1Section of Neurobiology, Division of Biological Sciences, University of California, San Diego, La Jolla, CA, 92093, USA
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
The capacity of an axon to regenerate is regulated by its external environment and by cell-intrinsic factors. Studies in a variety of organisms suggest that alterations in axonal microtubule (MT) dynamics have potent effects on axon regeneration. We review recent findings on the regulation of MT dynamics during axon regeneration, focusing on the nematode Caenorhabditis elegans. In C. elegans the dual leucine zipper kinase (DLK) promotes axon regeneration, whereas the exchange factor for Arf6 (EFA-6) inhibits axon regeneration. Both DLK and EFA-6 respond to injury and control axon regeneration in part via MT dynamics. How the DLK and EFA-6 pathways are related is a topic of active investigation, as is the mechanism by which EFA-6 responds to axonal injury. We evaluate potential candidates, such as the MT affinity-regulating kinase PAR-1/MARK, in regulation of EFA-6 and axonal MT dynamics in regeneration.
Corresponding author:
Andrew D. Chisholm
Competing interests:
The authors declare that they have no competing interests.
Grant information:
Work in the Andrew D. Chisholm and Yishi Jin lab on axon regeneration is supported by NIH R01 NS093588 (to Andrew D. Chisholm and Yishi Jin).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
More than a hundred years ago, Ramon y Cajal was the first to describe how individual axons respond to injury1. Many types of axons regenerate, including neurons in the peripheral nervous system (PNS), re-forming growth cones similar to those that Ramon y Cajal had characterized during development. In contrast, neurons of the mammalian central nervous system (CNS) often fail to regenerate, and their damaged ends form swollen, non-motile structures later termed retraction bulbs. These fundamental observations set the stage for subsequent exploration of why regenerative capacity varies drastically between the CNS and PNS. Many studies have focused on the inhibitory environment of the adult mammalian CNS2–4. However, it is becoming evident that cell-intrinsic processes are also key determinants of axon regeneration5. Among these intrinsic factors, regulation of axonal microtubule (MT) dynamics has emerged as a major influence on the capacity of an axon to regrow effectively6–10. Most strikingly, pharmacological stabilization of MTs by paclitaxel or related molecules enhances axon regeneration in vitro and in vivo, suggesting a potentially therapeutically significant role for MT dynamics in axon regeneration11–13.
The nematode C. elegans has long been used for studies in neuronal development and behavior, owing to its short life cycle, genetic tractability, and ease of in vivo imaging. The nervous system of an adult C. elegans hermaphrodite consists of 302 neurons of nearly invariant lineage14. A decade ago, a pioneering study showed that axons of mature C. elegans neurons can regenerate after precise laser axotomy15. Several kinds of neuron display robust axon regeneration; most work has focused on the mechanosensory and motor axons (Figure 1A)16,17. The genetic tractability and ease of imaging in vivo have made C. elegans a rising star in axon regeneration studies. Several laboratories have used large-scale genetic18, chemical19, and RNA interference (RNAi)20 screens to identify genes or molecules that regulate axon regeneration. These studies have identified several new players in axon regeneration, including the dual leucine zipper kinase (DLK) and mixed lineage kinase (MLK) mitogen-activated protein kinase (MAPK) pathways21,22, Notch signaling pathway23, insulin signaling pathway24, and microRNA25. Some pathways, such as DLK signaling, have been shown to function in axon regeneration in vertebrates26–29, suggesting that axon regeneration factors identified in C. elegans may be suitable for translational studies. Here, we review the role of MT dynamics in axon regeneration, primarily focusing on C. elegans.
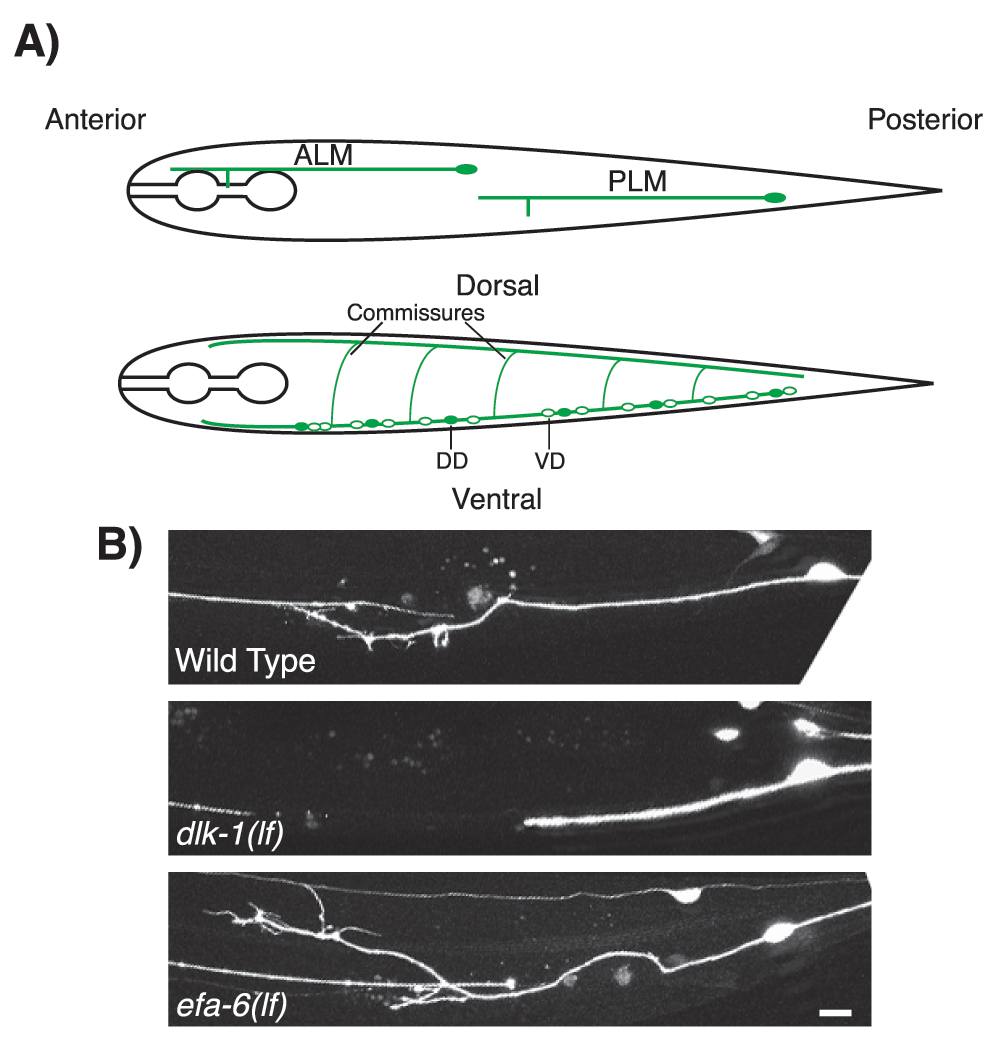
Figure 1. DLK-1 and EFA-6 regulate axon regeneration after injury in Caenorhabditis elegans.
