Keywords
Agricultural intensification, Agroecology, Biodiversity, Ecological intensification, Ecosystem services, Organic agriculture, Yield gap analysis
This article is included in the Ecology and Global Change gateway.
This article is included in the Agriculture, Food and Nutrition gateway.
This article is included in the Food Policy collection.
Agricultural intensification, Agroecology, Biodiversity, Ecological intensification, Ecosystem services, Organic agriculture, Yield gap analysis
Worldwide demand for food, fuel and fiber is expected to increase for at least the next four decades owing to continuing population growth (Godfray et al., 2010). Some potential pathways to address the future gap between supply and demand of agricultural production exist through reducing waste and altering consumer diets (Godfray et al., 2010; Tscharntke et al., 2012), but these may still be insufficient. We will therefore need to increase both land productivity and resource use efficiency of agricultural systems. This challenge for agriculture is set against global environmental changes and increasing pressure on natural ecosystems, some of which are driven by agriculture itself (e.g. forest clearance and reduced carbon sequestration capacity). The consequences of these problems of food production, along with water scarcity and declining energy resources, could produce significant global social unrest and destabilization in the coming years, which has been described as ‘The Perfect Storm’ (Beddington, 2009). Degradation of both land productivity and ecological processes, in turn, creates potentially dangerous feedbacks. Increasing desertification, soil salinization, and water eutrophication, for instance, all present long-term threats to agricultural production and the well-being of communities, significantly displace the boundaries of humans’ safe operating space (Rockström et al., 2009).
To cope with the challenge of ensuring global food security while preserving biodiversity and ecosystem integrity, the ‘ecological intensification’ of cropping systems paradigm has been proposed (Cassman, 1999). Advocates of the ecological intensification of agriculture presently propose two apparently contrasting ‘paradigms’ or perspectives of the future of farming. The first is focused on bridging yield gaps of the major commodity crops worldwide, through targeted breeding strategies of a few crop types, and precise applications of fertilizers and pesticides (Cassman, 1999; van Ittersum et al., 2013). This technology-based perspective renders management options that are simple to implement and reduce farmers’ uncertainty, but also externalize a range of environmental costs (Techno-simplicity). The second emphasizes the role of landscape complexity on biodiversity, supporting the ecological processes that underpin ecosystem services (Eco-complexity), such as pollination, pest control, and soil fertility (Bommarco et al., 2013; Doré et al., 2011). This perspective emphasizes the importance of spatial heterogeneity, especially landscape complexity, to preserve both biodiversity and the ecological functions related to ecosystem services (Chaplin-Kramer et al., 2011; Garibaldi et al., 2011).
Whilst eco-complex management options may be more sustainable than techno-simple practices, they are more complicated to implement, requiring co-ordination among farmers and local authorities, and carry higher levels of uncertainty and risk for farmers. Furthermore, the broader public is keen on preserving heterogeneous rural landscapes, particularly due to their scenic, recreational and cultural values, and complain when intensive agriculture leads to countryside homogenization. Thus, there is an urgent need to conserve biodiversity and rural landscapes, driven in part by public concern, while ensuring food security, particularly in poor countries where key areas of hot-spot biodiversity and severe poverty coincide (Fisher & Christopher, 2007). However, pesticides and transgenic crops are unpopular with the public, especially in some developed countries (Costa-Font et al., 2008), while both biotechnology and precision agriculture are helping to increase food supply and reduce environmental pollution worldwide. For instance, the use of insecticides in cotton has been markedly reduced since the adoption of Bt-transgenic varieties in Argentina and Australia (Qaim, 2009). Hence, attitudes and perceptions of benefits from techno-simplicity and eco-complexity perspectives greatly vary among the different societal actors, with a marked gap between where management interventions are implemented and where the benefits are perceived.
