Keywords
genetic algorithms, mutation rate, artificial intelligence, bioinformatics, genomics
genetic algorithms, mutation rate, artificial intelligence, bioinformatics, genomics
The title was modified to avoid giving a false sense of the subject matter of the study. The abstract was modified to focus on the computational findings rather than the applications. Implications mentioned in the Discussion were expanded to elaborate on potential uses for our algorithm in bioinformatics, among other fields. Additional references were added based on the new information.
See the authors' detailed response to the review by Panayiotis Benos
See the authors' detailed response to the review by Mark A. Ragan
See the authors' detailed response to the review by Christos Ouzounis
The last few years have seen an exponential increase in the size of databases, especially those in genetics, which catalog the basis of various diseases. Computing power has not kept pace with this rapid increase in bioinformatics. Therefore, it has become critical to develop clever algorithms to reduce the time needed to search such databases and arrive at solutions to the treatment of genetically determined diseases1. Genetic algorithms provide new hope to overcome this dilemma. Genetic algorithms attempt to copy the principle of “survival of the fittest”, using Darwin’s theory of evolution to find a satisfactory solution. In essence, a group of random solutions is created and ranked, after which the best solutions are allowed to “interbreed”. During interbreeding, small mutations are allowed to add an element of randomness, and in this way aid the genetic algorithm in finding the optimal solution2.
The applications of genetic algorithms (GAs) are well known and far ranging3. Because of their versatility, GAs have been widely and successfully used to optimize practical problems such as determining a long sequence of base pairs in a genetic database, scheduling drilling operations in a circuit-board factory, and data mining, among others4. However, one major problem faced by GAs is premature convergence, in which the algorithm becomes trapped in a local optimum and is unable to find the global optimum5.
This study focuses on the use of mutations in GAs. Normally, the mutations occur at a constant rate, known as the mutation rate. However, use of a fixed mutation rate can produce sub-optimal solutions5. We propose a new, variable mutation rate that uses a pseudoderivative to take into account the time that a GA can be stuck at a certain point. The longer the algorithm has been stuck at a local optimum, the more likely it is that a mutation will occur. This addition of a greater element of randomness to the algorithm allows it to move from the local optimum and look for better solutions. In this paper, we show that the proposed variable mutation rate outperforms fixed mutation rates and other common search algorithms in the effectiveness of their solutions.
Many real-life problems can be modeled as continuous, nonlinear optimization problems. Within a given search space S on the optimization function f, a global (absolute) optimum is sought. This may take the form of a global maximum or minimum, depending on the original problem. A typical 3-dimensional global optimization problem follows the form:
The challenge in solving a global optimization problem is in seeking the global optimum rather than becoming trapped in a local optimum, an issue that will be addressed in more detail later6. These optimization problems can be approached with a variety of techniques. One popular technique is the use of GAs, the focus of this study7.
GAs are population-based optimization techniques favored for their properties of self-learning, self-organization, self-adaptation, and implicit parallelism8. Based on the principles of Charles Darwin’s natural selection and meiotic cell division, GAs involve several components: a population, a measure of fitness, and a method of breeding2. The population forms the basis for the GA and is made up of many individuals, often called chromosomes. Over time, “chromosomes” breed with other “chromosomes” to form “children” that make up a new generation in the GA9.
Chromosomes can be made up of binary strings or of real values. Binary-coded GAs (BCGAs) have chromosomes of 0s and 1s. While BCGAs are the more traditional method and are adequate for small- to moderate-size optimization problems, they fail for high-dimensional problems because they require more computational time and memory. Real-coded GAs (RCGAs) utilize real values that allow for both improved computational time and memory as compared to BCGAs, making the optimization of multi-dimensional and high-precision problems more feasible5. Each chromosome (binary-coded or real-coded) has a certain fitness value or f value derived from its binary string or its real values, where the f value of a chromosome refers to its value when inputted into the function described in (1). The calculations for the f value of a chromosome vary by problem. In this respect, each chromosome represents a single solution to the optimization problem10. Following the principles of natural selection, chromosomes with higher fitness values give rise to children with high fitness values, so the GA “selects” for fitter chromosomes by giving them a higher probability of breeding and passing on their genes11.
The breeding stage involves two processes: crossover and mutation. In true meiotic cell division (the process by which a sperm or an egg cell is created), a crossover occurs when two chromosomes pair with each other and exchange portions of their length, forming two hybrid chromosomes. Similarly, chromosomes in a GA exchange values12. A simple example of crossover is illustrated in Figure 1.
