Hanemaaijer MJ, Houston PD, Collier TC et al. Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.12688/f1000research.13807.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
1Vector Genetics Laboratory, Department of Pathology, Microbiology and Immunology, University of California Davis, Davis, CA, 95616, USA 2Malaria Research and Training Center, University of Bamako, Bamako, E2528, Mali 3Mosquito Control Research Laboratory, Kearney Agricultural Center, Department of Entomology and Nematology, University of California Davis, Davis, CA, 93648, USA
Mark J. Hanemaaijer
Roles:
Conceptualization,
Formal Analysis,
Writing – Original Draft Preparation,
Writing – Review & Editing
Parker D. Houston
Roles:
Writing – Original Draft Preparation
Travis C. Collier
Roles:
Data Curation,
Software
Laura C. Norris
Roles:
Investigation
Abdrahamane Fofana
Roles:
Investigation
Gregory C. Lanzaro
Roles:
Writing – Review & Editing
Anthony J. Cornel
Roles:
Investigation,
Writing – Review & Editing
Yoosook Lee
Roles:
Conceptualization,
Data Curation,
Formal Analysis,
Writing – Original Draft Preparation,
Writing – Review & Editing
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
Here we report the complete mitochondrial sequences of 70 individual field collected mosquito specimens from throughout Sub-Saharan Africa. We generated this dataset to identify species specific markers for the following Anopheles species and chromosomal forms: An. arabiensis, An. coluzzii (The Forest and Mopti chromosomal forms) and An. gambiae (The Bamako and Savannah chromosomal forms). The raw Illumina sequencing reads were mapped to the NC_002084 reference mitogenome sequence. A total of 783 single nucleotide polymorphisms (SNPs) were detected on the mitochondrial genome, of which 460 are singletons (58.7%). None of these SNPs are suitable as molecular markers to distinguish among An. arabiensis, An. coluzzii and An. gambiae or any of the chromosomal forms. The lack of species or chromosomal form specific markers is also reflected in the constructed phylogenetic tree, which shows no clear division among the operational taxonomic units considered here.
Keywords
Mitogenome, species identification, Africa, malaria vector, mosquitoes, Anopheles, single nucleotide polymorphisms, phylogenomics
Corresponding author:
Yoosook Lee
Competing interests:
No competing interests were disclosed.
Grant information:
We thank University of California - Irvine, Malaria Initiatives (UCIMI) for their support.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Historically, mtDNA sequence has been used in taxonomy as a source of species diagnostic markers (Cronin et al. (1991); De Barba et al. (2014); Pegg et al. (2006)) or in population genetics and evolutionary studies (Fu et al. (2013); Harrison (1989); Llamas et al. (2016)). One advantage of using mitochondrial over nuclear DNA for such studies is that the mutation rate of mtDNA is about 10 times faster than nuclear DNA (Brown et al. (1979); Haag-Liautard et al. (2008)), hence amplifying the evolutionary trajectory of populations and species. In addition, mtDNA is easy to amplify, because there are more copies of mitochondrial DNA relative to nuclear DNA. Also, universal primers can be applied to a wide range of species. Widely used universal primers target the cytochrome b and cytochrome oxidase 1 genes (Tahir et al. (2016)), because both have conserved and highly variable regions. In addition to these, other genes as described in De Mandal et al. (2014), can also be used as markers. However, phylogenetic trees based on mtDNA can deviate from the ones that are derived from nuclear DNA (Phillips et al. (2013); Shaw (2002); Sota & Vogler, 2001).
The Anopheles gambiae species complex consists of eight morphologically identical species that can only be distinguished with molecular markers (Scott et al. (1993)) or, for some of the species, by cytological examination of polytene chromosomes (Green, 1972; Pombi et al., 2008). The currently used molecular markers are located within genomic islands of divergence located proximal to the centromeres (Lee et al. (2014); Turner et al. (2005)). Monitoring additional species-specific markers on mitochondrial DNA (mtDNA) could increase the ease of application and accuracy of species detection assays. In addition, mtDNA markers could enhance our understanding of divergence times among taxa within the complex.
