Keywords
Chasmosaurinae, Dinosaur Park Formation, Campanian
Chasmosaurinae, Dinosaur Park Formation, Campanian
AMNH, American Museum of Natural History, New York, New York; CMN, Canadian Museum of Nature, Ottawa, Ontario; NHMUK, Natural History Museum, London; ROM, Royal Ontario Museum, Toronto, Ontario; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta; UALVP, University of Alberta Laboratory for Vertebrate Paleontology; YPM, Yale Peabody Museum, New Haven, Connecticut.
The Chasmosaurinae is a diverse group of horned dinosaurs that flourished during the Late Cretaceous of western North America1,2. Chasmosaurines were especially diverse in the late Campanian. Different species are found up and down the coast of the Western Interior Seaway1,3–6, and multiple species frequently occur in a single habitat. This diversity and a rich fossil record make chasmosaurines an ideal system for studying dinosaur diversity and biogeography.
Among the best-known genera is the Chasmosaurus from the Dinosaur Park Formation (DPF) of Alberta. Chasmosaurus is known from numerous skulls and skeletons7–12 but despite abundant material there is little agreement about how many species, or even genera are present4–8,10,13.
Until recently, Chasmosaurus was split into three species: Chasmosaurus belli, Chasmosaurus russelli, and Chasmosaurus irvinensis7. The three succeed each other in stratigraphic section, with primitive species occurring at the base of the DPF and more derived forms appearing higher up7, suggesting a lineage evolving through time. Recently, however, it has been proposed that C. irvinensis represents a distinct lineage, and it was placed in a new genus, Vagaceratops4.
Here, a skull from the Yale Peabody Museum of Natural History, YPM 2016, is described. Until now, this specimen has only received cursory description11 but its unique morphology may help sort out the systematics of Chasmosaurus. YPM 2016 has previously been referred to C. belli5,7,8,10,11, but closer examination shows apomorphies that are absent in other specimens of this species. These features include five pairs of epiparietals, widely divergent lateral rami of the parietal caudal bar, and a caudally inclined premaxillary narial strut. These characters occur, however, in the highly derived C. irvinensis7.
This paper uses YPM 2016 as a starting point to examine Chasmosaurus’ systematics. Here, a revised taxonomy is proposed. Rather than starting with a taxonomy and then identifying diagnostic characters, this paper uses a bottom-up approach to taxonomy. Specimen-level phylogenetic analysis was conducted first, then phylogeny is used as a framework to delimit and diagnose species. Specimen-level phylogenies have previously been used to examine the systematics of other groups, including apatosaurine sauropods14, thescelosaurid ornithopods15, and the leptoceratopsid Montanoceratops16. Most recently, Scannella et al.17 have used specimen-level phylogenetic analysis to examine the systematics of Triceratops, and concluded that Triceratops prorsus represented the direct descendant of Triceratops horridus. This study represents the first application of this approach to Chasmosaurus. First, however, a history of the genus is presented.
The genus Chasmosaurus has a complex taxonomic history. Lambe12 originally described Monoclonius belli on the basis of a partial parietal (CMN 491). This species would then be referred to Hatcher et al.18 to the now defunct Ceratops, as Ceratops belli. Later, with more complete fossils, Lambe showed that the animal was distinct from previously known ceratopsids and created a new genus, Protorosaurus19. After Protorosaurus proved to be preoccupied, Lambe proposed Chasmosaurus20 as a replacement. And so, within the space of a dozen years, the species bore four different genus names. Despite being fragmentary, the holotype is diagnostic, as the morphology of the posterior bar of the parietal is unique to this species5.
Lambe named a second species in the 1902 paper, Monoclonius canadensis, which was later renamed Eoceratops canadensis21. This species was later placed in Chasmosaurus, as Chasmosaurus canadensis22,23. As discussed below, it appears to be distinct from Chasmosaurus, but the type is nondiagnostic.
New specimens led to new species. In 1933, Lull described a skull (ROM 839) as Chasmosaurus brevirostris11. The specimen has an unusually short rostrum, but this feature is typical of juvenile chasmosaurs24 and other features of C. brevirostris, including unfused epoccipitals and open cranial sutures24–26 indicate immaturity. As differences between C. brevirostris and C. belli likely reflect ontogeny, C. brevirostris is considered a junior synonym of C. belli8,10,22.
That year, another skull, AMNH 5401, was described as Chasmosaurus kaiseni27. AMNH 5401 has traditionally been referred to C. belli8,10, but the resemblance of the frill to that of C. belli is the result of plaster reconstruction. As discussed below, the skull appears to be distinct from Chasmosaurus and has been referred to a new genus, Mojoceratops5.
Sternberg9 named a fourth species, C. russelli. The type (CMN 8800) consists of a crushed skull. Sternberg interpreted the frill as having a V-shaped caudal margin; based on this and other characters, he regarded CMN 8800 as a distinct species. Sternberg referred three more specimens to C. russelli, with additional specimens referred by Godfrey and Holmes8, including CMN 2280, a largely complete skeleton. C. russelli has been regarded as a valid species by all recent studies1,4,5,7,8,10,13.
In 1989, a fifth species, Chasmosaurus mariscalensis, was named from bonebed material from the upper Campanian Aguja Formation of Texas22. Phylogenetic analyses fail to group this species and Chasmosaurus to the exclusion of other chasmosaurs3–6,28–30 however, and so it was referred to a separate genus, as Agujaceratops mariscalensis31.
In 2001, a sixth species, C. irvinensis, was described7. C. irvinensis is characterized by absence of brow horns, a straight posterior margin of the frill, and five pairs of epiparietals7 that fuse at their bases and curl forward. Holmes et al. interpreted C. irvinensis as closely related to C. belli7. The animal also occurs above C. belli in section7, raising the possibility that C. irvinensis descends from C. belli.
Placement of this species within Chasmosaurus has been contested, however. A species-level analysis by Sampson et al.4 recovered C. irvinensis as sister to Kosmoceratops richardsoni, an arrangement recovered in subsequent analyses using variants of the same character-taxon matrix13,29. Because this analysis placed C. irvinensis outside of the other species of Chasmosaurus, the animal was put in a new genus, as Vagaceratops irvinensis4. The phylogeny recovered appears well-supported in light of the similarity between the frills of C. irvinensis and Kosmoceratops. In each, the frill’s posterior margin bears ten epoccipitals, with the four middle pairs coalesced and hooked forward. Yet the frill as described by Holmes et al.7 differs from that of Kosmoceratops4 in its construction. In C. irvinensis, the back of the frill is formed by the parietal, which bears five epiparietals (P1–P5). In Kosmoceratops, the back of the frill is composed of parietal and squamosals, and the ornament consists of three epiparietals (P1–P3), an episquamosal (S1), and an epiparietal crossing the parietal-squamosal border (EPS)4. These differences suggest that resemblance between the two is convergent. However, the matrix used by Sampson et al.4 codes the posterior margin of the frill in C. irvinensis as being composed of the parietals and squamosals, as in Kosmoceratops. More recently, Longrich6 recovered irvinensis within Chasmosaurus.
In addition, Chasmosaurus’ systematics are further complicated by referral of a number of long-horned chasmosaurines to the genus. These include CMN 1254, holotype of Ceratops canadensis, AMNH 5401, holotype of C. kaiseni, and TMP 1989.83.25.1, a partial skull referred to C. russelli. Although traditionally referred to Chasmosaurus8,10,22,23,27 phylogenetic analyses show that these specimens represent a distinct lineage, named Mojoceratops5. The reader is referred to the literature5,28 for a discussion of this issue.
Finally, one specimen referred to Chasmosaurus, CMN 8801, does not appears to belong to that species. Instead, phylogenetic analysis recovers it as sister to K. richardsoni6.
YPM 2016 and all other specimens of Chasmosaurus are from the DPF of southern Alberta32. The formation is late Campanian in age, and is dated to 77.0 Ma - 75.47 Ma33. DPF strata are 70–80 m thick, and consist of river sands and fine-grained overbank deposits. Thick river sandstones are well-developed basally, while towards the top, mudstones and coals document swamps and estuaries formed during the transgression of the Bearpaw Sea34 which overlies the DPF.
At the time, the area was located further north than it is today35, at 58°N, roughly the same latitude as southeast Alaska. The climate would have been seasonal, but warm Cretaceous temperatures supported a diverse biota. The formation contains a rich herpetofauna of fish36, amphibians37, lizards38, turtles35, alligators and choristoderes39. Non-avian dinosaurs were especially diverse, with over 45 known species6, making this the most diverse known dinosaur assemblage. Ornithischians included hadrosaurs, ceratopsians, ankylosaurs, thescelosaurs, and pachycephalosaurs5,6,40,41 and theropods included tyrannosaurids, ornithomimids, caenagnathids, troodontids, and dromaeosaurs42–45 and eight species of ornithurine bird46. A diverse mammalian fauna occurs as well47,48.
While the formation contains a high diversity of dinosaurs, not all species coexisted7,40,49. Instead, many species have restricted stratigraphic distributions. Thus, the DPF assemblage is not a single fauna, but a succession of faunas spanning 1.5 million years.
The lowermost fauna is the Centrosaurus-Corythosaurus fauna; succeeded by a Styracosaurus-Lambeosaurus fauna, and finally a Lambeosaurus magnicristatus-Chasmosaurus/Vagaceratops irvinensis faunal zone. Chasmosaurus itself is known to show turnover through the formation. Holmes et al.7 suggested that three species occur in the formation- C. russelli near the base, C. belli higher up, and C. irvinensis at the top.
Reports of stratigraphic data for YPM 2016 conflict. Lull11 reports that YPM 2016 was “collected by C. M. Sternberg in 1919, on the south side of the west branch of Little Sandhill Creek, 60 feet below the overlying Pierre shales” i.e. the marine shales of the Bearpaw Formation, which overly the top of the DPF. Assuming that the formation is 70–80 m thick here, this puts the skull high up in section, in the muddy interval just below the Lethbridge Coal Zone (LCZ)34, and around 50–60 m above the base of the DPF. These data place YPM 2016 above C. belli, just below C. irvinensis in section7, and near the transition between the Styracosaurus-Lambeosaurus faunal zone and the L. magnicristatus-C. irvinensis faunal zone.
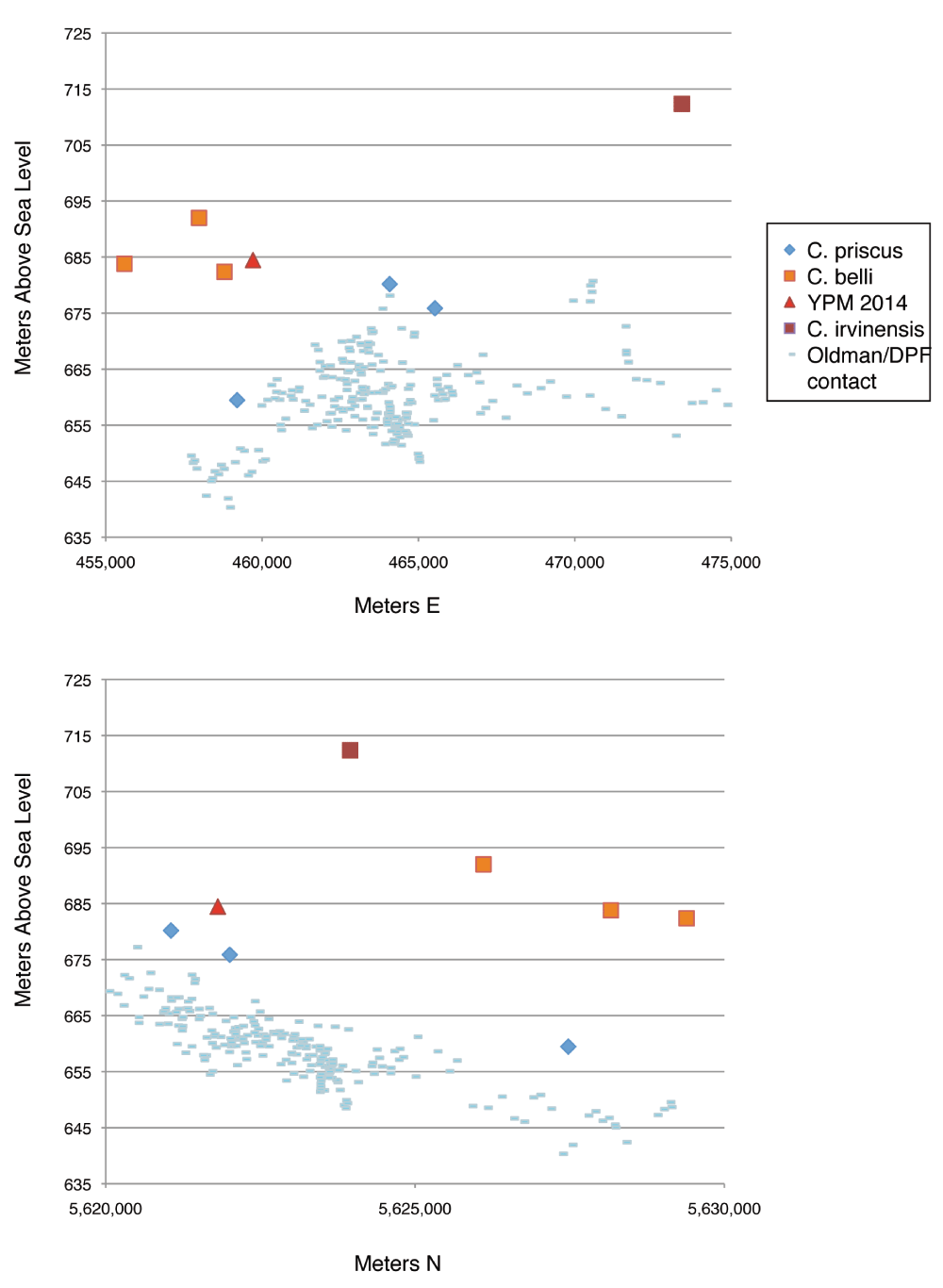
Later, in 1935 and 1936, Sternberg staked quarries and published maps of localities in 1935. YPM 2016 was reported as coming from Quarry 110. In contrast to the original reports, Quarry 110 lies low in section. Godfrey and Holmes8 place the quarry at 31.4 m above the base of the DPF contact, putting it several meters below the highest C. belli (CMN 2245, ROM 839, and ROM 843), and about 20 m below the level estimated in Sternberg’s notes. Currie and Russell32 report that the specimen comes from 684.5 meters above sea level (MASL), versus 682.4-702.7 MASL for other C. belli. This would put the specimen near the bottom of the Styracosaurus-Lambeosaurus faunal zone or at the top of the Centrosaurus-Corythosaurus faunal zone.
