitate the identification of unique lineages, map population structures, and develop DNA barcodes to accurately identify endangered species (Yin et al. 2025; Ma et al. 2025). Additionally, biosynthetic gene clusters (BGCs) are conserved genomic regions that encode specialized metabolites, which play a critical role in species adaptation, defense, and ecological interactions (Cawood and Ton 2025). Therefore, this study aims to characterize the chloroplast genome (cp genome) sequences, construct a phylogenetic tree, and identify the biosynthetic gene cluster (BGC) of D. littoralis.
Fresh leaf samples were collected from a single Dipterocarpus littoralis sapling ( Figure 1A) planted in 2022 at the area of Faculty of Forestry and Environment, Institut Pertanian Bogor (IPB University), located in Bogor Regency, West Java Province, Indonesia (6°33’21,775” S, 106°43’48,45781” E). The sapling was originally sourced from the West Nusakambangan Nature Reserve in Cilacap Regency, Central Java Province, Indonesia. Dipterocarpus littoralis exhibits unique morphological characteristics that distinguish it from other species in its genus. Notably, it features red velvet and hairy stipules ( Figure 1B), along with elliptical to ovate leaves of 16-25 cm x 10-18 cm (Newman et al. 1998) ( Figure 1C). The leaves are spirally arranged and clustered around the apices of the twigs (Hamidi and Robiansyah 2018). A herbarium specimen was collected and deposited in the Forest Genetics and Molecular Forestry Laboratory, the Department of Silviculture, Faculty of Forestry and Environment, at IPB University in Bogor Regency, West Java Province, Indonesia, under voucher number LGH.0369. The species name was verified by Fifi Gus Dwiyanti through comparison with the Dipterocarpus littoralis specimen number PL.VI-C.75 available at the Herbarium Center of the Center for Sustainable Forest Development, Ministry of Forestry of the Republic of Indonesia, Bogor City, West Java Province, Indonesia. This verification was also supported by prior research by Dwiyanti et al. (2014).

Pictures were taken by the author Fifi Gus Dwiyanti on 2026-02-12.
The laboratory work, which includes DNA extraction and sequencing, was conducted at the Laboratory of Forest Genetics and Molecular Forestry within the Department of Silviculture at the Faculty of Forestry and Environment, Institut Pertanian Bogor (IPB University) in Bogor Regency, West Java Province, Indonesia. A total of 100 mg of fresh leaf material was used for genomic DNA extraction and isolation using the modified Cetyltrimethylammonium Bromide (CTAB) protocol established by Doyle and Doyle (1990). The leaf sample was ground in a mortar and pestle with 1000 μL of 10% CTAB buffer (Cat. No. MB101-100G, HiMedia Laboratories). To eliminate proteins and RNA, 40 μL of 26% Polyvinylpyrrolidone (Cat. No. MB102-100G, HiMedia Laboratories), 5 μL of mercaptoethanol (Cat. No. 8.05740.0250, Merck), and 200 μL of NaCl (Cat. No.1.06404.1000, Merck) were added. The sample was then homogenized using a vortex MX-5 (Biologix) and subsequently incubated in a water bath (VWR Scientific) at 65°C for 30 minutes. The partitioning of lipids and cellular debris into the organic phase was completed by adding 500 μL of chloroform (Cat. No. 1.02445.2500, Merck)-isoamyl alcohol (Cat. No. 1.00979.1000, Merck) with a ratio of 24:1 and 10 μL of phenol, followed by centrifugation at 13,000 rpm for 3 minutes using Centrifuge (MPW Med. Instruments) to obtain purified DNA. The purified DNA was precipitated by adding an isopropanol (Cat. No. 1.09634.2500, Merck) solution at a 1:1 volume ratio to the supernatant, along with NaCl at a 1:4 volume ratio. The samples were then stored at -20°C for approximately 2 hours to allow DNA to precipitate. Subsequently, the DNA was washed with 500 μL of 70% ethanol (Cat. No. 1.00983.2500, Merck) and dried in a desiccator (Normax). Finally, 50 μL of TE buffer (Cat. No. 93283-500ml, Merck) was added to the genomic DNA before the samples were stored at -20°C.
