Keywords
neurodegenerative disease, brain rejuvenation, caloric restriction, exercise, blood plasma administration, healthspan,
neurodegenerative disease, brain rejuvenation, caloric restriction, exercise, blood plasma administration, healthspan,
Aging is the major risk factor for dementia-related neurodegenerative disease in the elderly. In the brain, aging is accompanied by a series of cellular and functional impairments that collectively drive vulnerability to neurodegenerative disease (Figure 1). It is important to take into account that, along with cellular and functional decline in the aged brain, organismal aging is broadly associated with global “hallmarks of aging” across all tissues in the body, such as stem cell dysfunction, genomic and protein instability, and altered intracellular communication, to name a few1. Consequently, age-related vulnerability to neurodegenerative disease occurs on a background of organismal-level functional decline, and as such interventions to counteract this vulnerability in the aged brain will benefit from a more systemic approach.
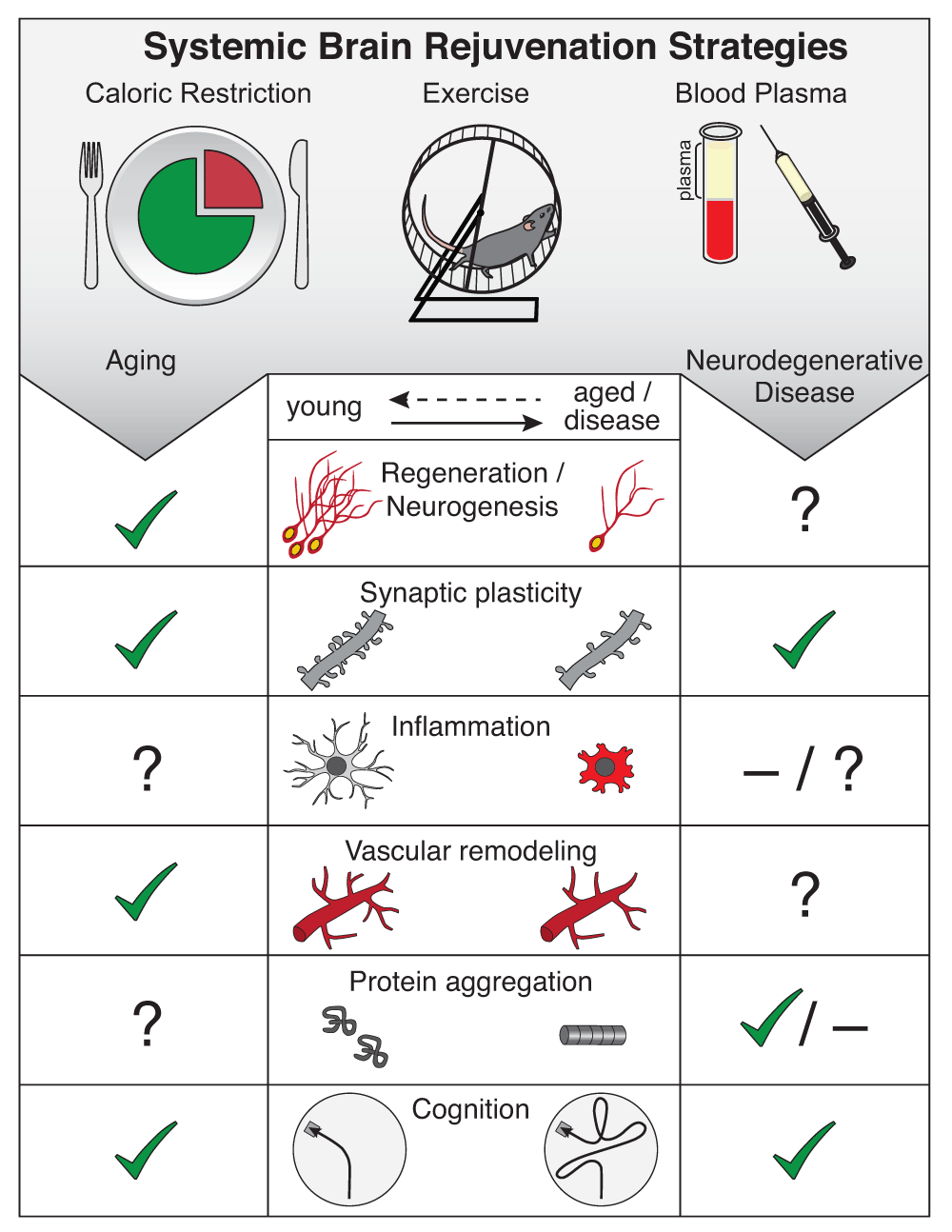
Hallmarks of brain aging amenable to rejuvenation (middle panel) include decreased regenerative capacity (neurogenesis), impaired synaptic plasticity, increased inflammation, vascular remodeling, increased protein aggregation, and impaired cognitive function. Systemic interventions (top panel), such as caloric restriction, exercise, and blood plasma administration, have been shown to rejuvenate hallmarks of brain aging (left panel) and ameliorate exacerbated pathology in models of neurodegenerative disease (right panel). Cellular or functional rejuvenation elicited by systemic interventions is denoted by a check (✔), lack of rejuvenation is denoted by a dash (–), and yet-to-be-determined effects are denoted by a question mark (?).
