Keywords
depression; social isolation; neuroplasticity; exercise; fluoxetine; corticosterone;; prefrontal cortex; male Wistar rats
This article is included in the Research Synergy Foundation gateway.
depression; social isolation; neuroplasticity; exercise; fluoxetine; corticosterone;; prefrontal cortex; male Wistar rats
Depression is a clinical progressive psychiatric disorder that significantly impairs the quality of life and functional capacity.1 Recently, the incidence of depressive disorders is increasing, resulting in enormous economic and social burdens.2,3 Ongoing research on biomarkers associated with major depression, as well as the development of new classes of antidepressants, has shown that only 60%–70% of patients with depression respond positively to antidepressant therapy, leaving a considerable number vulnerable to treatment-resistant depression.4 Moreover, the integration of psychopharmacology and non-pharmacology interventions focused on improving the quality of life, restoring the function of life, and minimizing disability.
Interactions mediate the cause of depression between genetic and epigenetic factors and the nature and nurture processes. Consequently, the inherent heterogeneity of this disease complicates its treatment through a singular modality.5 The monoamine hypothesis, which has dominated the field for over half a century, has provided a foundation for pharmacological treatment and the development of new antidepressant modalities; however, its limitations are underscored by the substantial proportion of patients who fail to achieve remission with monoaminergic antidepressants, thereby prolonging the recovery process on patient.6
One of the major challenges in understanding the pathophysiological mechanisms of depression is that no single hypothesis can fully explain all its signs and symptoms. Clinical and preclinical investigations have associated depression with the classical monoamine hypothesis,7–10 abnormalities in the second messenger system,9,11 neuroinflammation hypothesis,3,8,12,13 neurotrophic propositions,5,14 neuroplasticity and neurogenesis hypothesis,5,8,15 and mitochondrial dysfunction.16–20 This therapeutic gap has prompted a paradigm shift toward the neuroplasticity hypothesis and metabolism of energy through mitochondrial dysfunction, which suggests that depression results from impaired structural, autophagy, and functional plasticity in brain regions, particularly the prefrontal cortex (PFC) and hippocampus.16,17,20–23
The urgent need to identify novel pathways involved in depression arises from the limitations of existing antidepressant therapies, which primarily target monoamine neurotransmitter systems and the inflammation pathway; they often fail to achieve remission in a substantial proportion of patients.24,25 In light of the evidence suggesting a potential association between aberrant neuroplasticity and the underlying mechanism of MDD, further research aimed at disentangling the precise mechanisms by which neuroplasticity contributes to the pathological processes that lead to depression may provide insight for the identification of novel therapeutic targets.5,26 Converging evidence from clinical and preclinical studies has identified glutamatergic dysfunction as a fundamental neurobiological substrate of depression. Post-mortem analyses of individuals with depression have demonstrated altered glutamate transporter expression and a disrupted excitatory-inhibitory balance in the cortex.27–29
The vesicular glutamate transporter 1 (V-GluT1) facilitates the encapsulation of glutamate into synaptic vesicles before its release and serves as a molecular determinant of the efficacy of glutamatergic neurotransmission.30 Research In learned helplessness Sprague- Dawley rats model revealed a significant reduction in V-GluT1 expression within the hippocampus and cerebral cortex. This reduction corresponds to the anhedonic and helpless behavioral phenotype.31 Postsynaptic glutamate signaling via metabotropic glutamate receptors plays an equally critical role in neuroplasticity and mood regulation—metabotropic glutamate receptor 1 (mGluR1).31–33
The neurotrophic environment of the brain constitutes a significant axis of depression susceptibility. Among the neurotrophic factors associated with the pathophysiology of depression, FGF-2 has emerged as a particularly noteworthy molecule.5 Specifically, FGF-2 expression was downregulated in these cortical regions in patients with depression, and this reduction was attenuated by antidepressant treatment.34,35
A less explored yet increasingly recognized molecular system in the neurobiology of depression is the PA pathway. Multiple evidence support the involvement of the PA system in the pathophysiology of depression.36–43 Post-mortem analyses of individuals who completed suicide and had a history of major depression demonstrated significantly elevated levels of putrescine and spermidine in the cerebral cortex. This phenomenon is accompanied by a widespread downregulation of SAT1, the primary enzyme responsible for their catabolism.36,39,44
This study represents the first comprehensive analysis of the gene expression of sSrm, V-GluT1, mGluR1, and FGF-2, which are indicative of presynaptic glutamate dynamics, postsynaptic receptor-mediated plasticity, neurotrophic support, and polyamine-dependent cellular homeostasis, respectively, within the prefrontal cortex of a social isolation and chronic unpredictable stress Wistar rats depression model. This investigation offers a multidimensional molecular signature of neuroplasticity disruption and therapeutic restoration.
Research has established the positive effects of physical therapy, fluoxetine monotherapy, and combined exercise and pharmacological interventions on depression.45–47 Nevertheless, fluoxetine monotherapy has several limitations. One significant drawback is the delayed initiation of therapeutic effects, with approximately 30% of patients not reaching the remission phase.48 Additionally, the stigmatization associated with antidepressant use can lead to non-adherence and suboptimal treatment outcomes.49–51
Furthermore, a sedentary lifestyle is commonly observed in individuals with depressive disorders, contributing to a reduction in neurotrophin levels.46,52,53 Physical exercise has been identified as a significant non-pharmacological intervention for depression. A seminal Cochrane review (2024) and an extensive umbrella review of 27 systematic reviews, encompassing 190 experiments, have demonstrated that exercise yields a moderate effect size in alleviating depressive symptoms (Standarized Mean Difference = −0.67), comparable to both psychotherapy and antidepressant medications.54,55 Although extensive research has been conducted on the therapeutic efficacy of antidepressants and physical exercise for depression, their comparative and combined effects, as well as the underlying mechanisms, remain notably inconsistent. Theoretically, the combination of these two modalities should yield additive or synergistic benefits; however, empirical evidence does not consistently support this assumption.
