Keywords
MRM, Multiple Reaction Monitoring, targeted proteomics, Treponema pallidum, syphilis, biomarker discovery, antigen test, plasma
MRM, Multiple Reaction Monitoring, targeted proteomics, Treponema pallidum, syphilis, biomarker discovery, antigen test, plasma
Changes to the text were made to clarify the results along the lines suggested by the reviewers. These changes included:
- Further details clarifying the limit of detection
- Minor typographical errors
- An additional reference was added to reflect recent experimental efforts of Edmondson et al. to culture T. pallidum in vitro.
The interpretation of results and study conclusions remain unchanged.
See the authors' detailed response to the review by Timothy Palzkill
See the authors' detailed response to the review by Mohd M. Khan
hPTPs Isotopically labelled proteotypic surrogate peptides
LOD Limit of detection
MSM Men who have sex with men
PCR Polymerase chain reaction
TPPA Treponema pallidum Particle Agglutination test
RPR Rapid plasma reagin test
COG Clusters of Orthologous Groups
FA Formic acid
PBS Phosphate buffered saline
NSAF Normalized spectral abundance factor
IQR Interquartile range
SISCAPA Stable Isotope Standards and Capture by Anti-Peptide Antibodies
Treponema pallidum ssp. pallidum (T. pallidum), a culturable1 microaerophilic spirochete, is responsible for more than 8 million new cases of syphilis per year2. There has been a resurgence of syphilis in a number of world regions over the last two decades2–4. In Europe3 and North America4, this increase has been most marked in men who have sex with men (MSM). A striking feature of these outbreaks has been the increasing proportion of cases that are occurring in patients with a previous diagnosis of syphilis5,6. Patients with reinfections are more likely to present with asymptomatic or less symptomatic disease5, hence the diagnosis of reinfection is wholly dependent on subtle changes in serological tests7. Two types of serological tests are used to diagnose syphilis: treponemal tests detect antibodies to T. pallidum and non-treponemal tests, such as the Rapid plasma reagin (RPR) test, detect agglutination secondary to the presence of anti-lipoidal antibodies reactive to material released from damaged host cells and possibly cardiolipin released from T. pallidum8. Treponemal tests remain positive for life and are therefore of no use in the diagnosis of reinfection. Non-treponemal tests are used for syphilis post-treatment follow-up and diagnosis of reinfection. A wide range of factors can result in increases in test titers, causing syphilis to be over-diagnosed and unnecessarily treated7,9–11. Direct T. pallidum detection techniques, including various nucleic acid amplification tests, have been developed, but apart from testing of primary ulcer specimens the sensitivity of these techniques is low12. Even in the setting of secondary syphilis, when there is a high T. pallidum load in the blood13, the sensitivity of polymerase chain reaction (PCR) tests reaches only 52 % on serum specimens12,14.
The T. pallidum genome, through evolutionary reduction, is one of the smallest of the human bacterial pathogens, with a predicted 1044 open reading frames15. Approximately half of the predicted proteins have been detected through MS techniques16,17, including the semi-quantification of T. pallidum proteins using spectral counting17. A T. pallidum transcriptome study demonstrated that almost all genes were expressed during peak rabbit experimental infection18. This maximum utilization of the genome, well characterized proteome, and swift invasion of the organism into the bloodstream (within 24 hours after infection19) make this pathogen an ideal candidate for antigen diagnostic assay development. A variety of antigen tests against other pathogens have been designed for clinical samples such as blood, cerebrospinal fluid, faeces and urine; and these have proven their utility in the diagnosis and assessment of therapeutic response in a number of infections, including Helicobacter pylori20, Cryptococcus neoformans21, Cryptosporidium ssp.22, Entamoeba histolytica23, Ebola virus24 and Mycobacteria tuberculosis25. If a highly sensitive and specific test could be developed that is able to confirm the presence or absence of T. pallidum in the body then this would be of considerable utility in the diagnosis of syphilis reinfections and in assessing therapeutic response. It could also be useful for the diagnosis of neuro- and congenital syphilis – two diagnoses where contemporary tests are suboptimal26.
During the last decade, advanced MS-based proteomics platforms have emerged as mainstay bioanalytical tools for a broad range of clinical applications, including targeted protein identification27 and bacteria identification and typing28. Particularly the AQUA workflow29,30, with its use of stable isotopically labelled standard proteotypic peptides (henceforth referred to as ‘heavy’ PTPs or hPTPs) and selected/multiple reaction monitoring-mass spectrometry (SRM/MRM MS), has emerged as a powerful technique for the fast determination of multiple protein concentrations in highly complex sample matrixes such as urine (reviewed by Mermelekas et al.31) and plasma (reviewed by Pernemalm and Lehtiö32). Precise quantitation of proteins is possible by using hPTPs as internal standards that correspond to endogenous peptides created during the enzymatic digestion of the sample of interest. When combined, the endogenous and synthetic peptides elute together chromatographically and ionize with the same efficiency. Since the quantity of the labelled peptide is known, the absolute quantity of the targeted native protein can be determined by comparing MRM hPTP/endogenous peak areas. The precision and utility of this highly sensitive multiplexed method has been demonstrated on undepleted/ unenriched plasma for the detection of a panel of human cardiovascular disease33 and cancer34 biomarkers with a detection capability of four orders of magnitude (103–104 range in protein concentration) and up to femtomolar level sensitivity in plasma35. Recently, a panel of 136 cancer candidate biomarkers was interrogated in unenriched urine samples using MRM, revealing detection limits of up to 25 picogram/ml urine36.
With regards to infectious disease biomarker studies, MS-based approaches identified candidate biomarkers in urine for Leishmania sp.37, which has led to the development of a urine capture ELISA diagnostic test38. Considerable progress has also been made in Mycobacterium tuberculosis39–41 biomarker studies; recent advancements include the detection of M. tuberculosis in urine using IgG capture, immunodepletion and MRM methods42 and MRM assay of exosomes isolated from serum samples from patients with tuberculosis39.