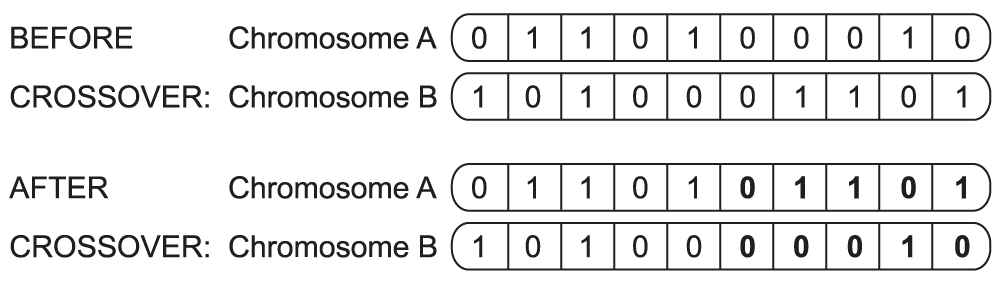
An example of crossover between a set of two binary-coded chromosomes where half the length of chromosome A is exchanged for half the length of chromosome B, resulting in a hybrid set of chromosomes.
The second operator of the breeding stage is mutation. Mutation is a naturally-occurring phenomenon that may come into play during any replication of chromosomes. Incorporation of mutation into a GA may be considered an optional step, but has generally been found to increase the diversity of the population13. This allows GAs to explore promising areas of the search space8. The major advantage of implementing mutations in a GA is in avoiding premature convergence, in which the GA becomes trapped in a local optimum14. However, the mutation operator has trade-offs in slowing down the learning process5. Mutations have in the past taken the form of point, frame-shift, and translocation mutations, all of which involve swapping, switching, or shifting binary bits, in the case of binary-coded chromosomes15. Random mutation has also been proposed, in which a gene is replaced with a random value within certain lower and upper bounds16, and may provide interesting insight into the mechanisms of mutation, since a small mutation in nature may cause a gene to take on a vastly different role17. Finally, the rate of mutation, rm, can play a key role in the effectiveness of a given GA7.
A variety of modifications have been made to GAs in the breeding stage. A large amount of attention has been devoted to developing improvements in crossover operators, but there have been fewer studies in improving mutation operators, which make up a very promising although largely unexplored branch5,15. Several studies have implemented adaptive mutation, or a changing mutation rate18,19. Adaptive mutation methods have ranged from individual gene-based mutations to Gaussian mutation operators based on the mean and the standard deviation of the Gaussian distribution20. This study proposes a novel adaptive mutation approach and applies it to two tests: the well-known N-queens problem and the maximization of a 3-dimensional function. This study also compares a GA implementing an adaptive mutation operator with other well-known search algorithms, such as the Nelder-Mead method, hill climbing technique, and random search.
Numerous other search algorithms besides GAs exist for solving similar optimization problems21. A basic method is the random search method whereby solutions are randomly chosen and evaluated for a certain amount of time, and the best solution found during the time span is returned22. A more advanced method implements the hill climbing technique. This method begins at a random point and uses a greedy algorithm to move to the best immediate neighboring solution until no neighboring solution is better than the current solution23. The final, and most robust, non-GA search algorithm being examined in this study is the Nelder-Mead algorithm. The Nelder-Mead method is a heuristic search that relies on approximating a local optimum for a problem with N variables by using a simplex (a polytope consisting of N + 1 vertices)24.
The N-queens problem is an interesting application for testing the accuracy of genetic algorithms due to the inherent difficulty of optimizing the problem under efficient time and memory constraints25. Given an N × N board, find a set of N-queens such that zero pairs of queens are in the same row, column, or diagonal (none of the queens are attacking each other). Although there are several approaches (brute force/recursion, simulated annealing, etc.) to solving this optimal placement problem26, the N-queens problem was chosen in this study as an early practical test of the proposed new mutation rate operator’s accuracy.
Finding the maximum or minimum value of a 3-dimensional surface is a more abstract but more visual problem. A surface with multiple local maxima and minima can simultaneously test a GA’s ability to avoid premature convergence and its ability to handle multidimensional optimization problems. Because GAs solve problems with implicit functions of N-dimensions, it is important that the newly proposed mutation operator can deliver reasonable speed and accuracy in the most primal form of the N-dimensional problem, optimizing a space function. While the N-queens problem is adequate for testing the accuracy of a GA, it is not adequate for testing its speed of convergence because there are multiple solutions. Therefore, maximization of a 3-dimensional surface with multiple local maxima but only one absolute maximum was chosen as a practical test of the new mutation operator’s speed of convergence, or number of generations needed to converge to the optimal solution. Furthermore, minimization of the Rastrigin function was chosen as a simple method for comparing accuracies of various search algorithms. The Rastrigin function is an optimal function to choose for such a comparison as the surface holds a myriad of local minima, but only one global minimum at f (0,0) = 0. The Rastrigin function models a difficult problem to optimize, where a non-robust search algorithm may easily become caught in a local optimum27.