This study aimed to assess the comparative therapeutic effects of fluoxetine therapy, physical activity, and their combination in the management of depression. Additionally, this research examined the molecular mechanisms by which combination therapies influence these pathways, with the objective of advancing evidence-based treatment strategies. This study investigated the relationship between antidepressants and exercise on neuroplasticity and stress levels in the brain following depression, as well as the interplay between HPA axis normalization and glutamatergic/neurotrophic restoration. Exploring this process may provide insights into the prevention and treatment of loneliness-induced depression resulting from social isolation and chronic unpredictable stress.
The study was performed in strict accordance with the guidelines specified in the Guide for the Care and Use of Laboratory Animals, with approval from the Ethics Committee of the Medical Faculty at Universitas Padjadjaran. The methodologies applied in this research align with the ethical standards of the relevant national and institutional bodies that oversee animal experimentation. The methodologies have received approval from the Research Ethics Committee at Universitas Padjadjaran (Number 1344/UN6.KEP/EC/2023).
In this study, 25 12-week-old male Wistar rats with an average weight of 200 g were housed in either experimental or control cages. The rats were procured from Biofarma Company, located in Bandung, Indonesia, and housed at the Animal Research Innovation center, Universitas Padjadjaran, Bandung, Indonesia.56,57 Acclimatization after transfer was 2 weeks. All animals were maintained under a 12-h light-dark cycle (lights activated at 19:00) under controlled environmental conditions (22–24°C). Food and water were provided ad libitum throughout the two-week acclimatization period.58
Experimental Procedure
Social Isolation Procedure and Unpredictable Chronic Mild Stress.56,57,59–64
All the Wistar rats that passed the acclimatization phase were randomly divided into groups. The Wistar rats were divided into 5 groups: a normal housing group (NCG), a positive control group (social isolation and chronic unpredictable mild stress (PCG), a positive control group with antidepressant fluoxetine treatment (PCG + FLX), a depression model group with moderate intensity continuous training (PCG + MICT), and a positive control group with combination antidepressant and moderate intensity continuous training (PCG + FLX + MICT).
The impact of antidepressants, continuous moderate-intensity training, and the combination of antidepressants with continuous moderate-intensity training on depressive-like behavior in Wistar rats was evaluated through weekly weight measurements, serum corticosterone levels, and the gene expression of Srm, V-GluT1, m-GluR1, and FGF-2 within the brain throughout the duration of the 8-week study. The body weight of the rats was recorded weekly using a digital scale (Jiuyue) to monitor changes throughout the intervention period. Figure 1 presents overview of experimental design and allocated treatment to each group. At the conclusion of the experiment, all groups of rats were euthanized using 5% isoflurane (Mersi) inhalation methods. Subsequently, the rats were dissected from the cranial region to facilitate the extraction of the brain tissue. The organs were placed in tubes according to their respective groups and stored at −80°C. (Thermo Scientific Forma 900 Series −86°C Upright Ultra-Low Temperature Freezers).
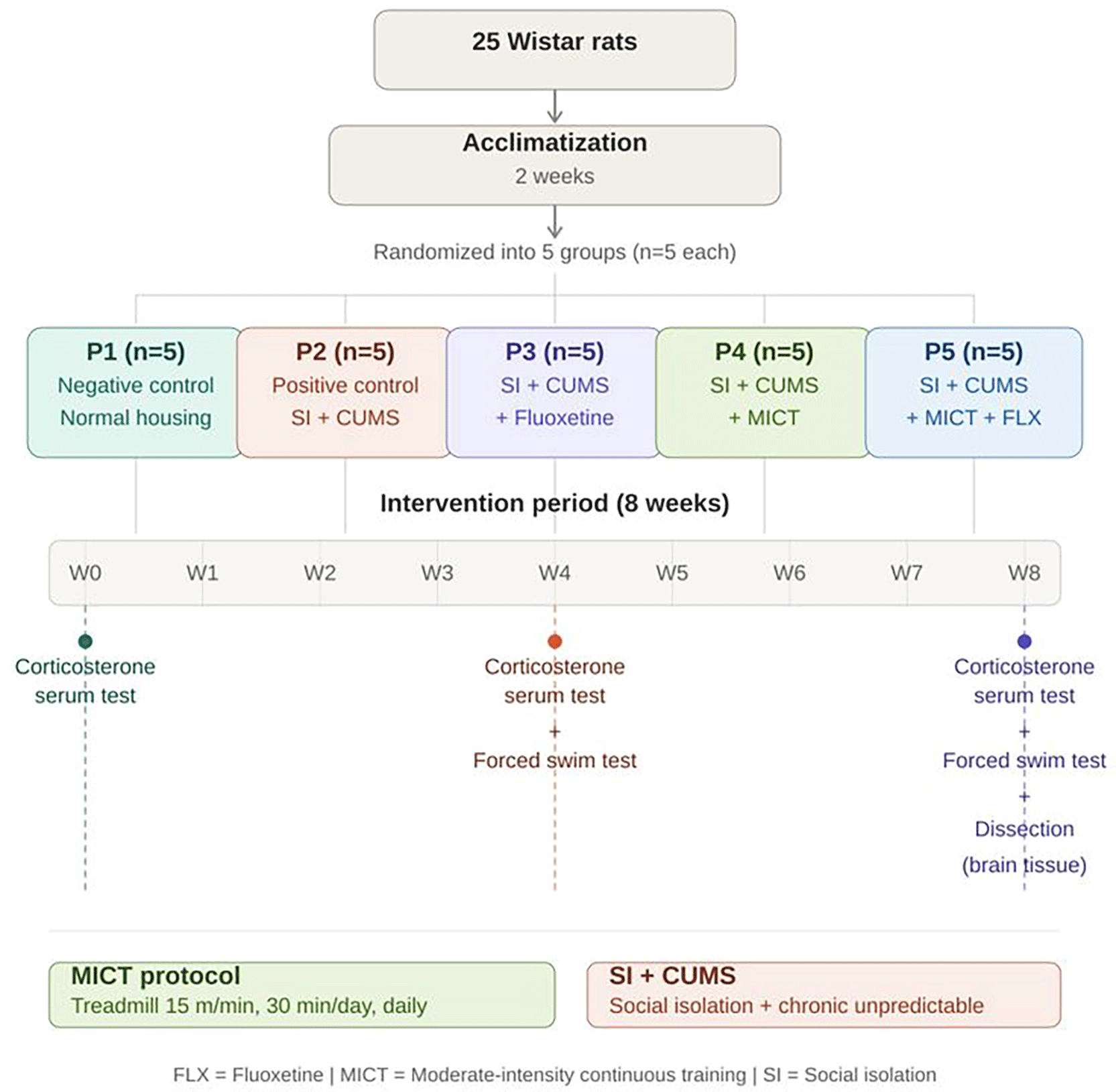
Twenty-five male Wistar rats were acclimatized for 2 weeks and randomly allocated into five groups. (n = 5 per group): P1 (negative control, normal housing), P2 (positive control, social isolation + CUMS), P3 (social isolation + CUMS + fluoxetine), P4 (social isolation + CUMS + MICT), and P5 (social isolation + CUMS + MICT + fluoxetine). MICT consisted of running on a treadmill at 15 m/min for 30 min daily. Serum corticosterone levels were measured at weeks 0, 4, and 8. Forced swim tests were conducted at weeks 4 and 8. All animals were dissected for brain tissue analysis at week 8.