(A) Illustration of the positions of the mechanosensory neurons anterior lateral microtubule (ALM) and posterior lateral microtubule (PLM) cells (top) and the GABAergic motor neurons dorsal D (DD) and ventral D (VD) (bottom). (B) Examples of PLM axon regeneration in wild-type, dlk-1(lf), and efa-6(lf) at 24 hours post-axotomy. dlk-1(lf) mutant (middle) shows decreased axon regeneration, whereas efa-6(lf) mutant (bottom) shows increased axon regeneration upon axotomy, compared with wild-type (top). Scale bar, 20 µm. DLK, dual leucine zipper kinase.
Axonal microtubule organization before and after injury
MTs are among the major cytoskeletal structures in cells. MTs are cylindrical and polarized polymers formed by αβ-tubulin heterodimers arranged in a head-to-tail configuration30. In vitro and in vivo MTs undergo rapid growth (i.e. polymerization) and shrinkage (i.e. depolymerization) at their plus ends, a behavior known as dynamic instability31. Minus ends of MTs are relatively stable but can also undergo polymerization and depolymerization. MT dynamics in vivo are influenced by many factors, including concentration of free tubulin monomers and tubulin post-translational modifications, and by MT-binding proteins. MT plus-end dynamics are regulated by a large cohort of plus end-tracking proteins (+TIPs)32; relatively few minus end-targeting proteins (−TIPs) have been identified that regulate MT minus ends33. Together, these proteins affect the frequency of catastrophe (switching from growth to shrinkage) and rescue (switching from shrinkage to growth) events.
In contrast to the highly dynamic behavior of MTs in dividing or migrating cells, axonal MTs of mature neurons are relatively stable, forming a consistent architecture that maintains neuronal polarity and allows directed axonal transport34. Axonal MTs in C. elegans were first characterized in the MT-rich mechanosensory neurons35,36. More recent imaging of the dynamics of plus end-binding proteins indicates that, as in other organisms, C. elegans axonal MTs are consistently arranged with plus ends away from the soma (‘plus end out’) but that dendritic MTs either are oriented with minus ends out or have mixed orientation37,38.
After axon injury, the stable axonal MTs become highly dynamic to allow axonal regrowth and establishment of a new growth cone6,7. In cultured Aplysia californica neurons, injury triggers rapid MT depolymerization followed by repolymerization with aberrant MT orientation39,40. Reversal of MT polarity after injury has been observed in Drosophila dendrites41,42. In addition, axotomy triggers an acute change of MT dynamics in Drosophila41,43,44. In C. elegans, axotomy of the mechanosensory posterior lateral microtubule (PLM) neuron triggers an increase in growing MTs locally at the injury site, followed by persistent growth of MTs that leads to formation of functional growth cones37. A mutation in mec-7/β-tubulin that hyperstabilizes MTs in touch neurons inhibits anterior lateral microtubule (ALM) axon regeneration, suggesting that precise regulation of MT dynamics is essential for axon regeneration45. Regenerating axon tips in severed mouse neurons display an acute increase in MT dynamics, followed by a sustained increase over several days46. Collectively, these findings suggest that axonal injury initiates an intricate series of changes in axonal MT organization.
The DLK-1 MAPK cascade promotes axon regeneration, in part via microtubule dynamics
The DLK MAPK pathway was identified several years ago as essential for axon regeneration in C. elegans motor neurons and in mechanosensory neurons21,22. Mutants lacking DLK-1 [dlk-1(lf)] display normal developmental axon growth but are unable to regenerate after injury, being blocked at the initial phase of growth cone reformation (Figure 1B). Conversely, overexpression of dlk-1 [dlk-1(gf)] enhances axon regeneration21,22. A mammalian DLK-1 homolog MAP3K13/LZK can functionally substitute for dlk-147, suggesting a high degree of conservation of the DLK pathway in axon regeneration. Indeed, in mammals, DLK is also required for axon regeneration after axonal injury26–29.