The research question this study attempted to answer was: what are the effects of an adaptive mutation rate, based on the derivative of the fitness function with respect to generations, on the efficiency and accuracy of a GA? We performed three experiments to answer this question. The first experiment in this study tested the null hypothesis that given a problem, which is not always solvable (to the global optimum) by a fixed-rate GA, a GA with an adaptive mutation rate (based on the derivative of the fitness function with respect to generations) would find an optimal solution significantly more often than one with a constant mutation rate. To reject the null hypothesis, we will need to show that an adaptive mutation rate can find an optimal solution significantly more often than one with a fixed mutation rate. The second experiment tested the null hypothesis that given a problem, which is always solvable (to the global optimum) by a constant mutation rate GA, there would be no significant difference in the efficiency between the constant mutation rate GA and a GA with an adaptive mutation rate, based on the derivative of the fitness function with respect to generation. The third experiment tested the null hypothesis that a GA with an adaptive mutation rate is not significantly more accurate in converging to a global optimum of a 3-dimensional function as compared to other search algorithms (i.e. Nelder-Mead, hill climbing, random).
The mutation rate function rm was the independent variable in the first study. Accuracy (how often a GA finds the optimum solution) measured in percent (%) with an error value of ±0.001, and efficiency (how long it takes to converge to the solution), measured in generations, were the dependent variables. The experiment was set up with three levels: a constant rm of 0% (no mutation, control), a constant rm of 20% (control), and an adaptive rm derived from a pseudoderivative and a sigmoid function. The independent variable of the second study was the specific search algorithm being run. The accuracy, measured in percent (%) with an error value of ±0.001, was the dependent variable. The experiment was set up with four levels (search algorithms): a random search, a search implementing the hill climbing technique, a search implementing the Nelder-Mead method, and a search implementing the proposed adaptive GA.
The adaptive mutation rate operator was tested on three problems: the N-queens problem, the maximization of a 3-dimensional surface, and the minimization of a different 3-dimensional surface (the Rastrigin function). The N-queens problem was used specifically to test the accuracy of the new adaptive mutation rate since a GA implementing a constant mutation rate does not have 100% accuracy in the N-queens problem. The maximization of a 3-dimensional surface problem was used to test the efficiency of the new adaptive mutation rate since a GA implementing the 20% constant mutation rate does have 100% accuracy in the 3-dimensional maximization problem. The minimization of the Rastrigin function was used to determine the robustness of the proposed adaptive GA compared to other well-known search algorithms. The Rastrigin function was selected due to a most diverse topography, including a myriad of local optima by which weak search algorithms may be forced to prematurely converge. In the first study, accuracy was defined to be the rate of successful optimization within 100,000 generations to the nearest ±0.001, in order to save computing power. Python 2.5 was used to create and run GAs for the N-queens and 3-dimensional maximization problems. Python 3.0 was used to create and run an adaptive GA, Nelder-Mead, hill climbing, and random search algorithms for the Rastrigin function minimization problem. A vector class was created to aid in simplifying the code for the latter experiment. Both GAs reported mutation rate, convergence, and best f values for later analysis. For both problems, the mutation rate operator function was a sigmoid function fit to the domain x in [0, ∞) and range y in [0, 1]. The sigmoid function is defined as:
with domain x in (-∞, ∞), range y in [0, 1], and sigmoid(0) = 0.5. The function used was thus a fit of the monotonic sigmoid function to the desired domain and range, resulting in
where x is given by:
with gc representing the current generation and go representing the oldest generation of the same best f value. x thus is inversely related to the derivative of the fitness function with respect to generation. This causes an increase in mutation rate when the fitness between generations is stagnant. A theoretical basis and further details of the adaptive mutation rate operator is included in the Theory section under the Discussion.
For the N-queens problem, the specific case of N = 8 was chosen. The fitness function was chosen for this problem to be the number of pairs of queens violating the problem specification. A program was written to run 200 trials of GAs maxed at 100,000 generations or until a solution was found (f = 0). The genetic code for each solution in the solution set was a sequence of eight integers, with each index representing a column and each value representing a row. Crossover locations were determined using random integers. All variables were held constant apart from mutation rates, which varied between constant mutation at 0% (no mutation), constant mutation rate of 20%, and the sigmoidally-determined adaptive mutation. For the full code, see Script 1 in the Supplementary materials.
Regarding the maximization of a 3-dimensional function, the proposed sigmoidal mutation operator was again compared to controls of a constant mutation rate of 20% and of no mutation (0%). The function chosen for maximization was
which represents a function with several local maxima and minima, which can be solved by the 20% mutation rate GA but not the 0% mutation rate GA. This allowed for a test of efficiency of the new sigmoidally-determined adaptation mutation rate GA against the 20% fixed mutation rate GA and a test of accuracy for the sigmoidally-determined adaptation mutation rate GA against the 0% fixed mutation rate (no mutation) GA. A graphical representation of the function is shown as Figure 2. The genetic code for each solution in the solution set for this problem was a sequence of two integers, the x and y coordinates. The chromosomes were implemented as real-coded chromosomes containing the actual coordinates rather than binary strings. For crossover, the x coordinate of one solution was chosen and the y coordinate of another solution was chosen. For mutation, a coordinate was replaced by a random number within the domain of the problem. Once again, all variables, except for mutation rates, were held constant throughout each experiment.
