Sexual reproduction is essential for many organisms to propagate themselves. It requires the formation of haploid female and male gametes: oocytes and sperms. These specialized cells are generated through meiosis, a particular type of cell division that produces cells with recombined genomes that differ from their parental origin. In this review, we highlight the end process of female meiosis, the divisions per se, and how they can give rise to a functional female gamete preparing itself for the ensuing zygotic development. In particular, we discuss why such an essential process in the propagation of species is so poorly controlled, producing a strong percentage of abnormal female gametes in the end. Eventually, we examine aspects related to the lack of centrosomes in female oocytes, the asymmetry in size of the mammalian oocyte upon division, and in mammals the direct consequences of these long-lived cells in the ovary.
All oocytes undergo induced arrest at the dictyate stage of prophase I during meiosis in the ovary. This arrest takes place after chromosome pairing and crossing-over formation between parental chromosomes. It can last months in mice and decades in humans. Upon hormonal surge, oocytes will exit the prophase I arrest and resume meiosis. All stages from meiosis resumption, starting with nuclear envelope breakdown (NEBD) until the next arrest where oocytes are fertilized, belong to the meiotic maturation process (Figure 1). This process terminates meiosis, allowing the gamete to go through two successive rounds of extremely asymmetric divisions in size. Between these divisions, there is no intervening DNA replication, truly rendering the gamete haploid after, at, or before the second meiotic arrest at fertilization. Indeed, depending on the species, this second arrest will take place at different cell cycle stages: coincident with meiosis resumption at NEBD in nematodes, in metaphase I in Drosophila, in metaphase II for most vertebrates, or after the end of the second meiotic division in starfish oocytes.
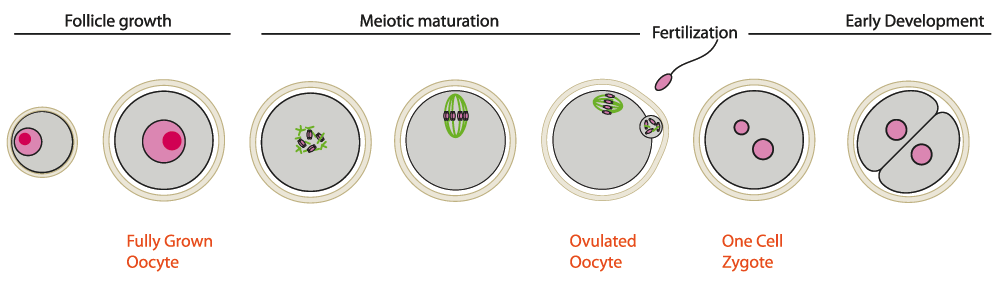
Figure 1. Meiotic maturation and first stages of embryo development in mammals.
Meiotic maturation starts with nuclear envelope breakdown (NEBD) and is followed by the first meiotic division where bivalents are separated, the first polar body is extruded and then, in vertebrates, arrest in metaphase of the second meiotic division (MII stage) occurs. The oocyte is ovulated at the MII stage. Sister chromatids will be segregated after fertilization. Zygotic development follows fertilization. Oocytes (in gray) are surrounded by a protective glycoprotein layer, the zona pellucida (beige). DNA is in pink, microtubules in green.
There are three essential features that we would like to highlight in this review: (1) meiotic divisions take place in the absence of centrosomes in most animals; (2) these divisions are highly asymmetric in size to produce large oocytes; and (3) in mammals, oocytes display an extreme longevity in the ovary.
We would like to propose that these three features, among other things, predispose oocytes to errors in chromosome segregation. In mammals, this aspect has been particularly well studied, especially in human oocytes, which present a basal rate of errors close to 20% in women younger than 35 years of age and which can be as high as 60% in older women1–3. Indeed, it remains puzzling that given the extreme parental investment in juvenile care in many species, such little attention is being paid to oocyte ploidy, as if errors in chromosome segregation were part of a selection process for gamete fitness.