In this study, we investigated if T. pallidum proteins could be detected in plasma and urine samples from individuals with syphilis using a targeted proteomics (MRM) approach. Successful development of a T. pallidum antigen test will most likely be contingent upon the simultaneous detection of multiple protein biomarkers to comprehensively cover different stages of disease. Eleven T. pallidum protein biomarkers were chosen based on a predicted specificity, high predicted abundance, and physiochemical properties. Thirty surrogate hPTPs were synthesized corresponding to eleven candidate T. pallidum biomarkers. Analysis of eighteen plasma and four urine samples revealed no detectable MRM signal for the endogenous peptides from the biomarkers of interest. This is likely due to the extremely low (femtomoles per liter) predicted concentration of bacterial proteins in the samples of interest, or the fact that the biomarkers are not expressed during infection. T. pallidum spiking experiments established a MS detection limit of 300 bacteria/ml in PBS; polyclonal anti-T. pallidum magnetic bead enrichment did not improve the protein detectability.
Between January 2014 and August 2015, 120 patients attending the Institute of Tropical Medicine Antwerp clinic, over the age of 17 years, and in whom a new diagnosis of syphilis was made and had not received antibiotics in the preceding thirty days, were recruited into the cohort study. Thirty HIV-positive controls, in whom the diagnosis of syphilis was excluded via serological and PCR testing, were also recruited. The diagnosis and staging of syphilis was according to the Centers for Disease Control and Prevention classification43, and treatment was administered according to European guidelines44. All patient sera were tested for syphilis using a RPR test (BD Macro-Vue RPR card test, Becton, Dickinson and Co., Sparks, MD, United States of America (USA)) and an antibody detection Treponema pallidum Particle Agglutination test (SERODIA-TPPA Fujirebio Inc., Tokyo, Japan). A PCR test targeting T. pallidum polA was also performed on serum45 and whole blood samples were tested for multiple gene targets46, as previously described. Selection criteria of participants from the cohort study for the MRM assay analysis included a range of syphilis clinical stages and prioritized predicted high bacterial loads, as demonstrated by positive PCR tests and/or high RPR titres. Patients with early stage syphilis (primary, secondary, early latent) that were plasma and/or whole blood PCR positive for T. pallidum were expected to have the highest bacterial load12,13.
Plasma was collected immediately before Benzathine Penicillin G intramuscular injection using 7.5 ml EDTA-coated blood collection tubes (Sarstedt Monovette, Nümbrecht, Germany). We refer to these samples as the pre-penicillin samples. A selection of randomly selected patients participated in an additional blood draw three hours after penicillin treatment since studies have shown penicillin to be fast acting on T. pallidum, leading to consequent cell lysis and antigen release47. These samples are termed the post-penicillin samples. Plasma was chosen for the MRM assay according to HUPO guidelines48. Protease inhibitors were not added to the plasma samples since previous studies did not demonstrate a significant higher protein yield with treated samples49 and peptides could inadvertently be modified50. Plasma were subjected to dual centrifugation in an Eppendorf 22331 centrifuge (Hamburg, Germany) in an effort to minimize cellular contamination: whole blood was centrifuged at 2000 g for 10 minutes at ambient temperature, followed by transfer of the plasma fraction to a 50 ml falcon tube and centrifugation at 2400 g for 15 minutes. All plasma were processed and aliquoted into cryovials for storage at -80 °C in a long-term freezer unit (Eppendorf U725-G Innova New Brunswick, Hamburg, Germany) until further testing. Mid-stream random-void urine samples were collected and processed following HUPO guidelines51, including centrifugation for 10 minutes at 2000 g at ambient temperature in order to remove insoluble contents such as cells and casts. Urine was aliquoted into 15 ml falcon tubes and stored at -80 °C until further testing. All plasma and urine samples were processed within three hours of collection and were only subjected to one freeze thaw cycle.
In a previous descriptive study we used non-gel based complementary MS techniques to characterize the proteome of in vivo rabbit cultured T. pallidum17. Candidate T. pallidum biomarker proteins for the MRM assay were chosen based on the following specific criteria: relative protein abundance (based on semi-quantitative spectral counting techniques17), Clusters of Orthologous Groups (COG) functional categorization, microarray transcriptome data18, protein size, physicochemical properties (i.e. previously detected by MS), predicted subcellular localization17 and literature review. Each of the candidate biomarkers were digested in silico by subjecting the FASTA-formatted sequences to tryptic digestion, assuming 100 % digestion efficiency. Proteotypic peptides (PTPs) corresponding to these proteins were determined using ESPPredictor52 and pBLAST53; analysis of the protein and PTPs was performed to determine possible homology with other bacterial species and human proteins. After PTP selection was finalized, isotopically labelled synthetic peptide standards (hPTPs) corresponding to the selected PTPs were synthesized (Heavy Peptide™ AQUA Basic with > 95 % purity; Thermo Fisher Scientific, Ulm, Germany).
Protein concentrations of urine and plasma samples were determined based on the area under curve at 214 nm using a RP-C4 column (Vydac 214TP5415; 4.6×150 mm, particle size 5 μm; Alltech Associates Inc., Lokeren, Belgium) coupled to an Alliance e2695 HPLC system equipped with a 996 PDA detector (Waters Corporation, Milford, MA, USA). For each sample, 250 µg of protein was precipitated by adding six volumes of ice cold LC-MS grade acetone (Biosolve, Valkenswaard, the Netherlands) followed by overnight incubation in freezer unit (Liebherr, Bulle, Switzerland) at -20 °C. In all cases, lo-bind Eppendorf tubes (Eppendorf, Hamburg, Germany) were used to ensure high recovery rates of proteins and peptides. Protein pellets were re-suspended in 50 mM Tris-HCl/6 M urea/5 mM dithiothreitol /10 % beta-mercaptoethanol (25 µL/100 µg protein) at pH 8.7. For the denaturation and reduction process all samples were incubated at 65 °C in a hot water bath for 1 hour. Subsequently, proteins in all fractions were diluted in 50 mM Tris-HCl/ 1 mM CaCl2 (75 µL/100 µg protein) and alkylated by adding 200 mM iodoacetamide (10 µL/100 µg protein) during 1 hour at ambient temperature and protected from light. Proteomics-grade modified trypsin (Promega, Madison, WI, US) was added at a 30:1 protein-to-enzyme ratio. After incubation at 37 °C in a hot water bath for 18 hours the digestion was stopped by freezing the samples. Protein digests were desalted by SPE using GracePure SPE C18-Max (50 mg) (W. R. Grace & Co., Columbia, MD, US) RP cartridges and a vacuum manifold. SPE cartridges were conditioned with 100 % methanol and equilibrated with 100 % LC/MS grade H2O and 0.1 % formic acid (FA). After loading the complete acidified (0.1 % FA) tryptic digest, peptides were washed with 10 % methanol and eluted with 40 % methanol/ 40 % acetonitrile (ACN) and 0.1 % FA. Eluted peptides were lyophilized and frozen at -20 °C until further analysis. Immediately before analysis, lyophilized digests were resuspended in 5 % ACN/0.1 % FA and spiked with a mixture of all hPTPs.