Abbreviations: CUMS = Chronic Unpredictable Mild Stress; FLX = Fluoxetine; MICT = Moderate-Intensity Continuous Training; SI = Social Isolation.
Rats were subjected to a protocol to establish a depressed state with chronic unpredictable mild stress and social isolation. The unpredictable stress was induced by multiple stressors, including food and drink restrictions. Another stress comes from a 12-hour light/dark cycle (three days a week (Wednesday – Friday – Saturday): a 12-hour light/dark cycle is changed and continuous lighting is applied on Tuesdays and Sundays (24 hours) – change every week. During splash testing, a 10% sucrose solution is randomly sprayed on the back of the mouse. Every Wednesday, Friday, and Sunday, 200 ml of water is added to the bedding (100 g) and left for 24 h. Every Tuesday for 30 minutes, the Wistar rat was positioned within a cylindrical apparatus with a diameter of 80 mm and a height of 315 mm (Taffhome). Every 10 minutes, the cage was sprayed with a solution of 10% peppermint and 10% lemongrass (RealEagle).63
Intervention and behavioral test
Continuous training with moderate intensity and fluoxetine medication were the intervention for this study. Continuous training consisted of daily moderate-intensity treadmill running (Ideas). The treadmill speed was set at 15 m/min for 30 min. The training was held at 09.00 am.65 The pharmacotherapy used in this research was Fluoxetine (Nopres, DexaMedica), given daily by oral gavage at a dosage of 10 mg/kg for 2 months, at 08.00–09.00 am.66 The behavioral assessment employed in this study was the FST in weeks 4 and 8 at 08.00–10 am. The FST commenced with a 20-minute acclimatization period in the room containing the cylinders, during which both animals underwent simultaneous testing. The Wistar rats were evaluated within a transparent glass cylinder, with a diameter of 20 cm, filled with water to a depth of 39 cm at a temperature of 24°C. A Swanbee B-650/DC cylinder was used, measuring 65 cm in height and 24 cm in width. This test was conducted to assess the rat’s escape attempts from the cylinder during weeks 4 and 8, with each session lasting 6 minutes. The assessment focused on the frequency and duration of behaviors such as swimming, climbing, and immobility. Fresh water was replenished for each rat. During the FST, the Wistar rats were visually isolated from one another by placing dark gray dividers between the beakers. Following the completion of the experiment, all rats were dried using dry towels and subsequently placed in a warm cage for a duration of 15 minutes.56,64,67,68 Documentation of treadmill running and swimming test is presented on Figure 2 and Figure 3, respectively.

Rats underwent treadmill running at a speed of 15 m/min for 30 minutes daily during the intervention period.

The test was conducted in a transparent water-filled cylinder to evaluate swimming, climbing, and immobility behaviors during 6-minute observation sessions.
Abbreviation: FST = Forced Swimming Test.
Real-time polymerase chain reaction
Approximately 10 mg of prefrontal cortex tissue was homogenized in a Sodium Dodecyl Sulfate (SDS) Lysis Buffer Solution (Promega Wizard). The homogenate was centrifuged at 10,000 g for 10 minutes at 4°C, after which the supernatant was analyzed by RT-PCR.(Bioline SensiFAST SYBR No-ROX One Step Kit, cat. no. BIO-72005) to know the gene expression of Fibroblast Growth Factor-2 (FGF-2), Spermidine Synthase (Srm), Glutamate Metabotropic Receptor1 (m-GluR1), Vesicular Glutamate Transporter1 (VGLUT1) in the brain (The primer manufactured by Integrated DNA Technologies). The level of FGF-2 (Gene ID 54250), Spermidine Synthase (Srm), m-GluR1, and V-GLUT 1 was evaluated relative to that of the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The design of the primers was using the NCBI Primer-BLAST tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) based on NCBI Reference Sequences (RefSeq). Primer specificity was verified against the Rattus norvegicus RefSeq mRNA database. (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 4 May 2023. The design of the primers was using the NCBI Primer-BLAST tool based on NCBI Reference Sequences (RefSeq). Primer specificity was verified against the Rattus norvegicus RefSeq mRNA database. The primer sequences used in this study are shown in Table 1.