A considerable body of research has highlighted that designing more diverse agricultural landscapes will be crucial for increasing yields and simultaneously conserving biodiversity to ensure food security (Loos et al., 2014). However, the apparent dichotomy between Eco-complexity and Techno-simplicity has led to an undue focus on how small, uncropped patches in landscapes can be better managed. Fields have been left to techno-simple solutions and the constraints of landscape research have limited the scales at which eco-complex management options can be tested. As a result, management actions for increasing diversity within the field are uncommon and, where they do exist, they have not been widely adopted and have had equivocal effects.
Here, we argue that integrating Techno-simplicity and Eco-complexity perspectives would deliver environmentally-friendly, intensive and food-secure agro-ecosystems, through the provision of more tractable management options for farmers. We stress that bringing about change of practices in intensified cropping systems is about making small changes to usual farming methods. In practice, each individual change may only lead to small gains in biodiversity. However, when actions are combined and implemented across large areas, marked increases in biodiversity and ecosystem services may emerge, expressed at broader spatial scales and over longer periods than a single growing season. We also claim that the knowledge needed is already available (Shennan, 2008), but needs to be properly integrated into intensively managed cropping systems to effectively design more multifunctional agricultural systems. We argue that achieving future food security through integrating eco-complex and techno-simple perspectives will require reconciling goals, costs and benefits perceived by social actors across scales, including farmers, city dwellers, practitioners and policy makers.
Modern agriculture has succeeded in increasing global food supply. This has been achieved through targeted breeding of a few, highly productive crops, providing resources through fertilization and irrigation, protecting crops by applying pesticides and, more recently, by genetic modifications to crops. This agricultural intensification has led to declines in biodiversity due to habitat loss, farmland homogenization and environmental pollution (Tscharntke et al., 2005). To overcome these negative environmental impacts, the paradigm of ecological intensification was initially coined to promote a site-specific, precision agriculture. This would increase yields of the main cereal crops, ensuring future global food security, while simultaneously reducing both soil degradation and pollution, derived from plowing and excessive fertilization (Cassman, 1999). This techno-simple perspective increased yields of the major commodity crops worldwide through breeding a handful of crop types and being highly precise in the applications of fertilizers and pesticides, but reducing crop diversity at local, regional and global scales (Khoury et al., 2014). Similar crop yields have been achieved in GM herbicide-tolerant varieties, where herbicides applications decline significantly in comparison to counterpart conventional crops (Champion et al., 2003). Ultimately, these crops were rejected in Europe both because of public concern and measurable impacts on biodiversity (Bohan et al., 2005), despite demonstration that these biodiversity effects could be reversed through modified management, such as herbicide timing (Dewar et al., 2003). However, in the Americas, where glyphosate-resistant crops have been rapidly adopted, biotypes of some major weeds have evolved resistance to this herbicide (Powles, 2008).
Techno-simplicity is, thus, directed at improving crops and their environment to bridge the gaps between actual and potential yields, through advancement in plant ecophysiology, molecular biology and soil science. As originally conceived, the ecological intensification paradigm is the latest reconceptualization of a long history of empirical research on resource use efficiency in agriculture (Cassman, 1999; de Wit, 1992; van Ittersum et al., 2013). The conceptual framework applied is based on yield gap analysis, which is aimed at identifying the key environmental and management factors defining the gaps between potential and actual yields (see Box 1). However, the techno-simple perspective of ecological intensification has not given proper attention to biodiversity and its benefits, through associated ecological processes and functions regulating the incidence of weeds, pests and diseases.
Crop yields, measured as the amount of harvestable production per land area unit (kg ha-1), are determined by factors influencing crop cycle length, resource use, growth rates and reproductive allocation (van Ittersum & Rabbinge, 1997; van Ittersum et al., 2013). Yield-defining factors determine the maximum yield levels (potential yields) for a given crop grown in a particular site, with no water and nutrients limitations, and effective control of biotic factors reducing yields. Potential yields are defined, on the one hand, by environmental factors that are location-specific, such as the growing season length (frost-free period), atmospheric CO2, and the annual variability in incoming solar radiation, day length, and temperature. On the other hand, potential yields are also determined by crop plant (genotypic) characteristics, such as developmental rates (type of response to day length), growth rates (photosynthetic pathway), and plant morphology and architecture (growth, reproductive allometry). Both environmental and genotypic attributes of crops can be managed to some extent in regular farming conditions. Hence, farmers make tactical decisions to maximize the use of resources available during the growing season by choosing well-adapted crop varieties and optimum sowing dates, thus determining the growing period to maturity, planting density and spatial arrangement.
