Keywords
Abiotic Stress; CRISPR/Cas9; Cytoplasmic Male Sterility; Hybrid Rice; Molecular Breeding.
This article is included in the Agriculture, Food and Nutrition gateway.
Abiotic Stress; CRISPR/Cas9; Cytoplasmic Male Sterility; Hybrid Rice; Molecular Breeding.
Increasing global food demand, climate change, and land resource degradation have made improving crop productivity and resilience to abiotic stress a major challenge in rice development. As the staple food for more than half of the world’s population, rice production plays a strategic role in maintaining global food security. The FAO (2025) projects that global rice consumption will increase by approximately 1% annually until 2034, with Asia accounting for nearly 69% of total consumption growth. At the same time, the increasing frequency of drought, salinity, extreme temperatures, and flooding associated with climate change continues to constrain rice productivity across major production regions (Aruneshwaran et al., 2025). These conditions highlight the urgent need for breeding innovations that enhance yield performance while strengthening crop adaptability under increasingly unstable environmental conditions.
Hybrid rice has emerged as one of the most promising breeding approaches for addressing these challenges. By exploiting heterosis, hybrid rice has been reported to achieve a 15–20% yield advantage over elite inbred cultivars (Virmani & Kumar, 2004). Commercial hybrid rice production is generally based on two major systems, namely the three-line and two-line systems. The three-line system, which relies on cytoplasmic male sterility (CMS), uses a cytoplasmic male-sterile line (A line), a maintainer line (B line), and a fertility restorer line (R line) to produce stable, genetically uniform hybrid seeds. In contrast, the two-line system utilizes genic male-sterile lines whose sterility is regulated by temperature and photoperiod, thereby simplifying hybrid seed production by eliminating the need for a maintainer line (Guo-hui & Long-ping, 2015). This system includes thermo-sensitive genic male sterility (TGMS), photoperiod-sensitive genic male sterility (PGMS), and photoperiod-thermo-sensitive genic male sterility (PTGMS) (Xu et al., 2023). The stability of male sterility expression in the female parent line is a critical determinant of successful hybrid seed production, as it prevents self-pollination and maintains the genetic purity of hybrid seeds.
Global climate change has further increased the complexity of hybrid rice development by intensifying abiotic stresses that affect plant growth, reproductive stability, and grain yield. Kandpal et al. (2020) demonstrated that hormonal regulation associated with abiotic stress responses influences peduncle elongation, spikelet fertility, and grain filling in male sterile lines. These findings suggest that the success of hybrid rice systems depends not only on heterotic yield potential but also on sterile lines’ ability to maintain reproductive stability under environmental stress. In recent years, advances in molecular breeding technologies, including marker-assisted breeding, QTL introgression, genomic selection, and CRISPR/Cas9 genome editing, have accelerated the incorporation of stress-tolerant traits into hybrid rice breeding programs (Gregorio et al., 2013). Nevertheless, information on the relationships among male sterility systems, hybrid performance, and physiological and molecular mechanisms of adaptation to abiotic stress remains fragmented across the literature and has not yet been comprehensively synthesized.
Furthermore, most existing studies focus on single-stress conditions in controlled environments, whereas combinations of stresses, such as drought, heat, and salinity, are increasingly common in modern agroecosystems (Suzuki et al., 2014; Altaf et al., 2021; Anwar et al., 2021). Limited multilocation evaluations and insufficient integration of multi-omics approaches have also restricted the understanding of hybrid adaptation stability under heterogeneous field conditions. Therefore, a systematic literature review is required to synthesize current research developments, identify emerging trends and knowledge gaps, and formulate future directions for the development of abiotic-stress-tolerant hybrid rice that is more adaptive and sustainable.
Based on this background, the present study aims to comprehensively analyze the role of male sterility systems in the development of abiotic stress-tolerant hybrid rice using a systematic literature review. The findings are expected to provide a scientific foundation for advancing breeding strategies and integrating modern molecular technologies to develop hybrid rice cultivars with improved productivity, stability, and resilience under global climate change.
This study employed a systematic literature review (SLR) to synthesise scientific evidence on the development of hybrid rice using male-sterility systems for abiotic stress tolerance. The review was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines to ensure methodological transparency and reproducibility (Donthu et al., 2021; Permatasari et al., 2025). The review process consisted of four stages: study identification, duplicate removal, title and abstract screening, and full-text eligibility assessment based on predefined inclusion and exclusion criteria.
A comprehensive literature search was conducted using the Scopus and PubMed databases because of their broad coverage of peer-reviewed publications in agricultural, biological, molecular, and environmental sciences. The search strategy combined controlled keywords and Boolean operators related to male sterility systems, hybrid rice, and abiotic stress. The following search syntax was applied across both databases: ((“male sterile” OR “cytoplasmic male sterility*” OR “genic male sterile*” OR “restorer” OR “maintainer”) AND (“rice” OR “Oryza sativa” OR “paddy” OR “hybrid rice”) AND (“abiotic stress” OR “environmental stress” OR “heat stress” OR “salinity stress” OR “drought stress” OR “salt stress” OR “water stress”))*.
The initial search retrieved 103 records. Additional filters were applied to include only English-language articles published between 2017 and 2026. In Scopus, the search was further restricted to the subject areas of Agricultural and Biological Sciences, Biochemistry, Genetics and Molecular Biology, and Environmental Sciences, and to original research articles published in peer-reviewed journals. After filtering, 56 articles met the preliminary eligibility criteria, comprising 49 records from Scopus and 7 from PubMed.
All records were consolidated into a single database, and seven duplicate articles were removed, leaving 49 studies for screening. Title and abstract screening was conducted independently by five reviewers (NM, AM, MKN, DC, and RS). Studies unrelated to hybrid rice, male sterility systems, or abiotic stress responses were excluded. Following screening, 39 studies proceeded to full-text assessment. All full-text articles were successfully retrieved and evaluated for conceptual relevance and methodological suitability. The final selection process followed the PRISMA framework, and the overall workflow is presented in Figure 1.