The reason for the discrepancy is unclear, but there are several possible explanations.
First, Sternberg may simply have committed an error with his original notes— perhaps misidentifying the top of the DPF, estimating the distance incorrectly, or simply recording the numbers incorrectly. Slumping of the overlying Bearpaw, for example, could create the impression that the contact with the Bearpaw was lower than it actually was.
Second, stratigraphy in the DPF is complicated by a number of factors. The position of the DPF-Oldman contact varies regionally and locally. Within a distance of 250 m, the height of the contact can vary by up to 5 m and it may vary by as much as 10 m over 1.5 km (Currie and Russell, 2005). The thickness of the formation also varies locally by up to 10 m, depending on whether the strata are composed of sandstones, which resist compaction, or mudstones, which readily compact34. These factors mean that distance above the DPF-Oldman formational contact does not correspond precisely with geological age. However, such factors seem unlikely to explain the 20–30 m discrepancy between Sternberg’s notes and more recent measurements. Another possibility is that the overlying marine strata are slumped down- making the contact appear lower than it is.
Third, Sternberg’s notes could be correct, but the specimen may be assigned to the wrong quarry. Quarry maps were not published until 1935 and staking efforts did not occur until 1935 and 1936, 16–17 years after the original excavation50. Given that (i) a number of quarries are known to be misidentified in the original map50, (ii) many known quarries cannot be assigned to any known specimen, and (iii) the original notes are inconsistent with the stratigraphic data for Quarry 110, a very real possibility exists that Sternberg’s original notes are correct and that the specimen does not come from Quarry 110. Sternberg may have simply misremembered where the quarry was. Further work is needed to resolve this issue. As discussed below there are different implications depending on whether YPM 2016 sits low or high in the formation.
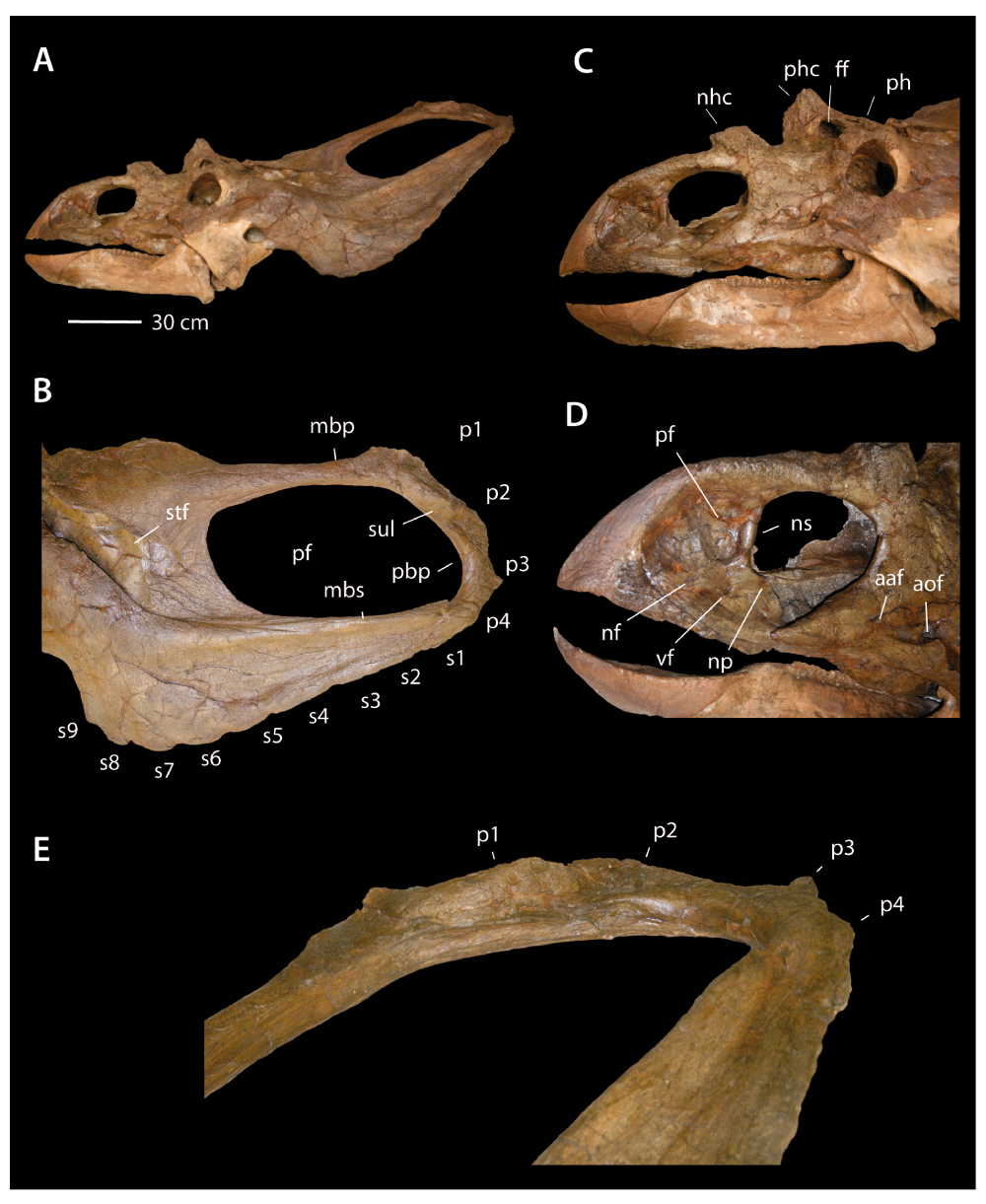
YPM 2016 is a virtually complete skull (Figure 1, Figure 2), but some details are obscured by plaster and many bones are fused, obscuring sutures. The skull is also crushed dorsoventrally, complicating comparisons. YPM 2016 is now on display, and so the palate could not be studied, but was figured in ventral view by Lull11. Throughout, comparisons with other chasmosaurines are based on a revision of chasmosaur systematics (Longrich 20146, this paper). A list of specimens examined and literature studied for this analysis is given in the Supplementary material.

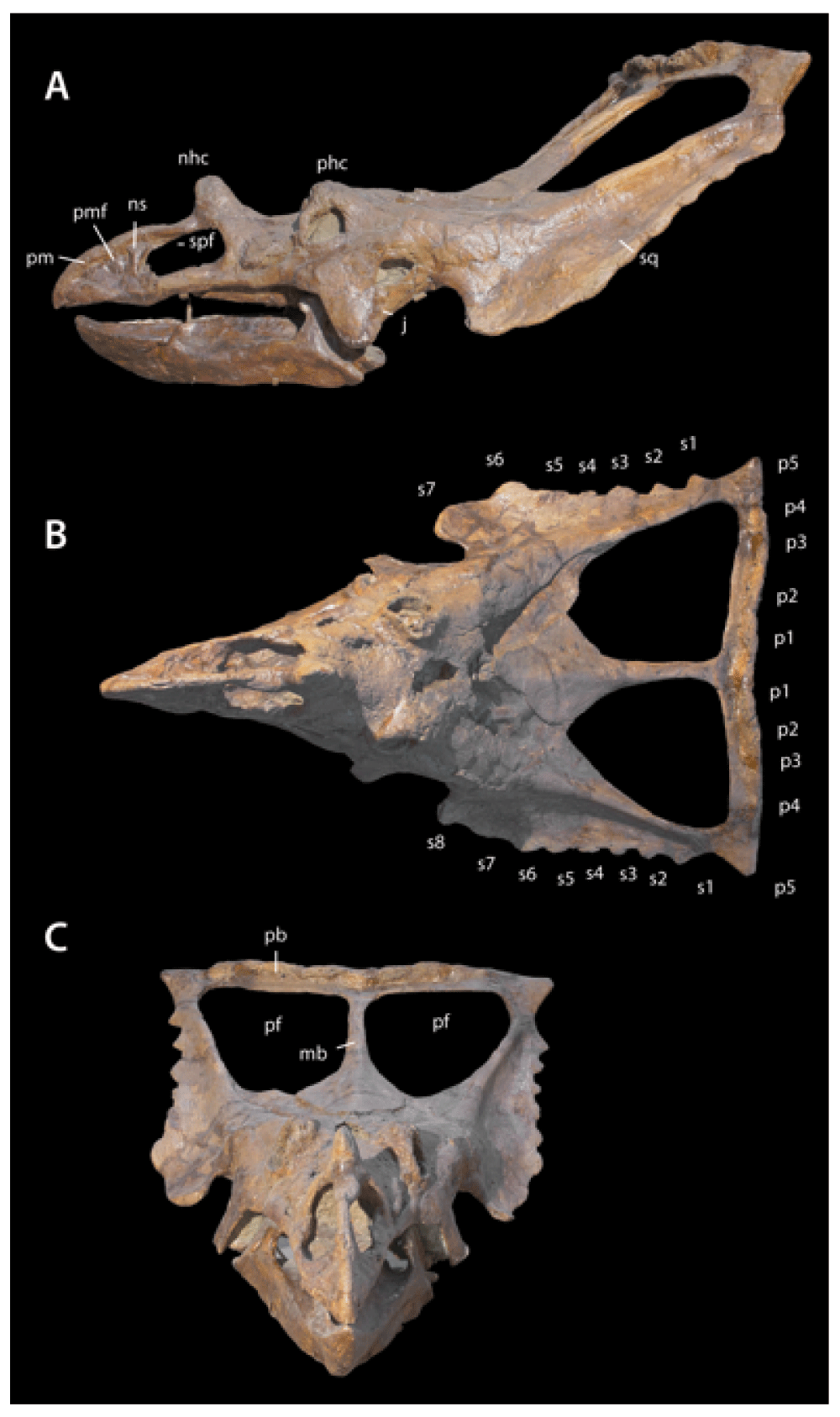
Chasmosaurus YPM 2016 in A, right lateral view; B, dorsal view; C, anterior view. Abbreviations: j, jugal; mb, median bar of parietal; nhc, nasal horn core; ns, narial strut; pb, posterior bar of parietal; pf, parietal fenestra; phc, postorbital horn core; pm, premaxilla; pmf, premaxillary fossa; spf, narial septum posterior flange; s1–s8, episquamosals s1–s8; p1–p5, epiparietals p1–p5.
Premaxillae and rostral. The premaxillae and rostral form a beak that is low, broad, and triangular in lateral view (Figure 3). The tip of the rostral projects anteriorly, similar to C. irvinensis CMN 413577. By comparison, the dorsal margin of the rostral is less triangular and more rounded in C. belli (e.g. AMNH 2402) and in CMN 2280, the snout is much deeper and the rostral has a distinctly rounded anterodorsal margin. These differences may be taxonomically significant. Holmes et al.7 diagnosed C. irvinensis in part on the basis of the low, broad rostrum, although the referred specimen (TMP 87.45.1) has a taller, narrower snout7.