Yield-limiting factors include water and nutrients, which determine the water- and nutrient-limited yields (van Ittersum & Rabbinge, 1997; van Ittersum et al., 2013). Water-limited yields are usually obtained from crops grown under rain-fed conditions, where water limitations result from climate (annual rainfall, dry season, evapotranspiration, vapor pressure deficit) and soil factors (runoff, infiltration, retention capacity, soil depth). Nutrient limitations are due to soil characteristics that may affect both soil nutrient availability and crop uptake efficiency, such as the organic matter content, soil reaction (pH) and salinity. Soil biotic activity may also influence nutrient availability for crops. Yield-reducing factors comprise biotic agents, such as weeds, pests and diseases, and abiotic stressors, such as pollutants, which reduce yields by decreasing or hindering either resource use, growth rates or reproductive partitioning. Actual yields at farm level result from combinations of limiting and reducing factors, which are defined by the actual supplies of water and nutrients and protection levels against biotic adversities. Farmers can increase actual yield levels by taking measures to overcome the effects of factors limiting and reducing yields. Yield-increasing measures consist of providing resources; water through irrigation and nutrients by applying synthetic fertilizers or managing soil fertility and structure (composting, green manuring) (Table 1, van Ittersum & Rabbinge, 1997). Yield-protecting measures are aimed at preventing and reducing the incidence of biotic factors reducing crop growth and yields (Table 2). These may involve direct control with specific tools or management strategies.
Supporting and regulating ecosystem services in agro-ecosystems closely interact with the measures farmers take for increasing and protecting yields. Supporting services are related to the biogeochemical processes governing organic matter decomposition, mineral and carbon cycling, soil porosity and structure. Although fertilization is the main practice against nutrient limitations, yield-increasing measures may involve strategies to sustain or enhance soil organic matter, such as diversified crop rotations (see Table 1 and Table 2, Bommarco et al., 2013; Shennan, 2008). These strategies are exclusively applied at field scale from a single growing season to several years. Regulating services have been mostly associated with the biological control of insect pests through predation and parasitism (Bommarco et al., 2013). Unlike supporting services, the influence of spatio-temporal heterogeneity (i.e. landscape complexity) is critical to ensure the provisioning of regulating services. Hence, most initiatives to promote regulating services in agro-ecosystems emphasize the role of semi-natural habitats for retaining biodiversity, thus promoting ecological processes in farmland mosaics. Moreover, abundance and activity of populations providing regulating services may closely interact with agrochemicals controlling target biotic adversities. Animal pollination, though considered a regulating ecosystem service, would actually be an essential yield-defining factor because it determines seed- and fruit-setting in many crops, and consequently crop yields.
Other management practices may involve a combination of both.
Other management practices may involve a combination of both.