Acentriolar divisions
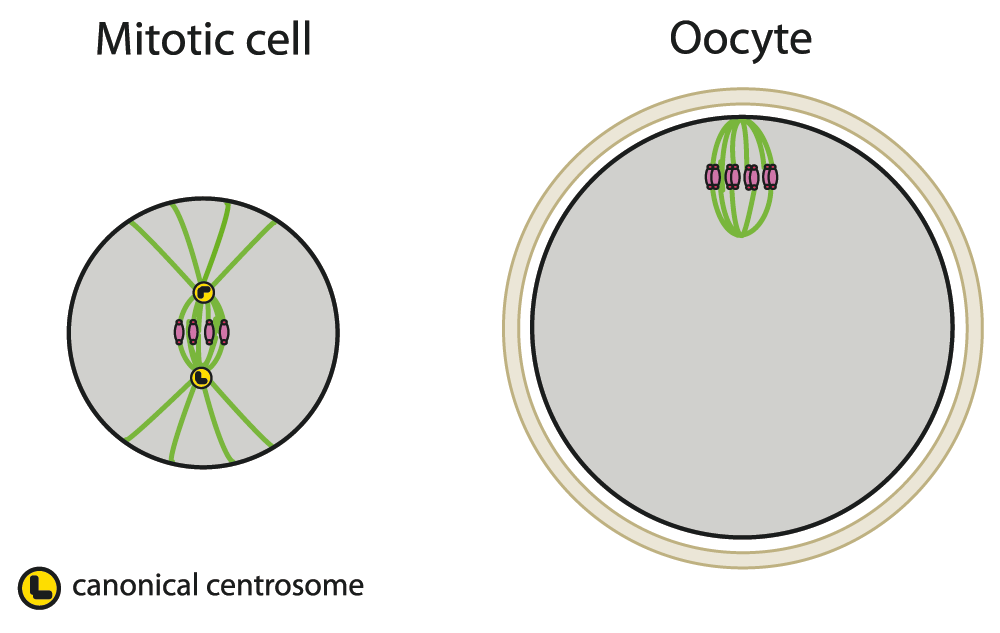
Centrosomes, consisting of a pair of centrioles surrounded by a cloud of pericentriolar material (PCM), are the major centers for microtubule assembly (microtubule-organizing center). Most oocytes lose their centrioles during oogenesis; if not, like in starfish, they are progressively eliminated and inactivated during meiotic divisions4–8. Even though centrosomes are not strictly required to segregate chromosomes (as shown in flies, planarians, or mice 9–12), they contribute to the coordination of spindle assembly and increase its robustness. The immediate consequence of centrosome loss is that meiotic spindles are devoid of astral microtubules and so lack the main connector between the spindle poles and the cell cortex (Figure 2). Hence, spindle positioning cannot rely on astral microtubules as it happens in most somatic cells13–16. Furthermore, centrosome-nucleated microtubules cannot capture chromosomes14. In mitotic cells, duplicated centrosomes are positioned on opposite sides of the nucleus such that in prometaphase the spindle axis is already set17–20. On the contrary, meiotic spindle bipolarity is not predefined by the position of the two centrosomes. Instead, during meiosis I, spindle bipolarity is progressively established and this can take about 40 minutes in Drosophila, 3 hours in mouse, and up to 6.5 hours in human oocytes21–25. Not only is meiotic spindle bipolarity an extremely slow process in meiosis I but it appears that assembly of K fibers (microtubule bundles that connect the kinetochores of chromosomes) is quite long: 50 minutes in Drosophila, 6 to 7 hours in mice, and about 16 hours in humans26–28. The biological significance of such a progressive K fiber assembly during oocyte meiosis I remains unknown, but it is clear that in both Drosophila and mice precocious stabilization of K fibers is deleterious for bivalent alignment, orientation, and segregation28,29.
Figure 2. Oocytes assemble and position their spindle in the absence of centrosomes.
Cells are in gray, and oocytes are surrounded by a protective glycoprotein layer, the zona pellucida (beige). DNA is in pink, microtubules in green, centrioles in black, and pericentriolar material (PCM) in yellow.
Oocytes use an inside/outside mode of spindle assembly, first promoting the assembly of microtubules around chromatin and then defining the spindle poles25,30–33. As a result, meiotic spindle poles in oocytes appear less robust, not being anchored into unique and well-defined centrosomes. In some species, like Drosophila, nematodes, Xenopus, and even human oocytes, microtubule minus ends at spindle poles are not even connected or anchored to discrete PCM foci25,34–38, unlike in rodents25,34–38. The lack of anchoring raises issues not only on spindle pole organization and maintenance but also on the nature of the force integration that allows all chromosomes to end up midway in between both poles, on the metaphase plate. As a consequence of having poles formed by more than one entity, pole integrity can be compromised and splitting of poles may occur, as in cancer cells with extra-centrosomes presenting unbalanced poles composed of multiple centrosomes23,31,39. It may not be so surprising that the rate of chromosome mis-segregation in oocytes is very high compared with most somatic cells in the presence of non-equilibrated spindle poles, which favor merotelic attachments not detected by the spindle assembly checkpoint (SAC)40 as shown in cells with extra-centrosomes41,42.