DLK-1 activity is required cell-autonomously at the time of regrowth, and DLK-1 itself is likely activated by injury signals. An axotomy-triggered Ca2+ transient has been implicated in DLK-1 activation47–49. In addition, the DLK pathway is sensitive to MT depolymerization. Mutations disrupting MTs trigger a DLK-dependent reduction of protein levels in touch neurons50. In Drosophila, loss of Short stop (shot), a member of the spectraplakin family that crosslinks actin and MT51,52, activates the DLK signaling pathway to promote axon regeneration53. Moreover, disruption of MTs by nocodazole in mammalian sensory neurons activates the DLK signaling pathway54. As yet, it remains unclear how MT polymerization is sensed by DLK.
Activation of the DLK pathway leads to two major outputs in C. elegans: a transcriptional response involving the CEBP-1 bZip transcription factor and CEBP-1-independent effects on axonal MT dynamics. The dlk-1(lf) mutant fails to increase persistent MT growth after axotomy, whereas dlk-1(gf) shows increased number of growing axonal MTs, both before and after axotomy37. Following laser injury, the DLK pathway promotes MT dynamics and growth, through downregulation of the kinesin-13 KLP-7 and upregulation of the cytosolic carboxypeptidase CCPP-637. Thus, the DLK cascade is closely interconnected with MTs, both as a sensor of MT integrity and as a regulator of MT dynamics, making it well placed to mediate regenerative reorganization of the axonal MT cytoskeleton after injury.
EFA-6, an inhibitor of axon regeneration acting via microtubule dynamics
The above studies of DLK-1 have helped spur efforts to identify additional factors that control MT dynamics during axon regeneration. Using a large-scale genetic screen, we identified the evolutionarily conserved protein EFA-6 (exchange factor for Arf-6) as a cell-intrinsic suppressor of axon regeneration18. Loss-of-function mutations of efa-6 [efa-6(lf)] enhance axonal regeneration, whereas efa-6 overexpression [efa-6(gf)] blocks regeneration (Figure 1B). Uniquely, efa-6(lf) partially bypasses the requirement for DLK-1 in axonal regeneration18, suggesting that EFA-6 and DLK-1 have antagonistic effects on a common process. Multiple lines of evidence suggest that EFA-6 inhibits axon regeneration through modulation of MT dynamics18,55.
The EFA-6/EFA6 protein family is conserved from yeast to mammals. EFA-6 contains a Sec7 domain that confers guanine exchange activity (GEF) for Arf6 GTPases56. Four EFA6 members (EFA6A–EFA6D) have been identified in mammals and three of them (except EFA6B) are expressed in neurons57,58. EFA6 localizes to the plasma membrane through its pleckstrin homology (PH) domain56. Furthermore, EFA6 can interact with filamentous actin in vitro through its PH domain and plays important roles in regulation of cortical actin cytoskeleton in vertebrate cells56,57,59–61. In C. elegans, efa-6 suppresses the embryonic lethality caused by mutations in dynein, a MT motor62, suggesting a functional linkage between actin and MT cytoskeletons at the cell cortex. In the one-cell C. elegans embryo, EFA-6 localizes to the plasma membrane, with enrichment in the anterior cortex in late one-cell embryo, to limit MT growth throughout the cell cortex62,63. The plasma membrane localization of EFA-6 is dependent upon the presence of its PH domain, whereas the intrinsically disordered N-terminal domain confers the enrichment at the anterior cell cortex63. The N-terminal region of EFA-6 contains a conserved 18-amino acid (18-aa) motif (Figure 2), which is essential for the MT growth-inhibiting activity.
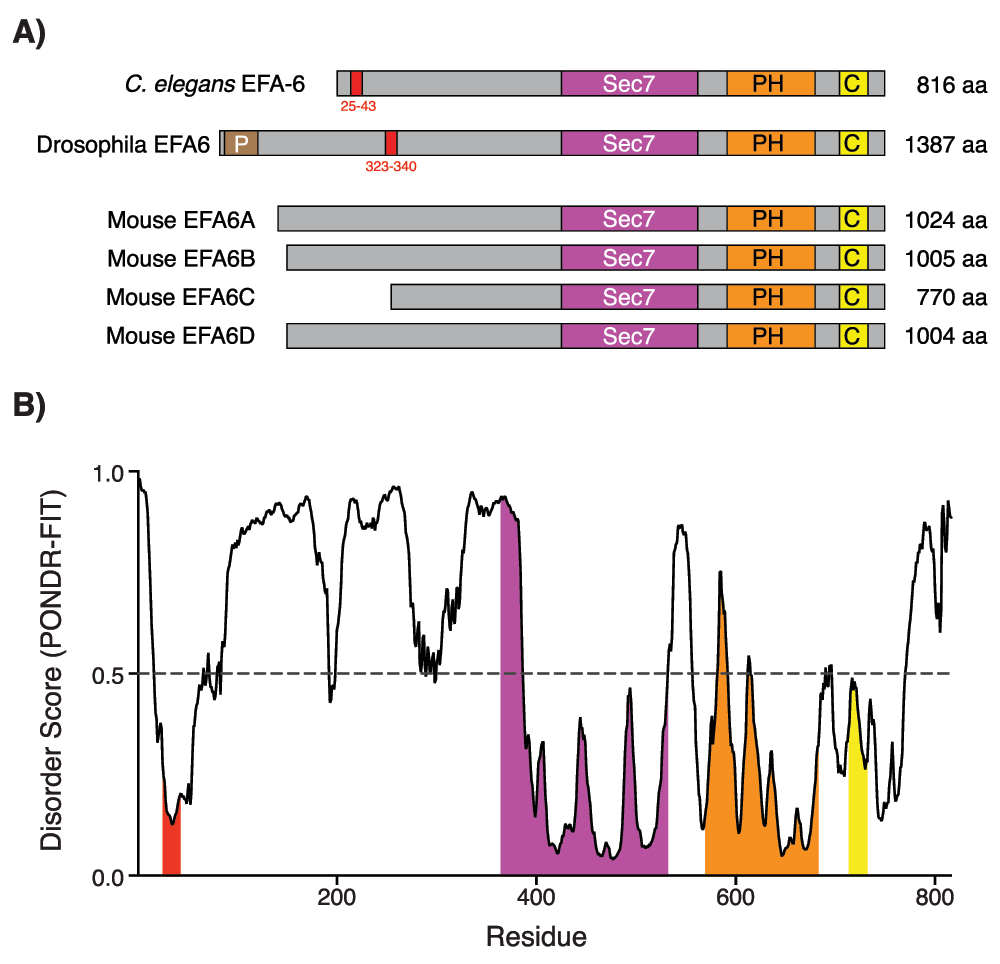
Figure 2. The intrinsically disordered N-terminal of EFA-6 contains a conserved 18-amino acid (18-aa) motif.