The high crop yields currently achieved in intensively managed agro-ecosystems are the successful outcome of a long process of techno-simplification of natural ecosystems. This process imposed increasing levels of spatiotemporal homogenization through standardized management and specialization in cropping systems (Figure 1). Hence, natural vegetation was cleared to grow crops, which promoted some desirable species while many others were controlled. Then, proportions of crop harvest and fallows intervals in the growing season were gradually reduced, whereas the natural regeneration of grasslands and forests was replaced by seeding and planting. In addition, recurrent disturbances during the growing season, such as plowing, maintained early successional conditions that promoted short-lived life-forms to the detriment of perennial ones. Moreover, many ecological functions have been progressively substituted by artificial inputs, such as the use of synthetic fertilizers in place of nutrient cycling by the soil biota (Figure 1). Therefore, achieving high monoculture yields relies on breeding high-yielding varieties and using high levels of petrochemical inputs, which consequently maintain biodiversity at low levels in intensively managed agro-ecosystems (Figure 1). Accordingly, croplands alone currently account for nearly half of human appropriation of global net primary productivity (Haberl et al., 2007). A substantial part of productivity, which in the past was biodiverse, has been transferred to (low diversity) crop yield productivity by farming intensification. Biodiversity has been actively selected against in these systems and has declined (Figure 1). Species composition and trophic structure have changed, thus modifying ecosystem processes of availability, uptake and use of soil resources, such as water and nutrients, among biotic communities (Tscharntke et al., 2005).
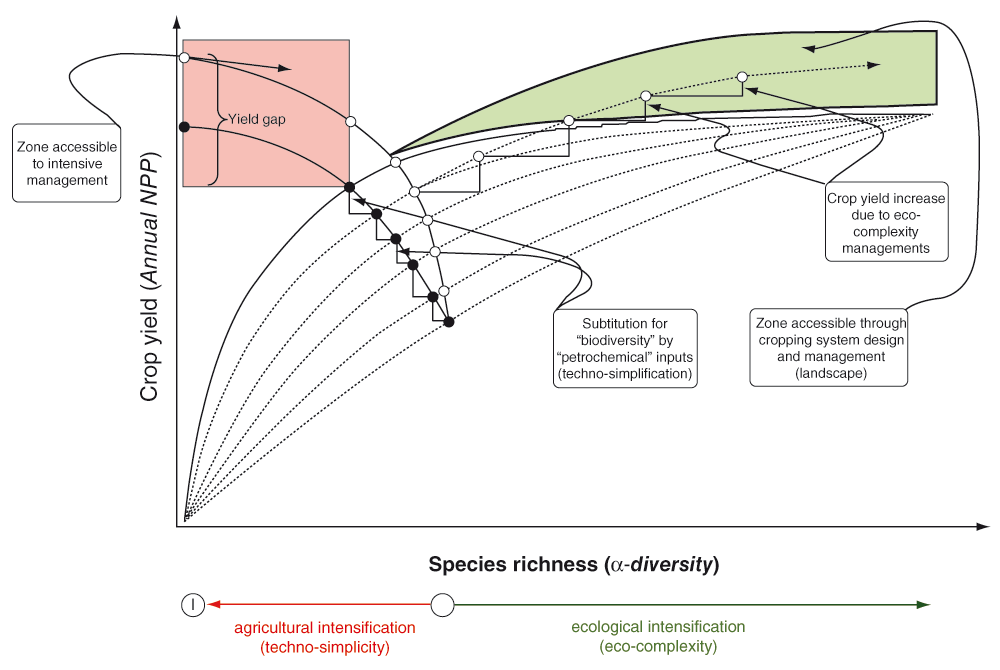
The “zone accessible to intensive management” (light red area on the upper left hand side) corresponds to intensively managed agro-ecosystems where high productivity levels can be achieved at very low species richness (ideally only one species in monocultures), but through substituting ecosystem functions (nutrients provided by SOM, biological regulation of pests) for the use of off-farm inputs (fuels, fertilizers, pesticides, and/or water resources; see Box 1 for further details). Curves in red on the left hand side are yield-input production functions showing how crop yields increase as more inputs are applied and species diversity decreases (i.e. agricultural intensification increases as techno-simplicity progresses) in rain-fed (solid line) and irrigated (dashed line) cropping systems (modified from Cassman, 1999). There is also a “crop yield zone accessible through the design of more diverse cropping systems or complex farmland mosaics” (light green area on the upper right hand side), which rather relies on ecological processes occurring across different spatial and temporal scales (i.e. by implementing eco-complexity measures; see Table 1 and Table 2 in Box 1 for further details).