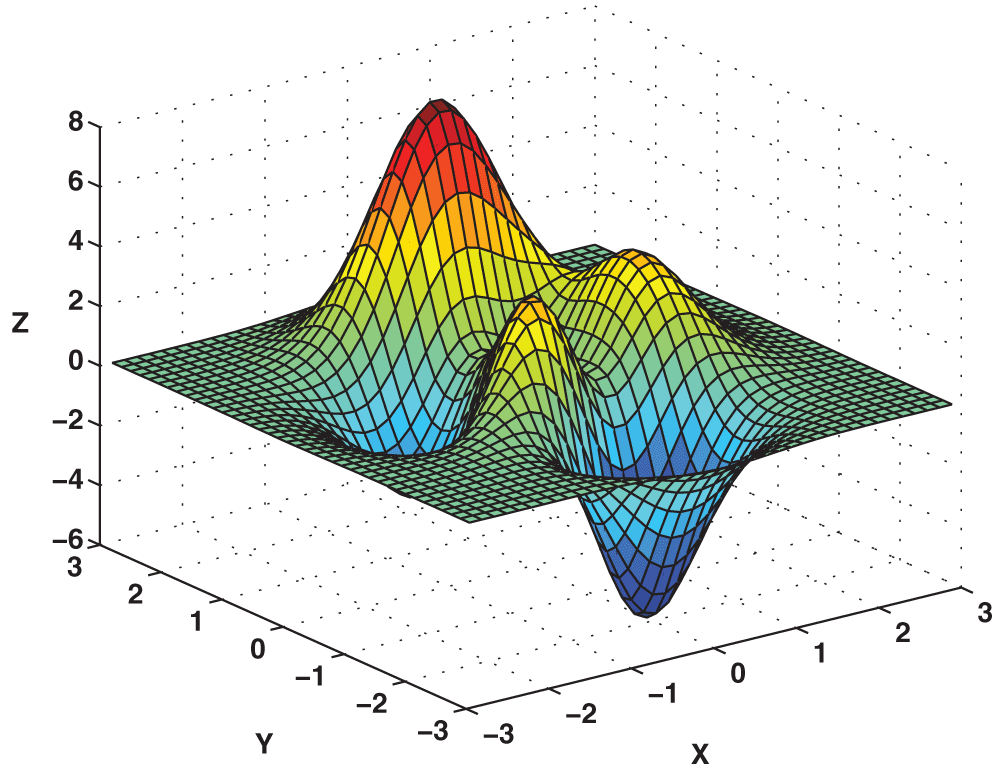
Visualized in MATLAB 6.5 Using the Command peaks(40). The three maximums of the surface were located at f = 8.1165 (global maximum), 3.5507, and 3.4652.
The minimization of the Rastrigin function, given by:
was the final test run, with a goal to learn how the proposed adaptive GA compares to non-GA search algorithms. All of the search algorithms were coded to report the run time to converge and the result of the respective convergence. In order for a result to be considered accurate, the returned answer had to be within 0.001 of the true answer. This ensures that the algorithm is converging to the correct minimum and also aids in simulating a difficult optimization problem. Since the random method does not converge, but rather runs for a given time span, the average run time for each GA trial was obtained and used for the run time of the random method search. This allows for direct comparison of the accuracies of the random method and the adaptive GA, as the run time for each method was identical, and thus, held constant. Each algorithm was run for 1,000 trials in order to obtain enough data for a robust analysis. For the full code, see Scripts 2, 3, and 4 in the Supplementary materials. A graphical representation of the Rastrigin function can be found as Figure 3 and Figure 4.