The quality of the genomic DNA was assessed via agarose gel electrophoresis utilizing a UV Transilluminator TFX-20 LM (Vilber Lourmat). Quantitative measurement of genomic DNA was performed using a Qubit 1.0 fluorometer (Invitrogen-Thermo Fisher Scientific) with the Qubit dsDNA BR assay kit (Cat. No. Q32850, Thermo Fisher Scientific). The high-molecular-weight genomic DNA extracted from D. littoralis was subsequently employed for library preparation to facilitate long-read sequencing. DNA libraries were prepared according to the Nanopore protocol using a ligation sequencing kit (Cat. No. SQK-LSK110, Oxford Nanopore Technologies), version GDE_9108_v110_revN_10Nov2020 (Oxford Nanopore Technologies). Long-read sequencing was then performed on a MinION Mk1B R9.4.1 flow cell (Cat. No. FLO-MIN106D, Oxford Nanopore Technologies), connected to MinKNOW software (Oxford Nanopore Technologies) to control the device, monitor flow cells, and perform real-time basecalling. All data analysis was performed at the Forest Genetics and Molecular Forestry Laboratory within the Department of Silviculture, Faculty of Forestry and Environment, IPB University, Bogor Regency, West Java Province, Indonesia.
The raw Fastq data were analyzed using NanoPlot v1.46.1 (De Coster et al. 2018) to calculate and visualize read distributions. De novo assembly of the organelle genome was performed using Flye v2.9.6 (Kolmogorov et al. 2019) to construct the plastome. The chloroplast genome was reconstructed from an assembly using GetOrganelle (Jin et al. 2020). The resulting chloroplast genomes were then annotated using GeSeq (https://chlorobox.mpimp-golm.mpg.de/geseq.html) (Tillich et al. 2017). The fully annotated circular genome was visualized using OrganellarGenomeDRAW (OGDRAW) v1.3.1 accessible through the MPI-MP Chlorobox platform (Greiner et al. 2019).
A total of 18 chloroplast genomes of taxa closely related to Dipterocarpus littoralis (from the Dipterocarpaceae family) were downloaded from GenBank (the National Center for Biotechnology Information/NCBI) and aligned with the obtained plastomes. A complete list of the accessions used is given in Table 1. Gyrinops verstegii (AP018453.1) was used as the outgroup. The alignment was performed using MAFFT v7.526 (Katoh and Standley 2013) using default parameters. A maximum-likelihood (ML) phylogenetic tree was inferred using the IQ-TREE2 (Trifinopoulos et al., 2016) with 1,000 bootstrap replicates. The phylogenetic tree was visualized using iTOL (Letunic and Bork 2024).
| No. | Taxon | GenBank accession number | Current taxon (Ashton and Heckenhauer, 2022)** |
|---|---|---|---|
| 1. | NC_058774.1 | Vatica rassak | - |
| 2. | NC_071231.1 | Vatica bantamensis | - |
| 3. | NC_058773.1 | Vatica xishuangbannaens | - |
| 4. | NC_054172.1 | Vatica odorata | - |
| 5. | NC_051531.1 | Vatica guangxiensis | - |
| 6. | NC_041485.1 | Vatica mangachapoi | - |
| 7. | NC_081465.1 | Dipterocarpus littoralis | - |
| 8. | NC_058777.1 | Dipterocarpus alatus | - |
| 9. | NC_067812.1 | Dipterocarpus retusus | - |
| 10. | NC_046842.1 | Dipterocarpus turbinatus | - |
| 11. | NC_065503.1 | Dipterocarpus hasseltii | - |
| 12. | NC_041191.1 | Neobalanocarpus heimii | - |
| 13. | NC_044642.1 | Hopea hainanensis | - |
| 14. | NC_052744.1 | Hopea reticulata | - |
| 15. | NC_053766.1 | Hopea chinensis | - |
| 16. | NC_057187.1 | Dryobalanops aromatica | - |
| 17. | NC_046579.1 | Parashorea chinensis | - |
| 18. | NC_064148.1 | Shorea macrophylla | Rubroshorea macrophylla |
| 19. | AP018453.1 | Gyrinops versteegii * | - |
* The plastome of Gyrinops versteegii, not a member of the Dipterocarpaceae family, was used as an outgroup.