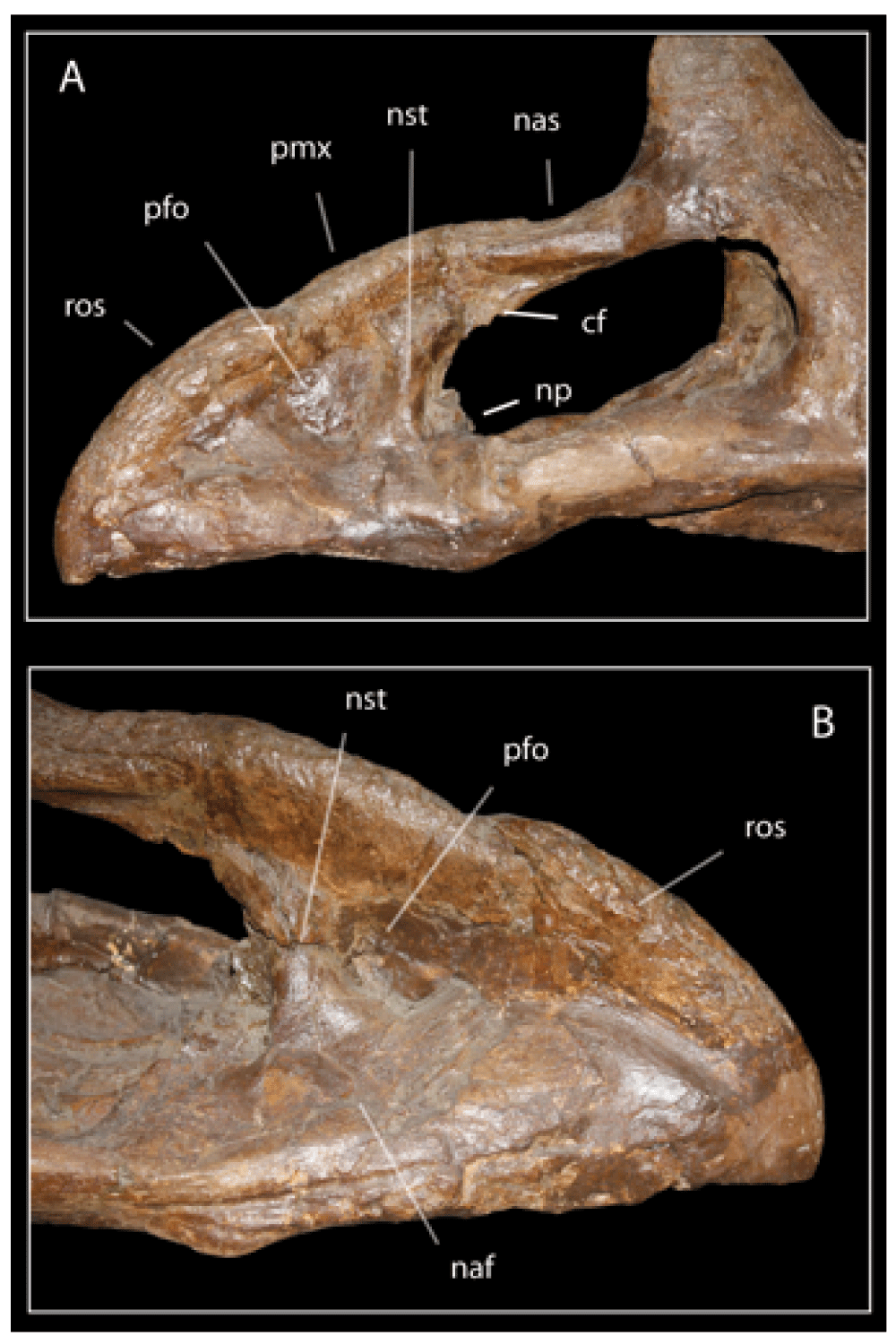
Rostrum of Chasmosaurus YPM 2016 in A, left lateral view, and B, right lateral view. Abbreviations: cf, caudal flange; naf, narial fossa; np, narial process; nst, narial strut; pfo, premaxillary fossa; pmx, premaxilla; ros, rostral.
Anteriorly, the rostral is fused to the premaxillae, a feature of mature chasmosaurines26. The premaxilla bears a broad narial fossa pierced by an ovoid premaxillary fossa, as in other chasmosaurs1. It is unclear if an interpremaxillary fenestra is present; this area is covered with plaster. Below the premaxillary fossa is a shallow anteroventral fossa, as in other chasmosaurines.
A narial strut separates the premaxillary fossa from the naris as in other chasmosaurs1 (Figure 3). In contrast to C. belli (e.g., AMNH 5402, ROM 839, CMN 8800) and CMN 2280, where the strut is anteriorly inclined, the strut is posteriorly inclined in YPM 2016. Posterior inclination of the narial strut is a derived feature shared with C. irvinensis7 and convergently in Arrhinoceratops51 and Anchiceratops13. While the rostrum is crushed, the orientation of the narial strut does not appear to result from distortion of the skull. Reorienting the strut from an anteriorly inclined orientation to a posteriorly inclined orientation would require rotating the premaxilla up and/or shearing the premaxilla, however the premaxillae do not appear to be either rotated upward or sheared.
The posterior margin of the narial strut bore a thin flange. As in other primitive chasmosaurs and centrosaurines, the flange is dorsoventrally extensive, running up the narial strut and onto the ventral margin of the nasal process. The posteroventral margin of the narial strut bears a narial process, a chasmosaurine apomorphy1. The flange is damaged, but apparently had the same subrectangular shape as other Chasmosaurus8.
The nasal processes of the premaxillae are long and slender, and insert into a narrow median slot formed by the premaxillary processes of the nasals. The nasal processes are horizontal anteriorly, but their posterior ends are upturned where they approach the nasal horn. The tips of the nasal processes are fused to the nasals posteriorly, but anteriorly the premaxilla-nasal suture is open.
The maxillary processes are long and slender, as in other chasmosaurines. The ventral margin of the maxillary process is medially inset, producing an anterior extension of the buccal fossa onto the premaxilla. Damage to the skull makes it difficult to ascertain details of the premaxilla-nasal contact.
Nasals. The nasals fuse to each other, the premaxillae, and the frontals. This extensive fusion is seen in other chasmosaurines, e.g. Triceratops26 but is generally absent in centrosaurs. The nasals arch transversely, forming a broad, rounded muzzle (Figure 2, Figure 4) as in other ceratopsids. Their dorsal surface is rugose and gnarled, but their lateral surfaces are smooth.

Skull roof of Chasmosaurus YPM 2016 in A, dorsolateral view and B, anterodorsal view. Abbreviations: ff, frontal fontanelle; nhc, nasal horncore; phc, postorbital horncore. C, anterodorsal view of the skull, showing the frontal fontanelle.
Anteriorly, each nasal bears a long, finger-like premaxillary process. These extend lateral to the nasal processes of the premaxillae to clasp the premaxillae between them, as in other ceratopsians. The maxillary process of the nasal forms the posterodorsal margin of the external naris, and has a long, fingerlike anteroventral extension as in other chasmosaurines.
Dorsally, the nasals bear a conical horn. In other chasmosaurs, the nasal horn develops from an epinasal ossification1,25. This was presumably the case here, but during ontogeny, the epinasal-nasal suture fuses, and so a discrete epinasal is not seen in adults. The nasal horn is shorter and blunter than in most specimens of C. belli (e.g., ROM 843, AMNH 5402) but taller and narrower than in C. irvinensis (CMN 41357).
Maxillae. Maxillae are typical of chasmosaurines in being subtriangular in lateral view (Figure 2). As in other ceratopsians, the toothrow is medially inset and the maxilla bears a broad buccal fossa. Dorsally, the maxilla bounds a small, narrow antorbital fossa. As in other Ceratopsidae, the antorbital fossa extends ventrally into the maxilla to form a dorsally opening pocket, concealing the antorbital fenestra.
The maxillae are long and low. Protoceratops and Centrosaurinae have proportionately shorter, taller maxillae, but chasmosaurs tend to have a low maxilla. Chasmosaurus takes this feature to an extreme7, resulting in an elongate rostrum, with the rostrum depth being 50% or less the skull’s preorbital length. This feature is exaggerated by crushing in YPM 2016, but even so the rostrum would have been elongate as in other Chasmosaurus.
Circumorbital bones. Circumorbital bones (Figure 4), including the prefrontals, frontals, and postorbitals, are fused and the sutures obliterated. The orbit is subcircular, as in most specimens of Chasmosaurus7, although some individuals do have an orbit that is taller than wide10. In almost all other ceratopsids, the orbit is taller than wide. The orbit’s anterior margin bears a rugose preorbital boss as in other ceratopsids.
The postorbitals lack a horn core; instead each bears a massive, gnarled supraorbital boss. The postorbital bosses seen in YPM 2016 also occur in the holotype of C. irvinensis7 and one specimen of C. belli (ROM 843). The holotype of C. russelli (CMN 8800) has a boss over the left orbit, but a short horncore with a resorbed tip over the right.
The top of each supraorbital boss bears a series of pits, up to 20 mm in diameter. Such pits also occur in C. irvinensis7 and in short-horned centrosaurines, e.g. Centrosaurus. They apparently result from resorption of the postorbital horncore as the animals grew52.
Skull roof. The frontals resemble those of other ceratopsids (Figure 4). They are platelike bones that slope down from the orbits toward the midline, making the skull roof dorsally concave. The frontals fuse to the postorbitals and nasals with no trace of a suture. Posteriorly, the frontals border a large frontal fontanelle. The frontal fontanelle is long, narrow, and expanded anteriorly to give it the keyhole shape that characterizes basal chasmosaurines such as Chasmosaurus8 and Mojoceratops5. Unusually, the frontal fontanelle is closed posteriorly by a small midline contact of the frontals (Figure 4B); no other specimen of Chasmosaurus exhibits this feature.
Posteriorly, the frontoparietal fontanelle communicates with the supratemporal fossae via a pair of narrow grooves, the dorsotemporal channels53. Posterior to this, the supratemporal fenestrae open upwards and the supratemporal fossae extend backwards onto the upper surface of the frill as a pair of broad, concave channels between the parietal and squamosal. These channels are partially roofed over by medial extension of the squamosals, which curve in over the supratemporal fossae as in other ceratopsids. The channels do not reach the parietal fenestrae; instead, the space between the parietal fenestrae and the fossae is covered by bone with a highly rugose, vascularized texture; presumably this part of the frill was covered by skin.
Cheek region. The jugal is typical of Ceratopsidae in being a triradiate element (Figure 2A, Figure 4A). It contacts the maxilla anteriorly, the quadratojugal posteroventrally, and the squamosal posterodorsally, and forms the ventral border of the orbit. The jugal is partly covered by paint and plaster, but apparently bore a prominent posterior flange that extends beneath the lateral temporal fenestra, a primitive feature seen in other Chasmosaurus. The epijugal hornlet is reduced to a blunt protuberance. This reduced epijugal is a derived character seen in most but not all other specimens of Chasmosaurus7,8, and convergently in Triceratops18,54.
The quadratojugal is overlapped by the jugal laterally, and overlaps the quadrate laterally. The jugal and quadratojugal, together with the squamosal, define a small lateral temporal fenestra. It is narrow and subtriangular, with its long axis inclined posteriorly, as in C. irvinensis7. The quadrate is similar to that of other chasmosaurs in being anteroposteriorly flattened and mediolaterally expanded to form a broad, platelike bone.
Squamosal. The squamosals (Figure 2, Figure 5A) resemble other chasmosaurines in being subtriangular, with and a long caudal blade and a broad ventrolateral wing. The base of the squamosal projects posteriorly from the back of the skull in lateral view, as in other specimens of Chasmosaurus8 and other basal chasmosaurs such as Mojoceratops5. By contrast, the base of the squamosal projects slightly downward in Pentaceratops where it contacts the back of the skull, and strongly downward in Triceratops, Torosaurus, and centrosaurines.
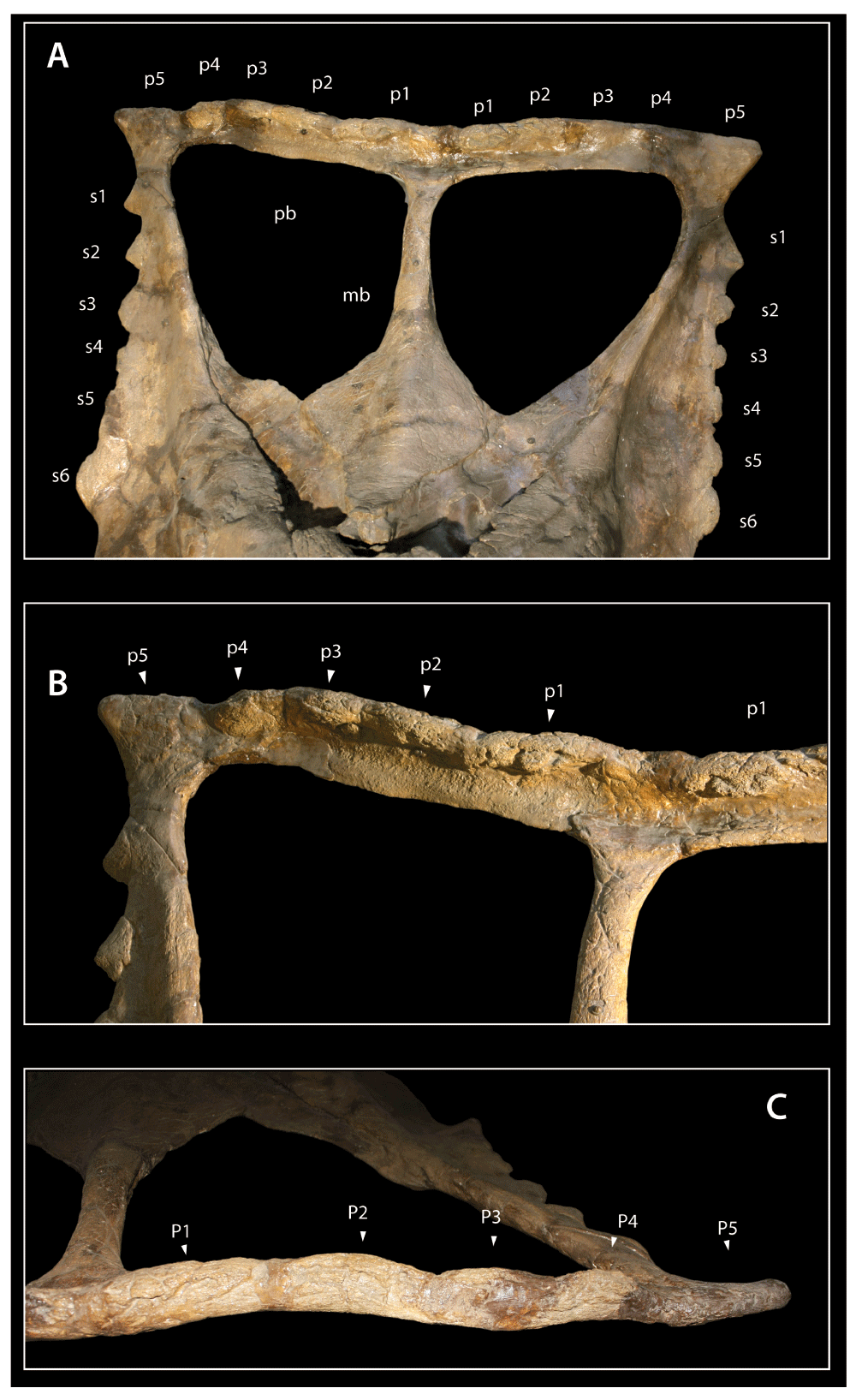
Chasmosaurus YPM 2016, A, frill in dorsal view; B, closeup of right ramus of posterior bar of parietal. Abbreviations: mb, median bar of parietal; pb, posterior bar of parietal; pf, parietal fenestra; s1–s6, episquamosals s1–s6; p1–p5, epiparietals p1–p5.