| Gene (Gene ID) | Official symbol | Direction | Primer sequence (5′ → 3′) | Product size (bp) | NCBI Accession No. |
|---|---|---|---|---|---|
| FGF-2 (54250) | Fgf2 | Forward | 5′-GGC TCT ACT GCA AGA ACG GC-3′, | 154 | NM_019305.2 |
| Reverse | 5′-TAC CGG TTC GCA CAC ACT CC-3′ | ||||
| Spermidine Synthase (84956) | Srm | Forward | 5′-TGG GTG GAC TCA GTC TCT GT-3′ | 100 | NM_053616.1 |
| Reverse | 5′ TTT GTT GGG TCA CAG GGC ATA-3′ | ||||
| mGluR1 (24414) | Grm1 | Forward | 5′-GAA CGG CTG CAG TCC TCT GA-3′ | 125 | NM_017011.1 |
| Reverse | 5′ TTT CAC TGG TGC CCG AAG GT-3′ | ||||
| V-GluT1 (116638) | Slc17a7 | Forward | [5′-GTG GTG CAG AAA GCC CAG TT-3 | 207 | NM_053859.2 |
| Reverse | 5-ACA GCC ATA GTG AAC ACG GG-3′ |
Blood was collected for corticosterone serum test from the tail of a rat between 9 and 10 am after.69,70 Using a serum separator tube, the samples were clotted for 30 min at room temperature before centrifugation for 15 min at 1000 × g. The blood serum was tested using the ELISA procedure for serum corticosterone level (ABClonal Corticosterone ELISA Kit 96 wells, cat. no. RK09054) in weeks 1, 4, and 8. The assays were performed in duplicate according to the manufacturer’s instructions.
Data analysis was conducted using the Statistical Package for the Social Sciences, version 24 (Chicago, IL, USA), with results expressed as mean ± SEM. The Kruskal-Wallis test was employed to determine statistical differences among the four groups, contingent on the normality test. The significance of the findings was further evaluated using the Mann–Whitney test and ANOVA, depending on the normality of the data. A p-value of 0.05 was considered statistically significant.
At baseline, body weight did not differ significantly among the five groups (p > 0.05), thereby confirming comparable initial conditions before the start of experimental procedures. The change in body weight (ΔBW) was determined by calculating the difference between BW at each subsequent time point and the baseline (H-0) for weeks 1, 4, and 8. Inter-group comparisons were conducted using the Mann–Whitney U test, with the PCG serving as the reference group.
By week 4, the divergent trajectory became apparent. The PCG displayed the highest cumulative weight gain among all groups (ΔBW: 57.80 ± 8.54 g), which was significantly greater than that of the NCG (ΔBW: 27.20 ± 5.51 g; p = 0).016). The treatment groups — PCG + FLX (ΔBW: 32.00 ± 14.94 g; p = 0.249), PCG + MICT (ΔBW: 47.20 ± 6.98 g; p = 0.421), and PCG + FLX + MICT (ΔBW: 39.80 ± 5.54 g; p = 0.310)—showed numerically lower weight gain compared to the PCG, but these differences did not reach statistical significance at this time.point.
At week 8, the inter-group divergence was most prominentnced. The PCG exhibited the greatest overall weight gain (ΔBW: 95.20 ± 10.37 g), significantly exceeding that of the NCG (ΔBW: 46.80 ± 5.46 g; p = 0).012). Notably, the combination therapy group (PCG + FLX + MCT) demonstrated significantly attenuated weight gain relative to the PCG (ΔBW: 70.80 ± 4.73 g; p = 0.036), whereas neither fluoxetine monotherapy (PCG + FLX; ΔBW: 64.60 ± 10.67 g; p = 0.059) nor exercise monotherapy (PCG + MICT; ΔBW: 82.40 ± 5.91 g; p = 0.143) reached significance individually. The near-significant trend observed in the PCG + FLX group (p = 0.059) suggests a potential weight-attenuating effect of fluoxetine that may be amplified when combined with exercise.
The results indicate that the combination of social isolation and the CUMS model resulted in significant weight gain in the PCG, which is consistent with stress-induced metabolic imbalances and possible overeating as a behavioral desperation symptoms.71 Changes in body weight across treatment groups over time are presented in Figure 4. The notable reduction in weight gain observed exclusively in the combination group at week 8—despite the lack of individual significance for either monotherapy—indicates a synergistic interaction between fluoxetine and CICT in normalizing stress-induced changes.
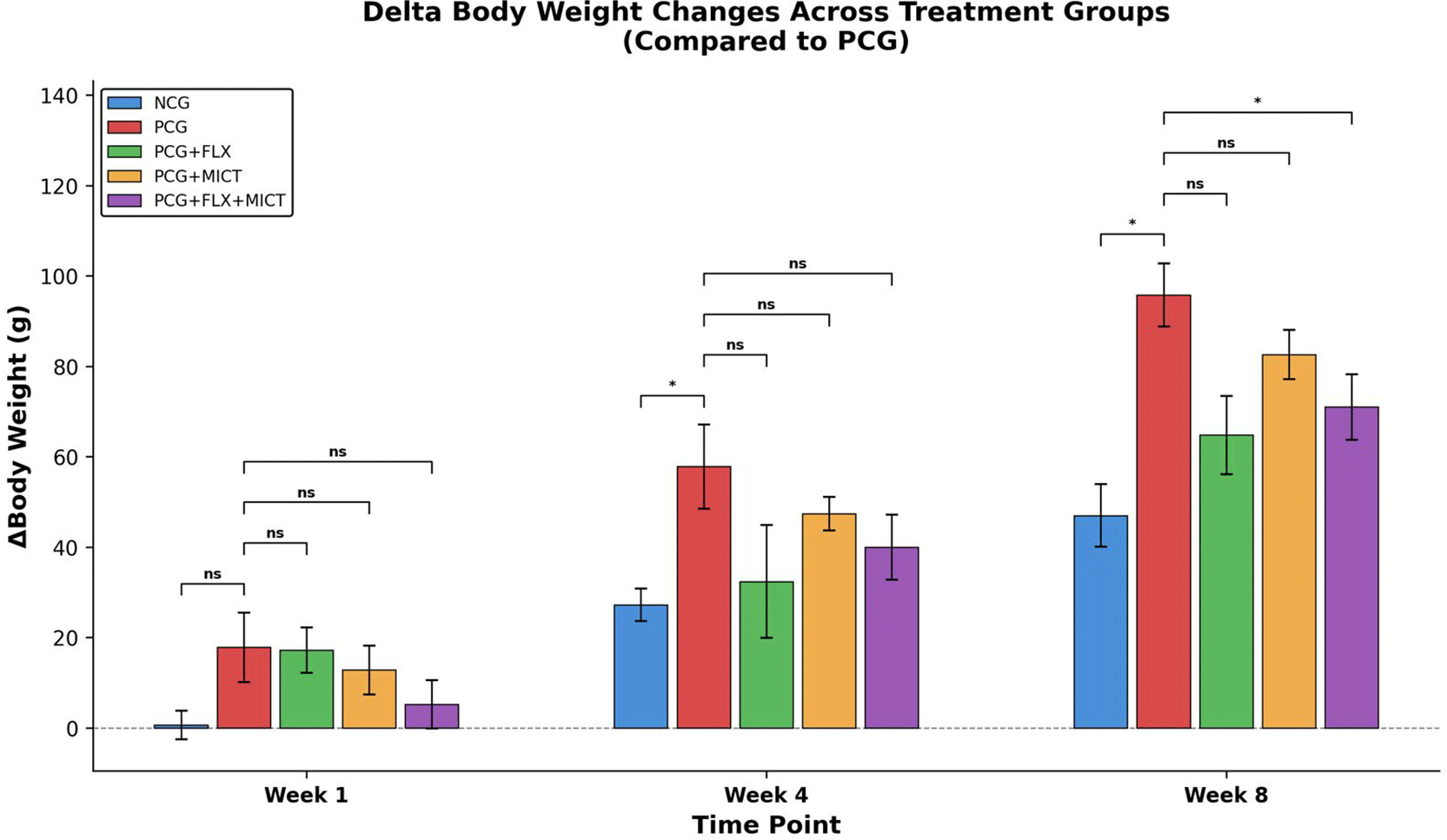
Abbreviations: NCG: Negative Control Group; PCG: Positive Control Group (depression model); PCG + FLX: Positive Control Group with Fluoxetine Treatment; PCG + MICT: Positive Control Group with Moderate Intervention Continuous Training; PCG + FLX + MICT: Model Depression Rat with Medical Intervention and Physical Activity Intervention.