In the United States, approximately 15% of the population, 46 million people, are over the age of 65. The number of older adults is predicted to increase to 24% of the population, about 98 million people, by 20602. Unfortunately, this increase in lifespan has been accompanied by a drastic rise in age-associated disease. For example, Alzheimer's disease, one of multiple dementia-related neurodegenerative diseases primarily seen in adults over the age of 65, affected 5 million people in 2013 and is predicted to affect 14 million by 20503 in the United States. Beyond the personal cost of living with dementia-related neurodegenerative diseases, the monetary cost to society is in the billions. Thus, while longer lifespan indicates successful scientific and medical progress, it requires a corresponding increase in healthspan, the years lived free from disease, to prove truly transformative for human quality of life. The capacity to rejuvenate the aged brain is emerging as a tantalizing prospect for the treatment of dementia-related neurodegenerative diseases of aging4. Results from animal studies involving systemic interventions that promote healthspan—including a decrease in caloric intake without malnutrition also referred to as caloric restriction (CR), exercise, and exposure to a young systemic environment—demonstrate the rejuvenation of regenerative and functional capacity in aged tissues (Figure 1). It is now evident that such rejuvenation even extends to the aged brain at the regenerative, functional, and cognitive level. Therefore, the application of proven systemic interventions that extend healthspan may prove key in our ability to restore cellular and functional decline in the aging brain to prevent onset, or even counteract the progression, of dementia-related neurodegenerative disease in the elderly.
In this review, we will focus on blood-based brain rejuvenation strategies and their application to animal models of normal aging and neurodegenerative disease. We will discuss future therapeutic applications to brain rejuvenation and highlight ongoing clinical trials applying these findings to human disease.
Major hallmarks of brain aging include decreased regenerative capacity, altered vasculature, increased neuroinflammation, and impairments in synaptic plasticity that culminate in cognitive dysfunction (Figure 1)5–7. Neurodegenerative disease exacerbates these aging-associated cellular and functional hallmarks and introduces gross cellular loss, archetypal of neurodegeneration8. Here we provide a brief overview of key hallmarks of brain aging.