The caudal blade has a concave dorsal face, with the medial edge curving upwards and the ventrolateral edge projecting down and out. This creates a trough-like concavity along the length of the blade, a derived feature of chasmosaurines. The caudal blade is thin but as in other chasmosaurs it has a thick lateral edge, and a thickened, rounded medial bar where it overlaps the squamosal. The squamosals are distinctly asymmetrical: the tip of the right squamosal is strongly tapered and thickened distally, and has a relatively flat dorsal surface. The end of the right squamosal is broader and thinner, with a more concave dorsal surface. This fluctuating asymmetry- i.e. random deviation from perfect bilateral symmetry- is commonly seen in other chasmosaurs such as Mojoceratops5 and also in pachycephalosaurs55 and is typical of structures under strong sexual selection55.
The left squamosal bears eight epoccipitals. There are seven or eight on the right side, but it is unclear whether the anterolateral corner bears two epoccipitals or one. Epoccipitals are large and crescentic anteriorly, decrease in size posteriorly, then become larger and more triangular towards the tip of the squamosal. The first few epoccipitals are completely fused to the squamosal with no trace of a suture. More posterior epoccipitals are fused to the frill but a suture is visible.
Parietal. The parietal posterior wing is long, broad, and triangular (Figure 5). As in other Chasmosaurus8, and other basal chasmosaurines, parietal fenestrae are expanded so that the frill is reduced to a frame of struts around them. Fenestrae are defined by a median, posterior, and lateral bars. The anterior end of the median bar is a broad, platelike element that is arched transversely, as in C. irvinensis. C. belli is similar, but the prominent dorsal ridge seen in C. belli (e.g. CMN 491) is absent in YPM 2016.
At its midlength, the median bar is wider than tall, measuring 52.5 mm wide by 47 mm tall. The bar becomes slightly narrower and thinner posteriorly. The dorsal surface of the bar is broadly rounded and rugose, and its lateral surface bears smooth, broad sulci. As it joins the caudal bar of the parietal, the median bar becomes wider again but continues to taper in lateral view, measuring 90 mm wide by 32 mm tall.
The lateral bar is incomplete, tapering posteriorly and then disappearing such that the squamosals participate in the parietal fenestrae. A similar condition occurs in CMN 2280. In other specimens referred to C. belli, the lateral bar may be either slender but well developed (CMN 2245; ROM 843) or reduced to a thin lamina (AMNH 5402), but is complete. The anterior part of the lateral bar bears a thin lamina of bone that projects inward to constrict the parietal fenestra, similar to other specimens of Chasmosaurus8.
The parietal caudal bar resembles that of other Chasmosaurus in being straplike in dorsal view. The lateral rami of the caudal bar meet along the midline at an angle of 165°. This angle is intermediate between that of C. belli, where the angle ranges from 150° to 160° (CMN 2245 and ROM 843, respectively), and C. irvinensis, where the lateral rami diverge at an angle of 180° (CMN 41357). As in both C. irvinensis and C. belli, the lateral rami are straight for most of their length, then near their lateral ends they bend anteriorly to underlie the posteromedial end of the squamosal, giving the lateral rami a distinctive ‘L’ shape. By contrast, the lateral rami are straight medially and strongly arched laterally in CMN 2280, giving them a ‘J’ or fishhook shape.
The caudal bar has the distinctive, comma-shaped cross section that characterizes Chasmosaurus. The anterior margin of the caudal bar is thin, measuring about 5–8 mm in thickness, but it becomes thicker posteriorly. It measures 10–12 mm thick a few centimeters behind the fenestra, then becomes much thicker along its caudal margin, measuring up to 25 mm thick near the posterolateral corner and 36 mm thick near the midline.
The posterior edge of the caudal bar curls upwards, such that the dorsal surface of the parietal bar is strongly concave, forming a trough across the midline. A similar condition is present in C. irvinensis7. In other specimens of C. belli, the posterior margin of the bar is curved upwards and there is a dorsal trough, but it is more poorly developed; for example in AMNH 5402, the trough extends across the midline but is relatively shallow, and in ROM 843 the trough is developed laterally but does not extend across the midline. In CMN 2280, the caudal margin of the bar curves upwards, but the dorsal surface is only weakly concave, rather than trough-like, and this concavity does not extend across the midline.
The parietal epoccipitals consist of a large, subtriangular lateral epoccipital, and a series of low epoccipitals medial to this. That the epoccipitals are low and tightly coalesced to the frill obscures their number and arrangement. Because the epoccipitals are upturned, they are best seen in posterior view, where four low, rounded epoccipitals (p1–p4) are bordered laterally by a triangular epoccipital (p5). The arrangement appears to have been similar on the left; p1 and p2 are developed in the same fashion as on the right, but p3–p5 are damaged. Thus, YPM 2016 bears five pairs of epoccipitals. This feature is unusual among chasmosaurs, occurring only in C. irvinensis7 and Torosaurus latus56. By comparison, C. belli bears four pairs of epoccipitals (AMNH 5402, ROM 843), and CMN 2280 has three pairs.
P1–P4 coalesce at their bases to form a continuous, rugose ridge, a derived condition seen in C. irvinensis (CMN 41357)7. The epiparietals are also coalesced at their bases in K. richardsoni4.
In mature C. belli, P1 and P2 fuse (AMNH 5402, CMN 491, ROM 843) but P3 is separate (AMNH 5402, ROM 843, condition unknown for CMN 491). The holotype of C. russelli (CMN 8800) appears to exhibit the same condition as in C. belli; however P1 and P2 are unfused in CMN 2280.
P1 is slightly pointed but P2–P4 are low and rounded. By contrast, P1–P4 are long and pointed in C. irvinensis. Both P1 and P2 are low and rounded in C. belli.
The forward curvature of the epiparietals is intermediate between the condition seen in C. irvinensis and that of C. belli. P1 hooks anteriorly over the frill; P2 and P3 are oriented anterodorsally, P4 projects dorsally, and P5 projects posteriorly. The curvature of the epoccipitals is less extreme than in C. irvinensis, where P1–P3 project anteriorly, and P4 projects anterodorsally. However, the curvature is more extreme than in C. belli and CMN 2280, where P1 and P2 project dorsally, but do not curve forward over the frill.
P5 is large and triangular. A large, triangular lateral epiparietal is seen in other specimens referred to C. belli (e.g. AMNH 5402, ROM 843, CMN 2245) although it appears that the position at which the character is expressed may vary (i.e. a ‘frame shift’): in AMNH 5402 the large epiparietal corresponds to P4; in ROM 843, P3 is enlarged.
Occiput. The occiput exhibits little variation in ceratopsids, and it is typical of ceratopsids in YPM 2016. The exoccipitals form the borders of an ovoid foramen magnum. Above the foramen magnum, the supraoccipital bears a pair of deep depressions, which are separated by a bony lamina. Lateral to the foramen magnum, the exoccipitals extend outwards to form a pair of broad paroccipital processes, contacting the parietals dorsally and the squamosals laterally. The paroccipital processes are expanded distally to give them a triangular shape, and twist to face posteroventrally. Below the foramen magnum, the exoccipitals and basioccipital fuse to form a ball-shaped occipital condyle that is offset from the braincase by a distinct neck. The basal tubera are robust and mediolaterally expanded.
To assess the relationships of Chasmosaurus, YPM 2016 was placed in a phylogenetic analysis along with specimens referred to Chasmosaurus and Mojoceratops, to create a specimen-level phylogeny. The matrix (Appendix 2) is modified from Longrich6 which in turn draws on several previous studies, primarily Dodson et al.1 and its derivatives3,5,28,57, but also Holmes7 and Sampson et al.4.
The matrix includes 203 characters; characters were treated as ordered where a transitional sequence could be identified (e.g., postorbital horns absent, short, moderately elongate, elongate). Character descriptions are given in the Supplementary material (Appendix 1). The character-taxon matrix (Appendix 2) was coded using Mesquite 2.7558. Following Longrich6, Ojoceratops, Nedoceratops, and Tatankaceratops are referred to Triceratops horridus and T. prorsus and excluded from analysis; in particular, characters previously used to diagnose Nedoceratops, including the orientation of the postorbital horns and the position of the squamosal59 are present only on one side of the skull (compare Farke 2011, Figure 1A,B vs. Figure 1C,D), indicating that they are artifacts resulting from postmortem distortion.
Analyses were conducted using PAUP* 4.0 b 1060 for phylogenetic analysis and TNT 1.1a61 to calculate support values. A specimen-level analysis was conducted including individual specimens referred to Chasmosaurus and Mojoceratops. Next, species were delimited based on the results of the specimen-level analysis and composite codings for each Chasmosaurus species were used for a species level-analysis.
Then, to test whether the results are contingent on the matrix used, the analysis of Sampson et al.4 was rerun with YPM 2016 included. For this study, the codings of the frill and epiparietals for C. irvinensis were revised. Following Holmes et al.7 and this study, the posterior margin of the frill in C. irvinensis is formed of the parietal, with five pairs of epiparietals. Characters were recoded for the matrix as follows: 60:2->1 (squamosal subequal to parietal->squamosal slightly shorter than parietal), 61:1->0 (squamosal forms part of posterior margin of frill, present->absent), 89:2->1 (P2 elongate->P2 D-shaped), 91:1->0 (EPS present->absent), 92:1->? (EPS is absent and therefore its shape cannot be scored), 93:0-1 (3 epiparietals->5 epiparietals). Characters 60, 61, 91, and 92 are recoded to reflect the fact that the posterior margin of the frill appears to be composed of the parietal (as reconstructed by Godfrey et al.) whereas Sampson et al. code the frill as being composed of the parietal and squamosal. The reader is referred to the discussion below for the rationale for these codings.
This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the International Commissions on Zoological Nomenclature. The ZooBank LSID (Life Science Identifier) is:
urn:lsid:zoobank.org:pub:DD4D2E55-7974-4C16-BE01-BD433ABD0000 (the information associated with this LSID can be viewed through any standard web browser by adding the LSID to the prefix http://zoobank.org/)
Phylum: Chordata
Subclass: Dinosauria
Family: Ceratopsidae
Subfamily: Chasmosaurinae
Genus: Chasmosaurus
Derivation of name: chasma, Greek, wide opening, sauros, Greek, lizard.
Diagnosis: Chasmosaurine ceratopsid characterized by the following derived characters. Supraorbital horncores short, length 200% basal diameter or less. Posterior bar of parietal with a thickened caudal margin and a thin lamina extending anteriorly. Posterior margin of parietal upturned and dorsal surface of posterior bar concave, epoccipitals P1 and P2 projecting dorsally or anterodorsally.
Species: priscus.
Derivation of name: priscus, Latin, ancient or primitive
Diagnosis: Rostrum rounded anteriorly in lateral view. Orbital horncores short, length 150%–200% basal diameter. Parietal bar with a well-developed caudal emargination, with lateral rami of caudal bar diverging at an angle of 120º. Lateral rami of posterior bar straight medially, but arched laterally, giving them a fishhook shape. Dorsal surface of lateral rami weakly concave, producing a shallow trough that does not extend across the midline. Three pairs of short, broad epiparietals, which do not coalesce at their bases. C. priscus is also distinguished by from C. belli and C. irvinensis by the absence of the derived features characterizing those species, described below.
Holotype: CMN 2280
Type locality: Dinosaur Provincial Park, Alberta
Specimen-level analysis. A specimen-level analysis was run using a heuristic search algorithm until 69201 trees were obtained. The resulting tree statistics were tree length (TL) = 389; consistency index (CI) = .5979; retention index (RI) = .8298.
In the strict consensus (Figure 6) Chasmosaurus-like skulls fall into two clusters. One includes chasmosaurines with long, anterolaterally projecting horns, an expanded, triangular frontal fontanelle, a deeply notched parietal, a parietal caudal bar with a concave anterior surface and tuberosities on the anterodorsal margin, and posteriorly projecting epiparietals. This clade corresponds to specimens previously referred to Mojoceratops5. Variation exists within this cluster5, but there is no resolution within this grouping, because so many specimens are fragmentary.