Baseline corticosterone concentrations were consistent across all five groups (one-way analysis of variance, F(4,20) = 0.462, p = 0.763), indicating a uniform pre-intervention HPA-axis status. By week 8, the PCG exhibited the highest corticosterone level (36,091.87 ± 278.83 ng/mL), representing a 58.62% increase from baseline, with notably low inter-individual variability. This finding aligns with a consistent and pronounced elevation in corticosterone levels induced by chronic stress, driven by the combined social isolation and CUMS paradigm (paired t-test, t = −35.54, p < 0.0001).72,73 The negative control group maintained a relatively stable corticosterone trajectory over the 8-week period (23,602.12 ± 10,515.81 ng/mL; Δ baseline = +14.5%, paired t-test p = 0.466), confirming the absence of pathological HPA-axis activation in non-stressed animals. Among the treatment groups, the combination therapy group (PCG + FLX + MCT) demonstrated the most effective reduction of stress-induced corticosterone elevation, with a week-8 mean of 23,797.63 ± 3,116.32 ng/mL, a value virtually indistinguishable from the non-stressed NCG (t = −0.040, p = 0.969) and significantly lower than the PCG (t = 8.786, p < 0.0001). The MICT-alone group (PCG + MCT) showed a moderate but significant reduction compared with PCG (29,649.05 ± 2,489.75 ng/mL; t = 5.750, p = 0.0004), while fluoxetine monotherapy (PCG + FLX) yielded a numerically lower but highly variable week-8 corticosterone level (30,828.76 ± 11,724.35 ng/mL; p = 0.345 vs. PCG), likely reflecting individual differences in SSRI responsiveness within the depression model. Figure 5 illustrates the level of corticosterone serum across treatment groups over time.
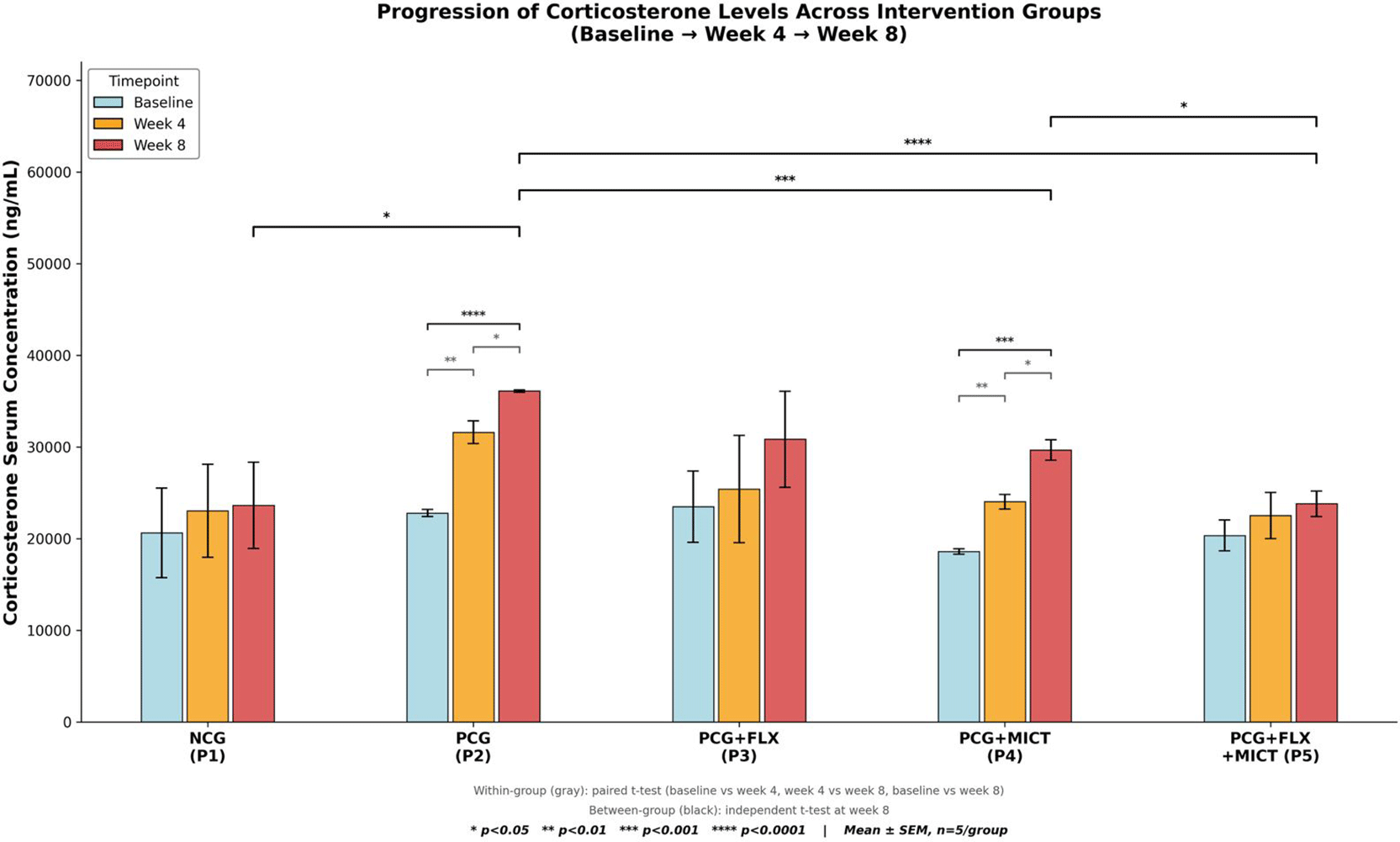
Bars represent mean ± SEM; individual data points are overlaid (n = 5 per group). Statistical comparisons were performed using t-tests with independent samples. *p < 0.05; ***p < 0.001; ****p < 0.0001.