Regenerative capacity in the brain is mediated by the generation of new neurons from neural stem cells, a process known as neurogenesis, which precipitously declines with age9–11. Neurogenesis predominantly occurs in two neurogenic zones: the dentate gyrus of the hippocampus12, a region involved in spatial and episodic learning and memory, and the subventricular zone lining the lateral ventricles13. Owing to known associations between adult neurogenesis and cognitive function, age-related loss in regenerative capacity has been proposed to contribute to cognitive decline, although the direct link between the two processes in the context of aging has yet to be resolved14,15. Neurogenesis is tightly linked to nutritive sources provided by the cerebrovasculature through the blood–brain barrier, an anatomical separation between the central nervous system and the periphery consisting of the vascular cells and glia. With age, cerebral blood flow and blood–brain barrier integrity also decline, compromising metabolic support for neurogenesis and proper neuronal signaling and promoting inflammatory responses by resident immune cells16,17. Correspondingly, increased activation of astrocytes and microglia underlies the neuroinflammatory hallmark of brain aging, accompanied by increased pro-inflammatory cytokine production8. Such pro-inflammatory changes have now been shown to contribute to vulnerability and advancement of neurodegenerative disease through processes such as accumulation of complement factors (a component of innate immunity) in microglia8,18. At a functional level, synaptic plasticity also declines with age19. For example, electrophysiology studies reveal an age-related decline in long-term potentiation (LTP), an electrophysiological measure correlated with learning and memory. Functional impairments are also accompanied by corresponding molecular changes in the expression of plasticity-related factors such as immediate early genes. In the context of neurodegenerative disease, an exacerbated decline in synaptic plasticity occurs20. Consistent with synaptic dysfunction, cognitive processes such as learning and memory also decline with age. Neurodegenerative disease magnifies all hallmarks of brain aging while also promoting the accumulation of damaged or misfolded protein aggregates and gross neurodegeneration8, which collectively exacerbate cognitive impairment21.
Multiple lines of evidence in animal models point to the malleability of lifespan22–31. The initiation of rejuvenating interventions functioning at the systemic level, for example CR, in later stages of life was first demonstrated to improve the overall lifespan of an aged organism over 30 years ago32–34. More recently, systemically mediated rejuvenating interventions such as CR, exercise, and heterochronic parabiosis (in which the circulatory systems of a young and old animal are joined) have succeeded in rejuvenating aged tissues—including muscle, liver, heart, pancreas, bone, spinal cord, and brain—to improve organismal healthspan35–42.
The most robust and reliably replicated systemic intervention to improve lifespan and/or healthspan in rodents and non-human primates is CR43,44. In primates, CR has proved to be protective against the development of neoplasia, cardiovascular disease, and glucoregulatory impairment as well as gray matter loss in the brain45. In rodent models, CR leads to beneficial metabolic changes, protection from oxidative stress, neurotrophic factor production, increased autophagy, and neurogenesis46–49. CR also promotes the maintenance of cerebral blood flow and white matter integrity with age50,51. While CR has been demonstrated to improve spatial learning and memory in aged mice51, global effects on cognition remain unresolved, as CR failed to improve other hippocampal-dependent cognitive processes52–54. It is suggested that such inconsistent results could be due to the varied macronutrient ratios in CR regimes, the age of CR onset, and the metabolic heterogeneity of the diverse cell types in the hippocampus55–57. Nevertheless, CR reliably improves overall organismal healthspan and likewise improves behavioral and cognitive metrics in animal models of neurodegenerative disease58,59. While the potential benefits of CR for human healthspan are evident, limitations of feasibility, adequate health monitoring, and adherence remain. Indeed, in a long-term CR study conducted by the CALERIE Research Group, low adherence decreased the average caloric deficit from the anticipated 25% to 11.7% over a 2-year period, offsetting major expected beneficial metabolic outcomes60. Consequently, limits to the application of CR need to be further addressed before widespread therapeutic application.