The importance of spatial heterogeneity, particularly landscape complexity, for preserving both biodiversity and ecosystem services has been the focus of many recent studies (Chaplin-Kramer et al., 2011; Garibaldi et al., 2011; Tscharntke et al., 2012). The seminal paper by Thies & Tscharntke (1999) was one of the earliest to empirically demonstrate that landscape structure is highly influential for mobile arthropod species in farmlands. Since then, numerous studies have shown that landscape complexity, in terms of the amount and configuration of habitat patches, can influence species dynamics and ecosystem services (e.g. Gagic et al., 2012; Thies et al., 2011). However, invertebrate pest species may be much more variable than other invertebrates, such as natural enemy taxa (Chaplin-Kramer et al., 2011; Jackson & Fahrig, 2014). As a consequence of this body of research, most biodiversity management initiatives in agro-ecosystems emphasize the role of non-crop habitats for retaining biodiversity and ensuring the provision of ecosystem services. This viewpoint advocates preserving, reintroducing and restoring semi-natural habitats to increase landscape complexity and, consequently, promote ecosystem services. This requires land-use changes at farm and landscape levels and, potentially, co-operation between landholders (landscape management in Figure 2). However, there is a growing appreciation that for certain species, local management changes may also show some benefits and, theoretically, they can be implemented at much smaller spatial scales (patch and field levels in Figure 2) (Gonthier et al., 2014).
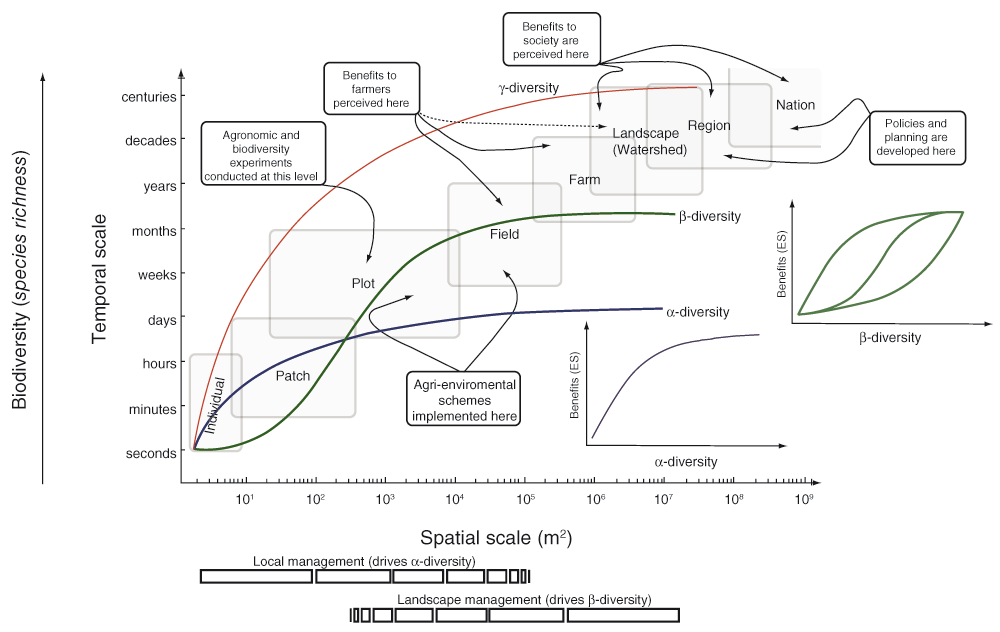
The two hypothetical relationships between management options and the benefits that may accrue are identified at the local (alpha diversity-driven) and landscape (beta diversity-driven) scales of implementation. Changes in biodiversity (species richness) at landscape (gamma, red line) and local (alpha, blue line) scales and species turnover (beta, green line) are also shown as a function of spatial scale. The range of implementation scales are shown in the light grey boxes (patch/plot – field – farm – landscape/watershed). Scales setting the upper (region/nation) and lower (individual plant/plant part-insect interactions) for implementing management options are depicted in grey boxes. Different beneficiaries are also identified. Farmers usually perceive the benefits at the same level at which they make decisions and do agriculture, field and farm levels, while society perceives the benefits from the landscape/watershed level to the regional and national levels.