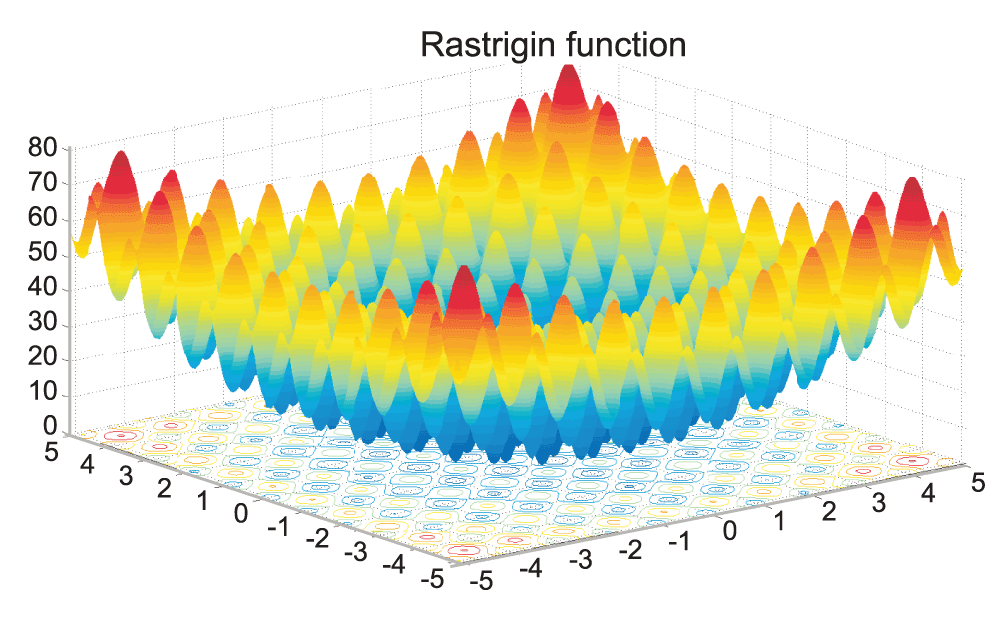
Visualized in MATLAB 6.5: f = @(x,y) 10*2 + x.^2 + y.^2 – 10*cos(2*pi*x) – 10*cos(2*pi*y). The global minimum of the Rastrigin function is f(0,0) = 0.
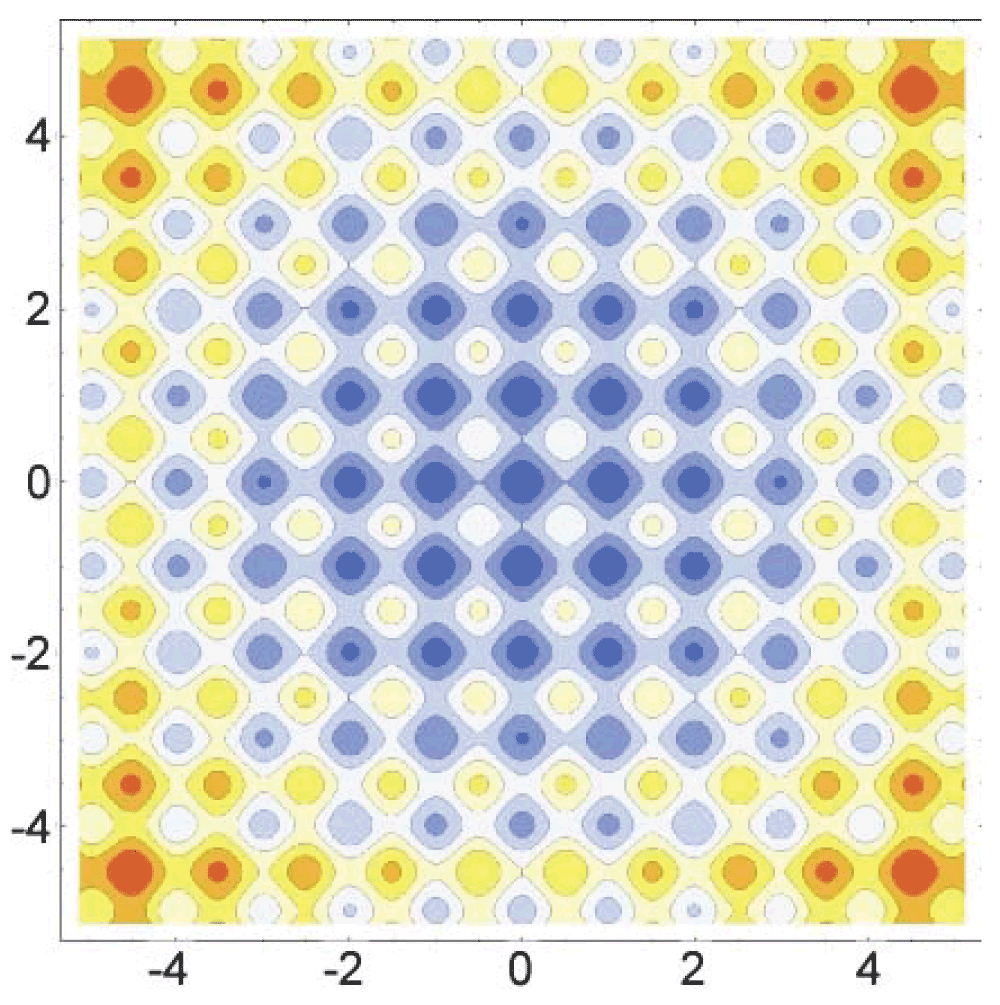
Visualized in Mathematica 8: ContourPlot[10*2 + x^2 + y^2 – 10 Cos[2 Pi x] – 10 Cos[2 Pi y], {x, -5.12, 5.12}, {y, -5.12, 5.12}, ImageSize -> 1000, Axes -> False, ColorFunction -> ColorData["TemperatureMap"]].