(A) EFA-6 protein domain organization in different organisms. Red boxes in the EFA-6 N-terminus highlight a conserved 18-aa motif, found in both Caenorhabditis elegans and Drosophila63. (B) Plot of intrinsic protein disorder score for C. elegans EFA-6. Different domains of EFA-6 are color-coded as in (A). Note that EFA-6 N-terminus has an overall high disorder probability, apart from the 18-aa motif. Figure adapted from 55.
In mature uninjured neurons, EFA-6 also localizes to the cell cortex via its C-terminal PH domain55. Upon axotomy, EFA-6 rapidly (within minutes) relocalizes to puncta close to sites containing MT minus ends, as marked by the minus end-binding protein PTRN-1/Patronin55,64. Relocalization of EFA-6 is dependent on the intrinsically disordered N-terminal domain and plays important roles in inhibition of axon regeneration (Figure 3A). In addition, the N-terminal domain of EFA-6 binds to the MT-associated proteins TAC-1/TACC (transforming acidic coiled-coil) and ZYG-8/DCLK (double-cortin-like kinase). Both TAC-1 and ZYG-8 are required for regenerative growth cone formation after axonal injury55. Although the roles of mammalian EFA6 family members in axon regeneration remain to be examined, Xenopus TACC3 promotes axon outgrowth in embryonic cultured neural crest cells65, and DCLK is required in mammalian axon regrowth66, suggesting potential functional conservation from C. elegans to mammals.
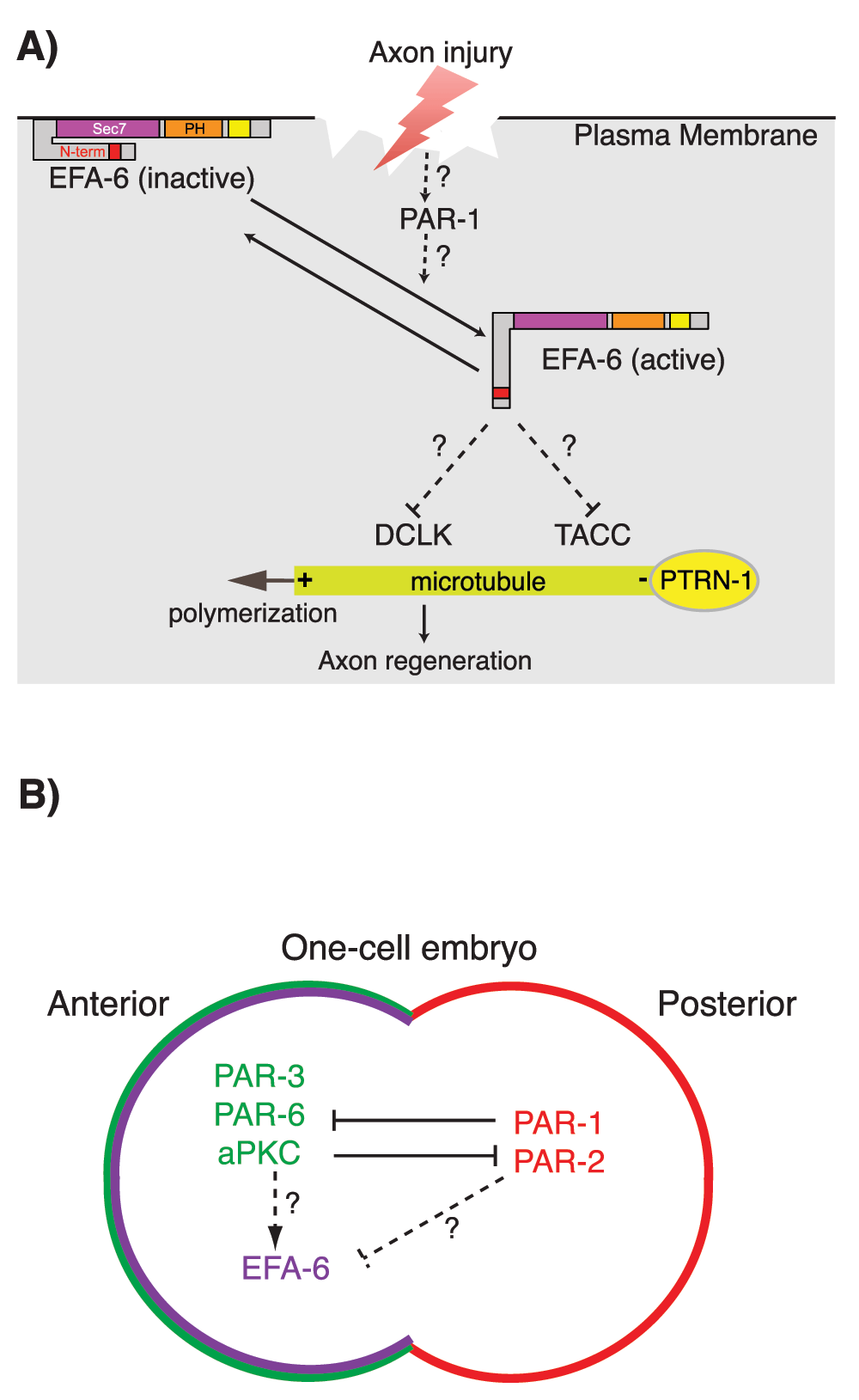
Figure 3. Functions of EFA-6 and PAR-1 in axon regeneration and polarity control in the early Caenorhabditis elegans embryo.