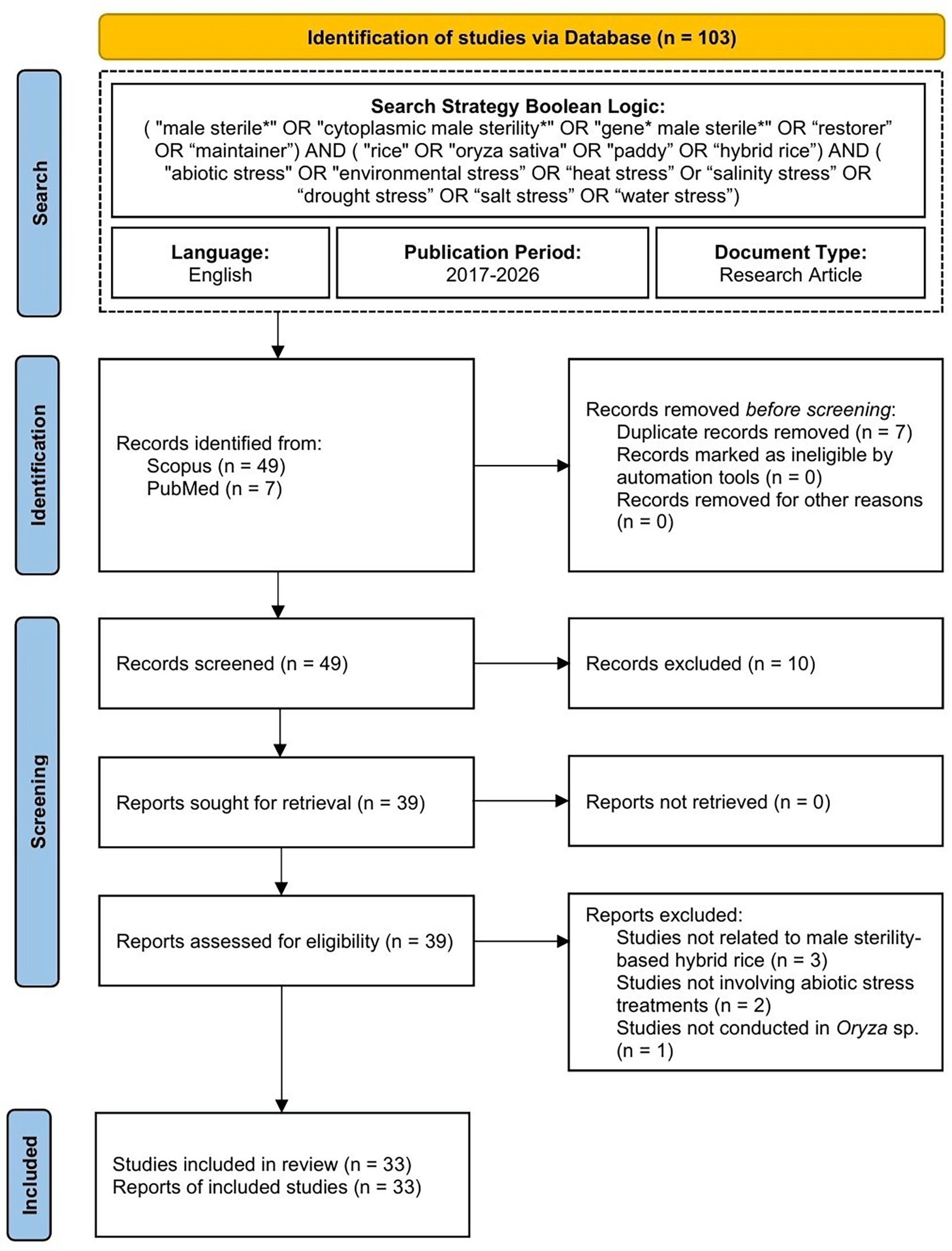
An eligibility assessment was conducted on 39 full-text articles to identify studies related to the development of abiotic stress-tolerant hybrid rice using male sterility systems. Study selection followed the PICO framework (Population, Intervention, Comparison, Outcome) proposed by Eldawlatly et al. (2018) with predefined inclusion and exclusion criteria summarised in Table 1. Articles were evaluated systematically based on their titles, abstracts, and full texts to ensure relevance to the objectives of the review.
Six studies were excluded because they involved non-rice commodities (Zhao et al., 2022), hybrid approaches unrelated to male sterility systems (Prasanth et al., 2017; Mo et al., 2023; Li et al., 2025), or focused solely on stress-response mechanisms without evaluating abiotic stress treatments in hybrid rice systems (Xia & Luo, 2023; Kandpal et al., 2020). Consequently, 33 articles were included in the final analysis. Data extraction was conducted systematically using a structured spreadsheet database. Extracted information included study characteristics, male sterility systems, genetic backgrounds, abiotic stress treatments, physiological and molecular responses, agronomic performance, and breeding strategies.
Due to substantial variation in experimental design, stress intensity, and evaluation parameters, data were synthesised using a narrative-qualitative approach rather than quantitative meta-analysis. The studies were grouped by abiotic stress type: drought, salinity, heat, low temperature, phosphorus deficiency, and combined stresses. Analyses focused on the effectiveness of male sterility systems and hybrid breeding strategies in improving stress tolerance, reproductive stability, and agronomic performance. The synthesis results are presented through summary tables, conceptual figures, and analytical narratives to identify current research trends and critical gaps in climate-resilient hybrid rice breeding.
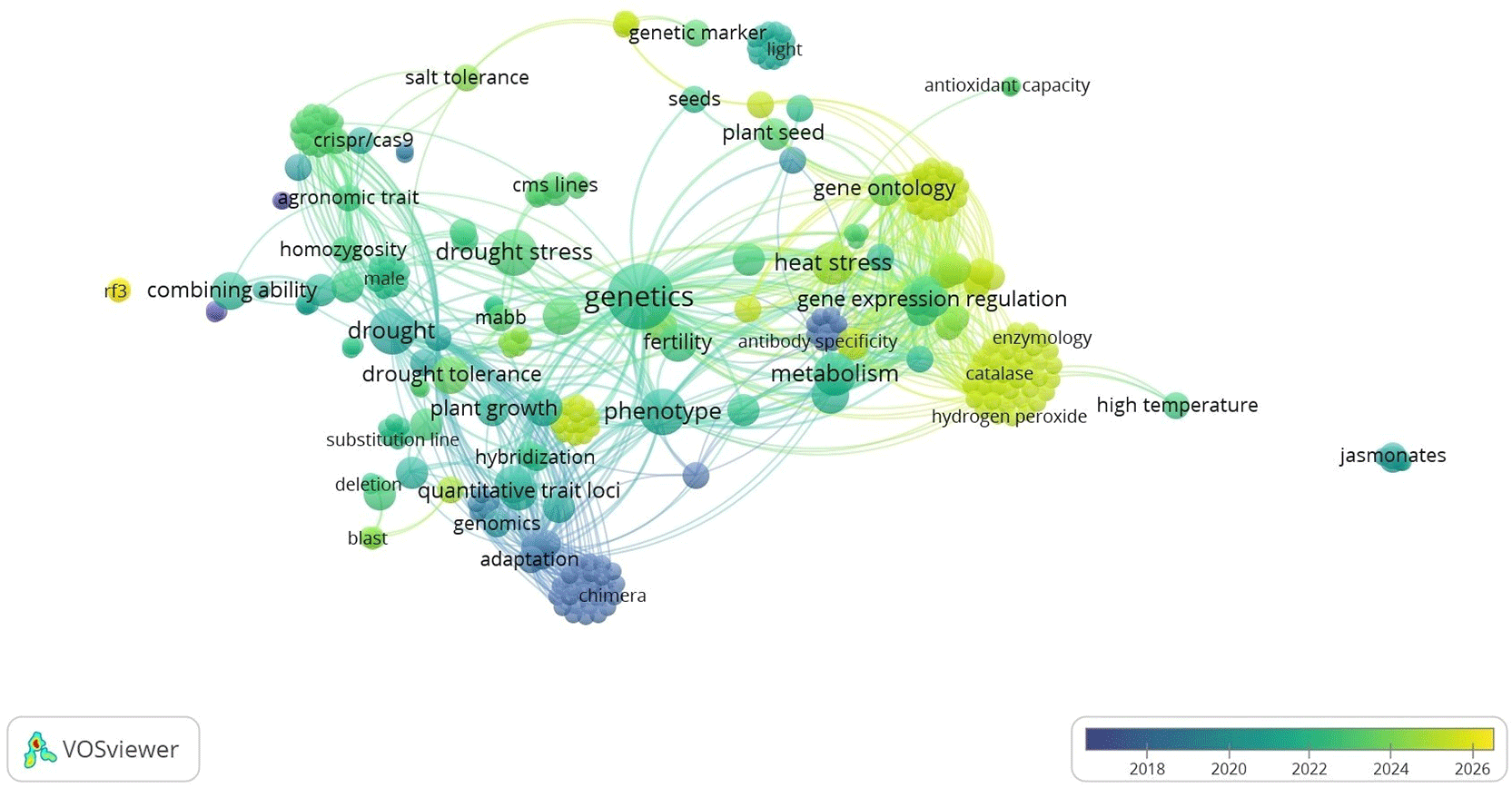
All literature included in this review met the eligibility criteria presented in Table 1 and comprised studies published between 2017 and 2026 on environmental stress tolerance in hybrid rice. Bibliometric visualisation using VOSviewer identified two major thematic clusters ( Figure 2): drought-related studies associated with CRISPR/Cas9, agronomic traits, and homozygosity, and more recent heat-stress research linked to metabolic regulation, catalase activity, and hydrogen peroxide scavenging. Node size indicated keyword frequency, with genetics, phenotype, drought stress, and gene ontology emerging as the most dominant themes. At the same time, network density highlighted strong co-occurrence among genetics, phenotype, environmental stress, and genetic engineering. Temporal overlay analysis further revealed a shift in research focus from foundational topics such as hybridisation, QTL mapping, genomics, and combining ability (2018) toward drought tolerance, CRISPR/Cas9, and heat stress (2020), followed by recent emphasis (2022–2026) on Rf3 genetics, gene expression, enzymology, antioxidant regulation, and high-temperature tolerance. Overall, these trends indicate a shift from conventional breeding toward molecular and physiological strategies to improve antioxidant defence systems and heat resilience in hybrid rice.
Hybrid rice (Oryza sativa) performance is strongly influenced by Cytoplasmic Male Sterility (CMS) systems, particularly Wild-Abortive CMS (WA-CMS), which are regulated by mitochondrial genes that disrupt pollen development and enable efficient hybrid seed production. Beyond reproductive control, CMS systems contribute to agronomic improvement through enhanced specific combining ability (SCA), heterosis, and hybrid vigor arising from interactions among CMS, maintainer, and restorer lines. CMS-derived hybrids also exhibit superior germination, stronger root architecture, and improved seedling establishment under drought stress, reflecting the importance of nuclear–cytoplasmic interactions and stress-responsive metabolic regulation.
As summarized in Table 2, hybrid rice breeding over the last decade has been predominantly centred on CMS-based three-line systems, with WA-CMS remaining the most widely utilized platform because of its stable sterility expression, broad fertility restoration capacity, and strong association with high yield performance (Hong-Bing et al., 2017; Widyastuti et al., 2017; Liao et al., 2019; Luo et al., 2019; El-mowafi et al., 2021; Madhusudan et al., 2022; Awad-allah et al., 2022; Li et al., 2023; Ghazy et al., 2023; Bhuyan et al., 2025). The recurrent use of elite CMS lines such as IR58025A and IR69625A further indicates the continued dependence of breeding programs on a limited number of reliable cytoplasmic donor sources. At the molecular level, CMS is associated with aberrant mitochondrial genes that induce maternally inherited male sterility, providing an efficient biological system for large-scale hybrid seed production without manual emasculation.