In addition to lacking robust spindle poles, many oocytes use actin-based propulsion forces to position their chromosomes43–50. In starfish oocytes, an actin fishnet is transiently formed at meiosis resumption, prior to the microtubule capture, to maintain all chromosomes spatially confined, avoiding their dispersal in the huge volume of the nucleus, beyond the reach of microtubules51–53. In mitotic cells, even though astral microtubules dictate the orientation of the spindle apparatus, they are also connected to F-actin, which helps transmit forces exerted by the cell environing tissue54–57. In mitosis, microfilaments cooperate in spindle assembly: they help separate the two centrosomes in prometaphase, hence promoting spindle bipolarization58, and they modulate spindle orientation and favor spindle assembly through mitotic cell rounding59. However, in mammalian oocytes, where spindle bipolarization in meiosis I can take hours, it seems important that microfilaments do not interfere with spindle assembly and are thus nucleated around the microtubule spindle only once the latter is robust enough60. It was very recently shown that the centrosome can also be a major filament-organizing center (it could be named an FTOC)61. The centrosome thus acts as a coordinator of microtubule and microfilament networks inside the cytoplasm. In oocytes, this coordination is missing. Therefore, it is conceivable that other mechanisms have emerged to avoid premature interference between the two meshes and also to modulate their interaction.
Extremely asymmetric divisions in size
Oocytes undergo extremely asymmetric divisions, leading to the formation of a large cell, the oocyte, and two minuscule polar bodies. This size asymmetry is essential to maintain the maternal reserves accumulated during oogenesis in order to sustain embryo development. For this, they rely on very asymmetric spindle positioning. In the case of the Xenopus oocyte, 1 mm wide (Figure 3), the asymmetry is clearly extreme where one spindle pole is anchored at the cortex while the other pole cannot reach the opposite cortex (the spindle being approximately 30 μm long). The asymmetric anchoring of the meiotic spindle to the cortex generates a strong imbalance of the forces experienced by each spindle pole, converted into asymmetric forces exerted on the chromosomes. How do oocytes achieve the equilibrium of tension on both sides of each bivalent (meiosis I) or univalent (meiosis II)? Moreover, when somatic cells enter mitosis, they round up and their cortical tension increases and this helps to equilibrate forces coming from each spindle poles to the chromosomes54,62–64. Unexpectedly, mouse oocytes experience a drop in cortical tension during meiosis and this is absolutely necessary for spindle positioning as well as for the asymmetry of the division65–67. One can easily understand that a soft and deformable cortex favors the extrusion of polar bodies tailored to the chromatin mass better than a stiff cortex, as in mitosis. However, it is difficult to conceive how spindle microtubules can transmit and propagate the tension to chromosomes when their poles are not symmetrically anchored and when one pole is actually anchored on a soft material. One has to imagine that pushing or pulling forces might be transmitted more locally or maybe via yet-to-be-discovered structures/mechanisms inside the meiotic spindle. In worm oocytes, a solution has emerged with extensive meiotic spindle pole depolymerization at anaphase I and with most microtubule forces required to separate bivalent chromosomes coming from local microtubule assembly at the chiasmata, allowing the chromosomes to be pushed apart68.