(A) Model for the regulation of EFA-6. In steady-state axons, EFA-6 localizes to the plasma membrane through its pleckstrin homology (PH) domain. Upon axon injury, EFA-6 and TAC-1/TACC (transforming acidic coiled-coil) relocalize close to the microtubule (MT) minus ends as defined by PTRN-1/Patronin puncta. The N-terminal intrinsically disordered region of EFA-6 is necessary and sufficient for relocalization and binding to TAC-1. (B) In one-cell embryos, PAR-1 and PAR-2 localize to the posterior cortex. This localization restricts the PAR-3/PAR-6/aPKC polarity complex to the anterior cortex. EFA-6 is enriched at the anterior cortex, dependent on the intrinsically disordered N-terminal domain. Such polarity complexes could regulate EFA-6 localization.
EFA-6, cell polarity proteins, and microtubule dynamics
Current evidence suggests that EFA-6 has bifunctional roles, as an MT destabilizing factor at the cell cortex in the steady state and relocalizing to the vicinity of MT minus ends after injury55. The molecular mechanisms involved in the transitions between these putative EFA-6 activity/localization states remain unknown. However, the intrinsically disordered N-terminal domain appears to be key to understanding the functions of EFA-6 with respect to MT dynamics. All EFA6 family members contain large N-terminal domains that are predicted to be intrinsically disordered (Figure 2A). Within the N-terminal domain, the sole region of primary sequence conservation is the 18-aa motif conserved in C. elegans and other invertebrate EFA6 family members and partly recognizable in some vertebrate EFA6 members. This motif contains potential phosphorylation sites, mutation of which abolishes the relocalization and regeneration-inhibiting activities of EFA-655. However, the identity of upstream kinases or phosphatases remains unknown; DLK-1 does not appear to be required for injury-triggered relocalization of EFA-6.
As the phosphorylation status of EFA-6 is tightly correlated to its MT dynamics-regulating activity55, the kinase or kinases responsible for EFA-6 phosphorylation may also have MT dynamics-regulating activity, directly or indirectly. Furthermore, like EFA-6, such kinases may also regulate MT dynamics in early embryo. Many candidates, including PAR-1/MARK (MT affinity-regulating kinase) and polo-like kinase, have been implicated in MT dynamics in embryos and neurons67,68, although none of these kinases has yet been associated with EFA-6 phosphorylation. Here, we focus on PAR-1/MARK and summarize the known functions of PAR-1/MARK in neurons and embryos that may be relevant to axon regeneration.
PAR-1/MARK is one of the PAR (partitioning defective) proteins, first identified in C. elegans for their roles in polarization of the early embryo69. PAR-1 encodes a serine/threonine kinase related to the MARKs70. Misregulation of PAR-1 and its phosphorylation targets has long been implicated in neuronal diseases such as Alzheimer’s disease and autism71,72. In mammalian neurons, MARK phosphorylates the MT-associated proteins (MAPs) tau, MAP2, and MAP4, causing these MAPs to dissociate from MTs and thereby destabilizing the MT network and increasing MT dynamics70. Furthermore, expression of MARK in cultured neurons promotes neurite outgrowth73. Neurite outgrowth involves a pioneer population of dynamic MTs that invades growth cones, followed by MT stabilization in axon extension74. These studies suggest that PAR-1/MARK plays key roles in MT plasticity during neurite outgrowth; less is known of its roles in axon regeneration.
PAR-1/MARK can regulate MT dynamics in many cell types75–79. In C. elegans one-cell embryos, PAR-1, together with its partner PAR-2, accumulates at the posterior cortex80. This localization prevents the anterior polarity complex, PAR-3/PAR-6/aPKC, from concentrating at the posterior cortex81,82 (Figure 3B). Intriguingly, MTs are more dynamic at the posterior end of an embryo, dependent on the asymmetric distribution of PAR proteins, including PAR-175. It is possible that PAR-1 regulates localization of EFA-6, causing an enrichment of EFA-6 at the anterior cortex (Figure 3B). Interactions between PAR network proteins and EFA-6 in the early embryo have not yet been tested but could affect regulation of cortical MT dynamics.
Concluding remarks
Recent studies have highlighted the importance of MT dynamics in regulation of axon regeneration. Several players, including DLK-1 and EFA-6, have been identified to regulate MT dynamics upon axon injury. However, many questions remain unexamined (see “Outstanding issues”). Importantly, pharmacological stabilization of MTs enhances axon regeneration both in vitro and in vivo, highlighting the therapeutic potential of MT dynamics regulation in axon regeneration. We anticipate that future studies should elucidate these mechanisms, which are potentially relevant to therapeutic interventions aimed at promoting regenerative axon growth.