Optimization of each PTP was performed on a triple quadruple mass spectrometer (Waters Xevo TQ, Waters Corporation, Milford, MA, US) in order to obtain the most intense transitions. The capillary voltage was tuned to approximately 2 kV with a source temperature of 150 °C. Desolvation temperature was set at 400 °C with a nitrogen gas flow of 800 L/h. Cone voltage, collision energy and dwell times were optimized for each of the PTPs. All PTPs were dissolved in mobile phase A (MP-A), containing 5 % ACN (LC/MS grade) and 0.1 % FA. For each of the peptides individually, the Limit of Detection (LOD) was determined by performing a dilution series in MP-A. Based on these concentrations, a mixture of all hPTPs was made. A balanced hPTP mixture has been shown to increase quantification accuracy and reproducibility compared to an equimolar mixture in previous studies35. To check for possible suppressive effects of the plasma matrix, the hPTP mixture was spiked into plasma from a control study subject. A balanced mixture of hPTP (concentrations detailed in Supplementary File 1) was spiked into 50 µg of plasma digest. Chromatographic separation of the plasma and urine samples was performed on a RP-C18 UPLC column (Waters, CSH 150 × 2.1 mm, 1.7 µm at 35 °C) connected to an Acquity UPLC system (Waters Corporation, Milford, MA, USA). In order to separate all peptides as best as possible, an optimized linear gradient of Mobile Phase B (MP-B) (0.1 % FA in 100 % ACN) was applied: 5 % MP-B during 1 min and from 5 to 35 % MP-B in 5 min, followed by a steep increase to 100 % MP-B in 1 min, all at a flow rate of 300 µL/min. Based on the specific retention times of each peptide, three scheduled MRM runs of 10 minutes were generated, each of them containing 20 MS1 channels (10 endogenous (T. pallidum) PTPs without isotopic label and 10 channels with a synthetic hPTP equivalent). At least three transitions (ion pairs) were selected for each peptide of interest. For each scheduled MRM analysis, 50 µg of peptides (injection loop of 5 µL) per plasma/urine sample were loaded onto the analytical column. In addition to an extensive needle wash after each injection, a blank run was performed between two subsequent clinical samples to prevent carry-over effects. Data acquisition was controlled by MassLynx version 4.1, while targeted datasets were analysed by TargetLynx, which is part of MassLynx (Waters Corporation, Milford, MA, USA). All Xevo TQ MS raw spectral files are available at PeptideAtlas54 with the identifier PASS00978.
T. pallidum protein enrichment was performed using magnetic beads (Dynabeads® M-270, Life Technologies, CA, USA) coated with biotin-conjugated polyclonal T. pallidum- specific antibodies (PA1-73103, Thermo Fisher Scientific, CA, USA) through streptavidin-biotin conjugation. According to the manufacturer’s protocol, 10 µg of antibody was used to bind 1 mg of beads (approximately 5 × 107 beads).
In vivo rabbit cultured purified T. pallidum DAL-1 strain extracts55,56 were kindly provided by the group of David Šmajs from the Masaryk University, Czech Republic. The original concentration of the T. pallidum extract was approximately 106 bacteria/ml as quantified under darkfield microscopy using a Olympus BX41 (Olympus Corporation, Tokyo, Japan) equipped with darkfield microscope condenser DCW 1.4-1.2; magnification 10×40. Samples were stored in 1 ml phosphate buffered saline (PBS) and only subjected to one freeze-thaw cycle. Two dilution series of T. pallidum were prepared, each time starting in 1 ml of PBS and finally equating to eight approximate bacterial concentrations: 104, 103, 300, 100, 33, 10, 3 and 0 bacteria/ml.
For one dilution series, each of the eight fractions were incubated with a constant amount (~105) of magnetic beads coated with polyclonal anti-T. pallidum antibodies. After incubation for two hours at 4° C and magnetic separation, the supernatant was discarded and beads were washed three times with PBS. To lyse the antibody bound bacteria, 1 ml of PBS was added to each bead sample, these were sonicated on ice using a Sonics Vibra Cell VC130 (Sonics and Materials Inc., Newtown, CT, USA) (two times 30 seconds with an amplitude of 50 %). The bead fraction was retained (retentant) after sonication by using magnetic separation. Released proteins were precipitated adding ice-cold acetone and incubated overnight at -20 °C. Tryptic digestion was performed, following the aforementioned procedure, on both the precipitated proteins (supernatant) and directly “on-bead” (retentate), to test for possible unreleased proteins during sonication. For the second dilution series (unenriched), 1 ml was directly drawn from each of the eight samples. The samples from this series were also sonicated on ice (two times 30 seconds with an amplitude of 50 %) to lyse the bacteria. Released proteins were then acetone precipitated and subsequently digested, in conformance with the other parallel series procedure.