| Material | Lines | System component | Functional role in hybrid system | Key findings | Genetic aspect | References | Main findings related to hybrid performance | Stress type |
|---|---|---|---|---|---|---|---|---|
| Oryza sativa | A line: IR58025A, APMS6A | WA-CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Priyadarshi et al. (2025) | Hybrids carrying Rf3/Rf4 showed higher fertility restoration, high SCA, and significant heterosis for grain yield | Cold |
| R-line: KMR 3; IR 93376- B- B- 130; IR 96322–34-223- B- 1-1- 1; IR 102774–31- 21- 2- 4-7; TDK 1; IR 99734:1–33- 69- 1- 12- LSM- 1; IR 99734:1–33- 69- 1- 9- LSM- 1; IR 99734:1–33- 69- 1- 39-6; IR 99734:1–33–69-1- 12-4; IR 102783:2–70-91- 2- 1-2 and IR83383- B- B (RP5333-12–2-1); KMR3 | ||||||||
| M-line : R 58025B | ||||||||
| Oryza sativa | A-line: IR58025A; IR80154A; IR80156A; GMJ13A; GMJ14A; GMJ15A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Widyastuti et al. (2017) | Certain hybrids maintained better germination and root vigor under drought stress | Drought & Waterlogging |
| R-line: PK90; R3; PK12; R32; BP11 | ||||||||
| Oryza sativa L. subsp. indica | R-line: R225 | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in CMS hybrids | Nuclear restorer gene (Rf gene) | Wang et al. (2025) | WT plants maintained higher yield stability than ts12 mutants under high temperature | Drought |
| Oryza sativa | R-line: Chenghui727 | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in CMS hybrids | Nuclear restorer gene (Rf gene) | Nabi et al. (2022) | Hybrid Deyou4727 showed superior yield and root performance under drought | Drought |
| Oryza sativa | R-line: 996 (HT 996); 343 (HS 343) | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in CMS hybrids | Nuclear restorer gene (Rf gene) | You et al. (2023) | Mixed nitrogen improved spikelet fertility and yield under heat stress | Drought |
| Oryza sativa | A-line: Gang46A (L1); IR69625A (L2) | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Awad-allah et al. (2022) | Several hybrids exhibited positive SCA and lower yield reduction under drought | Drought |
| R-line: NRL 2; NRL 9; NRL 10; NRL 11; NRL 12; NRL 29; NRL 42; NRL 43; NRL 44; NRL 47; NRL 50; Giza178 | ||||||||
| Oryza sativa | Wan2304S; S240; 2X5S; H03S; N95076S; N5088S.(etc) | ESGMS | Environment-dependent male sterility for two-line hybrid breeding | Conditional male sterility expression | Nuclear-controlled sterility gene | Zhang et al. (2021) | CMS and restorer lines showed high genetic purity suitable for hybrid breeding | Drought |
| O. sativa ssp japonica | A-line: oshsp60-3b male-sterile mutant | ESGMS | Environment-dependent male sterility for two-line hybrid breeding | Conditional male sterility expression | Nuclear-controlled sterility gene | Lin et al. (2023) | High temperature caused pollen sterility and reproductive abnormalities | Drought |
| Oryza sativa L. ssp japonica | A-line: Chinsurah Boro II | BT-CMS | Female parent in three-line hybrid breeding | Reliable cytoplasmic male sterility with effective fertility restoration by Rf genes | Cytoplasmic–nuclear interaction involving mitochondrial sterility genes | Zhang et al. (2017) | Fertility restoration was strongly associated with restorer gene combinations | Drought |
| Oryza sativa L. subsp. indica | R-line: R996, R4628 | Restorer | Pollen fertility restoration | Conditional male sterility expression | Nuclear-controlled sterility gene | Zhou et al. (2025) | Heat stress reduced reproductive performance and grain filling efficiency | Drought |
| Oryza sativa L. subsp. japonica | A-line : Shen 21A | BT-CMS | Male sterility induction | Reliable cytoplasmic male sterility with effective fertility restoration by Rf genes | Cytoplasmic–nuclear interaction involving mitochondrial sterility genes | Niu et al. (2025) | Heat stress disrupted reproductive tissue development | Salinity |
| Oryza sativa | R-line: KMR-3R; IL-1; IL-2; IL-3; IL-4; IL-5; IL-6; IL-7; IL-8 | Restorer | Pollen fertility restoration | Conditional male sterility expression | Nuclear-controlled sterility gene | Nagaraju et al. (2023) | Non-additive gene action contributed strongly to hybrid superiority | Drought |