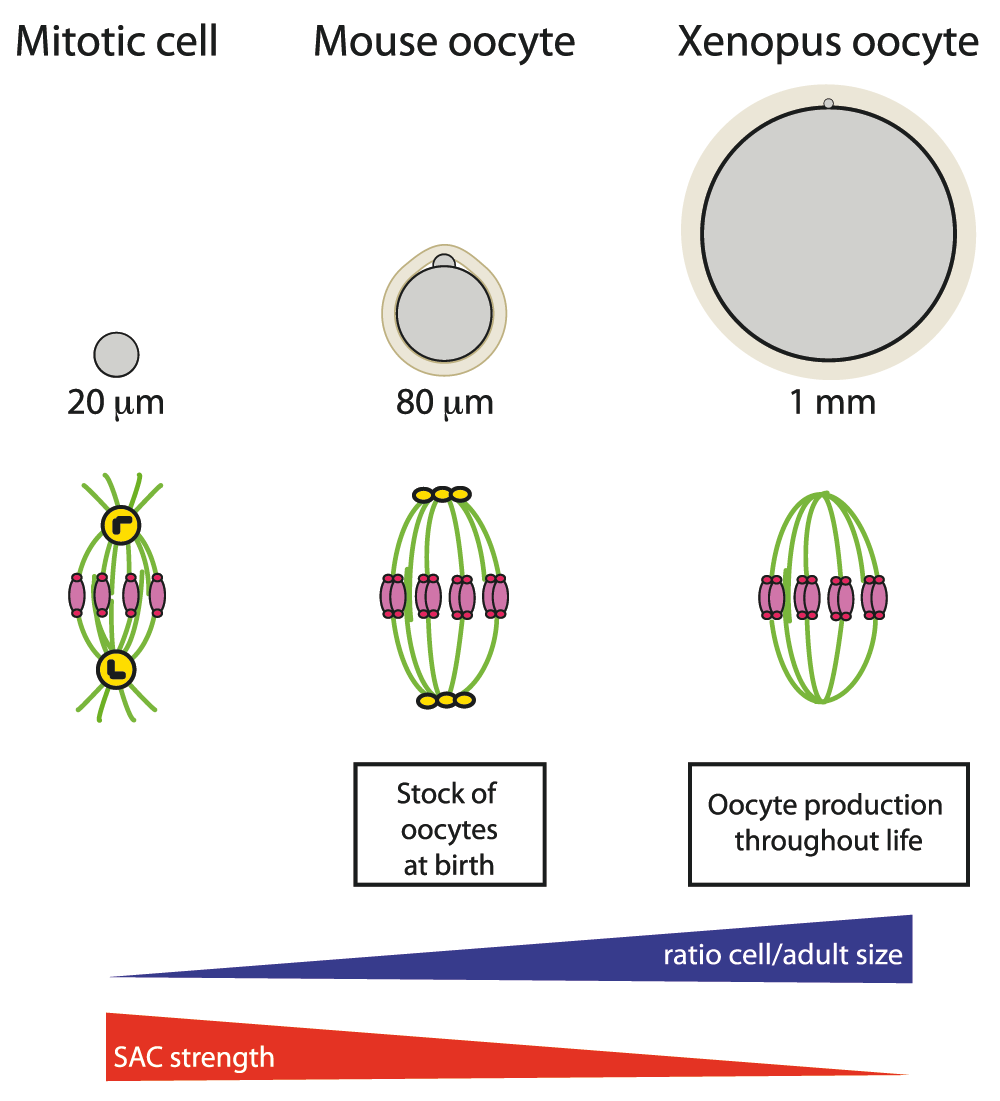
Figure 3. Spindle assembly checkpoint strength in different cells.
Cells are in gray and oocytes are surrounded by a protective glycoprotein layer, the zona pellucida (beige). DNA is in pink, kinetochores in dark pink, microtubules in green, centrioles in black, and pericentriolar material (PCM) in yellow.
Another feature which characterizes oocytes is the poor sensitivity of the SAC to errors in chromosome alignment or to a global drop in tension exerted on bivalents69–73. In nematodes and Xenopus oocytes, there is no SAC response, and no cell cycle arrest is observed in Caenorhabditis elegans mutants with severe meiotic spindle defects or after complete microtubule depolymerization in the frog74–77. Similarly, mutations in multiple SAC genes do not affect cyclin B levels or chromosome segregation in Drosophila oocytes78. In contrast, SAC-deficient Drosophila neuroblasts, genetically modified to lack centrosomes, present a higher incidence of chromosome segregation errors than acentrosomal neuroblasts with a functional SAC. This shows that, in Drosophila mitosis, a functional SAC is required, in the absence of centrosomes, contrary to what is observed in oocytes79. Interestingly, all three of the above species assemble meiotic spindles without discrete PCM foci at their poles and this might contribute to the absence of a SAC response (Figure 3).