** Ashton PS, Heckenhauer J. 2022. Tribe Shoreae (Dipterocarpaceae subfamily Dipterocarpoideae) Finally Dissected. Kew Bulletin. 77:885-903. doi:10.1007/S12225-022-10057-W.
All DNA sequence datasets from the assembly were annotated using BRAKER2 (without RNA sequencing) (Brůna et al., 2021) and then assigned to putative proteins and protein regions using GeneMark-ES. The predicted proteins are aligned with the NCBI protein database using DIAMOND to identify the target proteins. These proteins are subsequently aligned to the seed region using Spaln (a space-efficient spliced alignment tool). Through these steps, genome sequences, protein predictions, and the corresponding sequence regions were obtained. Afterward, the exon boundaries were predicted by identifying introns, start codons, and stop codons. The sequences are classified as high or low confidence based on the predictions' confidence levels. High confidence sequences can be processed further with AUGUST2, while those with low confidence are reviewed and reordered to enhance gene predictions using the EP+ gene tag. Upon completing all phases, the results from BRAKER2 yield DNA sequences with predictable genes, which are then clustered using the PlantiSMASH platform (plantismash.secondarymetabolites.org) (Kautsar et al., 2017).
Long-read sequencing of Dipterocarpus littoralis using the MinION ONT has produced a dataset of 40,219,030 reads, resulting in a total of 48.4 gigabase pairs (Gbp) of raw data. The mean read length was 1,204.4 base pairs (bp), and the mean raw data read quality score was 11.8. Following the filtering process, all reads have met the quality assessment criteria and retain the same quality score. The complete chloroplast genome of D. littoralis has been effectively assembled from long-read sequencing data, enabling the resolution of structural regions within the genome. The chloroplast genome exhibits a typical quadripartite structure ( Figure 2a) with a total length of 153,775 bp. The genome comprises a Small Single-copy Region (SSC: 20,232 bp) and a Large Single-copy Region (LSC: 85,079 bp), which are separated by two inverted repeat regions: Inverted Repeat A (IRA: 24,232 bp) and Inverted Repeat B (IRB: 24,232 bp) ( Figure 2a). The GC content of the D. littoralis sequence is 37 %. This finding exceeds the chloroplast genome size of Dipterocarpus turbinatus, as reported by Ci et al. (2019) using short-read sequencing on the Illumina HiSeq 2000 platform, which reported a size of 152,279 bp. However, it is lower than that of Dipterocarpus retusus, which was generated via short-read sequencing on the Illumina NovaSeq platform by Tao et al. (2024), at 154,303 bp. The findings support the conclusion that the plastome size in the genus Dipterocarpus is relatively conserved (typically in the 152–154 kb range).