Peptide mixtures were separated by RPLC on a Waters nano-UPLC system using a nanoACQUITY BEH C18 Trap column (100 Å, 5 μm, 180 μm × 20 mm) connected to a nanoACQUITY BEH C18 analytical Column (130 Å, 1.7 μm, 100 μm × 100 mm) (Waters Corporation, Milford, MA, USA). Peptides were dissolved in MP-A, containing 2 % ACN and 0.1 % FA and spiked with 20 fmol [Glu1]-fibrinopeptide B, which serves as an internal calibrant. A linear gradient of MP-B (0.1 % FA in 98 % ACN) from 2 to 45 % MP-B in 45 min, followed by a steep increase to 95 % MP-B in 2 min at a flow rate of 400 nl/min. The nano-LC was coupled online with a LTQ Orbitrap Velos (Thermo Scientific, San Jose, CA, US) mass spectrometer using a PicoTip Emitter (New Objective, Woburn, MA, US) linked to a nanospray ion source. The mass spectrometer was set up in a data dependent acquisition MS/MS mode where a full scan spectrum (350–2500 m/z, resolution of 60.000) was followed by a maximum of ten CID tandem mass spectra (100 to 2000 m/z). Peptide ions were selected as the twenty most intense peaks of the MS scan. CID scans were acquired in the LTQ ion trap part of the mass spectrometer with normalized collision energy of 32 %.
Obtained spectra were screened against the T. pallidum reference and resequenced databases (UniProt ID proteome UP00001425915 and UP00000081157 using the MASCOT search engine (Matrix Science; version 2.1.03) based on the digestion enzyme trypsin. Carbamidomethylation of cysteines was listed as a fixed modification, while methionine oxidation was set as a variable modification. A maximum of one missed cleavage was tolerated. Mass tolerance was set to 10 ppm for the precursors and 0.8 Da for the fragment ions. False discovery rate was set at 5 %. Scaffold Q+ (version 4.6.2, Proteome Software Inc., Portland, OR, US) was used to validate MS/MS-based peptide and protein identifications. Protein identifications were accepted if they could be established at greater than 95.0 % probability according to the protein prophet algorithm58.
All LTQ-Orbitrap MS/MS raw spectral data is available at PeptideAtlas54 with the identifier PASS00978.
Eighteen syphilis-infected study participants were selected for the MRM assay analyses (Table 1). All participants were male and identified as MSM. A third of the participants (6/18; 33 %) were HIV positive. Five (28 %) presented with primary, eleven secondary (61 %), and two early latent (11 %) stage disease. Thirteen participants were confirmed T. pallidum-positive by serum and/or whole blood PCR testing. Four participants had indeterminate PCR results, meaning their sample was weakly positive. A second confirmatory PCR was not performed on these samples. One patient was negative for both whole blood and serum PCR. All participants tested positive with both the RPR and TPPA tests. The median RPR value was 1/64 (Interquartile range (IQR): 1/16- 1/128). In total, 22 samples were analysed, including N = 12 pre-penicillin treatment plasma, N = 6 post-penicillin treatment plasma and N = 4 pre-penicillin treatment urine samples.
| Patient Number | HIV status | Syphilis stage | Sample type | Pre or post- treatment sampling# | PCR Whole Blood | PCR Serum | RPR titre | TPPA titre |
|---|---|---|---|---|---|---|---|---|
| 1 | Positive | Secondary | Plasma | Pre | Positive | Indet. | 1/512 | >1/20480 |
| 2 | Negative | Primary | Plasma | Pre | Negative | Indet. | 1/4 | 1/160 |
| 3 | Positive | Early latent | Plasma | Pre | Negative | Positive | 1/1 | 1/1280 |
| 4 | Positive | Secondary | Plasma | Pre | Positive | Positive | 1/128 | 1/20480 |
| 5 | Positive | Secondary | Plasma | Pre | Positive | Positive | 1/128 | >1/20480 |
| 6 | Negative | Secondary | Plasma | Pre | Negative | Positive | 1/128 | >1/20480 |
| 7 | Positive | Early Latent | Plasma | Pre | Positive | Positive | 1/64 | 1/10240 |
| 8 | Positive | Secondary | Plasma | Pre | Positive | Indet. | 1/32 | 1/1280 |
| 9 | Positive | Secondary | Plasma | Pre | Positive | Positive | 1/512 | >1/20480 |
| Urine | ||||||||
| 10 | Negative | Primary | Plasma | Pre | Positive | Indet. | 1/16 | 1/5120 |
| Urine | ||||||||
| 11 | Positive | Secondary | Plasma | Pre | ND | Indet. | 1/128 | >1/20480 |
| Urine | ||||||||
| 12 | Negative | Secondary | Plasma | Pre | Positive | Negative | 1/32 | >1/20480 |
| 13 | Positive | Secondary | Plasma | Post | ND | Indet. | 1/128 | >1/20480 |
| 14 | Negative | Primary | Plasma | Post | Positive | Indet. | 1/16 | 1/5120 |
| Urine | ||||||||
| 15 | Negative | Primary | Plasma | Post | Negative | Indet. | 1/8 | 1/1280 |
| 16 | Positive | Secondary | Plasma | Post | Positive | Negative | 1/64 | 1/20480 |
| 17 | Positive | Primary | Plasma | Post | Positive | Negative | 1/64 | >1/20480 |
| 18 | Positive | Secondary | Plasma | Post | Negative | Negative | 1/128 | >1/20480 |
Eleven T. pallidum proteins were selected as candidate biomarkers (Table 2). Most selected biomarkers had high normalized spectral abundance factor (NSAF) scores according to our previous study17 (median 4.02; IQR: 1.97-6.97) and high microarray signal ratios18 (median 3.05; IQR: 0.74-6.8). The median protein molecular weight was 39 kDa (IQR: 28-81). Two proteins were predicted to be located in the flagellum (TP_0249 and TP_0792), two in the ribosome (TP_0250b and TP_0244) and the subcellular localization of five proteins was unknown. Protein TP_0326, a BamA orthologue, has been experimentally shown59–61 to be localized in the outer membrane. A typical target for PCR assays is polA, coding protein TP_010562. One protein, Peptidyl-prolyl cis-trans isomerase (TP_0862) was found in a previous proteomics study where it demonstrated moderate reactivity during immunoblot experiments with human and rabbit T. pallidum infected serum16. Protein TprG (TP_0317) is part of the paralogous tpr gene family that encodes candidate virulence factors63 and is partially homologous to Tpr E/J. According to pBLAST analysis, all chosen biomarker proteins and corresponding PTPs did not demonstrate high homology with other pathogens, non-pathogenic commensal bacterial or human proteins (data not shown). One to three corresponding well-suited PTPs were selected for each biomarker, for a total of 30 PTPs. Details pertaining to these are provided in Table 2.