In this study we wished to identify species-specific markers within the mtDNA for Anopheles arabiensis, An. coluzzii and An. gambiae, including among the chromosomal forms currently subsumed under the designations An. gambiae and An. coluzzii, with the goal of adding these to our existing Anopheles species detection assay (Lee et al. (2014)). We sequenced the whole mitogenomes of 70 individual mosquito specimens collected throughout Sub-Saharan Africa. The raw Illumina sequencing reads were mapped to the AgamP4 reference sequence, which included both nuclear and mitochondrial sequences. We explore the relationship among An. arabiensis, An. coluzzii, An. gambiae and four of the sub-specific chromosomal form mitogenome sequences.
Methods
Sample collection
Anopheles arabiensis raw Illumina sequencing reads were obtained from our previous study (Marsden et al. (2014)). These included 20 samples from three villages in Tanzania collected in 2012 (Lupiro ((-8.38000°N, 36.66912°W), Sagamaganga (-8.06781°N, 36.80207°W), and Minepa (-8.25700°N, 36.68163°W) in the Kilombero Valley) and 4 samples from Cameroon collected in 2005 (9.09957°N, 13.72292°W). The An. gambiae and An. coluzzii samples were collected as resting adults using mouth aspirators in Kela, Mali (11.88683°N, -8.44744°W) in 2012 and Mutengene, Cameroon (4.0994°N, 9.3081°W) in 2011. We subdivided the An. coluzzii specimen into the Forest and Mopti chromosomal forms. Similarly, we did this for the An. gambiae Savannah and Bamako chromosomal forms. We used the same definitions and methods to characterize the chromosomal forms as in Lanzaro & Lee, 2013.
Genome sequencing
Sequencing methods for An. arabiensis samples are as described in Marsden et al. (2014). In short, individually barcoded Illumina paired-end sequencing libraries, with insert sizes of 320-400 basepairs (bp) using NEXTflex Sequencing kits (NOVA-5144) and barcodes (NOVA-514102)(Bio Scientific, Austin, TX, USA), were sequenced on an Illumina HiSeq2000 (Illumina, San Diego, CA, USA) with 100-bp paired-end reads using twelve samples per lane. For the An. coluzzii and An. gambiae samples we used the same methods as described in Norris et al. (2015) and Main et al. (2015). For the latter species, libraries were created using the Nextera DNA Sample Preparation Kit (FC-121-1031) and TruSeq dual indexing barcodes (FC-121-103)(Illumina) and the samples were sequenced on an Illumina HiSeq2500 with 100-bp paired end reads.
Data analysis
De-multiplexed raw reads were trimmed using Trimmomatic (Bolger et al. (2014)) version 0.36 and mapped to the mitogenome reference sequence of An. gambiae (Genbank accession number = NC_002084 (Beard et al. (1993))). Freebayes (v1.0.1) (Garrison & Marth, 2013) was used for mitochondrial variant calling assuming single ploidy and without population prior. Mapping statistics were calculated using qualimap version 2.2 (Okonechnikov et al. (2016)) and the data is represented in Table 1. Following the recommendation of Crawford and Lazarro (Crawford & Lazzaro, 2012), we used a minimum depth of 8 to call variants for each individual. Between positions 1-13,470bp of the mitogenome, we obtained consistently high quality reads for all samples, which were used for further analysis. An AT-rich region located between 13,471 and 15,388 suffers from low or zero coverage for sequences generated with the Nextera library preparation kit. Therefore, we excluded these regions from further analysis. The Vcf2fasta program (Danecek et al. (2011)) was used to extract mitogenome sequences from vcf file to fasta format. Geneious version 10.1.3 was used for mitogenome alignments. The phylogenetic tree was generated using the Jukes-Cantor genetic distance model and Neighbor-Joining tree methods available in Geneious version 10.1.3. We used scikit-allel (v1.1.9), a software package for Python (Miles & Harding (2017)), to identify species specific markers.
Table 1. List of samples that are used for the study.
Mapped reads indicates the reads that are mapped to the reference genome. Mean coverage indicates the average depth of reads on the mitochondrial DNA and standard deviation indicates the coverage deviation across the mitochondrial DNA.