Outstanding issues
1. What triggers EFA-6 relocalization upon axon injury?
EFA-6 rapidly relocalizes close to the MT minus ends to inhibit MT dynamics upon axon injury. This relocalization may be controlled by phosphorylation of the intrinsically disordered N-terminus of EFA-6. However, the signals (kinases/phosphatases) that might trigger EFA-6 relocalization or function remain to be discovered. Identification of these signals will bring crucial insights into how the activity of EFA-6 is regulated, possibly allowing precise manipulation of EFA-6 activity in a regrowing axon.
2. What are the cellular targets of DLK and EFA-6?
Upon axotomy, DLK-1 promotes MT dynamics and growth, whereas EFA-6 relocalizes close to the MT minus ends to inhibit MT dynamics. Both proteins seem to affect axon regeneration by regulating MT dynamics. Intriguingly, efa-6(lf) can partially bypass the requirement of DLK-1 in axon regeneration. However, how EFA-6 interacts with the DLK-1 pathway remains unclear. It is of great interest to investigate these issues further to provide a better understanding of how MT dynamics control axon regeneration.
3. Is EFA-6 a conserved inhibitor of axon regeneration?
Identification of EFA-6 as a cell-intrinsic inhibitor of axon regeneration in C. elegans raises the question of whether any of the four mammalian EFA6 family members are involved in mammalian axon regeneration or axonal MT dynamics or both. Like C. elegans EFA-6, mammalian EFA6 family members all contain large N-terminal domains that are predicted to be intrinsically disordered. EFA6A, EFA6C, and EFA6D are expressed in the nervous system, but their roles in axon regeneration have yet to be assessed. Given evidence that partial stabilization of MT dynamics can improve axon regeneration in vertebrates, manipulation of specific MT destabilizing factors such as EFA-6 might allow a more targeted approach to enhancing regrowth in a therapeutic context.
Competing interests
The authors declare that they have no competing interests.
Grant information
Work in the Andrew D. Chisholm and Yishi Jin lab on axon regeneration is supported by NIH R01 NS093588 (to Andrew D. Chisholm and Yishi Jin).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
We thank Fumio Motegi and members of the Andrew D. Chisholm and Yishi Jin labs for comments.
Faculty Opinions recommended
References
1.
Ramon y Cajal S, Defelipe J, Jones EG:
Cajal's degeneration and regeneration of the nervous system.
New York, Oxford University Press.
1991. Publisher Full Text
5.
Jin Y:
Unraveling the mechanisms of synapse formation and axon regeneration: the awesome power of C. elegans genetics.
Sci China Life Sci.
2015; 58(11): 1084–8. PubMed Abstract
| Publisher Full Text
| Free Full Text
6.
Chisholm AD:
Cytoskeletal dynamics in Caenorhabditis elegans axon regeneration.
Annu Rev Cell Dev Biol.
2013; 29: 271–97. PubMed Abstract
| Publisher Full Text
7.
Bradke F, Fawcett JW, Spira ME:
Assembly of a new growth cone after axotomy: the precursor to axon regeneration.
Nat Rev Neurosci.
2012; 13(3): 183–93. PubMed Abstract
| Publisher Full Text
9.
Liu K, Tedeschi A, Park KK, et al.:
Neuronal intrinsic mechanisms of axon regeneration.
Annu Rev Neurosci.
2011; 34: 131–52. PubMed Abstract
| Publisher Full Text
10.
Hur EM, Saijilafu, Zhou FQ:
Growing the growth cone: remodeling the cytoskeleton to promote axon regeneration.
Trends Neurosci.
2012; 35(3): 164–74. PubMed Abstract
| Publisher Full Text
| Free Full Text
14.
White JG, Southgate E, Thomson JN, et al.:
The structure of the nervous system of the nematode Caenorhabditis elegans.
Philos Trans R Soc Lond B Biol Sci.
1986; 314(1165): 1–340. PubMed Abstract
| Publisher Full Text
15.
Yanik MF, Cinar H, Cinar HN, et al.:
Neurosurgery: functional regeneration after laser axotomy.
Nature.
2004; 432(7019): 822. PubMed Abstract
| Publisher Full Text
18.
Chen L, Wang Z, Ghosh-Roy A, et al.:
Axon regeneration pathways identified by systematic genetic screening in C. elegans.
Neuron.
2011; 71(6): 1043–57. PubMed Abstract
| Publisher Full Text
| Free Full Text
19.
Samara C, Rohde CB, Gilleland CL, et al.:
Large-scale in vivo femtosecond laser neurosurgery screen reveals small-molecule enhancer of regeneration.
Proc Natl Acad Sci U S A.
2010; 107(43): 18342–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
22.
Yan D, Wu Z, Chisholm AD, et al.:
The DLK-1 kinase promotes mRNA stability and local translation in C. elegans synapses and axon regeneration.
Cell.
2009; 138(5): 1005–18. PubMed Abstract
| Publisher Full Text
| Free Full Text
26.
Itoh A, Horiuchi M, Bannerman P, et al.:
Impaired regenerative response of primary sensory neurons in ZPK/DLK gene-trap mice.
Biochem Biophys Res Commun.
2009; 383(2): 258–62. PubMed Abstract
| Publisher Full Text
27.
Shin JE, Cho Y, Beirowski B, et al.:
Dual leucine zipper kinase is required for retrograde injury signaling and axonal regeneration.
Neuron.