Abbreviation: SEM: Standard Error of the Mean.
By the eighth week, both social isolation and CUMS caused a notable decrease in four indicators of brain health. The positive control group (PCG) showed significantly lower levels compared to the normal control group (NCG) for all indicators: FGF-2 (0.186 ± 0.061 vs 1.931 ± 0.327, p = 0.008), spermidine synthase/SRM (0.161 ± 0.040 vs 1.213 ± 0.108, p = 0.012), V-GluT1 (0.571 ± 0.029 vs 3.478 ± 0.501, p = 0.012), and mGluR1 (0.501 ± 0.063 vs 1.521 ± 0.494, p = 0.008; Mann-Whitney U test). The Kruskal-Wallis test indicated significant differences among groups for all markers (FGF-2, p = 0.001; SRM, p < 0.001; V-GLUT1, p < 0.001; mGluR1, p = 0.011). All three treatments led to increased levels of FGF-2, SRM, and V-GLUT1 compared with PCG (all, p < 0.05). However, a difference was noted with mGluR1 levels: fluoxetine alone did not bring mGluR1 back to normal levels (1.072 ± 0.506 vs PCG 0.501 ± 0.063, p = 0.841), whereas moderate-intensity continuous training (MICT) (1.726 ± 0.330, p = 0.008) and combination therapy (2.527 ± 0.239, p = 0.008) did. This selective resistance of metabotropic glutamate receptor expression to SSRI monotherapy, despite successful restoration of neurotrophic (FGF-2) and presynaptic glutamatergic (V-GLUT1) markers in the same treatment group, indicates that fluoxetine-mediated serotonergic enhancement does not uniformly engage all neuroplasticity domains and that exercise-dependent activation is required for postsynaptic Group I mGluR signaling.
Expression of neuroplasticity biomarkers across treatment groups at week 8 is presented by Figure 6. The combination therapy (PCG + FLX + MICT) consistently exhibited the highest fold-change relative to PCG across all four biomarkers: FGF-2 (20.14-fold), SRM (16.78-fold), V-GLUT1 (5.59-fold), and mGluR1 (5.01-fold). The combination group was the only intervention that fully normalized V-GLUT1 expression to levels statistically indistinguishable from normal controls (3.198 ± 0.270 vs NCG 3.478 ± 0.501, p = 1.000), whereas fluoxetine alone (1.434 ± 0.241, p = 0.008 vs NCG) and MICT alone (1.811 ± 0.483, p = 0.056 vs NCG) remained significantly below physiological levels. This indicates that the complete restoration of vesicular glutamate packaging capacity necessitates the combined effect of both interventions. Furthermore, SRM expression in the combination group (2.752 ± 0.151) significantly exceeded normal control levels (1.213 ± 0.108, p = 0.012), indicating a supraphysiological upregulation not observed in any of the monotherapy groups. The overshoot pattern in SRM expression among all biomarkers analyzed suggests that the combination of fluoxetine and MICT converges on the polyamine biosynthesis pathway through distinct upstream mechanisms. This convergence results in an amplification effect that elevates the expression of spermidine synthase beyond the homeostatic set points. Collectively, these findings demonstrate a clear dose-response hierarchy across depression intervention modalities. (PCG < PCG + FLX < PCG + MICT < PCG + FLX + MICT) and the results suggest that each biomarker exhibits a distinct therapeutic response profile. Specifically, mGluR1 is uniquely reliant on exercise, V-GLUT1 requires combination therapy for full normalization, and SRM and FGF-2 are particularly responsive to synergistic induction.