Species
Banked_id
Year
Country
Village
Mapped bases
Mean coverage
Standard deviation
An. coluzzii-Forest
11MUTE470
2011
Cameroon
Mutengene
4265836
277.7
144.5
An. coluzzii-Forest
11MUTE472
2011
Cameroon
Mutengene
1862892
121.3
23
An. coluzzii-Forest
11MUTE476
2011
Cameroon
Mutengene
2130531
138.7
50.5
An. coluzzii-Forest
11MUTE477
2011
Cameroon
Mutengene
806611
52.5
16.7
An. coluzzii-Forest
11MUTE480
2011
Cameroon
Mutengene
804015
52.3
21
An. coluzzii-Forest
11MUTE483
2011
Cameroon
Mutengene
1702247
110.8
42.9
An. coluzzii-Forest
11MUTE487
2011
Cameroon
Mutengene
812839
52.9
21.2
An. coluzzii-Forest
11MUTE490
2011
Cameroon
Mutengene
1882088
122.5
52.4
An. coluzzii-Forest
11MUTE491
2011
Cameroon
Mutengene
1422997
92.6
46.6
An. coluzzii-Forest
11MUTE493
2011
Cameroon
Mutengene
627590
40.9
17.3
An. coluzzii-Mopti
12KELA022
2012
Mali
Kela
3695920
240.6
64.4
An. coluzzii-Mopti
12KELA024
2012
Mali
Kela
574282
37.4
30.8
An. coluzzii-Mopti
12KELA046
2012
Mali
Kela
4152520
270.3
87.2
An. coluzzii-Mopti
12KELA085
2012
Mali
Kela
10883282
708.4
345
An. coluzzii-Mopti
12KELA087
2012
Mali
Kela
3351158
218.1
79.8
An. coluzzii-Mopti
12KELA088
2012
Mali
Kela
1704283
110.9
91.3
An. coluzzii-Mopti
12KELA099
2012
Mali
Kela
349531
22.8
11
An. coluzzii-Mopti
12KELA112
2012
Mali
Kela
8550102
556.5
198.2
An. coluzzii-Mopti
12KELA161
2012
Mali
Kela
33794208
2199.7
629.3
An. gambiae-Savannah
12KELA210
2012
Mali
Kela
3007375
195.8
53.3
An. gambiae-Bamako
12KELA214
2012
Mali
Kela
26441050
1721.1
566.4
An. gambiae-Bamako
12KELA219
2012
Mali
Kela
3617355
235.5
130.2
An. gambiae-Savannah
12KELA228
2012
Mali
Kela
7783776
506.7
262.8
An. gambiae-Savannah
12KELA233
2012
Mali
Kela
7827363
509.5
138.6
An. gambiae-Savannah
12KELA234
2012
Mali
Kela
6721204
437.5
205.9
An. gambiae-Bamako
12KELA239
2012
Mali
Kela
6683521
435
126.4
An. gambiae-Bamako
12KELA240
2012
Mali
Kela
15131480
984.9
270.8
An. gambiae-Bamako
12KELA244
2012
Mali
Kela
12851754
836.5
306.5
An. gambiae-Savannah
12KELA285
2012
Mali
Kela
407888
26.6
119.8
An. gambiae-Savannah
12KELA321
2012
Mali
Kela
1034014
67.3
43.8
An. gambiae-Savannah
12KELA334
2012
Mali
Kela
20949015
1363.6
400.4
An. gambiae-Savannah
12KELA348
2012
Mali
Kela
12053890
784.6
280.9
An. gambiae-Bamako
12KELA367
2012
Mali
Kela
12109235
788.2
240.1
An. coluzzii-Mopti
12KELA400
2012
Mali
Kela
13707820
892.3
398.2
An. gambiae-Bamako
12KELA406
2012
Mali
Kela
17605437
1146
463.2
An. gambiae-Savannah
12KELA409
2012
Mali
Kela
10526480
685.2
259.1
An. coluzzii-Mopti
12KELA420
2012
Mali
Kela
31785953
2069
845.5
An. gambiae-Bamako
12KELA443
2012
Mali
Kela
25740781
1675.5
669.1
An. gambiae-Bamako
12KELA457