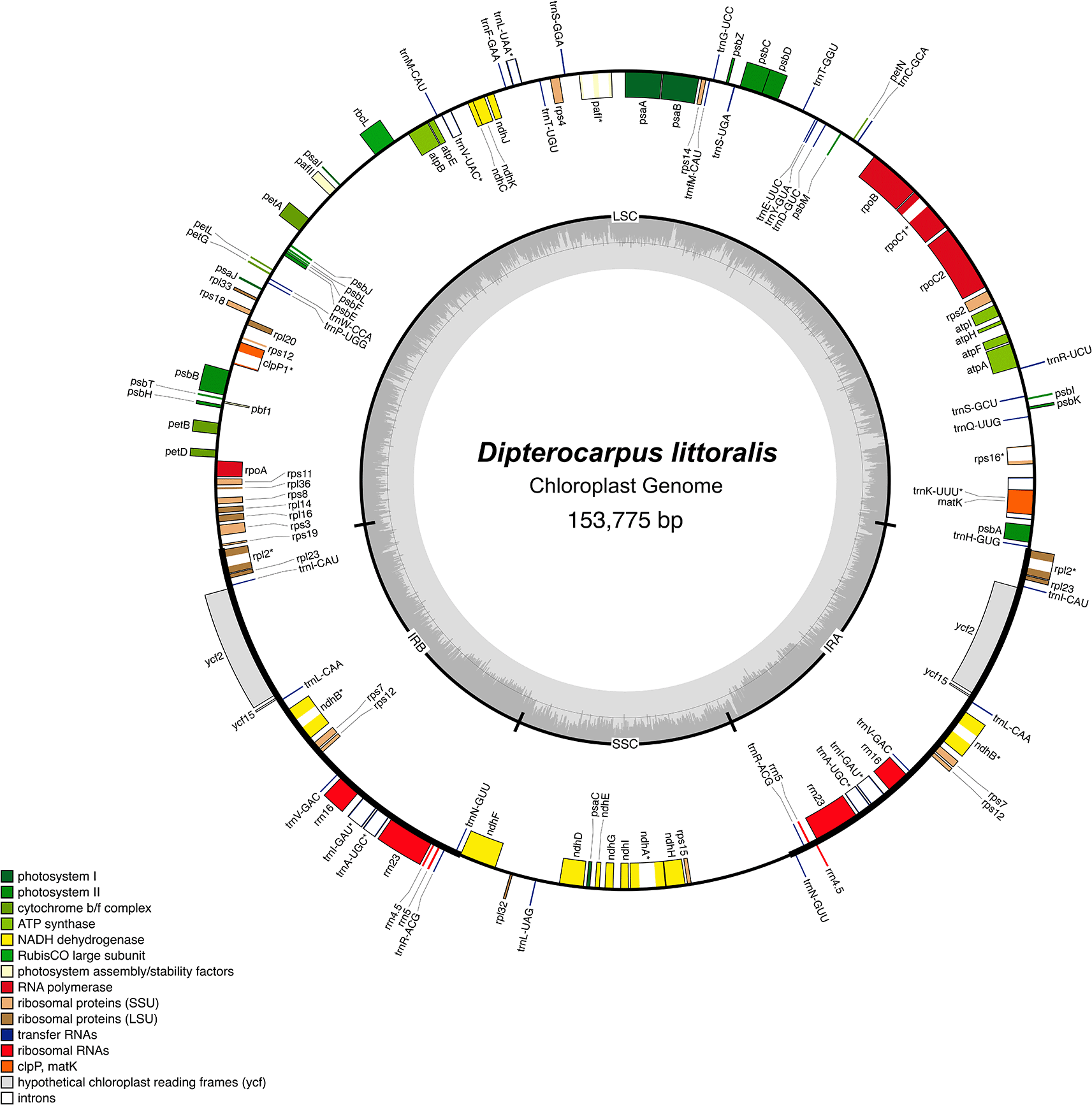
The D. littoralis chloroplast genome (cp genome) contains 111 genes, including 78 protein-coding genes, 29 transfer RNA (tRNA) genes, and 4 ribosomal RNA (rRNA) genes ( Table 2). These genes are categorized into four functional groups: self-replicating genes, photosynthetic genes, genes with other functions, and genes of unknown function ( Table 2). This composition is similar to that of D. retusus and D. turbinatus, whose chloroplast genomes encode 128 genes, including 84 protein-coding genes, 36 tRNA genes, and 8 rRNA genes (Ci et al., 2019; Tao et al., 2024).