Ecosystem functions derived from complex interactions among different species and their environment lie at the heart of ecology. However, the application of these concepts to agro-ecosystems, which are heavily influenced by intensive management, has only recently begun (Scherr & McNeely, 2008). Over the last 50 years the obvious and sustained loss of biodiversity in agricultural landscapes (Geiger et al., 2010) has given new impetus to the study of the relationship between species diversity and important ecosystem functions they support.
Theoretically, greater species diversity across a range of taxa has been shown to improve the way communities capture resources and convert these to biomass, thereby increasing productivity (Cardinale et al., 2012). Empirically, crop yields can be increased by intraspecific genetic diversity and crop diversity (Li et al., 2009). Diverse mixtures of native perennial grasses can produce more biomass for biofuel production (Tilman et al., 2006). A long-term grassland study showed that primary productivity in highly diverse plant communities has greater resistance to, and recovers more fully from, a major drought (Tilman & Downing, 1994). Species diversity has also shown to be important for a range of provisioning services. For instance, we know that herbivory can be reduced at higher crop diversity levels, while pest mortality can be increased at higher trophic diversity levels (Wilby & Thomas, 2002). However, whether this relationship is due to species diversity per se, or the result of species abundance and resource partitioning is dependent upon the system or the context (Northfield et al., 2012). There is consistent evidence that maintaining wild pollinator diversity improves pollination outcomes (Garibaldi et al., 2013), but for many other ecosystem functions the link to diversity is often unclear.
While complexity has been proved valuable at a range of spatial scales, methods for introducing complexity into intensive agricultural landscapes are still rare. Proponents of organic farming, which excludes synthetic pesticides and fertilizers, have long considered their systems to support more complex and diverse communities. Studies are starting to provide evidence supporting this theory, at least for some taxa (Fuller et al., 2005). Crowder et al. (2010) showed that organic farming promotes evenness of communities and enhances pest control. On just one organic farm, 1501 unique interactions among 560 plant and animal taxa have been shown (Pocock et al., 2012). Moreover, a few studies have demonstrated that organic farming can change the structure and function of interacting species networks (Lohaus et al., 2013; Macfadyen et al., 2009). Organic farming is typically adopted across an entire farm and consequently implements many different managements in a holistic way, going beyond the piecemeal implementation of individual managements that is typical in conventional agriculture. However, studies disentangling the influence of different organic management options suggest that some managements may be very much more influential than others (Puech et al., 2014).
Complexity of agricultural systems is beginning to be embraced and measured by researchers (Pocock et al., 2012). Eco-complexity management recommendations (see Table 1 and Table 2 in Box 1), which are derived from these research studies, emphasize the complex nature of agro-ecosystems and rely on the functioning of naturally occurring species. Some recommendations are intrinsically difficult and costly for individual landholders to adopt. Their implementation should be carried out at large spatial scales to make their full benefits evident (Figure 2). Despite those difficulties for landholders, many of these management options are attractive to consumers as they tend to have less negative environmental impacts over the long-term, and they also provide a marketing niche for some farmers.
Many management recommendations derived from eco-complex farming systems would be feasible in techno-simple cropping systems. Excessive emphasis has been put on agro-ecosystem complexity rather than on the simplicity of management interventions required to support complexity. For example, in many agricultural landscapes, we know that both pests and natural enemy species resort to various plant species throughout their life-cycle. Often a recommendation is made such as “greater diversity of native plant species in a landscape will increase natural enemy species diversity.” Although this may be technically correct, achieving this objective in highly altered agricultural landscapes will demand significant investment by landholders. A more manageable approach may be to identify a few key plant species supporting relatively more pest species than natural enemy species, thus targeting them for removal from semi-natural patches (Storkey et al., 2013). Similarly, in the cropping parts of landscapes, simple management recommendations can introduce significant differences in biodiversity, such as cover crops and intercropping (Shennan, 2008).