2012
Mali
Kela
1360654
88.6
36.6
An. coluzzii-Mopti
12KELA458
2012
Mali
Kela
153686
10
10.4
An. gambiae-Bamako
12KELA467
2012
Mali
Kela
10499093
683.4
249.1
An. gambiae-Savannah
12KELA468
2012
Mali
Kela
10315033
671.4
197.1
An. gambiae-Bamako
12KELA481
2012
Mali
Kela
20308589
1321.9
307.6
An. coluzzii-Mopti
12KELA496
2012
Mali
Kela
2975297
193.7
162.9
An. gambiae-Bamako
12KELA651
2012
Mali
Kela
376689
24.5
11.3
An. gambiae-Savannah
12KELA812
2012
Mali
Kela
799071
52
29.3
An. arabiensis
12LUPI001
2012
Tanzania
Lupiro
2843317
185.1
34.9
An. arabiensis
12LUPI007
2012
Tanzania
Lupiro
6288802
409.3
40
An. arabiensis
12LUPI024
2012
Tanzania
Lupiro
6328898
412
78.5
An. arabiensis
12LUPI056
2012
Tanzania
Lupiro
5440256
354.1
39.2
An. arabiensis
12LUPI059
2012
Tanzania
Lupiro
39721262
2585.5
801.8
An. arabiensis
12LUPI071
2012
Tanzania
Lupiro
3433158
223.5
59.2
An. arabiensis
12LUPI074
2012
Tanzania
Lupiro
10096062
657.2
100.5
An. arabiensis
12LUPI082
2012
Tanzania
Lupiro
5732773
373.2
69.6
An. arabiensis
12MINE001
2012
Tanzania
Minepa
7768923
505.7
66.9
An. arabiensis
12MINE040
2012
Tanzania
Minepa
2784428
181.2
54.9
An. arabiensis
12MINE100
2012
Tanzania
Minepa
10753877
700
93.9
An. arabiensis
12MINE101
2012
Tanzania
Minepa
5684230
370
41.9
An. arabiensis
12MINE105
2012
Tanzania
Minepa
1526829
99.4
32.8
An. arabiensis
12MINE111
2012
Tanzania
Minepa
5578562
363.1
76.3
An. arabiensis
12SAGA066
2012
Tanzania
Sagamaganga
12745079
829.6
142.3
An. arabiensis
12SAGA107
2012
Tanzania
Sagamaganga
14460217
941.2
259.2
An. arabiensis
12SAGA131
2012
Tanzania
Sagamaganga
15333239
998.1
282.9
An. arabiensis
12SAGA133
2012
Tanzania
Sagamaganga
3792945
246.9
62.5
An. arabiensis
12SAGA134
2012
Tanzania
Sagamaganga
2439101
158.8
34.5
An. arabiensis
12SAGA141
2012
Tanzania
Sagamaganga
3130504
203.8
33.3
An. arabiensis
05OKJ017
2005
Cameroon
Ourodoukoudje
9041052
588.5
78.8
An. arabiensis
05OKJ042
2005
Cameroon
Ourodoukoudje
148752684
9682.5
785.7
An. arabiensis
05OKJ045
2005
Cameroon
Ourodoukoudje
35514980
2311.7
262.8
An. arabiensis
05OKJ070
2005
Cameroon
Ourodoukoudje
22847478
1487.2
400.5
Dataset 1.Aligned FASTA file of mitogenome samples.
Results and Discussion
We identified a total of 783 single nucleotide polymorphisms (SNPs) over the entire mitogenome. The majority of these (58.7%) were singletons (found on one of the 70 mitogenomes). We did not identify any SNPs unique to the species or chromosomal forms (Table 2) and therefore conclude that mtDNA is not suitable for Anopheles gambiae complex species identification.
Table 2. List of SNP variants in the different Anopheles species and chromosomal forms.