| Oryza rufipogon Griff., Oryza sativa | R-line: R974 | Restorer | Pollen fertility restoration | Conditional male sterility expression | Nuclear-controlled sterility gene | Ding et al. (2022) | Heat stress disrupted reproductive tissue development | Drought |
| Oryza sativa | A-line: Peiai 64S; Shen 08S | PTGMS | Environment-sensitive male sterility for two-line hybrid breeding | Stable sterility–fertility transition under specific temperature and photoperiod conditions | Photo-thermo sensitive nuclear sterility gene | Chen et al. (2021) | Hybrid lines produced higher biomass and yield than parental lines | Drought |
| Oryza sativa L. subsp. indica | A-line: APMS-6A | WA-CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Sravanraju et al. (2024) | Several crosses showed positive heterosis and superior yield | Drought |
| R-line: RPHR-1005R, RPHR6339-4-16-14-2 (IL1); RPHR6339-4-16-14-3 (IL2); RPHR6339-4-16-14–7 (IL3); RPHR6339-4-16-14-8 (IL4); RPHR6339-4-16-14-16 (IL5); RPHR6339-4-16-14-22 (IL6) | ||||||||
| Oryza sativa L. subsp. Indica | M-line: 733Srr22-T1447–1 | GMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Sheng et al. (2023) | Heat stress reduced pollen viability and seed setting | Drought |
| R-line: HZrr22-T1349–3 | ||||||||
| Oryza sativa | R-line: Huhan 74S | PTGMS | Environment-sensitive male sterility for two-line hybrid breeding | Stable sterility–fertility transition under specific temperature and photoperiod conditions | Photo-thermo sensitive nuclear sterility gene | Liu et al. (2021) | Hybrids showed stronger stress-defense responses | Drought |
| Oryza sativa L. subsp. Nipponbare, Indica | R-line: ospks2–1, ospks2–2, ospks2–3 | MS-mutant | Male sterility induction | Stable sterility expression | Mitochondrial gene | Zou et al. (2018) | Hybrid rice maintained more stable productivity across environments | Drought |
| Oryza sativa L. | M-line: APMS6B | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Madhusudan et al. (2022) | High SCA values supported hybrid yield advantages | Drought & Salinity |
| Oryza sativa | R-line: Peiai 64S, Y58S | PTGMS | Environment-sensitive male sterility for two-line hybrid breeding | Stable sterility–fertility transition under specific temperature and photoperiod conditions | Photo-thermo sensitive nuclear sterility gene | Yang & Zhang (2020) | Stress reduced grain filling, but hybrids maintained better productivity | Heat |
| Padi (Oryza sativa L.). | R-line: Guihui5501 | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in three-line hybrids | Nuclear restorer gene (Rf gene) | Wei et al. (2024) | Heat stress significantly affected spikelet fertility | Heat |
| Oryza sativa | R-line: M.J.5460S, Giza177, Sakha105, Sakha106, GZ7768 | rTGMS (reverse thermo-responsive genic male sterile) | Temperature-dependent male sterility for two-line hybrid breeding | Reversible fertility alteration regulated by temperature changes | Nuclear gene-mediated reverse thermo-sensitive male sterility | Abdelrahman et al. (2021) | Hybrids activated stress-responsive pathways under abiotic stress | Heat |
| Oryza sativa | A-line: IR58025A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | El-mowafi et al. (2021) | Hybrid combinations showed superior agronomic performance | Heat |
| R-line: PR2, IR25571-31R, Giza-Basmati-201, PR1 | ||||||||
| Oryza sativa L. | A-line: CRMS31A, CRMS32A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Bhuyan et al. (2025) | Stress-tolerant hybrids maintained relatively stable productivity | Heat |
| R-line: IR 42266–29-3R | ||||||||
| Oryza sativa | A-line: Jiabuyu | DMS | Facilitation of hybrid breeding through dominant male sterility | Dominant inheritance of male sterility | Dominant nuclear sterility gene | Pang et al. (2017) | CMS-associated genes affected pollen fertility | Heat |
| R-line: 33, 84, 611, 9311, CDR22, Ce253, Cheng-Hui177, Duo-Xi 1 Hao, Fu838, Guang-Hui998, Gui99, Hang 1 Hao, Lu-Hui17, Mian-Hui725, Ming-Hui63, Ming-Hui86, Min-Hui3301, Mi-Yang46, R9308, Shu-Hui527, Wan-Hui057, Yan-Hui559, Yi-Hui1577, Zhe-Hui7954, Zhong-Hui8006 | ||||||||
| Oryza sativa | A-line: wsl3 mutant | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Hong-Bing et al. (2017) | Disruption in reproductive processes contributed to sterility | Heat |
| R-line: Zhenhui 714 (R714) | ||||||||
| Oryza sativa | A-line: IR69625A (Wild abortive (WA) CMSline; G46A (Gambiaca CMS line) | WA-CMS; | Male sterility induction for three-line hybrid breeding; Gambiaca Cytoplasmic male sterility for hybrid breeding | Effective sterility expression in hybrid rice system | Mitochondrial sterility-associated gene with nuclear–cytoplasmic interaction | Hamad et al. (2023) | Hybrid genotypes performed better under stress environments | Heat |
| R-line: Giza 178R | ||||||||