| Number | UniProt Accession Number* | TP Number/ gene | Protein Name | Peptide Number | Peptide Sequence# | Protein Weight (kDa) | Predicted Subcellular Location& | COG category function | Spectral Count NSAF value in T. pallidum17 | cDNA/ DNA signal ratio18 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | R9US76 | TP_0105/ polA | DNA-directed DNA polymerase | 16 | TSAVSGAIPIENR | 112 | NK | L | 2.55 | 0.283 |
| 17 | MALNTQIQSSAADIVK | |||||||||
| 18 | VHTSFVQIGTATGR | |||||||||
| 2 | O83346 | TP_0326/ tp92 | Putative outer membrane protein assembly factor | 19 | TEAGGVVVQFTIQEGK | 94 | Outer Membrane | M | 1.77 | 0.682 |
| 20 | EQWASSPGLAESFR | |||||||||
| 21 | LAFANTFTSPGGIPK | |||||||||
| 3 | R9UVD9 | TP_0249/ flaA1 | Flagellar filament outer layer protein | 22 | LATEVGFTPSGGAQR | 39 | NK | N | 7.47 | 16.05 |
| 23 | DESVLIDFAK | |||||||||
| 4 | O83834 | TP_0862/ fklB | Peptidyl-prolyl cis-trans isomerase | 24 | GTLLDGTVFDASR | 28 | NK | O | NF | 5.29 |
| 25 | KPGVQVTSSGLQYEVVK | |||||||||
| 27 | FYVPSSLGYGER | |||||||||
| 5 | O83892& | TP_0922 | Uncharacterized Protein | 26 | MPPSPCAVLR | 33 | NK | None | 7.80 | 1.599 |
| 30 | VASVVVISVDNR | |||||||||
| 28 | YFLPGECAGR | |||||||||
| 6 | R9USJ3 | TP_0250b/ rpsT | 30S ribosomal protein S20 | 29 | LYNGVFSSPEVVR | 11 | Ribosome | None | 5.28 | 3.39 |
| 7 | R9UU30 | TP_0244/ rpsG | 30S ribosomal protein S7 | 10 | TGEEPLPVFTK | 18 | Ribosome | J | 6.78 | 3.053 |
| 9 | ATAVGIMYDCLER | |||||||||
| 11 | LAAEILDAYHSTGTAFK | |||||||||
| 8 | O83337 | TP_0317 | Tpr protein G | 1 | VLDAVTAATETALQSR | 81 | NK | None | 1.32 | 0.743 |
| 8 | GNPMSLFNLPDQQK | |||||||||
| 2 | LTGSATLEWGISYGK$ | |||||||||
| 9 | P21991 | TP_0792/ flaB1 | Flagellar filament core protein FlaB | 6 | ELSVQAANGIYSAEDR | 31 | Flagellum | N | 6.97 | 13.82 |
| 7 | DAGDESVMNIDSPEK | |||||||||
| 12 | AYIGTMTAVAMGIR | |||||||||
| 10 | R9UTS8 | TP_0748/ cfpA/ tpn83 | Cytoplasmic filament protein A | 3 | GVNELETHTNSLLR | 79 | Cytoplasm | S | 2.75 | 6.79 |
| 4 | ADIGQSFASDGSADQK | |||||||||
| 5 | EYDDTDISNLPDER | |||||||||
| 11 | O83417 | TP_0402/ fliI | IIISP family Type III (Virulence- related) secretory pathway protein/ Flagellum-specific ATP synthase | 13 | EIGLASGELPATR | 48 | Flagellum; Cytoplasm | NU | 1.97 | 1.241 |
| 14 | SVIVSATSDESPLAR | |||||||||
| 15 | VGAYQQGSDAELDR |
Legend: *- UniProt proteome ID UP000014259; &- ORF was not annotated in the re-sequenced Nichols strain genome due to its length below the 150 bp limit15; #- underlined/bold amino acids indicate stable isotope labelled residues; $- peptide is homologous in Tpr E/G/J protein sequences; @- subcellular location as reported in Osbak et al.17; NK- not known; NSAF- normalized spectral abundance factor; COG- clusters of orthologous groups; COG categories: L- Replication, recombination and repair, M- Cell wall/membrane/envelope biogenesis; N- Cell motility; O- Posttranslational modification, protein turnover, chaperones; J- Translation, ribosomal structure and biogenesis; S- Function unknown; U- Intracellular trafficking, secretion, and vesicular transport.
The LOD for each peptide was determined individually by performing a dilution series of MP-A whereby the median LOD was 68.5 (IQR 14.2-176.7) picomoles. Once the peptide mixture composition was optimized based on the LOD, 2 µL of this mixture (Supplementary File 1) was spiked into 50 µg plasma from a control patient whereby no significant variations in the signal of the hPTP transitions could be detected, indicating that there was no evidence of transition interference from the plasma. After optimizing each of the PTPs, three different sets of transitions were combined in an MRM assay based on their chromatographic retention time, as detailed in Supplementary File 1. The experiments contained a total of 141 targeted ion pairs (transitions) corresponding to 30 PTPs from eleven T. pallidum proteins. Ten of the eleven proteins were represented by two or more (h)PTPs (Table 2/ Supplementary File 1). In total, three scheduled MRM assays of 10 minutes, each containing 20 peptides (10 endogenous (T. pallidum) peptides and 10 hPTP standards) were developed. These assays were evaluated based on a balanced mixture of all 30 hPTPs standards. Unfortunately, although each of the 30 spiked hPTPs could be detected, none of the selected endogenous T. pallidum peptides could be identified in any of the MRM assays (Figure 1; Supplementary File 2*).