2012; 74(6): 1015–22. PubMed Abstract
| Publisher Full Text
| Free Full Text
32.
Akhmanova A, Steinmetz MO:
Tracking the ends: a dynamic protein network controls the fate of microtubule tips.
Nat Rev Mol Cell Biol.
2008; 9(4): 309–22. PubMed Abstract
| Publisher Full Text
36.
Chalfie M, Thomson JN:
Structural and functional diversity in the neuronal microtubules of Caenorhabditis elegans.
J Cell Biol.
1982; 93(1): 15–23. PubMed Abstract
| Publisher Full Text
| Free Full Text
40.
Erez H, Spira ME:
Local self-assembly mechanisms underlie the differential transformation of the proximal and distal cut axonal ends into functional and aberrant growth cones.
J Comp Neurol.
2008; 507(1): 1019–30. PubMed Abstract
| Publisher Full Text
41.
Stone MC, Nguyen MM, Tao J, et al.:
Global up-regulation of microtubule dynamics and polarity reversal during regeneration of an axon from a dendrite.
Mol Biol Cell.
2010; 21(5): 767–77. PubMed Abstract
| Publisher Full Text
| Free Full Text
43.
Chen L, Stone MC, Tao J, et al.:
Axon injury and stress trigger a microtubule-based neuroprotective pathway.
Proc Natl Acad Sci U S A.
2012; 109(29): 11842–7. PubMed Abstract
| Publisher Full Text
| Free Full Text
45.
Kirszenblat L, Neumann B, Coakley S, et al.:
A dominant mutation in mec-7/β-tubulin affects axon development and regeneration in Caenorhabditis elegans neurons.
Mol Biol Cell.
2013; 24(3): 285–96. PubMed Abstract
| Publisher Full Text
| Free Full Text
49.
Pinan-Lucarre B, Gabel CV, Reina CP, et al.:
The core apoptotic executioner proteins CED-3 and CED-4 promote initiation of neuronal regeneration in Caenorhabditis elegans.
PLoS Biol.
2012; 10(5): e1001331. PubMed Abstract
| Publisher Full Text
| Free Full Text
51.
Applewhite DA, Grode KD, Duncan MC, et al.:
The actin-microtubule cross-linking activity of Drosophila Short stop is regulated by intramolecular inhibition.
Mol Biol Cell.
2013; 24(18): 2885–93. PubMed Abstract
| Publisher Full Text
| Free Full Text
52.
Alves-Silva J, Sánchez-Soriano N, Beaven R, et al.:
Spectraplakins promote microtubule-mediated axonal growth by functioning as structural microtubule-associated proteins and EB1-dependent +TIPs (tip interacting proteins).
J Neurosci.
2012; 32(27): 9143–58. PubMed Abstract
| Publisher Full Text
| Free Full Text
53.
Valakh V, Walker LJ, Skeath JB, et al.:
Loss of the spectraplakin short stop activates the DLK injury response pathway in Drosophila.
J Neurosci.
2013; 33(45): 17863–73. PubMed Abstract
| Publisher Full Text
| Free Full Text
54.
Valakh V, Frey E, Babetto E, et al.:
Cytoskeletal disruption activates the DLK/JNK pathway, which promotes axonal regeneration and mimics a preconditioning injury.
Neurobiol Dis.
2015; 77: 13–25. PubMed Abstract
| Publisher Full Text
| Free Full Text
55.
Chen L, Chuang M, Koorman T, et al.:
Axon injury triggers EFA-6 mediated destabilization of axonal microtubules via TACC and doublecortin like kinase.
eLife.
2015; 4: e08695. PubMed Abstract
| Publisher Full Text
| Free Full Text
56.
Franco M, Peters PJ, Boretto J, et al.:
EFA6, a sec7 domain-containing exchange factor for ARF6, coordinates membrane recycling and actin cytoskeleton organization.
EMBO J.
1999; 18(6): 1480–91. PubMed Abstract
| Publisher Full Text
| Free Full Text
57.
Derrien V, Couillault C, Franco M, et al.:
A conserved C-terminal domain of EFA6-family ARF6-guanine nucleotide exchange factors induces lengthening of microvilli-like membrane protrusions.
J Cell Sci.
2002; 115(Pt 14): 2867–79. PubMed Abstract
58.
Sakagami H, Suzuki H, Kamata A, et al.:
Distinct spatiotemporal expression of EFA6D, a guanine nucleotide exchange factor for ARF6, among the EFA6 family in mouse brain.
Brain Res.
2006; 1093(1): 1–11. PubMed Abstract
| Publisher Full Text
59.
Luton F, Klein S, Chauvin JP, et al.:
EFA6, exchange factor for ARF6, regulates the actin cytoskeleton and associated tight junction in response to E-cadherin engagement.
Mol Biol Cell.
2004; 15(3): 1134–45. PubMed Abstract
| Publisher Full Text
| Free Full Text
60.
Klein S, Partisani M, Franco M, et al.:
EFA6 facilitates the assembly of the tight junction by coordinating an Arf6-dependent and -independent pathway.
J Biol Chem.
2008; 283(44): 30129–38. PubMed Abstract
| Publisher Full Text
| Free Full Text
61.
Macia E, Partisani M, Favard C, et al.:
The pleckstrin homology domain of the Arf6-specific exchange factor EFA6 localizes to the plasma membrane by interacting with phosphatidylinositol 4,5-bisphosphate and F-actin.
J Biol Chem.
2008; 283(28): 19836–44. PubMed Abstract
| Publisher Full Text
62.