Robust benefits of exercise have also been consistently observed to increase healthspan, although its role in extending lifespan in mice remains obfuscated61–63. Exercise has been shown to improve a wide range of age-related cellular and functional impairments throughout the body. For example, frailty, which refers to a global decline in function that encompasses grip strength, activity, overall energy, and unintentional weight loss, is amenable to the effects of exercise17,63,64. In rodent models, exercise reduces age-associated frailty through increasing skeletal muscle function63 and has been shown to improve grip strength and nesting and burrowing behaviors even when initiated at middle age17. In the normal aged brain, exercise has been shown to enhance regenerative capacity in aged mice by promoting hippocampal neurogenesis, effects that correlated with improved learning and memory65. Furthermore, exercise also improved broad cellular hallmarks of brain aging in aged mice, including increased synaptic plasticity, improved neurovascular integrity, and decreased microglia activation17,66,67. In the context of neurodegenerative disease, researchers have utilized mouse models of Alzheimer's disease to demonstrate that exercise is able to suppress inflammatory cytokines, increase the expression of antioxidant enzymes, improve synaptic function in the hippocampus, and enhance hippocampal-dependent learning and memory68. Researchers have further explored the relationship between physical exercise, healthspan, and cognitive function in human populations. Epidemiological studies demonstrate that physical activity is associated with better overall survival and function in older adults compared to their sedentary counterparts69,70. It has been proposed that such benefits in elderly humans may be the result of counteracting muscle weakness and frailty as well as reducing circulating inflammatory markers71,72, consistent with animal studies. Exercise is also reported to reduce risk factors for cardiovascular disease, such as hypertension and metabolic syndrome71, as well as reduce the risk of mild cognitive impairment later in life73–76. From a neurodegenerative disease perspective, a 6-month program of physical exercise in adults over the age of 50 years who are at risk for Alzheimer's disease also showed significantly improved cognition over an 18-month follow-up period77. While the potential for human application exists, limitations should be noted, with evidence in humans indicating that the perception of physical frailty or poor health alone can decrease adherence in the elderly78. Notwithstanding, these studies point to the capacity of exercise to extend healthspan and rejuvenate cellular and functional hallmarks of brain aging, with direct relevance to neurodegenerative diseases, such as Alzheimer’s disease.
Recently, the classical model of heterochronic parabiosis has re-emerged as an experimental platform to explore the intricate interplay between the systemic environment and organismal aging. To date, heterochronic parabiosis experiments have been used to explore the effects of aging throughout the body in tissues including muscle, pancreas, bone, heart, and brain35–38,40,42,79,80. Consistent with beneficial effects observed with exercise, the exposure of an aged mouse to a youthful systemic environment through heterochronic parabiosis likewise rejuvenates muscle function through decreased fibrogenic potential and improved regenerative capacity36–38. Moreover, heterochronic parabiosis also reversed cardiac hypertrophy and restored bone healing capacity in aged mice35,42. Excitingly, rejuvenating effects of heterochronic parabiosis are observed in the aged brain, indicating blood-borne mechanisms can counteract cellular and functional age-related neuronal decline39,79–81.
Heterochronic parabiosis studies have also pointed to a role for old blood in driving brain aging79. Regenerative capacity in young heterochronic parabionts is impaired, with neurogenesis decreasing in both the hippocampus and the subventricular zone neurogenic niches after exposure to old blood79,80. Interestingly, the detrimental effects of old blood observed in young heterochronic parabionts are specific to blood derived from old, but not middle-aged, animals80. These data begin to tease apart the kinetics of circulating factors in old blood, defining ages at which to target pro-aging factors as a therapeutic approach. An independent approach to investigate the effects of blood exchange was recently developed using a microfluidic-based blood exchange device in rodents82. With the use of this device, mice received a single heterochronic blood transfusion with whole blood derived from young or old mice, equivalent to 50% of total blood volume. Short-term exposure to old blood corroborated previous observations in the heterochronic parabiosis model that show an aged systemic environment negatively affects hippocampal adult neurogenesis. Further evidence of the pro-aging effects of old blood come from direct systemic interventions with old blood plasma injections. Short-term systemic administration of old plasma recapitulates the effects of heterochronic parabiosis on adult neurogenesis, implicating circulating factors in the effects of old blood82. Moreover, long-term administration of old plasma over one month also resulted in impaired hippocampal-dependent learning and memory79.
Thus far, only a few isolated circulating factors have been identified to mediate the effects of old blood. For example, the pleiotropic cytokine transforming growth factor beta (TGF-β) was identified to increase with age in both the circulation and the brain83. Inhibition of TGF-β signaling systemically and locally by RNA interference or by pharmacological inhibition with TGF-β receptor kinase Alk5 inhibitor enhanced neurogenesis in the aged hippocampus83. Using a proteomics-based approach in combination with heterochronic parabiosis, additional immune-related systemic pro-aging factors have been identified79. In particular, the C-C motif chemokine 11 (CCL11) and beta-2 microglobulin (B2M) have been shown to negatively regulate neurogenesis and cognitive function in the hippocampus79,81. Moreover, using genetic knockout studies, researchers have shown that the loss of B2M prevents, in part, the aging-associated decline in neurogenesis and cognitive function at old age81. Altogether, these results open up the possibility that functional rejuvenation in the aged brain may be possible by targeting specific circulating factors in old blood, potentially through interventions such as small-molecule-mediated pharmacological inhibition or the administration of neutralizing antibodies.