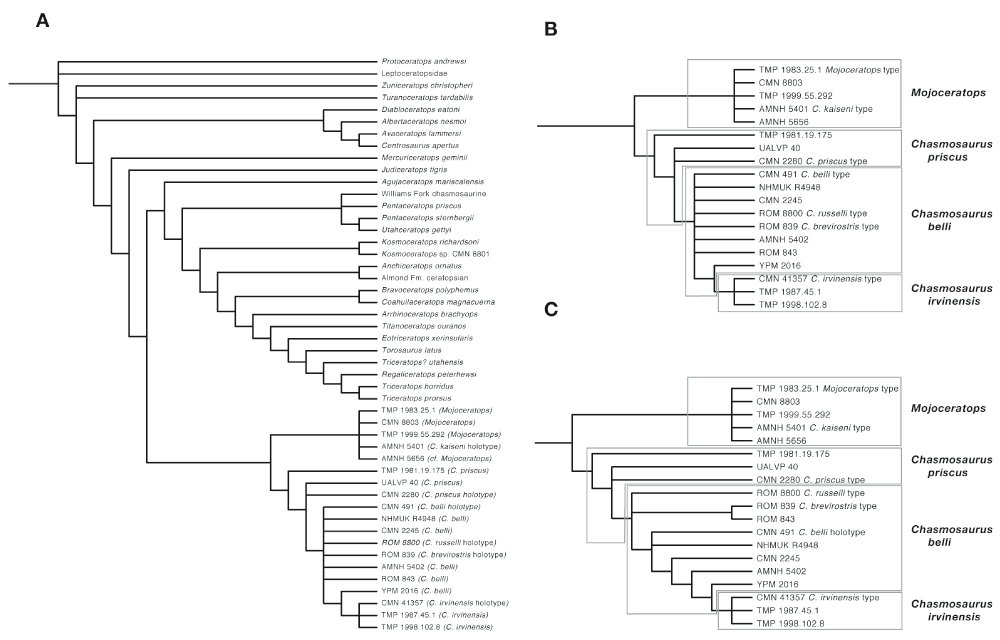
A, strict consensus tree showing all taxa; B, strict consensus showing Chasmosaurus specimens and proposed taxonomy; C, adams consensus showing the relationships of Chasmosaurus specimens. Treelength = Consensus of 50184 trees. Treelength = 389; CI = .5979; RI = .8298.
The second cluster contains chasmosaurines with short postorbital horns and a parietal caudal bar with a thickened, upturned posterior margin. This group corresponds to Chasmosaurus as defined by Longrich5. Within Chasmosaurus, phylogenetic structure exists in the strict (Figure 6B) and Adams (Figure 6C) consensus trees. Three main groupings can be identified. The first group, including the most primitive specimens, TMP 1981.19.175, CMN 2280 and UALVP 40, lie basal to all other Chasmosaurus. They have moderately short brow horns and in CMN 2280, strong caudal emargination of the parietal. The second group includes specimens previously referred to C. belli, including the holotype, CMN 491, and AMNH 5402, ROM 843, CMN 2245, and NHMUK R4948. This group also includes the holotypes of C. russelli, CMN 8800 and C. brevirostris, ROM 839. They are united by very short brow horns, weak caudal emargination of the parietal, an L-shaped lateral ramus of the caudal bar, and a low and coalesced P1 and P2.
The third group includes C. irvinensis, including CMN 41357, TMP 87.45.1, and CMN 41357. This clade is characterized by a posteriorly inclined narial strut, loss of brow horns, an expanded caudal end of the squamosal, a straight posterior bar of the parietal, and five parietal epoccipitals. The parietal ornament of these animals is specialized, with P1-P4 being fused, elongate, and turned forward over the frill.
The Peabody Chasmosaurus, YPM 2016, does not fit easily into any group. It lies just outside of the grouping formed by C. irvinensis, in both strict and Adams consensus trees, reflecting the intermediate nature of its morphology. It shares derived features with C. irvinensis, and plesiomorphic features with C. belli. Derived features include a posteriorly inclined narial strut and loss of the brow horns, and P1–P4 are fused. The parietal emargination is reduced compared to C. belli, and is therefore derived here, but not to the degree seen in C. irvinensis. Similarly, P1 and P2 are turned forward, a derived feature, but not P3 and P4, a primitive feature. P1 is slightly elongated, a derived feature, but P2–4 are not, which is primitive. P5 is smaller than in typical C. belli, but larger than in C. irvinensis. The parietal fenestrae are large and the median bar is subrectangular, both plesiomorphies shared with C. belli.
When all specimens are included, bootstrap and Bremer support indices are poor (Figure 7A). This seems to result from inclusion of a high number of incomplete specimens, rather than a lack of signal in the data: when the more incomplete specimens are excluded and the analysis is rerun, the resulting tree has high support values for most nodes, but especially for the inclusion of C. irvinensis within Chasmosaurus.
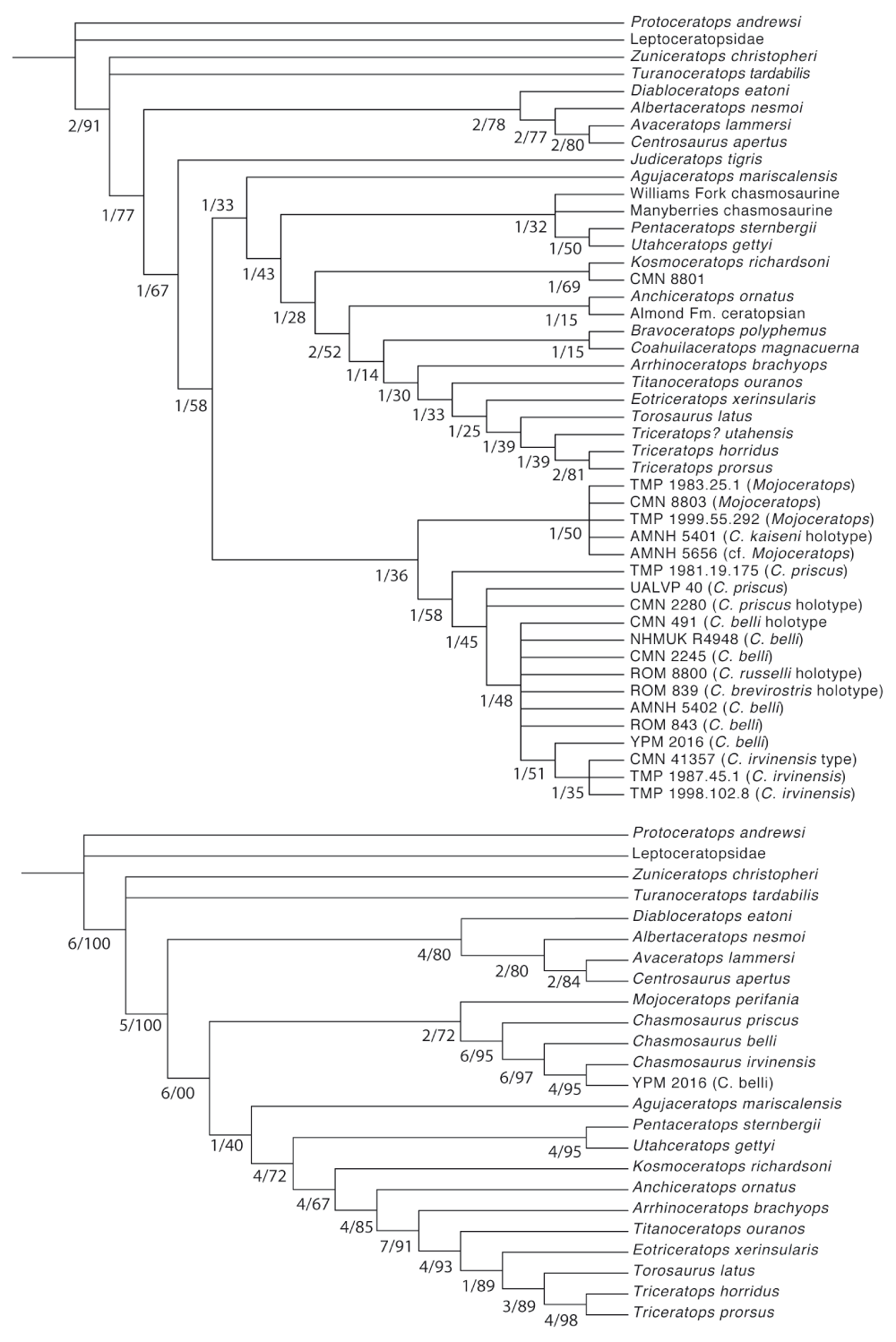
A, support values for specimen-level analysis; B, support values for analysis focusing on complete specimens. Low support values in the first analysis seem to result primarily from missing data, rather than conflicting data or lack of support for particular nodes.
Results of species-level analysis. The species-level analysis, with YPM 2016 included, produced 9 most parsimonious trees (Figure 8) (Treelength = 377, Consistency Index = .6435, Retention Index = .8184, Rescaled Consistency Index = .5266). Results do not differ from the specimen-analysis, but conducting the analysis at species rather than specimen level speeds up analysis and makes it possible to use a branch-and-bound search. Results broadly agree with a number of previous analyses1,3–5,13,28,29,57 in putting Chasmosaurus basal within the Chasmosaurinae.
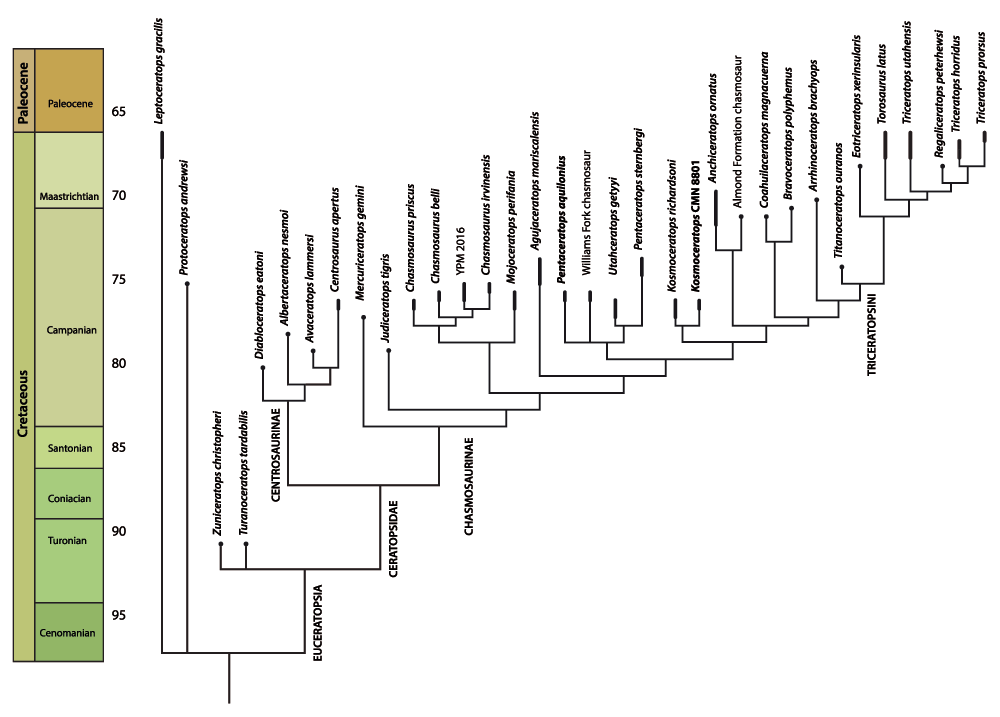
Strict consensus of 9 most parsimonious trees (TL = 377, CI = .6435, RI = .8184, RC = .5266).
Reanalysis of Sampson et al. (2010) matrix. Reanalysis of the Sampson et al. matrix (Figure 9) produced a total of 6 trees (Treelength = 270, Consistency Index = .6426, Retention Index = .7804, Rescaled Consistency Index = .5087). In this analysis, irvinensis clusters with Chasmosaurus (Figure 9B). This result is contingent on the inclusion of YPM 2016. When YPM 2016 is deleted (Figure 9A), C. irvinensis again clusters with Kosmoceratops, consistent with the original tree4.
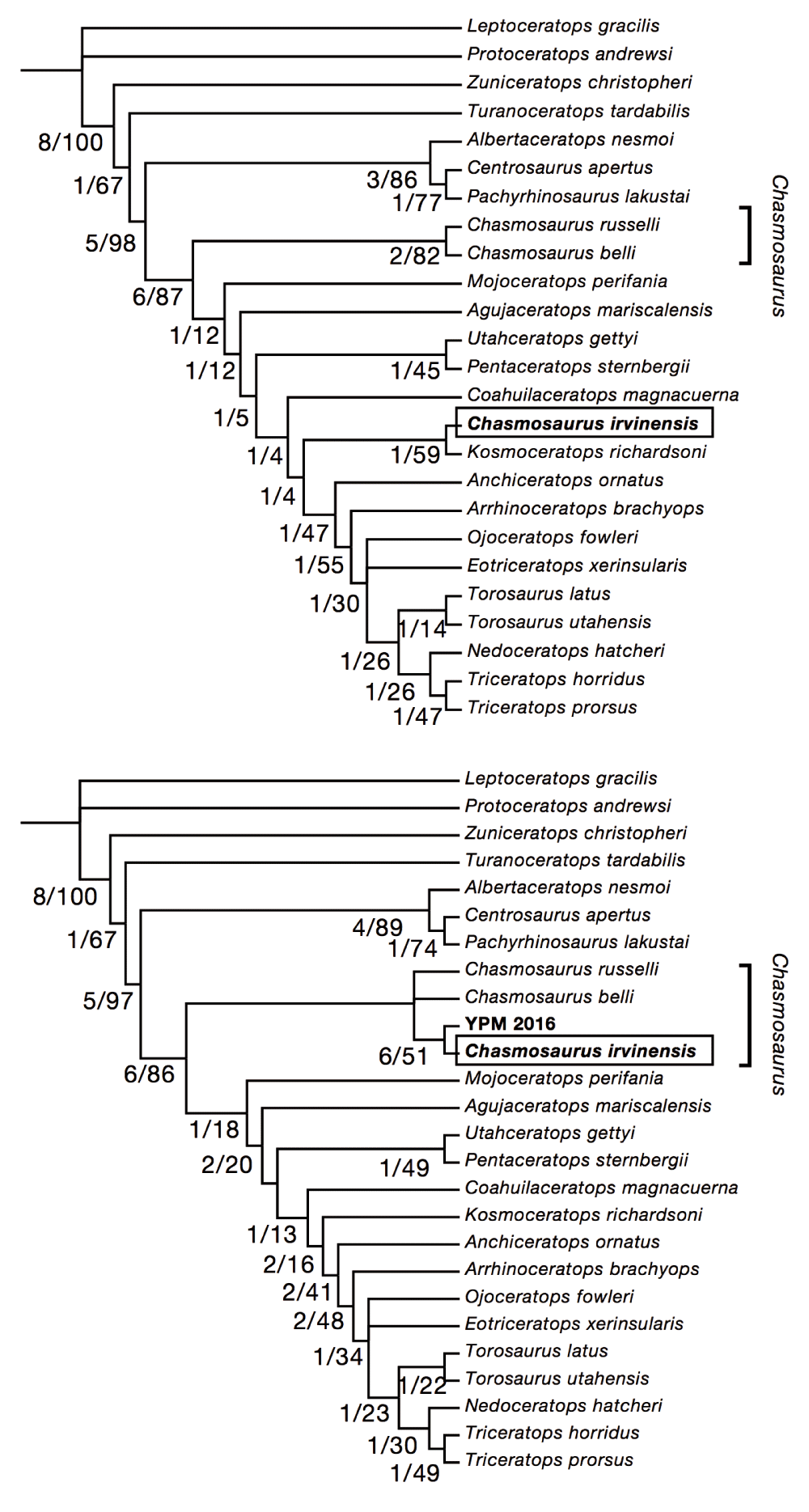
A, results of analysis using Sampson et al. (2010) with C. irvinensis recoded, with polyphyletic Chasmosaurus. Strict consensus of 6 most parsimonious trees; TL = 267, CI = .6500, RI = .7791, RC = .5136. B, results of analysis with YPM 2106 included, strict consensus of 6 most parsimonious trees; TL = 270, CI = .6426, RI = .7804, RC = .5087. Inclusion of the transitional morphology represented by YPM 2016 breaks up the long branch separating C. belli and C. irvinensis, resulting in a monophyletic Chasmosaurus.