| Oryza sativa | R-line: Zhonghui9308 | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in CMS hybrids | Nuclear restorer gene (Rf gene) | Anis et al. (2018) | Some hybrids showed broad adaptation across environments | P Deficiency |
| M-line: XieqingzaoB | ||||||||
| Oryza sativa | A-line: 36A; 52A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Liao et al. (2019) | CMS-related pathways affected fertility expression | P Deficiency |
| R-line: GXU16, GXU20, GXU28 (WT); GXU16–19, GXU20–8, GXU28–12 | ||||||||
| Oryza sativa L. subsp. indica | A-line: II-32A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Luo et al. (2019) | Fertility abnormalities were linked to disrupted pollen formation | Salinity |
| R-line: F6; BIL627 | ||||||||
| Oryza sativa | A-line: IR69625A; G46 | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Ghazy et al. (2023) | Certain hybrids maintained yield under limited water conditions | Salinity |
| R-line: R146; R727; RH103; R8258; HZ; R938; R4923; R1391 | ||||||||
| M-line: Dexiang074B (074B) | ||||||||
| Oryza sativa | A-line: Dexiang074A | CMS | Male sterility induction | Stable sterility expression | Mitochondrial gene | Li et al. (2023) | Oxidative stress influenced reproductive stability | Salinity |
| R-line: R146; R727; RH103; R8258; HZ, R938; R4923; R1391 | ||||||||
| M-line: Dexiang074B (074B) | ||||||||
| Oryza sativa L. subsp. indica | R-line: KMR3; IL50–13 | Restorer | Pollen fertility restoration | Effective restoration of pollen fertility in CMS hybrids | Nuclear restorer gene (Rf gene) | Thummala et al. (2022) | Hybrid combinations expressed significant heterosis for yield-related traits | Salinity |
Despite these advantages, several constraints continue to limit the long-term sustainability of CMS-based breeding. Narrow cytoplasmic diversity increases vulnerability to environmental and biological stresses, while sterility expression in certain CMS backgrounds remains sensitive to temperature and photoperiod fluctuations (Liu et al., 2021; Zhang et al., 2021; Chen et al., 2021; Lin et al., 2023). Another major challenge is the requirement for compatible nuclear restorer-of-fertility (Rf ) genes to achieve complete fertility recovery (Nabi et al., 2022; You et al., 2023; Wei et al., 2024; Wang et al., 2025). Consequently, recent research has increasingly shifted toward improving the efficiency of fertility restoration, diversifying restorer germplasm, and elucidating the genomic regulation of nucleus–cytoplasm interactions (Anis et al., 2018; Hamad et al., 2023; Nagaraju et al., 2023; Wei et al., 2024; Wang et al., 2025; Niu et al., 2025).
The strategic importance of restorer lines is evident from the repeated use of elite genotypes such as KMR, Guihui5501, Chenghui727, and Zhonghui9308, which exhibit stable fertility restoration and superior performance across multiple CMS backgrounds. However, the repeated deployment of these lines also indicates that current breeding programs still rely on a relatively narrow pool of elite donor germplasm. At the genetic level, fertility restoration is primarily regulated by nuclear-encoded Rf genes that suppress sterility-associated mitochondrial transcripts, representing a clear example of coordinated nucleus–cytoplasm coevolution in crop breeding systems (Anis et al., 2018; Nabi et al., 2022; Nagaraju et al., 2023; Wei et al., 2024; Wang et al., 2025).
To improve breeding efficiency, recent programs increasingly integrate molecular markers, high-throughput genotyping, and genomic selection to rapidly identify and develop superior restorer lines. These advances reflect the broader transition from conventional phenotype-based selection toward genomics-assisted hybrid breeding to enhance stress resilience, fertility stability, and sustainable yield performance under changing environmental conditions.
CMS-based hybrid rice exhibits integrated physiological and molecular adaptations that enhance tolerance to major abiotic stresses, including drought, heat, salinity, nutrient deficiency, and low temperature ( Figure 3). Physiological responses mainly involve improved water-use efficiency, carbohydrate regulation, and antioxidant defense systems (Zou et al., 2018; Liao et al., 2019; Yang & Zhang, 2020; Chen et al., 2021; Ghazy et al., 2023; Li et al., 2023; Wang et al., 2025). Under drought stress, SRL1/SRL2 mutants reduce stomatal density, conductance, and transpiration, thereby minimizing water loss (Liao et al., 2019). Hormonal treatments, such as kinetin and jasmonate, further improve chlorophyll retention, osmotic adjustment, and spikelet hydration under drought and heat stress (Yang & Zhang, 2020; Ghazy et al., 2023).