As suggested by pioneering work from Xenopus early development, the origin of the poor SAC response in oocytes might come from their large size (Figure 3)80. The SAC signal, which inhibits the activation of the anaphase-promoting complex/cyclosome (APC/C) and thus the degradation of two key substrates, cyclin B and securin that trigger the metaphase-to-anaphase transition, is produced by unattached kinetochores and might be diluted in the large cytoplasmic volume. It will be very interesting to reduce oocyte size and see whether this restores a mitotic-like SAC response. Alternatively, it may not be strictly the oocyte size per se, but rather its size with respect to the dimensions of the adult female (Figure 3). This ratio might relate better to the amount of energy invested by the species in its reproductive capacity. It could explain why, in nematodes and Drosophila, organisms a thousand times smaller than a mouse but producing eggs of comparable size as mammals, there is no SAC response during oocyte meiosis, as in Xenopus oocytes that lay eggs 12.5 times larger than mouse eggs for a comparable adult body size.
Exacerbated longevity of mammalian oocytes
Thanks to a renewable population of germ cells that supports gametogenesis in their ovaries, nematodes, Drosophila, and amphibian females produce oocytes during their whole life. On the contrary, eutherian mammals possess a finite reserve of germ cells that are formed and stored during embryogenesis. The different reproductive strategies used by these model organisms might also explain the differences in SAC sensitivity (Figure 3). Rapidly spawning a lot of eggs at the right season might have been selected to allow frog dissemination at the expense of gamete quality production. In contrast to species that lay eggs or embryos in the external milieu, in mammals, the longevity of oocytes from birth to ovulation can reach decades. This raises issues about chromatin architecture maintenance to sustain such a long-lived metabolism, in particular for the turnover of key elements involved in chromosome segregation. Meiotic spindle morphology is altered, with poor chromosome alignment and split poles in aging human oocytes obtained from normal naturally cycling women81. Also, it has been clearly established that the amount of maternal mRNA encoding for genes involved in the SAC response, spindle integrity, and spindle positioning decreases in aged mouse oocytes82,83. More importantly, the number of proteins maintaining chromosome pairing reaches a critical low level in aged mouse and human oocytes, which impedes the integrity of chiasmata84–86. Furthermore, artificially abolishing one key meiotic cohesin, SMC1β, by gene targeting already has profound effects on the integrity of bivalents, and aged oocytes deficient for SMC1β display evidence of chiasma terminalization87. The reduction in cohesin levels can be attributed to their lack of turnover. Indeed, genetic studies aimed at assessing the turnover of key cohesin subunits, such as the meiotic cohesin Rec8 or SMC1β, have demonstrated that these cohesins are not replenished after birth in the growing follicles of the mouse88,89. The progressive deterioration of cohesion could potentially be a leading cause for the increase in errors in chromosome segregation (in particular, errors in meiosis I) observed with age1,3,90. Nonetheless, in Drosophila oocytes, evidence for cohesin rejuvenation during oogenesis has been shown91. Whether this aspect is specific to Drosophila or can be extended to mammals is currently unknown.
The effect of cohesin deterioration can be further amplified by the poor sensitivity of the SAC, at least in mice, to a reduction in tension on bivalents71. Recently, direct observations made on human oocytes have shown that sister kinetochores tend to split prematurely during meiosis I and that this effect increases with age92,93. Premature splitting potentially favors precocious bivalent dissociation into univalent, contributing to aneuploidy.
Conclusions
Amazing progress has been made since the pioneering review that put into the limelight the fact that human oocytes are error prone and that the rate of errors increases with the age of the mother1. This review challenged scientists in the field to try to understand a major societal issue of our modern societies where women postpone childbearing and to which female scientists were maybe already particularly exposed. Advance has come from studies on diverse model systems presenting both similarities and differences with human oocyte meiosis. Importantly, observations made in human oocytes have been challenged by hypotheses tested in model systems where genetics or biochemistry can be performed. Obviously, it is a combination of factors that seem to predispose oocytes to aneuploidy: their confounding fragile mode of spindle assembly and positioning, coupled to the pressure to preserve maternal stores in a gigantic cell via extreme asymmetry in size of their divisions as well as their longevity. Of course, many other factors not highlighted here, such as the distribution as well as the rate of recombination between homologues, which have a direct influence on chromosome segregation, can also predispose for aneuploidy in oocytes3. The major discovery of PRDM9, a key factor controlling the distribution of hot spots for recombination in some species but not others, will certainly help us to understand the impact of recombination on aneuploidy in oocytes94–96. Indeed, meiotic maturation is embedded in a continuous process that started in the embryo and that resumes later in the life of an animal. Meiotic maturation also prepares the gamete for fertilization and early development; hence, gamete quality does greatly influence the chances of a successful pregnancy.