The phylogenetic results presented here provide a framework for interpreting the taxonomy of Chasmosaurus. Critical issues addressed here include the assignment of C. irvinensis to the genus Vagaceratops, the diagnosis of C. belli, validity of C. russelli, and the identity of CMN 2280.
Phylogenetic analysis places the species irvinensis not as the sister taxon of C. belli but nested within C. belli. Numerous synapomorphies unite specimens referred to C. belli and C. irvinensis, including reduced brow horns, a frill that is weakly inclined in lateral view, a parietal posterior bar that is thickened posteriorly and upturned such that the bar is concave dorsally, and an upturned P1 and P2. It shares with C. belli further reduction of the orbital horns, the L-shaped lateral bars of the parietal, reduced midline emargination of the frill, and coalesced P1 and P2. Finally, C. irvinensis shares with YPM 2018 the further reduction of the posterior emargination of the frill, the five pairs of parietal epoccipitals, coalescence of P1–P4 at the bases, and strong forward curvature of P1 and P2.
The relationships found here do not result from the characters selected for this study. As discussed above, when C. irvinensis is recoded in the Sampson et al.4 matrix following the original description and YPM 2016 is included, irvinensis is again recovered with Chasmosaurus.
Part of the argument for uniting C. irvinensis with Kosmoceratops concerns frill morphology. As discussed above, in the description of C. irvinensis, the back of the frill was interpreted as being composed of the parietals, with five epiparietals7, as in the Peabody Chasmosaurus. However, Sampson et al.4 interpreted the frill as being composed of the parietals and squamosals, with three epiparietals, as in Kosmoceratops. The suture between the parietal and squamosal is difficult to trace distally in C. irvinensis, but it appears to curve laterally towards the end of the frill (Figure 10), such that the parietal forms the back of the frill, as suggested by Holmes et al.7. The interpretation advanced by Sampson et al.4 would instead require that the ends of the squamosals make a 90° bend and hook in medially behind the parietal fenestrae. No other ceratopsid exhibits such a configuration; more importantly, there is no evidence for the parietosquamosal suture here.

Abbreviations: par, parietal; sq, squamosal; p1–p5, epiparietals 1 – 5.
The results of the phylogenetic analysis mean that referring irvinensis to Chasmosaurus is a reasonable taxonomic decision given that the animal nests within specimens previously referred to Chasmosaurus. That being said, the animal is also very distinct from C. belli, to the point that the relationship between the two has been controversial. Given this, treating the animal as a distinct genus, Vagaceratops is also reasonable. One emphasizes the continuity of the lineage, the other approach emphasizes the marked differences.
Neither is more correct, in a strictly scientific sense, than the other. Given the current tree topology, the difference between C. irvinensis and V. irvinensis is semantic, not scientific. There is no objective way of deciding which name to use, because there is no objective, scientific criterion for deciding how distinctive a species must be to merit its own genus. The choice is up to the taxonomist; it is an issue of taxonomic bookkeeping, and not a scientific debate. Here, Chasmosaurus is used because it emphasizes the continuity of the lineage, in part because of tradition, and in part because it is simply more euphonious. Admittedly, these are not scientific reasons, but the argument is not a scientific argument. The debate will need to be settled through the consensus of the palaentological community. For this paper, I have chosen to use Chasmosaurus. Other paleontologists can either follow this approach or not, as they see fit.
Including Vagaceratops within a clade of Chasmosaurus does have an interesting consequence: Chasmosaurus becomes paraphyletic. Although some authors argue that genera should be monophyletic62, others do not recognize monophyly as necessary or desirable; Lehman30 presented a phylogeny in which Chasmosaurus and Torosaurus were both paraphyletic genera. In fact, the International Code of Zoological Nomenclature does not require monophyletic taxa and, as discussed below, paraphyletic taxa are an inevitable consequence of a ranked taxonomy.
Specimens previously referred to C. belli are recovered as a paraphyletic grouping. No derived features were found to unite C. belli that are not also found in C. irvinensis. Instead, C. belli is diagnosed only by the absence of apomorphies diagnosing C. irvinensis. Furthermore, if YPM 2016 is included in C. belli, then C. belli ranges in form from primitive animals that retain short orbital horns and strongly emarginate parietals with four epoccipitals, to derived animals with orbital bosses, and weakly emarginated parietals with five epoccipitals. Excluding YPM 2016 from C. belli and referring it to C. irvinensis would result in a more limited range of variation in the species, but would not result in a monophyletic C. belli. The only way to create a monophyletic C. belli would be to create new species for all forms that lie either above the holotype or below the holotype on the tree.
As with the decision to refer C. irvinensis to Chasmosaurus, the decision to leave YPM 2016 in C. belli is largely arbitrary. Here, YPM 2016 is referred to C. belli because it lacks many of the apomorphies of C. irvinensis. Yet one could just as easily refer YPM 2016 to C. irvinensis because it exhibits a number of the apomorphies of this species that are absent in C. belli.
Chasmosaurus russelli (Figure 11) emerges as a problematic taxon in the current analysis. Sternberg9 diagnoses C. russelli as follows:
A, left lateral view; B, close up of frill; C, close up of face; D, close up of rostrum, E, close up of posterior margin of frill. Abbreviations: aaf, accessory antorbital fenestra; aof, antorbital fenestra; ff, frontal fontanelle; mbs, medial bar of squamosal; nf, narial fossa; nhc, nasal horn core; np, narial process; pbp, posterior bar of parietal; pf, parietal fenestra, pmf, premaxillary fossa; phc, postorbital horn core; stf, supratemporal fossa; sul, sulcus; vf, ventral fossa; p1–p4, epiparietals 1–4; s1–s9, episquamosals 1–9.
“1) Skull large, 2) relatively high and short in front of orbits; 3) rostral straight inferiorly, not hooked downward at tip, 4) nasal horncore massive, broad between orbits; 5) no brow horncores; 6) well-developed epijugal 7) parietals deeply indented posteriorly, 8) squamosal border not strongly scalloped, 9) epoccipitals small, 10) mandible massive.” (numbers are inserted for the reader).
Character 1 applies to all ceratopsids. Character 2 is possibly due to crushing. Character 3 cannot be assessed in the type. Character 4 does not differ markedly from the condition in C. belli. Character 5 is found on the left side of the skull only, and is seen in some C. belli and C. irvinensis. Character 6 is developed to a comparable degree in some specimens of C. belli (e.g., ROM 843). Character 7, parietal emargination, is debatable. The skull is crushed almost flat, with the squamosals displaced to lie posterior to the parietals, rather than lateral to them, and the right horn lies ahead of the left, showing that the entire skull has been not only crushed but sheared, with the left side of the skull moved posteriorly relative to the right. It is therefore impossible to know if the emargination is real, or simply a result of crushing. Character 8 is also seen in C. irvinensis. Character 9, reduction of the epiparietals, is seen in the median epiparietals of C. belli, although the reduction of the lateral epiparietals seems to be unique to this specimen. Character 10 is not preserved in the holotype.
There are real differences between CMN 8800 and characteristic C. belli, including the weakly scalloped squamosal and small lateral epiparietals, but they do not appear to lie outside the range expected for intraspecific variation, and so the differences identified by Sternberg appear insufficient to separate the type from C. belli.
C. russelli has continued to be treated as a distinct species, however. The reason is that CMN 2280 was referred to C. russelli, and the species has then been diagnosed based on this highly complete and well-described specimen8. Godfrey and Holmes diagnose C. russelli as follows: “1) Posterior margin of parietal frill broadly arched on either side of median emargination. 2) Each side bears three low, triangular roughly equal-sized epoccipitals. 3) The lateral ramus of the parietal is reduced and does not completely encircle the fenestra in all but one specimen, permitting the squamosal to form a part of its lateral border” (numbers added here for clarity). Again, this diagnosis is based on CMN 2280, not on the holotype of C. russelli. CMN 8800, the holotype, clearly lacks characters 1 and 2 (Figure 11). Character 3 cannot be assessed in the holotype because the skull is embedded in the matrix; at any rate it is present in C. belli (e.g. YPM 2016) and therefore does not differentiate C. russelli.
Longrich5 provided the following diagnosis: “Chasmosaurus exhibiting the following combination of characters: 1) lateral rami of parietal posterior bar weakly arched in dorsal view, 2) well-developed caudal emargination of the frill, with an angle between the lateral rami of 120°, 3) three broad, moderately elongate parietal epoccipitals.” Again, the diagnosis is based on CMN 2280, not the holotype, CMN 8800, and characters 1 and 3 (= Godfrey and Holmes’ characters 1 and 2) are absent in CMN 8800, and as discussed above, the emarginated parietal of CMN 8800 may be a preservational artifact. While these problems were recognized during the writing of the 2010 paper, it was felt that dealing with Chasmosaurus russelli was beyond the scope of that particular project.
Last, Maidment and Barrett10 diagnose C. russelli as follows: “taxon displaying the combination of characters unique to the genus Chasmosaurus along with the following features: 1) parietal posterior bar bearing a median emargination and is broadly arched either side so that it forms an ‘M’ shape with the parietal median bar; 2) each side of the posterior parietal bar bears three roughly equally sized epiparietals (after Godfrey & Holmes, 1995). Both characters are autapomorphic for C. russelli within Chasmosaurus.” This diagnosis follows Godfrey and Holmes8, and again is based on CMN 2280, not the type of C. russelli.
Sternberg’s diagnosis of C. russelli does not provide a basis for recognizing a distinct species, and the holotype cannot readily be separated from C. belli. Meanwhile, the diagnoses of Godfrey and Holmes8, Longrich5 and Maidment and Barrett10 do potentially diagnose a distinct species, but the diagnosis is based on CMN 2280, and the holotype of C. russelli, CMN 8800, does not fit this diagnosis, as it has reduced medial epiparietals and lateral episquamosals. CMN 8800 instead appears to represent a crushed C. belli, and the reduction of the epoccipitals may be related to the age of the specimen, as seen in other chasmosaurs24. This conclusion is supported by phylogenetic analysis (Figure 6). However, the diagnoses provided by Godfrey and Holmes8, Longrich5 and Maidment and Barrett (2011) do diagnose a distinct taxon, in that they separate CMN 2280 from the remaining Chasmosaurus.
A number of plesiomorphies distinguish CMN 2280 from C. belli and C. irvinensis (excellent figures and a description of CMN 2280 were presented by Godfrey and Holmes; so readers are referred to that paper for a thorough description). First, while the brow horns are reduced in CMN 2280, they are markedly longer than in C. belli (e.g. AMNH 5402). Although horn length changes with age24,52, this feature is difficult to explain in terms of ontogeny. Because CMN 2280 is near maturity: the frill is rugose and some of the epoccipitals are fused, with only the rostral and some epiparietals remaining unfused. By comparison, AMNH 5402 is comparable in terms of maturity, but has shorter horns.
The posterior margin of the frill also has a distinct M-shape, similar to that of Mojoceratops, although not developed to the same degree. The posterior bar of the parietal is straight medially, but strongly curved laterally, such that each ramus of the bar is shaped like a ‘J’ or a fishhook. By contrast, the posterior bar is straight along most of its length in C. belli, then sharply bends forward near the parietal-squamosal junction, giving it a shape like an L or a 7. The arching of the posterior bar also means that the parietal projects well posterior to the squamosals, approaching the condition in Mojoceratops. By contrast, the parietal does not project as far in C. belli. The emargination of the parietal is also well developed; the lateral rami of the posterior bar diverge at an angle of 120°, versus 150° or more in C. belli.