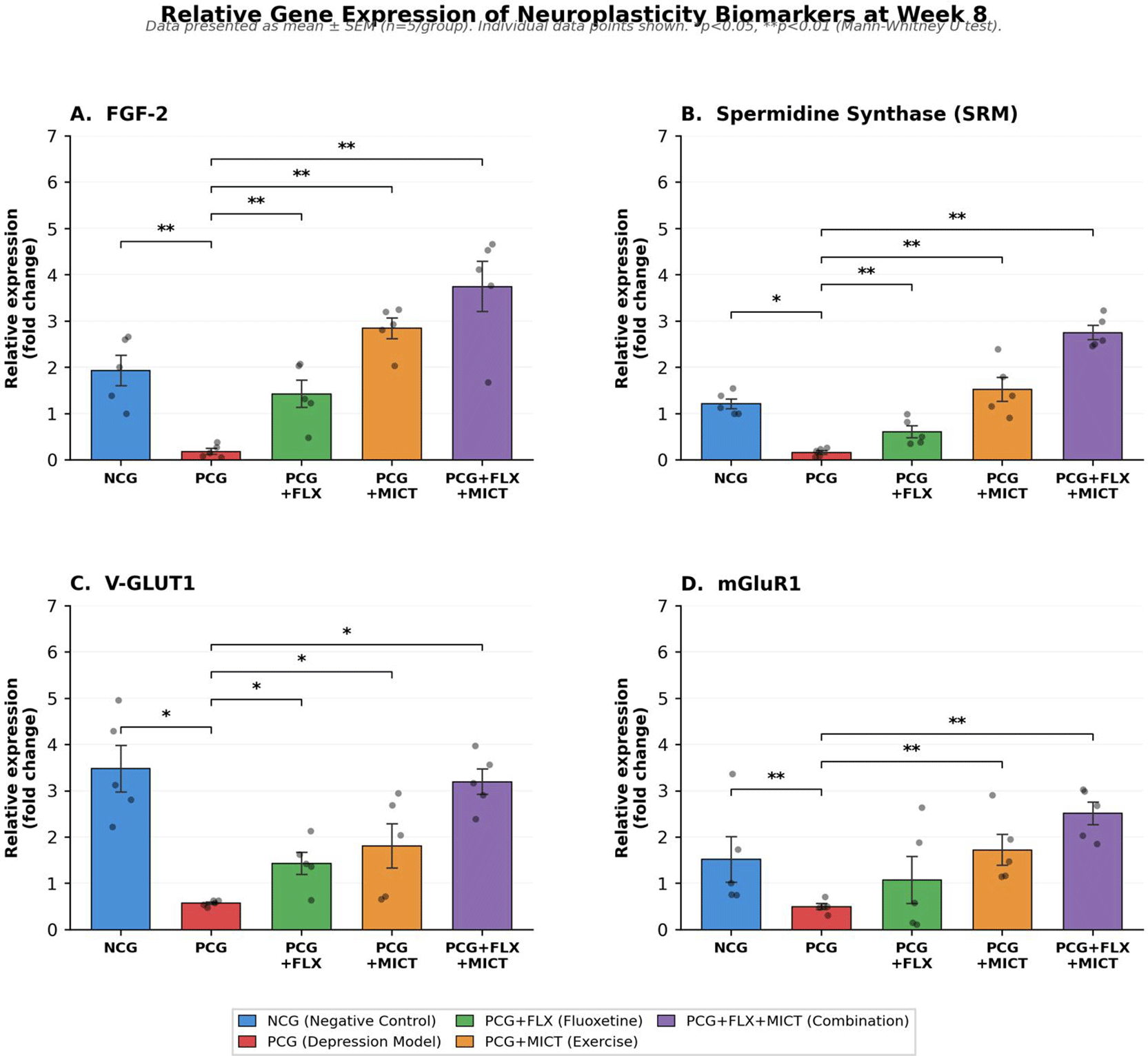
Bar graphs represent the relative expression of mRNA (A) Fibroblast Growth Factor-2 (FGF-2), (B) Spermidine Synthase (SRM), (C) Vesicular Glutamate Transporter 1 (V-GLUT1), and (D) Metabotropic Glutamate Receptor 1 (mGluR1), measured at the end of the experimental period, in the 8th week. Values are expressed as fold-change relative to the housekeeping gene and presented as mean ± SEM (n = 5 per group). Individual data points are displayed for each animal. NCG = negative control group (non-isolated); PCG = positive control group (social isolation depression model, untreated); PCG + FLX = depression model treated with fluoxetine; PCG + MICT = depression model treated with MICT; PCG + FLX + MICT = depression model treated with combined fluoxetine and MICT. Omnibus analysis was performed using the Kruskal-Wallis test (FGF-2: H = 18.44, p = 0.001; SRM: H = 21.76, p < 0.001; V-GLUT1: H = 19.01, p < 0.001; mGluR1: H = 12.96, p = 0.011). Post hoc pairwise comparisons were conducted using the Mann–Whitney U test (two-tailed). Significance brackets indicate statistically significant differences: *p < 0.05, **p < 0.01. Note the absence of a significance bracket between PCG and PCG + FLX in panel D, indicating that mGluR1 expression was not significantly restored by fluoxetine monotherapy (p = 0.841). The combination therapy (PCG + FLX + MICT) achieved the highest fold-change relative to PCG across all four biomarkers: FGF-2 (20.14×), SRM (16.78×), V-GLUT1 (5.59×), and mGluR1 (5.01×).
Abbreviations: FGF-2: Fibroblast Growth Factor-2; SRM: Spermidine Synthase; V-GLUT1: Vesicular Glutamate Transporter 1; mGluR1: Metabotropic Glutamate Receptor 1; SEM: Standard Error of The Mean; NCG: Negative Control Group; PCG: Positive Control Group (depression model); PCG + FLX: Positive Control Group with Fluoxetine Treatment; PCG + MICT: Positive Control Group with Moderate Intervention Continuous Training; PCG + FLX + MICT: Model Depression Rat with Medical Intervention and Physical Activity Intervention.
The present study demonstrated that the dual-stressor paradigm, which combines CUMS with CUMS, resulted in a robust, progressive, and statistically significant increase in serum corticosterone levels in the PCG. Notably, the remarkably low coefficient of variation in the PCG at week 8 (0.77%) indicates a homogeneous and saturated hyperactivation of the hypothalamic-pituitary-adrenal (HPA) axis across all subjects, thereby confirming the reliability and reproducibility of this combined depression model. This stepwise escalation is in stark contrast to the NCG, which maintained stable corticosterone levels throughout the experimental period. These findings confirmed that chronic social isolation in adult Wistar rats results in sustained elevations in plasma corticosterone levels, accompanied by alterations in synaptic plasticity markers.74
The concurrent application of social isolation and unpredictable mild stress resulted in a depressive-like phenotype compared to the effects of each individual stressorally. Prolonged social isolation can adversely affect the stress response system of the body, leading to elevated corticosterone levels, brain atrophy, and prefrontal cortex.75 This phenotype was characterized by both behavioral despair and sustained corticosterone levels. Previous research has demonstrated that both pharmacological and exercise interventions can mitigate the heightened stress response associated with social isolation and chronic stress.76,77 The comparable efficacy of MICT and FLX treatments is significant as it supports the notion that exercise may serve as a viable non-pharmacological alternative with effects akin to conventional antidepressant therapies.78,79
This pattern was successfully replicated in the present study, thereby enhancing the face and construct validity of our dual-stressor model for producing a depressed male Wistar model. Rats. Our findings are consistent with the mechanistic framework proposed by previous research, which demonstrated that chronic glucocorticoid excess induces a harmful mineralocorticoid receptor (MR)/glucocorticoid receptor (GR) imbalanceance. This imbalance impairs the negative hippocampal feedback loop, thereby perpetuating hyperactivation and HPA ax dysfunctionis.80 Temporal corticosterone profiles across intervention groups reveal a critical dissociation between pharmacological, physical, and combination therapies in their capacity to normalize HPA-axis function. The combination therapy group (PCG + FLX + MCT) demonstrated the most synergisticcome.