Position
Reference nucleotide
Alternative nucleotides
SNP variant counts per group (first value is count of reference allel)
An. arabiensis
An. coluzzii- Forest
An. coluzzii- Mopti
An. gambiae- Bamako
An. gambiae- Savannah
55
C
T/-/-
21/3/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
71
C
T/-/-
0/24/0/0
0/11/0/0
0/13/0/0
1/11/0/0
0/11/0/0
117
A
G/-/-
23/1/0/0
9/2/0/0
13/0/0/0
12/0/0/0
11/0/0/0
126
T
C/-/-
11/13/0/0
1/10/0/0
0/13/0/0
0/12/0/0
1/10/0/0
205
C
T/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
223
T
C/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
248
G
A/-/-
24/0/0/0
11/0/0/0
12/0/0/0
12/0/0/0
10/1/0/0
250
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
298
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
340
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
373
A
G/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
391
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
396
C
T/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
409
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
441
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
469
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
488
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
502
A
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
503
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
514
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
520
T
A/C/-
22/0/2/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
526
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
10/1/0/0
562
A
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
572
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
10/2/0/0
11/0/0/0
583
A
G/-/-
24/0/0/0
11/0/0/0
10/3/0/0
12/0/0/0
11/0/0/0
611
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
665
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
685
A
G/-/-
24/0/0/0
11/0/0/0
12/0/0/0
12/0/0/0
10/1/0/0
688
T
A/-/-
24/0/0/0
10/1/0/0
12/0/0/0
11/1/0/0
11/0/0/0
700
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
706
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
712
A
C/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
757
A
G/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
763
A
G/-/-
11/13/0/0
3/8/0/0
0/13/0/0
2/10/0/0
1/10/0/0
787
A
G/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
788
T
C/-/-
24/0/0/0
9/1/0/0
12/1/0/0
12/0/0/0
10/1/0/0
796
A
G/-/-
22/2/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
811
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
821
T
C/-/-
11/13/0/0
1/9/0/0
1/12/0/0
0/12/0/0
1/10/0/0
832
A
G/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
887
T
C/-/-
16/8/0/0
10/0/0/0
12/0/0/0
12/0/0/0
10/1/0/0
908
A
G/-/-
18/6/0/0
10/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
934
G
A/-/-
22/2/0/0
9/1/0/0
12/0/0/0
12/0/0/0
10/1/0/0
949
A
G/-/-
24/0/0/0
10/0/0/0
11/2/0/0
11/1/0/0
11/0/0/0
955
T
C/-/-
12/12/0/0
5/5/0/0
9/4/0/0
4/8/0/0
2/9/0/0
1012
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1029
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1042
T
C/-/-
24/0/0/0
11/0/0/0
11/1/0/0
12/0/0/0
11/0/0/0
1045
A
G/-/-
24/0/0/0
11/0/0/0
11/1/0/0
12/0/0/0
11/0/0/0
1114
C
T/-/-
24/0/0/0
11/0/0/0
12/0/0/0
11/1/0/0
11/0/0/0
1150
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1153
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1155
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1163
T
C/-/-
0/24/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
1189
G
A/-/-
24/0/0/0
9/2/0/0
11/2/0/0
11/1/0/0
11/0/0/0
1198
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1308
T
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1465
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1474
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1486
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1504
A
G/-/-
23/1/0/0
9/2/0/0
13/0/0/0
11/1/0/0
10/1/0/0
1507
G
A/-/-
23/1/0/0
9/2/0/0
11/2/0/0
11/1/0/0
11/0/0/0
1523
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1538
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
1540
A
T/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
1543
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1546
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1549
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
1558
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1561
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
1567
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1618
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
1624
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1636
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1639
A
G/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1642
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1645
A
G/T/
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
10/0/1/0
1648
G
A/-/-
23/1/0/0
10/0/0/0
9/4/0/0
12/0/0/0
10/1/0/0
1681
A
C/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1693
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1729
G
A/-/-
23/1/0/0
7/4/0/0
13/0/0/0
10/2/0/0
11/0/0/0
1735
T
C/-/-
23/1/0/0
9/2/0/0
13/0/0/0
10/2/0/0
11/0/0/0
1738
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1751
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1765
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1780
G
A/-/-
23/1/0/0
10/1/0/0
11/2/0/0
12/0/0/0
11/0/0/0
1792
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1807
T
C/-/-
23/1/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1837
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/1/0/0
10/1/0/0
1876
A
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1879
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1903
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1906
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
1918
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1930
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
1933
G
A/-/-
23/1/0/0
10/0/0/0
11/2/0/0
7/5/0/0
7/4/0/0
1964
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1972
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
1978
G
A/-/-
22/2/0/0
11/0/0/0
9/4/0/0
8/4/0/0
11/0/0/0
2026
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2068
A
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2071
T
C/G/-
23/1/0/0
11/0/0/0
12/0/1/0
12/0/0/0
10/0/1/0
2080
T
C/-/-
22/2/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
2098
T
C/-/-
23/1/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2101
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2110
T
C/-/-
23/1/0/0
10/1/0/0
11/2/0/0
11/1/0/0
10/1/0/0
2116
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2149
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2164
T
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2179
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
2182
G
A/-/-
22/2/0/0
6/5/0/0
11/2/0/0
4/8/0/0
8/3/0/0
2206
A
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2209
G
A/-/-
19/5/0/0
10/1/0/0
12/1/0/0
11/1/0/0
9/2/0/0
2227
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2233
A
G/T/-
24/0/0/0
11/0/0/0
13/0/0/0
11/0/1/0
11/0/0/0
2239
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2260
C
T/-/-
2/22/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
2269