Evaluation of brain aging hallmarks in old heterochronic parabionts has further bolstered our appreciation for the inherent plasticity of the aged brain. Notably, regenerative capacity in the aged hippocampus and subventricular zone of heterochronic parabionts was enhanced after exposure to a young systemic environment39,80. This rejuvenating potential was maintained at a cell autonomous level with neural stem cells from old heterochronic parabionts showing enhanced self-renewal potential in vitro80. Vascularization of the neurogenic niche is known to influence neural stem cell function84,85, and correspondingly heterochronic parabiosis also restored blood vessel volume in the aged subventricular zone neurogenic niche to youthful levels80. Beyond regenerative function, exposure to a young systemic environment also enhanced synaptic plasticity, eliciting an increase in the expression of immediate early genes and the density of dendritic spines as well as enhancements in LTP39. At a cognitive level, long-term administration of young plasma over one month was sufficient to reverse cognitive impairments in hippocampal-dependent learning and memory in old mice through increased activation of the transcription factor cAMP response element binding protein (CREB)39. Consistently, old heterochronic parabionts that were separated from their young partner also demonstrated improvements in their olfactory discrimination ability compared to separated old isochronic controls. In a pre-clinical experiment, human umbilical cord-blood-derived plasma was demonstrated to similarly enhance immediate early gene expression, increase LTP, and improve cognitive function in aged mice86. This demonstration of the rejuvenating potential of human umbilical cord blood in mice strengthens the possibility that human blood can also be used to elicit brain rejuvenation in humans.
Thus far, growth differentiation factor 11 (GDF11), colony-stimulating factor 2 (CSF2), and tissue inhibitor of metalloproteinases 2 (TIMP2) have been identified as rejuvenating factors in young adult and/or juvenile blood35,86. While its role in cardiac and skeletal muscle rejuvenation is currently under discussion35,38,87, GDF11 was demonstrated to enhance neurogenesis and cerebral blood flow in aged mice when systemically administered80. Hinting at the multifactorial nature of young plasma, TIMP2 or CSF2 administration was also shown to enhance immediate early gene expression, LTP, and cognitive performance in aged mice86. Despite their shared rejuvenating effects, GDF11 was identified in young adult mouse plasma, while TIMP2 and CSF2 were identified in human umbilical cord blood, a comparatively early developmental stage. These distinctions again point to the importance of kinetics in determining circulating factor changes35,86. As more insight into the rejuvenating potential of young blood is obtained, it becomes evident that research is necessary to identify a breadth of factors by which to reverse global hallmarks of brain aging. While the burgeoning field of rejuvenation research is fast growing, and mounting evidence supports the translation of these interventions to humans, fundamental questions pertinent to therapeutic applications remain unexplored—in particular, how long lasting are the rejuvenating effects of young blood in the aged brain?
Current neurodegenerative disease animal models recapitulate many pathologies observed in humans, including amyloid plaque deposition, tau phosphorylation, increased neuroinflammation, altered synaptic plasticity, and cognitive impairments88. Despite their success in dissecting cellular and molecular changes involved in disease progression, current models are limited by the increasing array of transgenes they rely on, complicating interpretations and translation to human disease88. Additionally, such brain-centric models have also precluded the possibility of investigating the systemic contribution to neurodegenerative disease. This is especially limiting when one considers the number of systemic diseases, such as diabetes and atherosclerosis, for which neurodegeneration is a common co-morbidity89. Nevertheless, combining neurodegenerative disease animal models with systemic interventions, such as heterochronic parabiosis, has yielded promising results indicating the potential of young blood to counteract a number of neurodegenerative pathologies (Figure 1).