Competing interests
The authors declare that they have no competing interests.
Grant information
The author(s) declared that no grants were involved in supporting this work.
Acknowledgments
We wish to thank Maria Almonacid (CIRB, Collège de France, Paris) for her help with the iconography of figures and Agathe Chaigne (Laboratory for Molecular Biology, University College London, UK), Julien Dumont (Institut Jacques Monod, Paris), and Renata Basto (Institut Curie, Paris) as well as all the members of the Verlhac lab for constructive comments on this review. We would like to apologize to those whose work we have not cited here, owing to space constraints.
Faculty Opinions recommended
References
1.
Hassold T, Hunt P:
To err (meiotically) is human: the genesis of human aneuploidy.
Nat Rev Genet.
2001; 2(4): 280–91. PubMed Abstract
| Publisher Full Text
2.
Hassold T, Hall H, Hunt P:
The origin of human aneuploidy: where we have been, where we are going.
Hum Mol Genet.
2007; 16(R2): R203–8. PubMed Abstract
| Publisher Full Text
4.
Szollosi D, Calarco P, Donahue RP:
Absence of centrioles in the first and second meiotic spindles of mouse oocytes.
J Cell Sci.
1972; 11(2): 521–41. PubMed Abstract
5.
Tamura M, Nemoto S:
Reproductive maternal centrosomes are cast off into polar bodies during maturation division in starfish oocytes.
Exp Cell Res.
2001; 269(1): 130–9. PubMed Abstract
| Publisher Full Text
6.
Manandhar G, Schatten H, Sutovsky P:
Centrosome reduction during gametogenesis and its significance.
Biol Reprod.
2005; 72(1): 2–13. PubMed Abstract
| Publisher Full Text
7.
Shirato Y, Tamura M, Yoneda M, et al.:
Centrosome destined to decay in starfish oocytes.
Development.
2006; 133(2): 343–50. PubMed Abstract
| Publisher Full Text
8.
Januschke J, Gervais L, Gillet L, et al.:
The centrosome-nucleus complex and microtubule organization in the Drosophila oocyte.
Development.
2006; 133(1): 129–39. PubMed Abstract
| Publisher Full Text
13.
Kirschner M, Mitchison T:
Beyond self-assembly: from microtubules to morphogenesis.
Cell.
1986; 45(3): 329–42. PubMed Abstract
| Publisher Full Text
14.
Heald R, Khodjakov A:
Thirty years of search and capture: The complex simplicity of mitotic spindle assembly.
J Cell Biol.
2015; 211(6): 1103–11. PubMed Abstract
| Publisher Full Text
| Free Full Text
15.
Gönczy P:
Mechanisms of spindle positioning: focus on flies and worms.
Trends Cell Biol.
2002; 12(7): 332–9. PubMed Abstract
| Publisher Full Text
16.
Kotak S, Gönczy P:
Mechanisms of spindle positioning: cortical force generators in the limelight.
Curr Opin Cell Biol.
2013; 25(6): 741–8. PubMed Abstract
| Publisher Full Text
17.
Rattner JB, Berns MW:
Centriole behavior in early mitosis of rat kangaroo cells (PTK2).
Chromosoma.
1976; 54(4): 387–95. PubMed Abstract
| Publisher Full Text
18.
Aubin JE, Osborn M, Weber K:
Variations in the distribution and migration of centriole duplexes in mitotic PtK2 cells studied by immunofluorescence microscopy.
J Cell Sci.
1980; 43: 177–94. PubMed Abstract
19.
Waters JC, Cole RW, Rieder CL:
The force-producing mechanism for centrosome separation during spindle formation in vertebrates is intrinsic to each aster.
J Cell Biol.
1993; 122(2): 361–72. PubMed Abstract
| Free Full Text
23.
Schuh M, Ellenberg J:
Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes.
Cell.
2007; 130(3): 484–98. PubMed Abstract
| Publisher Full Text
26.
Brunet S, Maria AS, Guillaud P, et al.:
Kinetochore fibers are not involved in the formation of the first meiotic spindle in mouse oocytes, but control the exit from the first meiotic M phase.
J Cell Biol.