| Functional category | Gene group | Gene name |
|---|---|---|
| Self-replication | Ribosomal RNA | rrn16d, rrn23d, rrn4.5d, rrn5d |
| Transfer RNA | trnA-UGCd*, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-UCC, trnH-GUG, trnI-CAUd, trnI-GAUd*, trnK-UUU*, trnL-CAAd, trnL-UAA*, trnL-UAG, trnM-CAU, trnN-GUUd, trnP-UGG, trnQ-UUG, trnR-ACGd, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GACd, trnV-UAC*, trnW-CCA, trnY-GUA | |
| Large subunit ribosomal proteins (LSU) | rpl2d*, rpl14, rpl16d*, rpl20, rpl23d, rpl32, rpl33, rpl36 | |
| Small subunit ribosomal proteins (SSU) | rps2, rps3, rps4, rps7d, rps8, rps11, rps12d*, rps14, rps15, rps16d*, rps18, rps19 | |
| DNA-dependent RNA polymerase | rpoA, rpoB, rpoC1*, rpoC2 | |
| Photosynthesis | Subunits of ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI |
| Subunits of NADH-dehydrogenase | ndhA*, ndhBd*, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Subunits of photosystem I | psaA, psaB, psaC, psaI, psaJ, pbf1 | |
| Subunits of photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, psbZ | |
| Subunits of cytochrome b/f complex | petA, petB, petD, petG, petL, petN | |
| Subunit rubisco | rbcL | |
| Other function | Subunit of acetyl-CoA-carboxylase | accD |
| C-type cytochrome synthesis gene | ccsA | |
| Protease | clpP1*, pafI**, pafII | |
| Maturase | matK | |
| Envelope membrane protein | cemA | |
| Translation initiation factor | infA | |
| Unknown function | Conserved open reading frames | ycf1d, ycf2d |
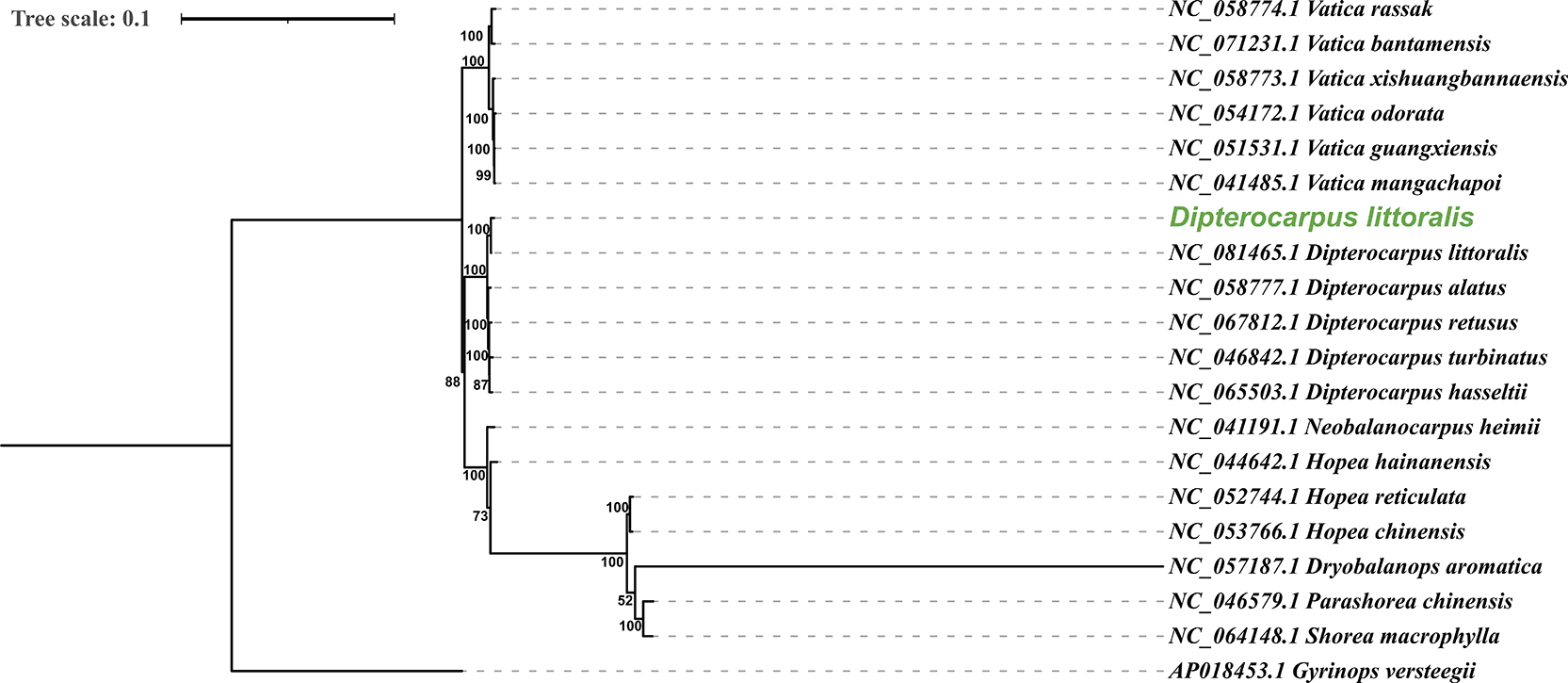
The phylogenetic tree of D. littoralis based on the complete chloroplast genome showed that the studied D. littoralis was in the same clade as D. littoralis (NC_081465.1) with a bootstrap value of 100% ( Figure 3). Additionally, the studied D. littoralis formed a monophyletic group with other species within the same genus, specifically D. alatus (NC_058777.1), D. retusus (NC_067812.1), D. turbinatus (NC_046842), and D. hasseltii (NC_065503.1), also showing a bootstrap value of 100% ( Figure 3). This pattern aligns with the findings of Tao et al. (2024), which reported a monophyletic clade for the phylogenetic tree of Dipterocarpus, indicating a specialized evolutionary lineage. The present study contributes valuable insights for future studies on the molecular identification and evolutionary dynamics of D. littoralis and related species, enhancing understanding of the phylogeny of Dipterocarpus and the family Dipterocarpaceae.