The results of experimentation are summarized in Table 1, Table 2, Table 5, and Table 7. In the N-queens problem, the proposed sigmoid mutation operator displayed both the fastest convergence and greatest accuracy with convergence after an average of 25,455.4 generations and an accuracy of 95.5%. In the 3-dimensional surface maximization problem, there was no significant difference between the speeds of the successful GAs. Accuracies were tied between 20% constant and adaptive mutation, as both techniques consistently identified the global maximum within the 100,000-generation limit. In the Rastrigin function minimization problem, the proposed sigmoid mutation operator GA performed significantly better than the three other search algorithms in terms of accurately converging to the global minimum. See Figure 5–Figure 7 for a summary of the results and significant differences.
| Constant (0%) mutation | Constant (20%) mutation | Adaptive mutation | |
|---|---|---|---|
| Overall accuracy (%) | 0 | 53.5 | 95.5 |
|
Average convergence (generations) | 100000 | 50661.49 | 25455.4 |
| Constant (0%) mutation | Constant (20%) mutation | Adaptive mutation | |
|---|---|---|---|
| Overall accuracy (%) | 0 | 100 | 100 |
|
Average convergence (generations) | 100000 | 67460.99 | 67779.92 |
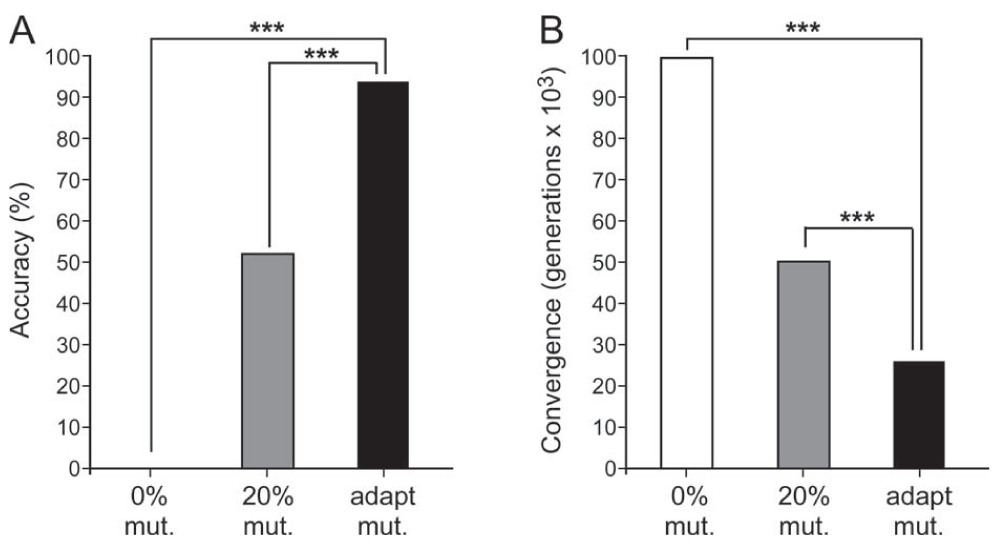
A. Graph of the N-queens problem accuracies among 0% constant mutation, 20% constant mutation, and adaptive mutation. ***p < 0.001. B. Graph of the N-queens problem convergences among 0% constant mutation, 20% constant mutation, and adaptive mutation. ***p < 0.001. mutation (mut.), adaptive (adapt.).
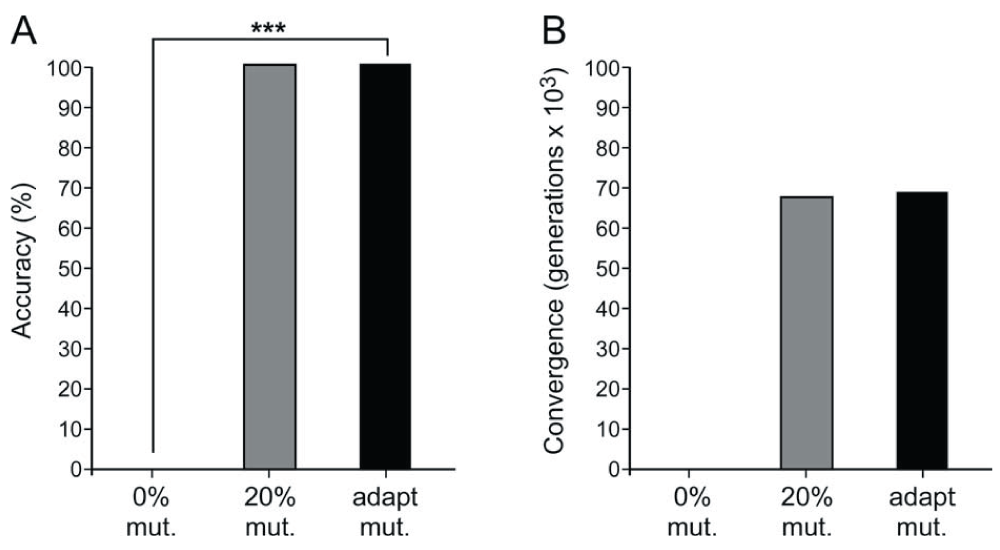
A. This is a graph of the 3D surface maximization problem accuracies among 0% constant mutation, 20% constant mutation, and adaptive mutation. B. This is a graph of the 3D surface maximization problem convergences among 20% constant mutation and adaptive mutation. 0% constant mutation was not statistically analyzed because convergence was not achieved. ***p < 0.001. mutation (mut.), adaptive (adapt.).