1999; 146(1): 1–12. PubMed Abstract
| Publisher Full Text
| Free Full Text
30.
Heald R, Tournebize R, Blank T, et al.:
Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts.
Nature.
1996; 382(6590): 420–5. PubMed Abstract
| Publisher Full Text
31.
Breuer M, Kolano A, Kwon M, et al.:
HURP permits MTOC sorting for robust meiotic spindle bipolarity, similar to extra centrosome clustering in cancer cells.
J Cell Biol.
2010; 191(7): 1251–60. PubMed Abstract
| Publisher Full Text
| Free Full Text
33.
Ohkura H:
Meiosis: an overview of key differences from mitosis.
Cold Spring Harb Perspect Biol.
2015; 7(5): pii: a015859. PubMed Abstract
| Publisher Full Text
34.
Maro B, Howlett SK, Webb M:
Non-spindle microtubule organizing centers in metaphase II-arrested mouse oocytes.
J Cell Biol.
1985; 101(5 Pt 1): 1665–72. PubMed Abstract
| Publisher Full Text
| Free Full Text
35.
Gard DL:
Microtubule organization during maturation of Xenopus oocytes: assembly and rotation of the meiotic spindles.
Dev Biol.
1992; 151(2): 516–30. PubMed Abstract
| Publisher Full Text
36.
Gard DL:
Gamma-tubulin is asymmetrically distributed in the cortex of Xenopus oocytes.
Dev Biol.
1994; 161(1): 131–40. PubMed Abstract
| Publisher Full Text
37.
Srayko M, Buster DW, Bazirgan OA, et al.:
MEI-1/MEI-2 katanin-like microtubule severing activity is required for Caenorhabditis elegans meiosis.
Genes Dev.
2000; 14(9): 1072–84. PubMed Abstract
| Free Full Text
| Faculty Opinions Recommendation
38.
Cullen CF, Ohkura H:
Msps protein is localized to acentrosomal poles to ensure bipolarity of Drosophila meiotic spindles.
Nat Cell Biol.
2001; 3(7): 637–42. PubMed Abstract
| Publisher Full Text
40.
Cimini D, Howell B, Maddox P, et al.:
Merotelic kinetochore orientation is a major mechanism of aneuploidy in mitotic mammalian tissue cells.
J Cell Biol.
2001; 153(3): 517–27. PubMed Abstract
| Publisher Full Text
| Free Full Text
42.
Silkworth WT, Nardi IK, Scholl LM, et al.:
Multipolar spindle pole coalescence is a major source of kinetochore mis-attachment and chromosome mis-segregation in cancer cells.
PLoS One.
2009; 4(8): e6564. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Longo FJ, Chen DY:
Development of cortical polarity in mouse eggs: involvement of the meiotic apparatus.
Dev Biol.
1985; 107(2): 382–94. PubMed Abstract
| Publisher Full Text
44.
Verlhac MH, Lefebvre C, Kubiak JZ, et al.:
Mos activates MAP kinase in mouse oocytes through two opposite pathways.
EMBO J.
2000; 19(22): 6065–74. PubMed Abstract
| Publisher Full Text
| Free Full Text
46.
Prodon F, Chenevert J, Sardet C:
Establishment of animal-vegetal polarity during maturation in ascidian oocytes.
Dev Biol.
2006; 290(2): 297–311. PubMed Abstract
| Publisher Full Text
47.
Dumont J, Million K, Sunderland K, et al.:
Formin-2 is required for spindle migration and for the late steps of cytokinesis in mouse oocytes.
Dev Biol.
2007; 301(1): 254–65. PubMed Abstract
| Publisher Full Text
48.
Azoury J, Lee KW, Georget V, et al.:
Spindle positioning in mouse oocytes relies on a dynamic meshwork of actin filaments.
Curr Biol.
2008; 18(19): 1514–9. PubMed Abstract
| Publisher Full Text
49.
Schuh M, Ellenberg J:
A new model for asymmetric spindle positioning in mouse oocytes.
Curr Biol.
2008; 18(24): 1986–92. PubMed Abstract
| Publisher Full Text
50.
Amiel A, Leclère L, Robert L, et al.:
Conserved functions for Mos in eumetazoan oocyte maturation revealed by studies in a cnidarian.
Curr Biol.
2009; 19(4): 305–11. PubMed Abstract
| Publisher Full Text
57.