Corticosterone data present compelling evidence that the combination of fluoxetine and moderate-intensity continuous training exerts a synergistic normalizing effect on hypothalamic-pituitary-adrenal (HPA) axis dysregulation induced by social isolation combined with chronic unpredictable mild stress (CUMS). This finding underscores the dual-stressor paradigm’s efficacy in producing a sustained neuroendocrine hallmark of depression, aligning with previous reports that chronic social isolation amplifies glucocorticoid release through impaired negative feedback at the hippocampal mineralocorticoid and glucocorticoid receptor levels.81–83
Furthermore, the polyamine pathway and molecular neuroplasticity are disrupted in depression caused by social isolation and chronic stress.21,40,84,85 This finding matches a recent study that found imbalanced polyamine levels, neuroplasticity problems, and autophagy dysfunction in depression exposed to chronic stress and social isolation.23,40,72,74 The study showed that the expression of spermidine synthase, the enzyme catalyzing the final step of spermidine biosynthesis, was decreased in the depression condition and recovered after the combinationtion. This study also showed that the combination supports the hypothesis that pharmacological restoration of serotonergic tone by fluoxetine and exercise-mediated upregulation of BDNF, FGF-2, and hippocampal neurogenesis converge on complementary mechanisms to restore glucocorticoid receptor sensitivity and reinstate HPA-axis negative feedback.34,86–89 This recovery in the combination group suggests a possible complementary synergistic effect of exercise boosts polyamine production by increasing ornithine decarboxylase activity, and SSRIs help restore spermine N1-acetyltransferase (SSAT) expression, which is usually low inon.90
Glutamate release mediated by VGLUT1 and the subsequent activation of membrane GluA1 may be key to rapid-antidepressant-like efficacyeits. VGluT1 acts as the main vesicular glutamate transporter in cortical neurons, enabling the loading of glutamate into presynaptic vesicles, a step essential for glutamatergic excitatory transmisssion.30,91 Meanwhile, M-GluR1 is essential for inducing synaptic plasticitycess. Research involving VGLUT1 heterozygous mice has demonstrated that short-term chronic mild stress, in conjunction with reduced VGLUT1 expression in VGLUT1−/− mice, modifies glutamate transmission,92,93 resulting in depression-like behaviors and memory recognition deficits.91,94 In the present study, we showed that the interventions produced graded recovery of VGluT1 in the prefrontal cortex with the combination of FLX and MICT, which nearly restored NCG levels. The VGluT1 response in the MICT group is consistent with a previous study that showed that antidepressant agents selectively increase cortical VGluT1 mRNA, indicating that VGluT1 upregulation is a common marker of antidepressant efficacy.91,95 This study showed that a combination of moderate-intensity continuous training exercise and pharmacotherapy synergy to the glutamate vesicular packaging systemstem. The observed coordinated upregulation of both V-GluT1 and M-GluR1 in the intervention groups suggests a comprehensive restoration of glutamate cycling in the prefrontal cortex, encompassing both presynaptic vesicular packaging and postsynaptic/glial reuptake mechanisms.
These findings showed that in the management of depression, a single intervention approach is insufficient for dealing with the magnitude of depression disorder in the brain. The multimodal approaches, especially pharmacotherapy and exercise training, have synergistic effects on the recovery from depression and HPA axis normalization.
Chronic social isolation triggers and CUMS-sustained hyperactivation of the HPA axis in male Wistar rats, which in turn suppresses neurotrophic factors (FGF-2), disrupts the polyamine–autophagy axis (spermidine), and impairs glutamatergic neurotransmission at both vesicular packaging (V-GluT1) and M (M-GluR1) levels. This represents a comprehensive “molecular cascade of depression” that extends beyond the traditional monoamine hypothesis to encompass glutamatergic, neurotrophic, and metabolomic dimensions.
The combined moderate-intensity continuous training exercise and fluoxetine therapy produces synergistic effects in depression, achieving more than normal spermidine synthase and FGF-2 gene expression levels and near-complete normalization of V-GluT1 and M-GluR1 gene expression1.
Several limitations of this study should be acknowledged. First, the research exclusively used male Wistar rats; given the well-documented sex-dependent differences in HPA axis regulation and neuroplasticity, this may limit the generalizability of the findings to female subjects.96,97 SecondMoreover, although gene expression was quantified at the mRNA level, protein-level validation was not conducted, and post-translational modifications could influence the activity of functional transporters or growth factors. Third, the mechanisms underlying the synergistic effects of combined therapy remain speculative and require further investigation through pathway-specific pharmacological blockade or genetic manipulation. Future research should incorporate both sexes, protein-level analyses, option from other antidepressant medication, comprehensive behavioral testing, and dose-response exercise paradigms to further elucidate the optimal therapeutic window for combined exercise–pharmacotherapy interventions in depression.
During the intermediate stage of the editorial review, various phrasing and wording alternatives were crafted using the generative AI tool Paperpal Pro, while the figure was developed with the assistance of Claude Pro. All original revisions were selected, modified, and reviewed by the authors to verify their accuracy and precision. The authors assume full responsibility for the final paper and affirm that the incorporation of AI did not affect the study’s outcomes or scientific integrity.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)