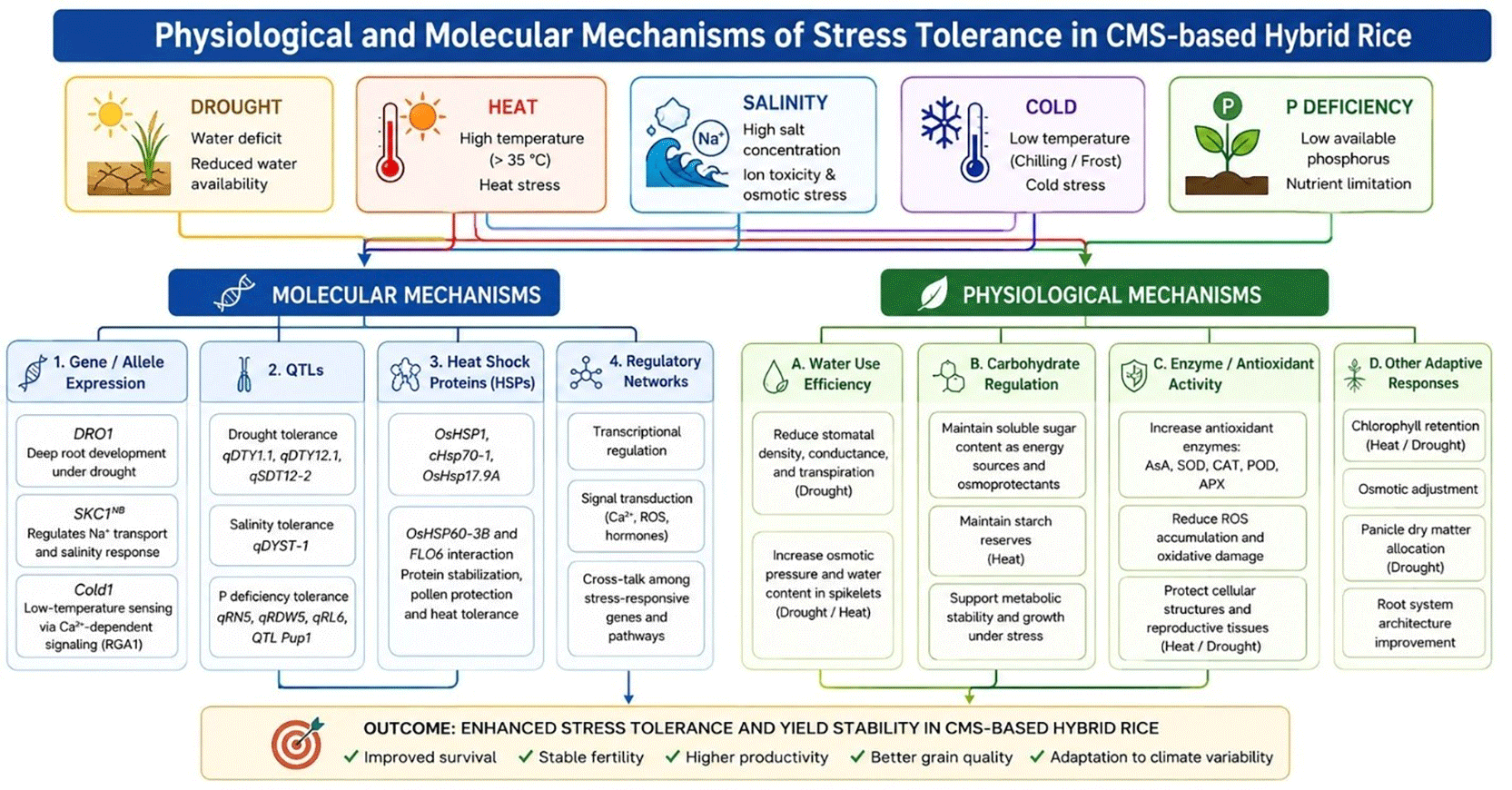
Stress adaptation is also supported by maintenance of carbohydrate homeostasis and enhanced antioxidant activity. The sterile line Dexiang 074A maintained soluble sugar content and panicle dry matter allocation under drought conditions, while the restorer line R996 preserved soluble sugars and starch reserves during heat stress, supporting osmoprotection and metabolic stability (Li et al., 2023; Zhou et al., 2025). Increased activities of ascorbic acid (AsA), superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and ascorbate peroxidase (APX) reduce ROS accumulation and oxidative damage under stress conditions (Liao et al., 2019; You et al., 2023; Wang et al., 2025). Application of methyl jasmonate (MeJA) further strengthens antioxidant defense by enhancing AsA and CAT activity, thereby protecting reproductive tissues from heat-induced injury (Chen et al., 2021).
At the molecular level, stress tolerance is regulated through gene expression and QTL introgression. The DRO1 gene promotes deep root development under drought stress, while qDTY1.1, qDTY12.1, and qSDT12–2 contribute to maintaining plant architecture and yield stability under water limitation (Luo et al., 2019; Nabi et al., 2022; Nagaraju et al., 2023). Heat tolerance is strongly associated with heat shock proteins (HSPs), including OsHSP1, cHsp70–1, OsHsp17.9A, and OsHSP60-3B, which stabilize proteins and reduce oxidative damage, thereby preserving pollen viability under high temperatures (Lin et al., 2023; Zhou et al., 2025).
Under salinity stress, the qDYST-1 locus and SKC1NB allele regulate transcriptional responses and Na + transport to reduce ion toxicity (Ding et al., 2022; Niu et al., 2025). Tolerance to phosphorus deficiency is associated with qRN5, qRDW5, qRL6, and Pup1, which improve root development and phosphorus uptake efficiency (Anis et al., 2018; Madhusudan et al., 2022). Cold tolerance remains comparatively underexplored, although the Cold1 gene has been identified as a regulator of low-temperature sensing through Ca2 + –dependent signaling pathways involving RGA1 (Wei et al., 2024). Overall, these findings demonstrate that modern CMS-based hybrid rice breeding increasingly integrates physiological adaptation with molecular and genomic regulation to enhance stress resilience and yield stability under adverse environments.
CMS-based hybrid rice exhibits coordinated morphological, physiological, and reproductive adaptations to abiotic stress ( Figure 4), with drought and heat stress representing the most extensively studied constraints. Drought tolerance is primarily associated with enhanced root architecture, seminal root length, biomass accumulation, and seedling vigor, which collectively improve water uptake and sustain vegetative growth under limited water availability (Widyastuti et al., 2017; Hong-Bing et al., 2017; Liao et al., 2019; Luo et al., 2019; Nabi et al., 2022; Ghazy et al., 2023). In contrast, heat stress predominantly disrupts reproductive processes, including spikelet fertility, pollen viability, another development, and seed set formation, with spikelet fertility consistently identified as the principal determinant of yield stability under high-temperature conditions (Zhang et al., 2017; Chen et al., 2021; Lin et al., 2023; You et al., 2023; Wei et al., 2024; Zhou et al., 2025). Salinity, phosphorus deficiency, and cold stress remain comparatively less explored but are generally linked to changes in root development, biomass accumulation, seedling vigor, and reproductive performance (Anis et al., 2018; Thummala et al., 2022; Niu et al., 2025).