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2284
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2329
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2359
G
A/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
2374
G
A/-/-
23/1/0/0
11/0/0/0
10/3/0/0
11/1/0/0
10/1/0/0
2401
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2410
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2411
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2413
G
A/-/-
0/24/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
2426
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2432
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2434
G
A/-/-
22/2/0/0
9/2/0/0
11/2/0/0
11/1/0/0
10/1/0/0
2449
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2473
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2474
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2482
A
G/-/-
20/4/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2521
C
T/-/-
24/0/0/0
10/1/0/0
12/1/0/0
11/1/0/0
10/1/0/0
2530
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2558
T
C/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2569
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2599
A
G/T/-
23/1/0/0
10/1/0/0
13/0/0/0
11/0/1/0
11/0/0/0
2608
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2612
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2615
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2635
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2692
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2719
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2722
A
G/-/-
23/1/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2728
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2731
A
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2737
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2752
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2755
C
A/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
2758
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2774
G
A/-/-
23/1/0/0
11/0/0/0
11/2/0/0
11/1/0/0
11/0/0/0
2782
T
A/C/-
23/0/1/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
2791
T
A/C/-
23/0/1/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2797
C
T/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
2806
C
T/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
2818
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2848
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2851
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2859
C
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2872
T
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2902
C
T/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
2908
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
2938
G
A/-/-
23/1/0/0
10/1/0/0
13/0/0/0
11/1/0/0
10/1/0/0
2945
C
T/-/-
15/9/0/0
9/2/0/0
13/0/0/0
11/1/0/0
11/0/0/0
2975
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
3055
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
3124
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3136
T
C/-/-
24/0/0/0
9/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
3199
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3202
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3205
A
G/-/-
24/0/0/0
9/2/0/0
12/1/0/0
12/0/0/0
11/0/0/0
3238
T
A/C/
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3277
A
T/-/-
24/0/0/0
11/0/0/0
11/1/0/0
11/1/0/0
8/3/0/0
3286
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3304
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3307
A
C/G/T
24/0/0/0
11/0/0/0
9/0/1/3
12/0/0/0
11/0/0/0
3310
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
3325
G
A/-/-
24/0/0/0
11/0/0/0
11/2/0/0
10/2/0/0
11/0/0/0
3331
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
3334
T
C/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3337
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3352
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
3400
A
G/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3401
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3403
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
3412
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3413
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
3424
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3451
C
T/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
3457
G
A/T/
12/12/0/0
6/4/1/0
12/1/0/0
3/9/0/0
5/6/0/0
3458
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3466
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3508
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
11/1/0/0
10/1/0/0
3511
C
A/T/-
22/1/1/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3514
C
T/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
3523
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3544
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
3595
A
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3613
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
3619
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3652
G
A/-/-
22/2/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
3773
A
G/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3838
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
10/1/0/0
3870
C
T/-/-
12/12/0/0
3/8/0/0
6/7/0/0
1/11/0/0
3/8/0/0
3882
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3924
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
3960
T
C/-/-
24/0/0/0
10/0/0/0
10/3/0/0
11/1/0/0
11/0/0/0
3976
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4002
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
4011
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
4034
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4037
G
A/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
4052
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
4100
C
T/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4104
G
T/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
4112
C
T/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4115
A
G/-/-
11/13/0/0
1/9/0/0
0/13/0/0
0/12/0/0
2/9/0/0
4121
A
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4145
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4157
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4181
T
A/-/-
3/21/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
4190
A
G/-/-
0/24/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
4196
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4206
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4208
A
G/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
4214
T
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4220
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4256
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
4259
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4292
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4310
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
4332
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
4352
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4367
A
G/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4368
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4373
T
C/-/-
22/2/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
4430