In this study, 8,399 contigs encoding biosynthetic gene clusters (BGCs) of D. littoralis and 77 genes coding for BGCs were identified, comprising 44 terpene clusters, 6 alkaloid clusters, 1 lignan cluster, 4 saccharide clusters, 1 Saccharide-Polyketide cluster, 1 saccharide-terpene cluster, and 20 putative clusters ( Table 3), indicating a high potential for secondary metabolism. At 44 BGC terpene, D. littoralis was found to contain gene-producing protein domain including Terpene synthase (involved in sesquiterpene and monoterpene pathways) and SQHop cyclase (involved in triterpenoid pathways) (Yang et al., 2018). Six D. littoralis alkaloid BGCs contain gene-producing protein domains, namely, Copper amine oxidase (involved in the biosynthesis of pyrrolizidine/PA alkaloids), Pictet-Spengler enzymes, and Strictosidin synthase-like (Bet v1), both of which are involved in the biosynthesis of monoterpene indole alkaloids (MIA) (Maresh et al., 2008). This information enables an overview of D. littoralis by identifying the Pictet-Spengler enzyme and the Strictosidine synthase-like (Bet v1) gene.
A total of four D. littoralis saccharide BGCs contain gene-producing protein domains, namely glycosyltransferases (GTs), which are carbohydrate-active enzymes (CAZy) and are the primary catalysts in the biosynthesis and modification of plant cell walls (Hao et al., 2012). Although D. littoralis has only one cluster of BGC lignans, this cluster acts as a catalyst for bimolecular coupling reactions that result in regiochemical and stereochemical control (Pickel and Schaller, 2013). Hybrid BGCs that identify saccharides joined to other clusters were also identified in this study, namely, one saccharide-polyketide BGC cluster and one saccharide-terpene BGC cluster ( Table 3). The sub-clusters in the saccharide-polyketide BGCs were Stilbene synthase domains and UDPGT2, and the sub-clusters in the saccharide-terpene BGCs were Terpene synthase and UDPGT2. Both BGCs identified the unique characteristics of the secondary metabolites in D. littoralis. In addition to these clusters, D. littoralis also has 20 putative clusters with sub-clusters, such as CoA-ligase, Fatty acid desaturase, Dioxygenase, Cellulose synthase-like, cytochrome 450, Dioxygenase, Methyltransferase, COesterase, BAHD acyltransferase, Methyltransferase, Polyprenyl synthetase, Oxidoreductase, Lipoxygenase, Aminotransferase, Squalene epoxidase, Aminotransferase, and Prenyltransferase.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)