(a) synthetic hPTPs, even numbers and (b) endogenous (T. pallidum) PTPs, odd numbers; gradient 1 of 3. For each peptide the number of selected transitions (channels) is reported. The x-axis shows the chromatographic retention time of the corresponding peptide while the y-axis shows the relative intensity of the MS2 signal. Note: Signal fluctuations present in the ‘endogenous’ PTP chromatogram are always the result of just one transition, often coupled with a shift in retention time and differing m/z-values differ from the hPTP run, thus these are considered to be noise.
Two T. pallidum spiking dilution series were prepared in PBS and subjected to LTQ-Orbitrap MS/MS analysis in order to estimate the LOD of MS detection. One of the series was subjected to an additional polyclonal antibody coupled magnetic bead enrichment step, including sonication of the beads and subsequent separate measurement of the lysate and on-bead digestion retentate (Figure 2).

In total, eight different concentrations of T. pallidum (from 104 to 0 bacteria/ml PBS) were treated in three different ways i) T. pallidum was enriched using magnetic beads coated with polyclonal anti-T. pallidum antibodies and lysed by sonication for release of T. pallidum proteins in the supernatant. Acetone precipitated proteins were trypsinized; ii) In order to detect any remaining protein on the beads, the beads were also trypsinized (retentant on-bead trypsinization); iii) As a control, non-enriched samples were sonicated and immediately trypsinized. *-proteins selected as candidate biomarkers in this study. All samples were analysed by an LTQ-Orbitrap mass spectrometer.
Two unique T. pallidum proteins, Cytoplasmic filament protein A (TP_0748) and Lipoprotein antigen Tp47 (TP_0574), were found in the 300 bacteria/ml fraction in the enriched and unenriched samples, respectively (Figure 3; Supplementary File 3). Therefore, the LOD based on a high-resolution LTQ-Orbitrap instrument was approximately 300 bacteria/ml PBS for both the antibody enriched and unenriched samples, meaning there was no significant improvement in LOD using bead enrichment. No proteins were detected in any sample concentrations for the enriched bacterial lysate (supernatant) fraction. Possibly, the sonication conditions were not harsh enough to lyse the bacteria on the beads and lysis was mainly the results of trypsin treatment under denaturing conditions. In total, eight unique T. pallidum proteins were found in both the unenriched and enriched retentate dilution series in one or more of the concentrations analyzed: 60 kDa chaperonin (TP_0030), Flagellar filament outer layer protein flaA1 (TP_0249), Alkyl hydroperoxide reductase (TP_0509), Lipoprotein antigen Tp47 (TP_0574), Galactose ABC superfamily ATP binding cassette transporter, binding protein (TP_0684), Cytoplasmic filament protein A (TP_0748) and the Flagellar filament core proteins flaB1/B3 (TP_0792/TP_0870). Four proteins, Lipoprotein, 15 kDa (TP_0171), 10 kDa chaperonin (TP_1013), Elongation factor Tu (TP_0187) and Tp34 lipoprotein (TP_0971) were only found in the unenriched and enriched series, respectively. Ten unique T. pallidum proteins were found in the highest concentration (104 bacteria/ml) for in the enriched retentate sample (N = 10) and non-enriched sample (N = 10), two proteins detected were unique to either the enriched or unenriched samples (Figure 3). Five unique T. pallidum proteins were found in the 103 bacteria/ml sample, including N = 4 in the unenriched and N = 4 in the retentate fractions. A peptide (LSGGVAVIK) related to 60 kDa chaperonin (TP_0030) was detected in the low concentration (100/ 33/ 10/ 3 bacteria/ml) and in the negative control samples of the enriched sample series. This was likely a false-positive non-specific peptide secondary to rabbit protein contamination since this short peptide sequence is closely homologous to the Oryctolagus cuniculus (rabbit) 60 kDa heat shock protein, or could have originated from the beads or antibodies. As a result, it has been excluded from the analysis. Three T. pallidum proteins detected in both the enriched and unenriched sample series were also biomarker candidates tested in the MRM assay experiments: Flagellar filament core protein flaB2 (TP_0792), Cytoplasmic filament protein A (TP_0748) and the Flagellar filament outer layer protein flaA1 (TP_0249). Detailed information about the identified proteins, peptides, coverage and search parameters can be found in Supplementary File 3. Rough concentration calculations estimated that our target PTPs would be present in the femtomoles per liter range in human T. pallidum infection (calculations presented in Supplementary File 4).
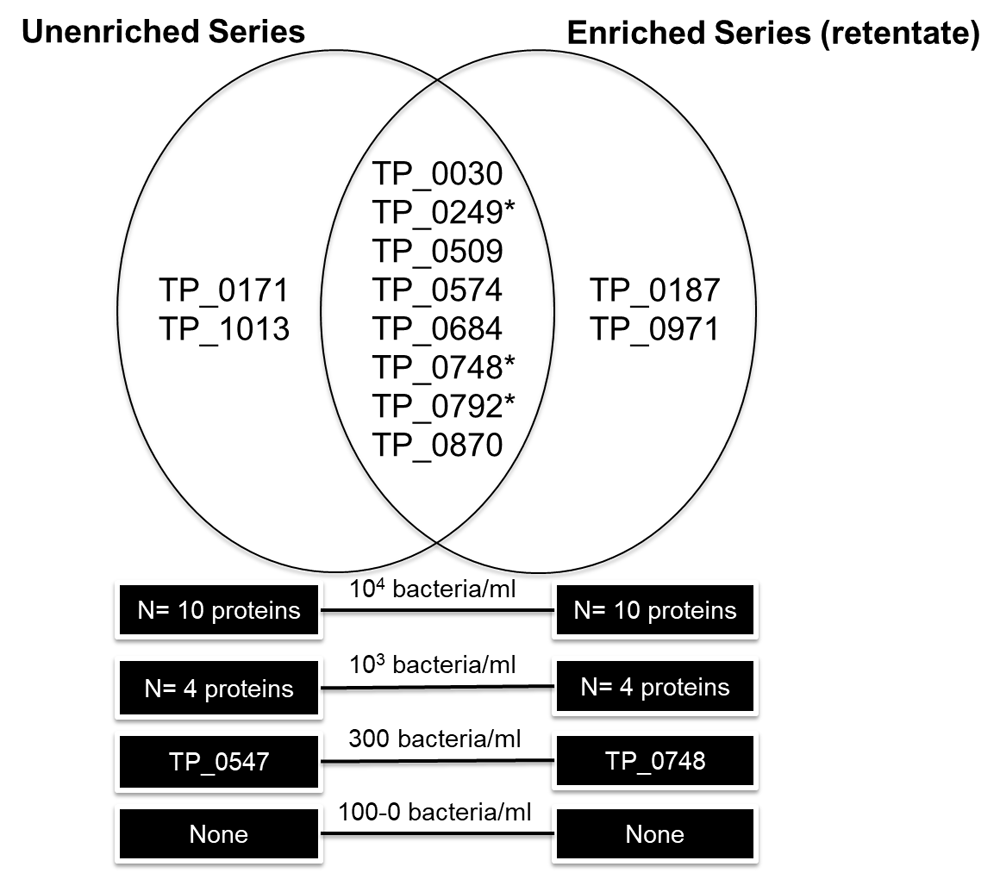
*-proteins selected as candidate biomarkers in this study.