Kwon M, Bagonis M, Danuser G, et al.:
Direct Microtubule-Binding by Myosin-10 Orients Centrosomes toward Retraction Fibers and Subcortical Actin Clouds.
Dev Cell.
2015; 34(3): 323–37. PubMed Abstract
| Publisher Full Text
| Free Full Text
58.
Agircan FG, Schiebel E, Mardin BR:
Separate to operate: control of centrosome positioning and separation.
Philos Trans R Soc Lond B Biol Sci.
2014; 369(1650): pii: 20130461. PubMed Abstract
| Publisher Full Text
| Free Full Text
63.
Carreno S, Kouranti I, Glusman ES, et al.:
Moesin and its activating kinase Slik are required for cortical stability and microtubule organization in mitotic cells.
J Cell Biol.
2008; 180(4): 739–46. PubMed Abstract
| Publisher Full Text
| Free Full Text
64.
Roubinet C, Decelle B, Chicanne G, et al.:
Molecular networks linked by Moesin drive remodeling of the cell cortex during mitosis.
J Cell Biol.
2011; 195(1): 99–112. PubMed Abstract
| Publisher Full Text
| Free Full Text
66.
Chaigne A, Campillo C, Gov NS, et al.:
A soft cortex is essential for asymmetric spindle positioning in mouse oocytes.
Nat Cell Biol.
2013; 15(8): 958–66. PubMed Abstract
| Publisher Full Text
67.
Chaigne A, Campillo C, Gov NS, et al.:
A narrow window of cortical tension guides asymmetric spindle positioning in the mouse oocyte.
Nat Commun.
2015; 6: 6027. PubMed Abstract
| Publisher Full Text
70.
Lane SI, Yun Y, Jones KT:
Timing of anaphase-promoting complex activation in mouse oocytes is predicted by microtubule-kinetochore attachment but not by bivalent alignment or tension.
Development.
2012; 139(11): 1947–55. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
73.
Sebestova J, Danylevska A, Novakova L, et al.:
Lack of response to unaligned chromosomes in mammalian female gametes.
Cell Cycle.
2012; 11(16): 3011–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
74.
Clark-Maguire S, Mains PE:
Localization of the mei-1 gene product of Caenorhaditis elegans, a meiotic-specific spindle component.
J Cell Biol.
1994; 126(1): 199–209. PubMed Abstract
| Publisher Full Text
| Free Full Text
75.
Matthews LR, Carter P, Thierry-Mieg D, et al.:
ZYG-9, a Caenorhabditis elegans protein required for microtubule organization and function, is a component of meiotic and mitotic spindle poles.
J Cell Biol.
1998; 141(5): 1159–68. PubMed Abstract
| Publisher Full Text
| Free Full Text
76.
Peter M, Castro A, Lorca T, et al.:
The APC is dispensable for first meiotic anaphase in Xenopus oocytes.
Nat Cell Biol.
2001; 3(1): 83–7. PubMed Abstract
| Publisher Full Text
78.
Batiha O, Swan A:
Evidence that the spindle assembly checkpoint does not regulate APCFzy activity in Drosophila female meiosis.
Genome.
2012; 55(1): 63–7. PubMed Abstract
| Publisher Full Text
80.
Newport JW, Kirschner MW:
Regulation of the cell cycle during early Xenopus development.
Cell.
1984; 37(3): 731–42. PubMed Abstract
| Publisher Full Text
81.
Battaglia DE, Goodwin P, Klein NA, et al.:
Influence of maternal age on meiotic spindle assembly in oocytes from naturally cycling women.
Hum Reprod.
1996; 11(10): 2217–22. PubMed Abstract
82.
Hamatani T, Falco G, Carter MG, et al.:
Age-associated alteration of gene expression patterns in mouse oocytes.
Hum Mol Genet.
2004; 13(19): 2263–78. PubMed Abstract
| Publisher Full Text
92.
Patel J, Tan SL, Hartshorne GM, et al.:
Unique geometry of sister kinetochores in human oocytes during meiosis I may explain maternal age-associated increases in chromosomal abnormalities.
Biol Open.
2015; 5(2): 178–84. PubMed Abstract
| Publisher Full Text
Verlhac MH and Terret ME. Oocyte Maturation and Development [version 1; peer review: 2 approved]. F1000Research 2016, 5(F1000 Faculty Rev):309 (https://doi.org/10.12688/f1000research.7892.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)