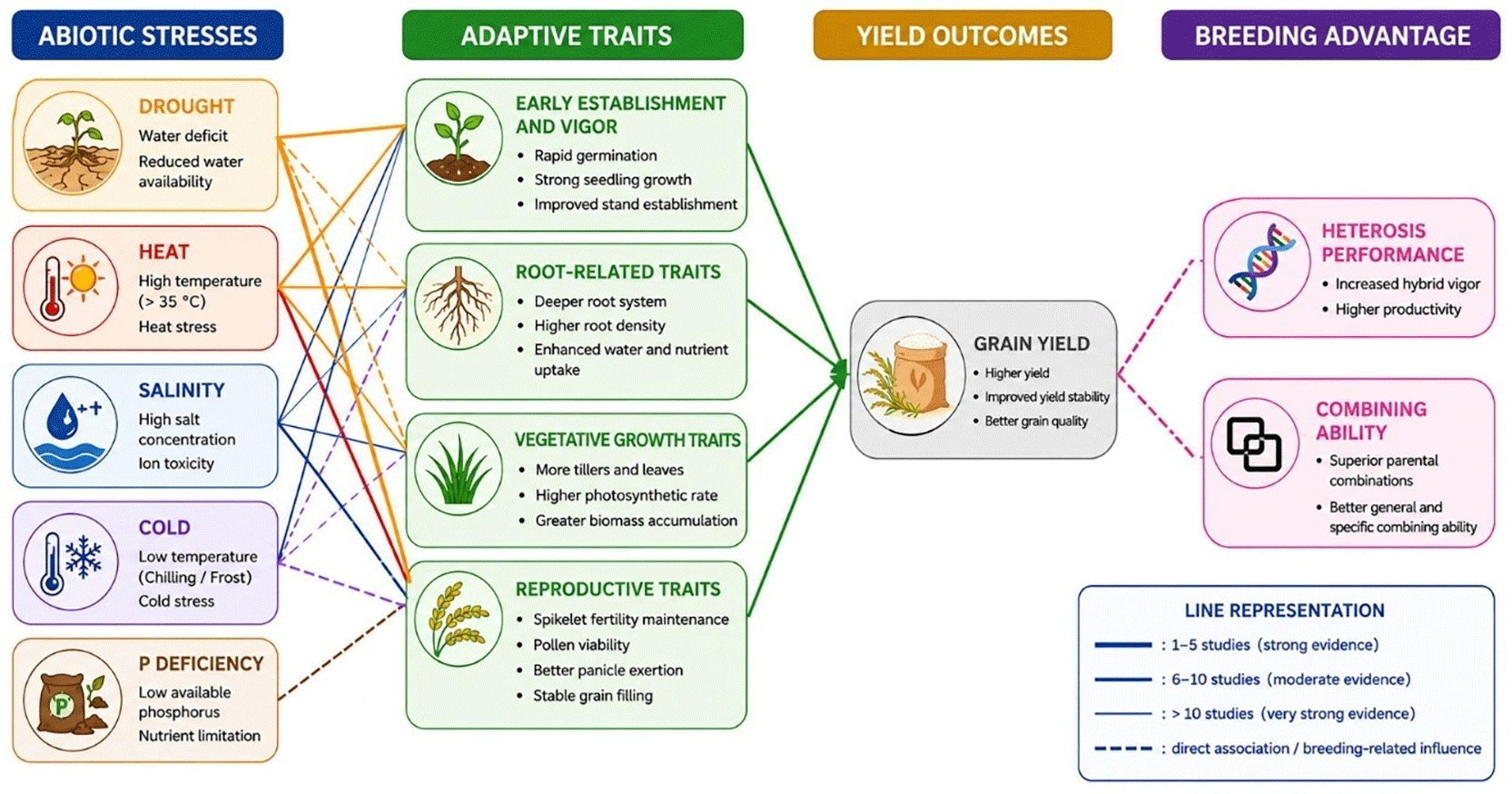
Despite these adaptive responses, physiological tolerance does not always translate into superior productivity, indicating a complex relationship between stress-resilient traits and agronomic performance. Several drought-tolerant hybrids with improved germination and root traits, for example, did not consistently achieve high grain yields, while maintaining reproductive viability under heat stress was sometimes insufficient to sustain seed production (Widyastuti et al., 2017; Chen et al., 2021). In addition, stress-responsive traits are frequently governed by non-additive gene action, reflected by high specific combining ability (SCA) and strong heterotic effects, emphasizing the importance of parental genetic interactions and optimal hybrid combinations in the development of climate-resilient hybrid rice (Widyastuti et al., 2017; Pang et al., 2017).
Based on the reviewed studies, hybrid rice breeding strategies are increasingly directed toward developing abiotic stress-tolerant genotypes while maintaining high yield potential. As shown in Figure 5, the three-line system remains the dominant breeding approach because of its stable male sterility and well-established seed production system. However, its effectiveness is constrained by the limited availability of compatible restorer lines, leading recent breeding efforts to focus on improving both fertility restoration and stress tolerance simultaneously (Chen et al., 2021). Restorer lines, therefore, serve not only as fertility recoverers but also as important sources of adaptive traits under stress conditions (Sravanraju et al., 2024; Wei et al., 2024; Bhuyan et al., 2025). Drought-tolerant restorers were reported to maintain fertility and productivity under water-deficit conditions (Nagaraju et al., 2023), while Guihui5501 combined high yield, grain quality, and cold tolerance (Wei et al., 2024).
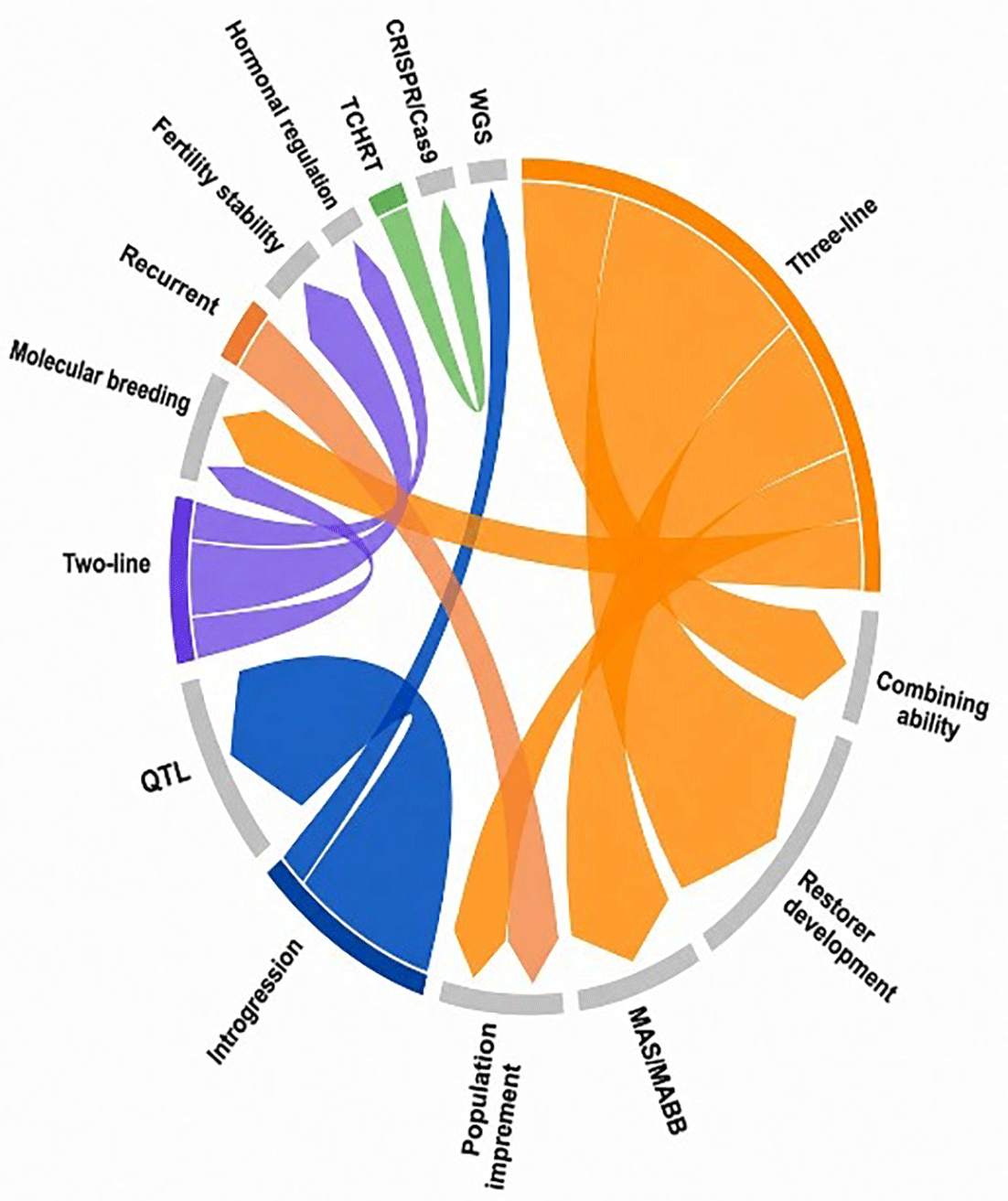
The development of stress-tolerant hybrids is also strongly influenced by parental combining ability. General combining ability (GCA) and specific combining ability (SCA) analyses are widely used to identify superior hybrid combinations under stress environments. Several studies reported strong SCA effects for drought-tolerant hybrids rice (Widyastuti et al., 2017; El-mowafi et al., 2021; Awad-allah et al., 2022), whereas IL50–13 was identified as a promising salinity-tolerant restorer with favorable combining ability for yield traits (Thummala et al., 2022). These findings indicate that non-additive genetic interactions between parental lines largely govern stress tolerance in hybrid rice.
To accelerate breeding efficiency, molecular approaches are increasingly integrated into hybrid rice improvement programs. Marker-assisted selection targeting fertility restoration genes such as Rf3 and Rf4 has been widely applied for identifying stress-tolerant parental lines (Priyadarshi et al., 2025), while QTL introgression remains the primary strategy for improving tolerance to drought, salinity, heat, and phosphorus deficiency (Anis et al., 2018; Luo et al., 2019; Abdelrahman et al., 2021; Bhuyan et al., 2025; Niu et al., 2025). More advanced technologies, including whole-genome sequencing and CRISPR/Cas9-based genome editing, are further enhancing the precision of stress-tolerance improvement and accelerating the development of resilient hybrid rice varieties adapted to changing environmental conditions (Thummala et al., 2022; Sheng et al., 2023).
Although significant progress has been made in developing abiotic stress-tolerant hybrid rice through CMS-based systems, maintaining stable sterility expression and fertility restoration under stress conditions remains a major challenge, particularly under high-temperature stress, which severely disrupts reproductive development and spikelet fertility (Widyastuti et al., 2017; Chen et al., 2021). Most existing studies also focus on single-stress tolerance, whereas combined-stress responses and environmental stability remain insufficiently explored. Pang et al. (2017). demonstrated the potential of DMS-assisted recurrent selection for developing drought- and salinity-tolerant lines, although evaluations were still conducted under limited environmental conditions.