To date, young blood studies have focused on animal models of Alzheimer’s disease. Changes in Alzheimer’s disease-related pathology were first investigated in parabiosis studies in which young transgenic mice carrying the human amyloid precursor protein (APP) gene containing Swedish mutations and the human presenilin (PS1) gene encoding the deleted exon 9 mutation (APPswe/PS1dE9) were joined with age-matched young wild-type animals90. Exposure to a young wild-type systemic environment resulted in decreased plaque accumulation in APPswe/PS1dE9 transgenic mice after six months of parabiosis90. Additionally, pro-inflammatory cytokines, levels of tau phosphorylation, and gliosis were reduced in transgenic mice following exposure to a young wild-type circulatory system90. While this was not a direct effect of rejuvenation, as all parabionts were age-matched isochronic pairs, it does suggest a significant role for the systemic environment in the progression of neurodegenerative disease. Correspondingly, the use of peritoneal dialysis to filter amyloid-beta from the blood of APPswe/PS1dE9 transgenic mice also ameliorated Alzheimer’s-associated phenotypes, further demonstrating a role for the systemic environment in disease pathogenesis91. More recently, the effect of young blood in the context of aging was investigated using heterochronic parabiosis in a complementary transgenic mouse model of Alzheimer’s disease carrying the human APP gene harboring human familial London and Swedish mutations92. This study demonstrated increased expression of the synaptic marker synaptophysin and the pro-survival calcium binding protein calbindin in aged APP heterochronic parabionts exposed to a young systemic environment92. No changes in amyloid plaque deposition or microglia activation were observed92. Failure of heterochronic parabiosis to reverse plaque deposition and neuroinflammation after significant disease progression at old age suggests that it may be necessary to initiate systemic interventions prior to significant disease progression. Additionally, the duration of parabiosis used in the two studies above differed greatly from a more long-term six-month duration90 to a shorter five-week duration92, indicating that different disease pathologies may also prove amenable to improvements at vastly different timeframes. Lastly, at a cognitive level, injections of young wild-type plasma into aged APP transgenic mice also elicited improvements in hippocampal-dependent learning and memory92. Of note, the benefits of administering specific pro-youthful factors in neurodegenerative disease models have yet to be tested. However, treatment with neurotrophic compounds has previously been demonstrated to attenuate neurodegenerative disease pathology in a triple-transgenic Alzheimer’s disease mouse model harboring human APPswe, PS1, and tau mutations93,94. These emerging studies raise significant translational potential for blood-based therapeutic approaches to counter neurodegenerative disease progression at a functional and cognitive level.
Systemic interventions, including CR and exercise, have been previously evaluated in clinical trials, demonstrating beneficial effects on human healthspan and cognition (reviewed above). Notwithstanding, physical and technical barriers to adherence remain in the therapeutic application of exercise and CR. Coupled with promising results from young blood studies in animal models, particularly the recent demonstration of the rejuvenating effects of human umbilical cord plasma, researchers are now working to translate alternative blood-based systemic interventions to the clinic. Currently, a handful of ongoing clinical trials are seeking to identify biomarkers of healthspan and cognitive decline as well as exploring the potential of blood-based therapeutic interventions for the treatment of dementia-related neurodegenerative disease.
The Genetic and Epigenetic Signatures of Translational Aging Laboratory Testing (GESTALT) trial has taken a holistic approach to identifying biomarkers of healthspan95. The researchers aim to correlate biomarkers from blood, muscle, and skin with performance in a range of physical and cognitive exams for a 10-year period in healthy adults over the age of 20. They will assess peripheral blood mononuclear cells compared to muscle and skin biomarker data to assess systemic versus tissue-derived age-related changes and investigate relationships between changes in biomarker levels and hallmarks of aging, such as cognitive decline and increased inflammation. A second trial run by the Stanford Memory and Aging Study aims to identify proteins in blood and cerebrospinal fluid (CSF) of healthy older adults (60–90 years old) that correlate with changes from magnetic resonance imaging (MRI), neuropsychological and neurological testing, and memory performance96. Stanford University is also conducting a longitudinal study, the Healthy Brain Aging Study, that seeks to identify blood and CSF biomarkers associated with MRI and cognitive testing in patients with dementia-related neurodegenerative diseases97. Uniquely, this study aims to follow enrollees over time, ending in eventual brain donation to obtain a more complete assessment of disease progression. Collectively, these biomarker studies will be crucial in identifying potential indicators of human aging and early development of neurodegenerative disease-related cognitive decline.