In addition, the parietal is more primitive in the weak development of the distinctive upturned edge of the frill. The posterior margin of the parietal bar is slightly upturned in CMN 2280 and its dorsal surface is weakly concave. By comparison, the posterior margin is strongly upturned in C. belli, resulting in a trough along the dorsal surface of the parietal posterior bar.
Finally, the parietal ornament is more primitive. There are only three parietal epoccipitals, versus four or more in C. belli. P1 and P2 are also prominent and separate, versus low and coalesced in C. belli.
Thus, there appears to be a primitive form of Chasmosaurus present in the DPF, which is represented by CMN 2280. TMP 1981.19.175 and UALVP 40 may also represent this species, as they retain relatively long brow horns. TMP 1981.19.175 may be more primitive than CMN 2280, because the parietal fenestrae appear to extend further anteriorly, as in Mojoceratops, whereas there is a broad sheet of bone behind the supratemporal fenestra in CMN 2280, as in C. belli and C. irvinensis.
The differences seen here appear to be sufficient to warrant a distinct species, but the referral of CMN 8800, the holotype of C. russelli, to C. belli leaves this species without a name. Accordingly, the name C. priscus is proposed as a replacement. Following the results of the analyses presented above, the following is proposed as a revised taxonomy of the genus.
Taxonomy
Dinosauria Owen 1842
Ornithischia Seeley 1888
Ceratopsia Marsh 1890
Euceratopsia new taxon
Ceratopsidae Marsh 1888
Chasmosaurinae Lambe 1915
Chasmosaurus Lambe 1914
Euceratopsia is a node-based clade defined as the most recent common ancestor of Zuniceratops, Turanoceratops, and Triceratops, and all of its descendants.
Chasmosaurus Lambe 1914
Chasmosaurine ceratopsid characterized by the following derived characters. Supraorbital horncores short, length 200% basal diameter or less. Posterior bar of parietal with a thickened caudal margin and a thin lamina extending anteriorly. Posterior margin of parietal upturned and dorsal surface of posterior bar concave, epoccipitals P1 and P2 projecting dorsally or anterodorsally.
Type species. Chasmosaurus belli Lambe 1902.
Distribution. Dinosaur Park Formation, Alberta, Canada.
Chasmosaurus priscus sp. nov.
Diagnosis. Rostrum rounded anteriorly in lateral view. Orbital horncores short, length 150%–200% basal diameter. Parietal bar with a well-developed caudal emargination, with lateral rami of caudal bar diverging at an angle of 120°. Lateral rami of posterior bar straight medially, but arched laterally, giving them a fishhook shape. Dorsal surface of lateral rami weakly concave, producing a shallow trough that does not extend across the midline. Three pairs of short, broad epiparietals, which do not coalesce at their bases. C. priscus is also distinguished by from C. belli and C. irvinensis by the absence of the derived features characterizing those species, described below.
Holotype. CMN 2280.
Referred specimens. TMP 1981.19.175 (?), UALVP 40 (?)
Distribution. Lower Dinosaur Park Formation, Alberta, Canada.
Chasmosaurus belli Lambe 1902
Synonyms
Monoclonius belli Lambe 1902
Ceratops belli Hatcher 1907
Protorosaurus belli Lambe 1914
Chasmosaurus brevirostris Lull 1933
Chasmosaurus russelli Sternberg 1940
Diagnosis. Chasmosaurus distinguished from C. priscus by the following derived characters. Rostrum with subtriangular beak in lateral view. Orbital horncores very short or absent, with length subequal to or less than basal diameter. Parietal short, with limited projection posterior to squamosals. Caudal emargination of parietal reduced, lateral rami of caudal bar diverging at an angle of 140°–165°. Lateral rami of posterior bar straight medially and strongly bent at the corner of the frill, giving them the shape of the numeral 7. Lateral rami of caudal bar with a strongly concave dorsal surface, which may or may not extend across the midline. At least four parietal epoccipitals. Epiparietals P1 and P2 short and their bases coalesced. Large, subtriangular lateral epiparietals. C. belli is typically distinguished from C. irvinensis by the absence of the derived features characterizing that species, which are described below. However, some of these features are in fact found in YPM 2016.
Holotype. CMN 0491
Referred specimens. AMNH 5402, CMN 2245, NHMUK R4948, ROM 839 (holotype of “Chasmosaurus brevirostris”), ROM 843, CMN 8800 (holotype of “Chasmosaurus russelli”), YPM 2016.
Distribution. Lower Dinosaur Park Formation, Alberta, Canada.
Chasmosaurus irvinensis Holmes et al. 2001
Synonyms
Vagaceratops irvinensis Sampson et al. 2010
Diagnosis. Narial strut of premaxilla inclined posteriorly. Orbital horncores reduced to low, rugose bosses. Squamosal broad distally. Parietal with limited projection beyond squamosals. Parietal fenestrae reduced, broader than long. Caudal emargination of parietal absent, posterior bar of parietal straight. Caudal margin of parietal strongly upturned, defining a deep, trough-like concavity developed on the dorsal surface of the lateral rami of the parietal bar and extending across the midline across the junction of the caudal bar and median bar. Five pairs of parietal epoccipitals. Parietal epoccipitals strongly curved forward, with apices of median parietal epoccipitals projecting anteriorly. Medial parietal epoccipitals elongate.
Holotype. CMN 41357.
Referred specimens. TMP 87.45.1, TMP 98.102.8
Distribution. Upper Dinosaur Park Formation (Lethbridge Coal Zone), Alberta, Canada.
Speciation of Chasmosaurus could potentially involve anagenesis, cladogenesis, or some combination of the two. The two different mechanisms make distinct predictions about both tree structure and stratigraphic distribution. Overall, the structure of the tree found in the current analysis (Figure 6) is consistent with the hypothesis that specimens referred to Chasmosaurus represent a single, evolving population, i.e. anagenesis. Rather than forming discrete clusters as expected for reproductively isolated populations, the specimens form a pectinate array, with primitive C. priscus specimens branching off near the base, followed by more derived C. belli specimens, then YPM 2016, until the highly derived C. irvinensis appears.
Stratigraphic distribution of Chasmosaurus specimens is largely consistent with the idea of anagenesis (Figure 13), with more derived specimens tending to occur higher in section. Specimens referred to C. priscus lie low in section in the DPF. C. belli lies above, and finally, C. irvinensis lies at the top7,32.
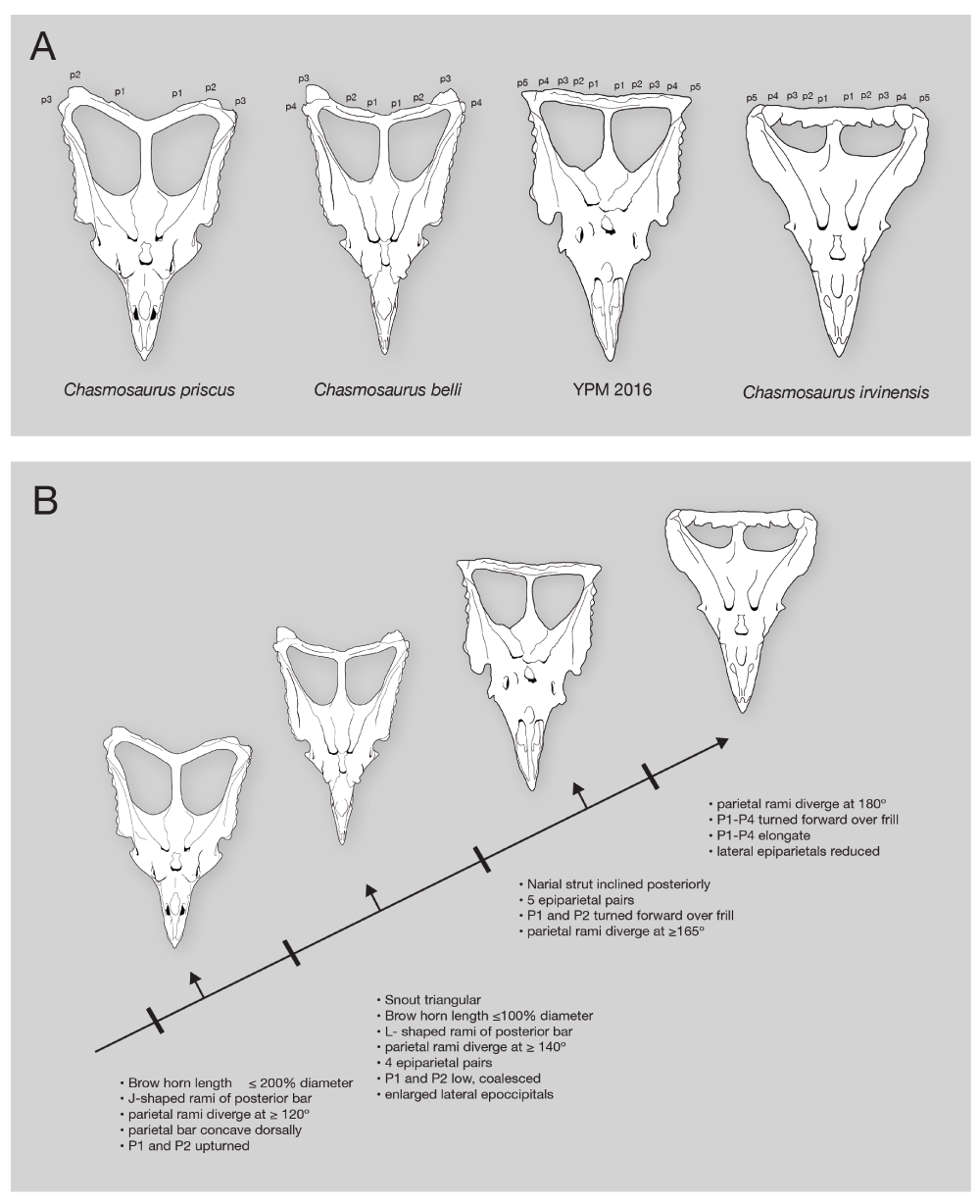
A, Skulls of the three Chasmosaurus species and YPM 2016 compared. Abbreviations: p1–p5, epiparietals 10–5. B, hypothesized transitional series and transformations.
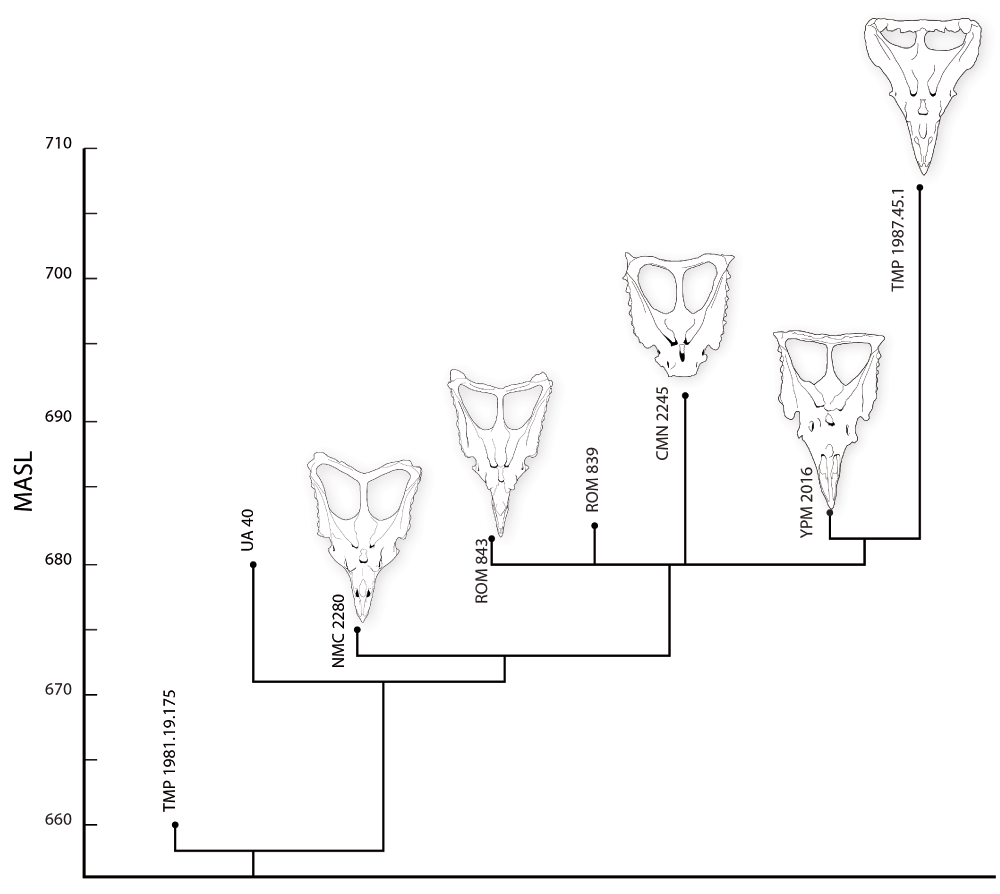
A, Phylogeny of Chasmosaurus specimens calibrated against elevation (meters above sea level, MASL), above, and meters above the Oldman, below. In general, the more advanced specimens tend to lie higher in stratigraphic section, but the stratigraphy of Dinosaur Park is complicated by the fact that the formational contact dips from East to West, and from South to North.
Where YPM 2016 fits into this picture is unclear. In terms of phylogenetic position, YPM 2016 lies between classic C. belli and C. irvinensis morphotypes. If C. irvinensis is the product of anagenesis, then YPM 2016 should also lie between the two in terms of stratigraphy. The original data reported by Sternberg place the specimen relatively high in section, which is consistent with the hypothesis of anagenesis.