Another key limitation is the weak integration between molecular breeding and agronomic management. Molecular studies largely emphasise QTL mapping, introgression breeding, and candidate gene identification for stress tolerance (Anis et al., 2018; (Thummala et al., 2022), whereas agronomic approaches, such as growth regulator applications, are generally investigated separately (Ghazy et al., 2023; Hamad et al., 2023). This indicates the need for more integrated strategies that combine molecular genetics, plant physiology, and crop management to simultaneously improve stress tolerance, yield stability, and hybrid seed production efficiency.
Future progress in stress-tolerant hybrid rice breeding will likely depend on the utilisation of novel germplasm and advanced genomic technologies. Wild rice species such as Oryza rufipogon provide valuable sources of adaptive alleles for drought and salinity tolerance (Luo et al., 2019; Thummala et al., 2022), while whole-genome sequencing and genome editing offer opportunities for precise identification and transfer of stress-responsive genes. These advances could accelerate the development of climate-resilient hybrid rice that maintains fertility, productivity, and stability under increasingly complex environmental conditions. Accordingly, future research should prioritise multi-location and multi-season evaluations, combined-stress analyses, and the development of environmentally stable sterility systems to support sustainable hybrid rice production under climate change.
The reviewed studies demonstrate that CMS-based hybrid rice breeding remains the most effective and widely adopted strategy for improving rice productivity under abiotic stress conditions. The dominance of WA-CMS and related three-line systems highlights their continued importance in ensuring stable hybrid seed production, efficient fertility restoration, and high yield performance. Advances in physiological, molecular, and genomic research have substantially improved the understanding of stress adaptation mechanisms associated with drought, heat, salinity, nutrient deficiency, and low-temperature tolerance. These mechanisms involve coordinated regulation of water-use efficiency, antioxidant defense, carbohydrate metabolism, root system architecture, and stress-responsive gene expression, collectively contributing to improved resilience and yield stability under adverse environments.
The review further demonstrates that integrating elite CMS lines, superior restorer germplasm, QTL introgression, and genomics-assisted breeding approaches has accelerated the development of stress-tolerant hybrid rice. Increasing application of molecular markers, high-throughput genotyping, whole-genome sequencing, and genome-editing technologies reflects the transition from conventional phenotype-based breeding toward precision breeding frameworks with greater selection efficiency and predictive accuracy. In particular, the strategic role of restorer lines and non-additive genetic effects emphasizes the importance of parental interactions in maximizing heterosis and environmental adaptability in hybrid rice systems.
Despite these advances, important limitations remain, including the narrow genetic diversity of CMS and restorer lines, instability of sterility expression under environmental fluctuations, and the limited evaluation of combined-stress responses across diverse field conditions. Therefore, future hybrid rice breeding should prioritize the development of multi-stress-tolerant genotypes, environmentally stable sterility systems, and broader germplasm utilization integrated with advanced genomic technologies. Such approaches will be essential for developing climate-resilient hybrid rice that maintains fertility, productivity, and yield stability under increasingly unpredictable environmental conditions and future climate change scenarios.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)