The T. pallidum MRM assay designed in this study failed to detect any of the 30 targeted proteotypic peptides related to eleven candidate T. pallidum protein biomarkers in eighteen plasma and four urine samples from individuals with syphilis. A number of explanations are possible. The foremost is the extremely low predicted concentration of bacterial proteins compared to host proteins. To a large extent our estimates of T. pallidum bacterial load in blood are based on molecular studies. In one of the largest studies, Tipple et al. found that median copy numbers of Lipoprotein antigen Tp47 (TP_0574) DNA detectable per milliliter of whole blood was 127, 516 and 70 in primary, secondary and latent syphilis, respectively13. Other studies have produced comparable results47,64,65, with the exception of a recent study that found a median of 1.4 × 105 T. pallidum/ml in whole blood from patients with secondary syphilis66.
The concentration of T. pallidum in blood according to these PCR-based studies is lower compared to our estimated LOD in a shotgun experiment on diluted samples (300 T. pallidum/ml) since we would need a 500x higher concentration (same amount of proteins from 300 T. pallidum in 1 ml vs. 2 µl) to detect the 300 T. pallidum/ml (see Supplementary File 4). Despite this outcome, we were hoping to detect T. pallidum proteins in the plasma or urine of some syphilis patients because i) MRM measurements are generally more sensitive than shotgun experiments since scanning times are drastically reduced and ii) the amounts from Tipple et al.13 were averages so we hypothesized that some patients (especially those with secondary syphilis) might have high T. pallidum levels detectable by MRM. These results could then motivate us to develop an (immuno)assay, capable of detecting the proteins even at low concentrations.
Little difference in T. pallidum abundance has been found between whole blood, plasma or serum12. Not much is known about the persistence of T. pallidum in the human urinary tract and to our knowledge no studies have quantified T. pallidum in the urine of syphilis-infected patients. However, even if T. pallidum does not consistently persist in the urinary tract, bacterial proteins present in the blood could be filtered through the glomerulus, ending up in the urine either intact or as peptide fragments, depending on the size of the protein and state of proteolysis67.
These considerations suggest that detection of T. pallidum proteins in human biofluids may not be possible without additional steps such as front-end immunoaffinity depletion68, two-dimensional LC separation69 and/or selective enrichment of target proteins/peptides (as reviewed by Shi et al.70). These techniques, or combinations thereof, have allowed the detection of low abundance proteins up to the low- to sub-nanogram/ml level70,71 in clinical samples. For example, to reduce the wide dynamic range of plasma proteins, multicomponent single-step immunoaffinity depletion of high-abundant (host) proteins can allow up to a 10-20-fold enrichment of low-abundant proteins due to the depletion of 90–95 % of the total protein mass68. However, of particular concern with this approach is the possibility of concomitant removal of low-abundance proteins due to protein binding to the antibodies or high-abundant proteins, as shown in a study that systematically analysed the antibody bound (high-abundant) protein fraction which found that this fraction contained 101 proteins at a high degree of confidence72. T. pallidum has a high binding affinity for constituents of serum and host cells, including laminin73, fibronectin74,75 and albumin76, which may lead to unintentional depletion of targeted proteins if human protein specific immunodepletion would be applied. Furthermore, targeted mass spectrometric immunoassays (MSIA) that use surface-immobilized antibodies to affinity retrieve proteins from biological samples have proven their utility for clinical applications77–79. In our study, magnet bead coupled polyclonal anti- T. pallidum antibodies failed to significantly detect more T. pallidum proteins compared to the unenriched dilution series. Antibody effectivity is dictated by binding affinity; we used commercial antibodies that were to our knowledge not previously characterized as to their binding affinity or targeted proteins. Furthermore, it is unlikely that the polyclonal antibodies would bind a large range of proteins since few (<5 %) T. pallidum proteins are immunogenic16,80. The fact that T. pallidum can remain in ‘plain sight’ without invoking immune defences81, together with the very low amount of outer membrane proteins compared to other human pathogens82, also suggests that antibody enrichment of whole organisms and/or proteins would probably not be an effective strategy. Peptide-level immunoenrichment, also known as the ‘Stable Isotope Standards and Capture by Anti-Peptide Antibodies’ (SISCAPA) method developed by Anderson et al.83 has shown considerable promise as a high-throughput, automated, highly multiplexed approach for protein biomarker quantification, with MRM application detection limits in the low picogram/ml range of protein concentration in plasma84. If a selection of T. pallidum peptides could be definitively demonstrated to be present in plasma or urine, then this could be an attractive analytical approach with a strong potential for yielding the detection capabilities and precision needed for clinical applications.