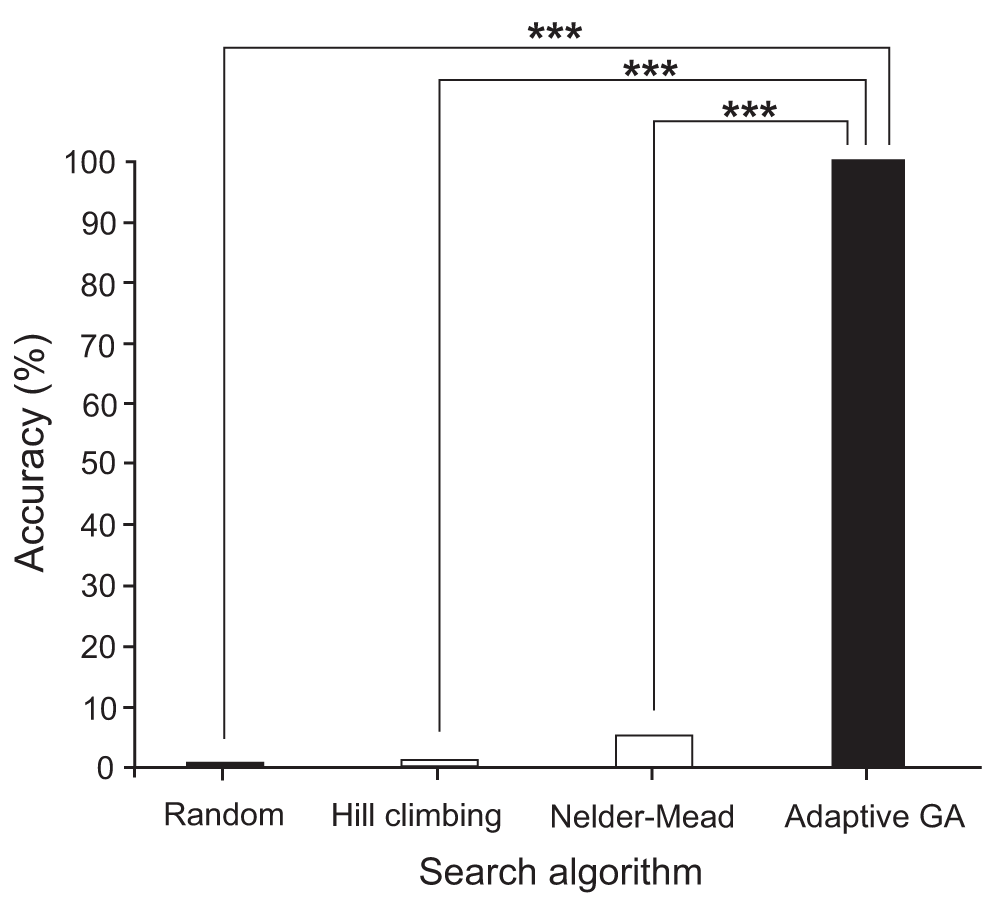
Graph of the Rastrigin function minimization problem accuracies among random, Hill climbing, Nelder-Mead, and adaptive GA methods. ***p < 0.001. genetic algorithm (GA).
In the N-queens problem, a series of unpaired, single-tailed Student’s t-tests were used to test the following null hypothesis at the 0.05 level of significance: the accuracy of the adaptive mutation operator will not be significantly higher than that of 20% constant mutation or that of 0% constant mutation. The null hypothesis was rejected for the 20% constant mutation rate (p = 6.108 × 10-25 < 0.05) and the 0% constant mutation rate (p = 2.572 × 10-214 < 0.05). See Table 3 for a summary of the N-queens statistics.
In the surface maximization problem, a series of unpaired, single-tailed Student’s t-tests were used to test the following null hypothesis at the 0.05 level of significance: the efficiency of the 20% constant mutation rate GA will be significantly higher than that of the adaptive mutation operator GA. The null hypothesis was rejected (p = 0.448 > 0.05) for the 20% level. Because the constant 0% mutation rate GA does not find the global maximum, it was compared to the adaptive mutation operator on the basis of null hypothesis 1 at the 0.05 level of significance: the accuracy of the adaptive mutation operator will not be significantly higher than that of 0% constant mutation rate. The null hypothesis was rejected (p = 0 < 0.05). See Table 4 for a summary of the surface maximization statistics.