To explore the translational potential of young plasma as a treatment for neurodegenerative disease-related cognitive decline, two major studies were independently initiated. The first study, PLasma for Alzheimer SymptoM Amelioration (PLASMA), was initiated in 2014 by Stanford University in collaboration with Alkahest, a company based in San Carlos, California98. This study enrolled patients with mild-to-moderate Alzheimer's disease for infusions of one unit of plasma derived from males aged 30 years or younger once weekly for four weeks. While the primary outcome is to determine the feasibility and safety of young plasma administration, the researchers will assess memory performance, psychological status, and MRI analysis. Concurrently, they will analyze plasma levels of factors previously associated with neurodegenerative disease. The second study, Young Donor Plasma Transfusion of Age-Related Biomarkers, is being conducted by the company Ambrosia, a startup based in Monterey, California99. In contrast to the PLASMA study, enrollees in the Ambrosia trial can be healthy or disease-affected individuals over 35 years of age. Enrollees will receive a single infusion of plasma from young donors aged 16–25. Researchers will then assess a specific panel of plasma factors previously associated with aging and neurodegenerative hallmarks, such as inflammation, neurogenesis, and amyloid plaque deposition. Of note, a number of concerns have been raised with the design of the Ambrosia-led study, including the omission of an independent placebo control group100. Lastly, the potential of using young plasma interventions is also being explored in a broader context of neurodegenerative disease, including Parkinson's disease101, progressive supranuclear palsy102, and acute stroke103. While any of these studies has yet to yield results, the outcomes from the young plasma administration trials, coupled with the perspective gained from the extensive biomarker studies, will provide a more complete picture of potential pro-aging and rejuvenating circulating factors that can be targeted to increase healthspan and ameliorate neurodegenerative disease in the elderly.
Systemic interventions, such as CR, exercise, and young blood administration, have reliably ameliorated many facets of functional and cognitive decline in rodent models of aging and neurodegenerative disease by targeting molecular and cellular pathways conserved across species (Figure 1). Collectively, the rejuvenating power of these systemic interventions has peaked interest in the public at large for their potential therapeutic translation to humans. Coupled with the most recent reports that human cord-blood-derived plasma proteins can also counteract age-related cognitive impairments in aged mice, the possibility for brain rejuvenation strategies in humans is increasingly strengthened. To that end, multiple clinical trials are now underway to test the safety and efficacy of systemic brain rejuvenation strategies in the elderly under normal aging and neurodegenerative disease conditions. While current clinical trials have limitations, they are based on a strong foundation of animal research, with much reason for excitement in the potential to develop effective systemic strategies for the treatment of dementia-related neurodegenerative disease. Looking toward the future, barriers posed by the idiopathic nature of many neurodegenerative diseases, unknown mechanisms driving the rejuvenating effects of blood, and added inter-individual variability introduced by human subjects must now be overcome in the pursuit to fully tap into the therapeutic potential of systemic brain rejuvenation strategies. Additionally, remaining questions as to how pro-aging and rejuvenating factors in blood can be manipulated in concert to maximize the potential therapeutic benefit to patients affected by neurodegenerative disease must also be addressed, providing a rich scientific field of inquiry to explore. Broadly, future research investigating the potential for brain rejuvenation may not only impact human health but also yield fundamental understanding of the biological mechanisms governing the aging process itself.
APP, amyloid precursor protein; B2M, beta-2 microglobulin; CR, caloric restriction; CSF, cerebrospinal fluid; CSF2, colony-stimulating factor 2; GDF11, growth differentiation factor 11; LTP, long-term potentiation; MRI, magnetic resonance imaging; PS1, human presenilin 1; TGF-β, transforming growth factor beta; TIMP2, tissue inhibitor of metalloproteinases 2.
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)