However, apart from the low abundance in plasma or urine, other factors could explain why the T. pallidum proteins were not detected in our MRM experiments:
1. The LOD T. pallidum spiking experiments were performed in PBS buffer as opposed to a highly complex plasma or urine matrix background.
2. Variations in gene expression and structural components of proteins could also account for the lack of T. pallidum protein detection. Fluctuations in gene expression may explain why we did not find TprG, a protein implicated in phase variation which has been shown to be expressed at varying levels during infection due to changes in the number of guanine nucleotide repeats immediately upstream of its transcriptional start site85. Heterogeneous T. pallidum protein sequence sites15,17,86 could also confound rigid MRM assay detection parameters. Such heterogeneity has been shown17 to be present in one candidate biomarker, TP_0922, although this variable site was not present in the PTPs incorporated in this MRM assay. Poor proteolytic cleavage can stem from structural features of the protein, different digestion kinetics and post-translational modifications. For example, phosphorylated residues within two amino acids of the point of cleavage can hinder proteolysis87. Little is known about the extent of T. pallidum protein post-translational modification aside from a study that demonstrated glycosylation of the Flagellar core proteins (FlaBs) as reported by antibody and glycan staining techniques88, however, the exact modification sites and extent of modification remain unknown. Other proteomics studies of L. interrogans have demonstrated likely roles for protein acetylation and methylation in virulence mechanisms89,90.
3. We only tested eleven out of more than a thousand predicted proteins in the T. pallidum proteome57, a selection largely based on spectral counting17 as an estimation of protein abundance. We cannot assume, however, that this indirect manner of quantifying T. pallidum protein levels in a rabbit testicle model directly recapitulates T. pallidum protein expression levels in plasma samples of syphilis-infected patients. One of the reasons for this is that protein expression may vary according to host and disease stage. Antigen detection during latent stage disease will be especially challenging since T. pallidum has been shown to sequester itself in protected niches such as eyes, hair follicles and nerves91. Other T. pallidum proteins may be more suitable diagnostic biomarkers, given that they are reflective of the disease stages studied and that they are consistently present in the biofluids of interest. For example, Lipoprotein Tp47, which could still be identified in the most diluted T. pallidum sample (300 T. pallidum/ml) in this study, could be an interesting biomarker for future studies.
4. Various technical limitations such as a possible suboptimal chromatographic gradient length, modifiable proteotypic residues and protein degradation secondary to sample processing could have impeded biomarker detection. Other studies have reported chromatographic gradient lengths of 30 minutes or longer33,34,36,39, thus implementation of longer gradients could be considered in future studies in order to improve peptide resolution. In this study, chromatographic separations were performed in triple using shorter 10-minute gradients in order to optimize the sample throughput without the loss of MS sensitivity due to overlapping transition windows. Therefore, co-eluting peptides were split over different chromatographic runs since plasma protein availability was not a limiting factor. Oxidizable proteotypic residues, namely cysteine, methionine and tryptophan, can cause artifactual modifications during processing or storage resulting multiple forms of targeted peptides. With this said, the PTP selection process also requires a necessary balance between many different parameters, whereby selection of peptides containing suboptimal amino acid residues can sometimes remain the most favourable option. Ribosomal protein TP_0250b was only represented by one PTP, which may have limited detectability, thus future assays could ideally incorporate more than one peptide per protein.
5. Sample processing may have also contributed to protein degradation; therefore prompt analysis of fresh non-frozen biological specimens, if possible, is recommended. Moreover, alternative sample processing procedures, such as the use of molecular weight cut off filters to concentrate urine could improve protein detectability40.
6. Lastly, only a limited amount of clinical samples were analysed, especially urine and the study was a single centre study with only MSM participants, therefore it is not generalizable. An improvement for future studies would be the incorporation of isotopically labelled (non-T. pallidum) reference standards, which have been shown to improve analytical precision, detect variations in instrument performance and aid in detecting chemical interferences92.
Targeted MS approaches are only able to search for a limited amount of pre-selected biomarker candidates. A more comprehensive approach would be to take a step backwards to conduct broader shotgun proteomics in plasma and urine samples of individuals with syphilis. Shotgun approaches identifying M. tuberculosis antigens in urine have been previously successful40,41. A compelling study from Eyford et al. used a ‘deep-mining’ proteomics approach and were able to detect 254 Typanosoma brucei rhodesiense proteins in plasma from African sleeping sickness patients93. Quantitative data- independent acquisition modes of MS analysis, including SWATH-MS94, are also very promising avenues for clinical applications95,96.
In an effort to identify promising T. pallidum diagnostic biomarkers, we designed a scheduled MRM assay incorporating 141 MRM ions pairs correlated to 30 PTPs/ 11 T. pallidum proteins. Factors such as the extremely low (femtomoles per liter) predicted T. pallidum protein concentration in biofluids, possible variable protein expression according to host/disease stage and potential presence of protein post-translational modifications likely contributed to the lack of signal detection for all candidate biomarkers investigated. Since the proteins targeted in this study were likely buried in the proverbial haystack of plasma proteins, alternative sample preparation and analysis strategies are warranted. With the rapidly progressing innovations of MS applications and technology, we believe clinical proteomics is far from its pinnacle of potential.
The datasets supporting the conclusions of this article are available in the PeptideAtlas54 repository, with the identifier PASS00978, in addition to being provided within the article and its supplementary files.
The prospective observational cohort study (SeTPAT ClinicalTrials.gov # NCT02059525) that provided the clinical samples used in this study was approved by the Institutional Review Board of the Institute of Tropical Medicine Antwerp and the Ethics Committee of the University of Antwerp (13/44/426), Belgium. Written informed consent for publication of the participants’ anonymized details was obtained from the participants. The T. pallidum ssp. pallidum DAL-1 strain used in this study was propagated in rabbits at the Veterinary Research Institute in Brno, Czech Republic. The handling of animals in the study was performed in accordance with the current Czech legislation (Animal Protection and Welfare Act No. 246/1992 Coll. of the Government of the Czech Republic). These specific experiments were approved by the Ethics Committee of the Veterinary Research Institute (Permit Number 20– 2014).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Comments on this article Comments (0)