However, measurements for Quarry 110 place the site low in stratigraphic section. Assuming this is correct, it implies cladogenesis, not anagenesis. If YPM 2016 lies low in section, then one lineage must split off and ultimately produce C. irvinensis, while one or more morphologically conservative lineages of C. belli persist, before ultimately being replaced by the C. irvinensis lineage at the top of the formation. Thus, Chasmosaurus may not have been a single, evolving population but a collection of many populations evolving more or less in isolation, with one lineage- the one leading to C. irvinensis- ultimately winning out.
As discussed above, Sternberg’s stratigraphic and provenance data are in conflict; YPM 2016 is reported as coming from high in section but Quarry 110 lies low in section. In light of conflicting data, we cannot be certain that the specimen comes from Quarry 110- although additional study of the matrix, palynology, or the quarry might help settle the issue. DPF stratigraphy is complicated, such that elevation and stratigraphic position are not perfect proxies for age. In general, specimens lying higher up in elevation lie higher up in section, but the base of the DPF dips from east to west and south to north32, such that specimens lying at similar elevations might not lie at the same stratigraphic position. Measuring distance from the formational contact is also problematic, for two reasons. First, the contact is not even; even adjacent boundary sections may differ in elevation by almost 10 m (Figure 15), which raises questions about whether the onset of deposition was simultaneous across the park. Second, the thickness of the formation varies34 locally. Because sandstone resists compaction better than mudstones, sequences with large basal sandstones will tend to be thicker than those with more mudstones34. In effect, the compression of the mudstones will compress time.
This might help explain the apparent discrepancy between the original stratigraphic data, putting the specimen 20 m below the Bearpaw, and more recent data which put it 31.4 m above the base of the formation. Additional stratigraphic work will be required to resolve this issue, but is beyond the scope of this paper.
The case of Chasmosaurus provides a case study in the evolution of dinosaurs. Altogether, the Chasmosaurus lineage spans a period of roughly 1.8 million years34. Given this, it would be unsurprising if the lineage adapted to selective pressures exerted by the environment, flora, predators, and other members of the species. One of the more striking changes is seen in the rostrum. In C. priscus (TMP 1981.19.175; CMN 2280) the snout is broadly rounded in lateral view. In C. belli (e.g., AMNH 5402, YPM 2016) and especially C. irvinensis (CMN 41357) the rostrum is more tapered in lateral view, with a more triangular rostral. YPM 2016 and C. irvinensis also differ in having a posteriorly inclined narial strut, which would presumably alter the ability of the beak to resist bending and shearing stresses. Such changes imply changes in dietary preferences and/or feeding strategies. Given that the flora underwent rapid changes during the Campanian63, it would be unsurprising to find that the jaws responded to shifts in floral composition, much as the beak of Galapagos finches adapts to changes in available food64,65. Similar patterns are seen in Triceratops, where changes in beak morphology distinguish the derived T. prorsus from the primitive T. horridus66, with the short-beaked T. prorsus appearing higher in section.17. Insofar as the beaks of extinct dinosaurs (like the beaks of living dinosaurs such as Galapagos finches) evolved rapidly in response changes in diet, dinosaur beaks may be useful for species-level diagnosis44,66, as much or more than the cranial ornament.
The horns in Chasmosaurus also evolved rapidly. The ancestor of Chasmosaurus would have had moderately long postorbital horns, as in Mojoceratops. In C. priscus, the horns are shortened; in primitive C. belli they are reduced to blunt protuberances, and in advanced C. belli and C. irvinensis the horns are replaced by low bosses. Horns in modern animals- including deer, bovids, and rhinos - are used for combat, sometimes against predators, but primarily against conspecifics67 and the same presumably held for horned dinosaurs as well68. Long-horned chasmosaurines presumably engaged in combat by locking their horns together and engaging in wresting and shoving matches68. Reduction of the brow horns suggests that Chasmosaurus did not engage in such bouts; however the robust nasal horn, extended by a horny sheath, would have remained an effective weapon for goring the flanks of opponents or defending against predators.
Finally, and most conspicuously, the shape of the frill and parietal ornament underwent marked changes. From C. priscus to C. irvinensis, the frill evolved from a heart-shaped structure to a subrectangular shield. The epiparietals increased in number; the medial epoccipitals were fused, reduced, then were elongated again. The frill of ceratopsians is likely to have evolved in response to sexual selection69 as a display structure. Sexually selected structures tend to exhibit high variability between species69 which indicates rapid evolution; the rapid evolution of the frill structure can therefore be explained by- and also be considered evidence of- sexual selection.
Taken together, the information suggests that Chasmosaurus represents a lineage evolving through time. Few other examples are known for dinosaurs, although recent work on Triceratops has presented compelling evidence for evolution in this genus17. Although such examples are rare, they are hardly unexpected. Dinosaurs appear to have evolved rapidly, given the profusion of species seen in a short period of time; it stands to reason that dense sampling of a taxon through a single formation could reveal evidence of evolution within that taxon.
Along with previous, specimen-level analyses14–17,66, the results presented here show the advantages of applying phylogenetic analysis to individual specimens. Many studies have previously taken on the taxonomy of Chasmosaurus5,7,8,10,22,70, but consensus has been elusive. The traditional approach has been to focus on supposed key characters such as frill shape8,10,22,70; characters inconsistent with the proposed taxonomy, such as horn length, are then interpreted as intraspecific variation. The problem with this approach should be obvious: how can we know that parietal emargination captures the true signal, rather than horn length, or some other character entirely? With this approach, homology of the characters in question is assumed, instead of tested.
Phylogenetic analysis instead examines all characters and then produces the most parsimonious relationship given the data. Taxonomic groupings and diagnostic characters are therefore inferred from the phylogeny, rather than defined beforehand. This approach is not foolproof: the results will depend on which characters are included, how characters are defined, and depend on accurate coding and identifying informative characters. The advantage of the approach is that it is explicit and repeatable; evidence and assumptions used to classify specimens will be evident, along with any errors.
A potential issue is that ontogenetic changes in morphology could confound the phylogenetic analysis. Ceratopsians experience marked morphological changes as they mature24–26,52, and clustering analysis using ontogenetic characters will group specimens on the basis of maturity, rather than common ancestry26. It follows that either juveniles should be excluded, or ontogenetically variable characters must only be coded for adults. This problem is not unique to a phylogenetic approach; classic approaches to taxonomy are likewise misled when ontogeny is not taken into account (e.g., misidentification of the juvenile Chasmosaurus “brevirostris” as a distinct species).
The use of a specimen-level phylogeny reveals a conflict between evolution and taxonomy. Species are identified by the existence of discontinuities between populations71 either in terms of reproductive isolation or morphological differences71. Yet if evolution proceeds incrementally via the successive accumulation of slight variation71,72, we should expect variation to be continuous, rather than discrete, and the boundaries between species should be blurry.
It follows that when discontinuities do exist such that an extant population is separated from other populations, this must be an artifact of extinction, while discontinuities in the fossil record must be an artifact of sampling72. But where the fossil record is well sampled over a long period of time, we should expect intermediates between recognized species. The endpoints of a lineage, such as C. belli and C. irvinensis, may look distinct, but intermediates such as YPM 2016 will prove difficult to classify, and as additional intermediates are discovered, species boundaries will only become more blurred.
Where the lines are drawn is somewhat arbitrary. One could reasonably place the boundary between C. irvinensis and C. belli such that the Peabody specimen fell into either. Including YPM 2016 in C. irvinensis emphasizes that it forms a monophyletic assemblage with irvinensis specimens to the exclusion of other Chasmosaurus, and would emphasize their apomorphies. Including YPM 2016 in C. belli emphasizes the plesiomorphies shared with that species. All species concepts are ultimately statements about evolutionary lineages73, but to be more specific, all species concepts involve identifying lineages and then ranking them. Biologists or paleontologists must first identify lineages, and then decide whether these lineages differ enough from other segments of the tree to merit rank as a species. Phylogenies are a useful tool for the taxonomist because they allow us to assign organisms to lineages. However, this does nothing to resolve the issue of ranking. The question of how different one segment of a lineage must be from another to merit species or genus rank is still up to the taxonomist, and it remains as much an art as a science, subjective rather than objective, and always will.
The divisions are not, however, wholly arbitrary. As taxonomists, we want our names to apply to real entities. In the Linnaean worldview, these ‘real’ entities reflected the underlying logic of a Creator, who (like his human creations) thought about the world in terms of discrete categories. In a post-Darwinian worldview, groupings such as genera and species reflect common ancestry72. While the boundaries chosen between species may be human constructs, they must reflect real patterns of descent; species and genera must represent groups united by common ancestry74.
Another issue raised by specimen-level analysis is that recognized species and genera may fail to emerge as monophyletic. Two of the three species of Chasmosaurus form paraphyletic assemblages in the current analysis. Instead, the three species lie inside one another like Russian dolls, with C. irvinensis nesting inside of C. belli, which in turn lies inside of C. priscus.
If one advocates a taxonomy based on monophyletic taxa74,75, one can attempt to either split or lump lineages to create monophyletic species. In the case of Chasmosaurus, one approach would be to name the side branches that diverge on the way to C. irvinensis; the Peabody skull would then be given its own species. However, splitting the Peabody specimen off fails to render the remaining C. belli monophyletic (Figure 6C); instead all of the lineages branching off below C. irvinensis must become species to create a monophyletic taxonomy. Furthermore, each time a new specimen was recognized as either intermediate or an outgroup, another species would be necessary, and so on.
Another approach is to lump the specimens and treat the entire clade as one species, C. belli, evolving through time. This approach might better characterize the diversity of the DPF, in that there may have been only one species of present at any given time7. However, it obscures the marked differences in the frill, horns, and jaws within Chasmosaurus, and if more primitive or more derived members of Chasmosaurus were identified, these would have to be included to maintain monophyly, no matter how distinct they appeared. If an even more primitive specimen was discovered- part of the ancestral population that gave rise not only to Chasmosaurus but also to the lineage including Pentaceratops and Triceratops- then it would be necessary to subsume all the genera into one to maintain monophyly.
Splitting or lumping to enforce monophyly may be effective up to a point, particularly when the record is poorly sampled. But both are half-measures and fail to deal with the fundamental problem: enforcing the criterion of monophyly in a classificatory scheme74,75 is inherently incompatible with ranked taxa such as genera and species76,77. The heart of the issue is the existence of ancestors. Species and genera must descend from other species and other genera72. The ancestral taxon, by definition, does not include its descendants, and so is paraphyletic. Ancestors may be difficult to identify with confidence but this does not mean that they did not exist. They must exist if our understanding of evolution is correct, and the completeness of the fossil record is such that ancestor-descendant pairs should be relatively common78.
Among chasmosaurines, potential ancestors include C. priscus as an ancestor for C. belli, and C. belli as an ancestor to C. irvinensis. T. horridus lacks autapomorphies26,66 and T. prorsus is nested within T. horridus17, and therefore may be ancestral to T. prorsus. Titanoceratops ouranos lacks autapomorphies3 and therefore could potentially be ancestral to the remainder of the Triceratopsini. Among modern dinosaurs, the common Canada Goose (Branta canadensis) is an extant ancestral taxon: mitochondrial DNA indicates that various populations of the species are paraphyletic with respect to a Hawaiian clade containing the Nene (B. sandvicensis) and two extinct flightless species of Branta79. The existence of such ancestors means that without abandoning ranked taxa – including genera and species- it is logically impossible to create a complete taxonomy based on the criterion of monophyly76,77.
Finally, the taxonomy presented also demonstrates that the common practice of using autapomorphies to diagnose taxa is impractical. The nature of an ancestral species is that it will be distinguished from its descendants by plesiomorphies. C. belli for example, can be distinguished from C. priscus by apomorphies, but can only be distinguished from C. irvinensis by plesiomorphic characters. Similarly, the features used to distinguish T. horridus from T. prorsus26,66 are all plesiomorphies. Ancestral taxa will be diagnosed by a unique combination of apomorphies and plesiomorphies, rather than a list of autapomorphies.
As shown by the case of Chasmosaurus, specimen-level phylogenetic analysis is a powerful tool for creating phylogenies, which can in turn be used for delimiting and diagnosing species. The best approach to taxonomy is to start with phylogeny; the tree should come first, and names and diagnoses after. The inevitable consequence of this approach is that it will create paraphyletic taxa. A degree of arbitrariness is unavoidable; there is no scientific way to determine how distinct a taxon must be to merit its own genus, and the boundaries between species will only be distinct insofar as our record of evolution is poor.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
I just have a couple of very short question: In the text you mention 69201 MPTs, but I don't understand ... Continue reading Hey Nick, nice to see this already online, very interesting paper!
I just have a couple of very short question: In the text you mention 69201 MPTs, but I don't understand from your formulation if you stopped the tree search once you reached this number, or if these were all shortest trees found by PAUP. Also, in the figure caption of Fig. 6, you mention that you calculated the strict consensus based on 50184 MPTs, how comes?
I'm currently reviewing the several specimen-level phylogenetic analyses published, so I'm particularly interested in these details. I'd be glad to hear your comments about these small issues.
Best,
Emanuel
I just have a couple of very short question: In the text you mention 69201 MPTs, but I don't understand from your formulation if you stopped the tree search once you reached this number, or if these were all shortest trees found by PAUP. Also, in the figure caption of Fig. 6, you mention that you calculated the strict consensus based on 50184 MPTs, how comes?
I'm currently reviewing the several specimen-level phylogenetic analyses published, so I'm particularly interested in these details. I'd be glad to hear your comments about these small issues.
Best,
Emanuel