In the Rastrigin function minimization problem, a series of unpaired, single-tailed Student’s t-tests were used to test the following null hypothesis at the 0.05 level of significance: the accuracy of the adaptive mutation operator GA will not be significantly higher than those of the other search methods (Nelder-Mead, hill climbing, random). The null hypothesis was rejected for random method (p = 2.53 × 10-223 < 0.05), for the hill climbing method (p = 9.61 × 10-268 < 0.05), and for the Nelder-Mead method (p = 3.04 × 10-152 < 0.05). Furthermore, a series of unpaired, single-tailed Student’s t-tests were used to test if there were significant differences in the run times of the hill climbing and Nelder-Mead methods when the algorithm accurately converged and when the same algorithm did not accurately converge. Both the hill climbing method (p = 0.140 > 0.05) and the Nelder-Mead method (p = 0.576 > 0.05) showed no significant differences in run time when accurately converging compared to inaccurately converging. These results point to the possibility that these two methods were only accurate when a fortunate random point was selected to start running the search algorithm from. No extra time being needed to converge correctly versus incorrectly, points towards the idea that no extra computation was needed either. See Table 6 and Table 7 for a summary of the Rastrigin function minimization statistics.
| Convergence | Hill climbing | Nelder-Mead | |
|---|---|---|---|
| Run time (s) | Accurate | 0.65205 | 0.01852 |
| Inaccurate | 0.59429 | 0.01830 | |
| Trials | Accurate | 35 | 57 |
| Inaccurate | 4965 | 943 | |
| p-value | 0.13960 | 0.57576 |
The mutation rate operator presented in this study uses randomness to guide a genetic algorithm (GA) towards the optimal solution. A GA with less randomness leads to faster convergence towards local optima; however, by limiting randomness it limits the search space, which in turns hinders the search for the global optimum. Conversely, a GA with more randomness hinders progress towards local optima, but allows for a wider search space, aiding the search for the global optimum8,28. Therefore, randomness should be inversely related to the derivative of the fitness function f. Because f is implicit in most problems, the proposed heuristic attempted to look at the derivative of f with respect to generation, creating a pseudo, inversely related function to the derivative that could be used to calculate an adaptive mutation rate. The heuristic subtracted the first generation at which the most optimal fitness value appeared from the current generation to obtain a result inversely related to the derivative, and thus directly related to the randomness or mutation rate. Therefore any monotonically increasing function with domain bounded by x in [0, ∞) and y in [0, 1] should provide an appropriate amount of randomness to either find a local optimum or increase the search space from the local optima in order to find the global optimum.
The implications of a new, pseudoderivative-based adaptive mutation rate are considerable. The implementation of this new technique within a genetic algorithm could provide increased accuracy in optimization of all GA problems. For example, the treatment of complex diseases requires the discovery of new drug combinations that are hard to come by solely on the basis of empirical clinical knowledge. Search algorithms, including GAs, have begun to provide promising results in identifying optimal drug combinations, e.g. for destroying human cancer cells as well as for minimizing the physiological decline associated with aging. Moreover, these approaches required only one-third of the number of tests employed in the classic method for the discovery of optimal drug combinations. This approach has greatly reduced the risk and expense of clinical trials29. The availability of a more sophisticated class of GAs will also allow search algorithms that are based on GAs to be improved. For example, swarm-based optimization algorithms, such as the Bees Algorithm, can improve the efficiency with which optimal and suboptimal solutions can be discovered within a given search space30.
Our results agree with results reported in other articles on adaptive mutation accuracy and efficiency5,31. The method described in this study also has the added benefit of low computational complexity while still being able to guide the GA out of local optima and towards the global optimum. Studies in adaptive mutation have ranged widely from individual gene-based mutations, to Gaussian operators, to polynomial operators20. However, to our knowledge, past adaptive mutation techniques have never used the derivative of f with respect to generation to determine a changing mutation rate. This new method of implementing adaptive mutation may open up entirely new areas for implementing search algorithms based on improved GAs.
GAs are powerful tools that can optimize overwhelmingly complex real-life problems, including speedy diagnoses of complex diseases. Previous GAs faced a trade-off between speed and accuracy, as more random GAs sacrificed speed in exchange for a better chance of optimization or less random ones gave up accuracy for speed. The solution of an adaptive mutation operator based on the derivative of f with respect to generation allows for increased accuracy without the loss of speed. Suggestions for research that would expand upon current findings include determining an optimal monotonically increasing function for the mutation operator, such as a sigmoid, inverse tangent, or scaled linear function, as well as comparing the current GA to other commonly used search methods in current bioinformatics problems.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)