O'Rourke SM, Dorfman MD, Carter JC, et al.:
Dynein modifiers in C. elegans: light chains suppress conditional heavy chain mutants.
PLoS Genet.
2007; 3(8): e128. PubMed Abstract
| Publisher Full Text
| Free Full Text
63.
O'Rourke SM, Christensen SN, Bowerman B:
Caenorhabditis elegans EFA-6 limits microtubule growth at the cell cortex.
Nat Cell Biol.
2010; 12(12): 1235–41. PubMed Abstract
| Publisher Full Text
| Free Full Text
64.
Chuang M, Goncharov A, Wang S, et al.:
The microtubule minus-end-binding protein patronin/PTRN-1 is required for axon regeneration in C. elegans.
Cell Rep.
2014; 9(3): 874–83. PubMed Abstract
| Publisher Full Text
| Free Full Text
65.
Nwagbara BU, Faris AE, Bearce EA, et al.:
TACC3 is a microtubule plus end-tracking protein that promotes axon elongation and also regulates microtubule plus end dynamics in multiple embryonic cell types.
Mol Biol Cell.
2014; 25(21): 3350–62. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
66.
Nawabi H, Belin S, Cartoni R, et al.:
Doublecortin-Like Kinases Promote Neuronal Survival and Induce Growth Cone Reformation via Distinct Mechanisms.
Neuron.
2015; 88(4): 704–19. PubMed Abstract
| Publisher Full Text
67.
Zitouni S, Nabais C, Jana SC, et al.:
Polo-like kinases: structural variations lead to multiple functions.
Nat Rev Mol Cell Biol.
2014; 15(7): 433–52. PubMed Abstract
| Publisher Full Text
69.
Kemphues KJ, Priess JR, Morton DG, et al.:
Identification of genes required for cytoplasmic localization in early C. elegans embryos.
Cell.
1988; 52(3): 311–20. PubMed Abstract
| Publisher Full Text
70.
Drewes G, Ebneth A, Preuss U, et al.:
MARK, a novel family of protein kinases that phosphorylate microtubule-associated proteins and trigger microtubule disruption.
Cell.
1997; 89(2): 297–308. PubMed Abstract
| Publisher Full Text
71.
Delacourte A, Buée L:
Tau pathology: a marker of neurodegenerative disorders.
Curr Opin Neurol.
2000; 13(4): 371–6. PubMed Abstract
72.
Drewes G, Trinczek B, Illenberger S, et al.:
Microtubule-associated protein/microtubule affinity-regulating kinase (p110mark). A novel protein kinase that regulates tau-microtubule interactions and dynamic instability by phosphorylation at the Alzheimer-specific site serine 262.
J Biol Chem.
1995; 270(13): 7679–88. PubMed Abstract
| Publisher Full Text
73.
Biernat J, Wu YZ, Timm T, et al.:
Protein kinase MARK/PAR-1 is required for neurite outgrowth and establishment of neuronal polarity.
Mol Biol Cell.
2002; 13(11): 4013–28. PubMed Abstract
| Publisher Full Text
| Free Full Text
74.
Conde C, Cáceres A:
Microtubule assembly, organization and dynamics in axons and dendrites.
Nat Rev Neurosci.
2009; 10(5): 319–32. PubMed Abstract
| Publisher Full Text
75.
Labbé JC, Maddox PS, Salmon ED, et al.:
PAR proteins regulate microtubule dynamics at the cell cortex in C. elegans.
Curr Biol.
2003; 13(9): 707–14. PubMed Abstract
| Publisher Full Text
76.
Doerflinger H, Benton R, Shulman JM, et al.:
The role of PAR-1 in regulating the polarised microtubule cytoskeleton in the Drosophila follicular epithelium.
Development.
2003; 130(17): 3965–75. PubMed Abstract
| Publisher Full Text
77.
Sapir T, Sapoznik S, Levy T, et al.:
Accurate balance of the polarity kinase MARK2/Par-1 is required for proper cortical neuronal migration.
J Neurosci.
2008; 28(22): 5710–20. PubMed Abstract
| Publisher Full Text
79.
Sapir T, Shmueli A, Levy T, et al.:
Antagonistic effects of doublecortin and MARK2/Par-1 in the developing cerebral cortex.
J Neurosci.
2008; 28(48): 13008–13. PubMed Abstract
| Publisher Full Text
80.
Guo S, Kemphues KJ:
par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed.
Cell.
1995; 81(4): 611–20. PubMed Abstract
| Publisher Full Text
81.
Munro E, Nance J, Priess JR:
Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo.
Dev Cell.
2004; 7: 413–24. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
Work in the Andrew D. Chisholm and Yishi Jin lab on axon regeneration is supported by NIH R01 NS093588 (to Andrew D. Chisholm and Yishi Jin).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Tang NH and Chisholm AD. Regulation of Microtubule Dynamics in Axon Regeneration: Insights from C. elegans [version 1; peer review: 3 approved]. F1000Research 2016, 5(F1000 Faculty Rev):764 (https://doi.org/10.12688/f1000research.8197.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
He Z. Reviewer Report For: Regulation of Microtubule Dynamics in Axon Regeneration: Insights from C. elegans [version 1; peer review: 3 approved]. F1000Research 2016, 5(F1000 Faculty Rev):764 (https://doi.org/10.5256/f1000research.8817.r13598)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
He Z. Reviewer Report For: Regulation of Microtubule Dynamics in Axon Regeneration: Insights from C. elegans [version 1; peer review: 3 approved]. F1000Research 2016, 5(F1000 Faculty Rev):764 (https://doi.org/10.5256/f1000research.8817.r13598)
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)