Designing policies and initiatives for conserving biodiversity in agro-ecosystems, as well as encouraging environmentally friendly farming, are strongly influenced by the social, not purely economic, attitudes of people towards biodiversity and agriculture, which may differ starkly between city dwellers and landholders. On the one hand, urban consumers usually express positive attitudes towards organically grown food, which are associated with environmental concern and a rejection of GM food (Dreezens et al., 2005). These attitudes are usually based on emotion, rather than scientific knowledge (Martín-López et al., 2007), such as the appreciation of charismatic and useful species as well as the aesthetic and cultural values of landscapes (Martín-López et al., 2007). On the other hand, farmers are keen to adopt technological innovations aimed at facilitating crop management and increasing yields, such as transgenic crops (Qaim, 2009), while their willingness to implement conservation schemes is influenced by many factors interacting in complex social and natural networks (Siebert et al., 2006).
Relationships between both the goals and costs of implementing eco-complex management options, as well as the benefits that may accrue, can be described at the local (alpha diversity-driven) and landscape (beta diversity-driven) scales of implementation (Figure 2). Techno-simplicity and eco-complexity perspectives may appear distinct, from a consideration of their goals and approaches to achieve ecological intensification. However, a synthesis of both can be created because they are applied in the same systems and face trade-offs between costs and benefits across similar scales of management implementation and outcomes perception (Figure 2). Articulating goals, costs and benefits of any management intervention will be, therefore, critical for designing farming systems that synthesize both techno- and eco- perspectives for ensuring food security. The conceptual framework needed to articulate these three components can be articulated with insights that have been made from integrating different management strategies and tactics involved in crop protection systems (Cardina et al., 1999; Irwin et al., 2000). To be operational, such a synthetic framework should also properly integrate processes and contexts at different space and time scales (i.e., biological, ecological, agricultural, socioeconomic, and political domains).
Practically, within our synthetic framework, the main components can be conceptually defined for any management intervention (whether agronomic, restoration or conservation). Firstly, both the space and time scales in which interventions are applied should be specified (Figure 2). For instance, herbicides to control weeds are applied to the entire field area, while grass strip schemes are installed only along field margins. Secondly, the scale at which the expected benefits from a particular intervention are effectively perceived should be identified. Hence, actions applied at a particular scale may express their beneficial effects directly or indirectly at the same or different scales. Finally, implementing any intervention has a cost, which can vary across scales, not only in monetary terms, but also in the complexity and nature of tasks involved and the time required to be implement (e.g. installing a grass strip along the margin of a particular field vs. implementing grass strip schemes in a region).
Designing more diverse cropping systems is crucial for increasing yields and, simultaneously, conserving biodiversity to ensure the functioning of agro-ecosystems and improvements to future food security. We argue that space and time dimensions require special attention. Doing so would allow us to synthesize a more comprehensive and practical ecological intensification from the techno-simple and eco-complex paradigms. We highlight that the knowledge needed is already available in the different disciplines of agroecology, crop ecophysiology, food-web ecology and landscape ecology, to build a more integrative and effective cropping system design. This could lead to the design of inter-connected habitats, which contain both managed and naturally-occurring species and their services and are thereby multi-functional. Moreover, we argue that developing eco-complex agricultural systems requires proper articulations of goals, costs and benefits of any management intervention. This demands the integration of processes and contexts at different space and time scales. Giving special attention to the social actors involved at different domains, such as farmers, city dwellers and policy makers is also relevant. Effective communication among these social actors about their perceptions of goals, costs and benefits is fundamental for succeeding in the implementation of any program aimed at achieving future food security through reconciling eco-complex and techno-simple perspectives.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)