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4442
G
A/-/-
14/10/0/0
10/1/0/0
11/2/0/0
12/0/0/0
9/2/0/0
4451
G
A/-/-
24/0/0/0
10/1/0/0
12/1/0/0
11/1/0/0
11/0/0/0
4484
A
T/-/-
20/4/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4487
T
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
4490
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
4494
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4526
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4529
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
4538
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4559
A
G/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
4562
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
4565
T
A/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
4566
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
4579
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4603
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4620
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4637
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4649
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
4652
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
4715
G
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
4747
C
A/T/-
24/0/0/0
10/0/1/0
13/0/0/0
10/1/1/0
11/0/0/0
4750
A
G/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
4756
C
T/-/-
21/3/0/0
9/2/0/0
13/0/0/0
9/3/0/0
10/1/0/0
4774
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4777
A
C/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
4795
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4846
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
4847
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4867
G
A/-/-
24/0/0/0
9/1/0/0
12/1/0/0
11/1/0/0
11/0/0/0
4885
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
4969
A
C/-/-
16/8/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5011
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5020
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5029
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5044
A
G/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5056
T
C/-/-
23/1/0/0
10/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
5059
A
T/-/-
11/13/0/0
1/9/0/0
0/13/0/0
0/12/0/0
1/10/0/0
5062
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5065
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5072
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5074
A
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5077
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5080
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5128
A
G/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5131
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5137
C
T/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5165
G
A/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5167
T
A/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5173
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5182
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5215
A
G/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5216
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5221
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/1/0/0
10/1/0/0
5254
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5257
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
5272
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
5278
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5279
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5287
A
G/-/-
19/5/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5308
A
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5314
C
T/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
5320
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5323
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5359
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/1/0/0
11/0/0/0
5398
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5428
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5431
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5513
A
G/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5540
C
T/-/-
23/1/0/0
7/4/0/0
7/5/0/0
7/5/0/0
9/2/0/0
5564
T
C/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
5572
A
G/-/-
21/3/0/0
10/1/0/0
11/1/0/0
11/1/0/0
11/0/0/0
5622
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5652
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5661
A
G/-/-
20/4/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5664
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5670
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5679
T
C/-/-
24/0/0/0
11/0/0/0
10/3/0/0
12/0/0/0
11/0/0/0
5682
G
A/-/-
23/1/0/0
7/4/0/0
5/8/0/0
6/6/0/0
11/0/0/0
5688
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
5691
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5700
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5712
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5850
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
5865
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5871
C
T/-/-
16/8/0/0
10/1/0/0
13/0/0/0
11/1/0/0
10/1/0/0
5883
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
5898
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
5941
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6021
A,T
A/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6054
G
A/-/-
21/3/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6222
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6252
A
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6277
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6289
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
10/1/0/0
6315
A
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6388
A
G/-/-
24/0/0/0
10/1/0/0
11/2/0/0
12/0/0/0
11/0/0/0
6421
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6425
A
G/-/-
9/15/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
6430
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6439
C
T/-/-
24/0/0/0
10/0/0/0
12/1/0/0
11/1/0/0
11/0/0/0
6442
T
C/G/-
14/9/1/0
10/0/0/0
12/1/0/0
12/0/0/0
8/3/0/0
6450
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6466
A
G/-/-
16/8/0/0
7/4/0/0
13/0/0/0
7/5/0/0
6/5/0/0
6469
G
A/-/-
21/3/0/0
9/2/0/0
11/2/0/0
12/0/0/0
10/1/0/0
6482
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6488
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6501
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6517
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6529
G
A/-/-
12/12/0/0
1/10/0/0
0/13/0/0
0/12/0/0
1/10/0/0
6551
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6556
G
A/-/-
9/15/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
6563
T
C/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
6571
G
A/-/-
23/1/0/0
9/2/0/0
13/0/0/0
11/1/0/0
10/1/0/0
6578
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6580
G
A/-/-
21/3/0/0
7/4/0/0
11/2/0/0
10/2/0/0
6/5/0/0
6616
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6619
G
A/-/-
21/3/0/0
9/2/0/0
13/0/0/0
11/1/0/0
7/4/0/0
6628
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/1/0/0
10/1/0/0
6631
T
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6637
G
A/-/-
23/1/0/0
9/2/0/0
11/2/0/0
10/2/0/0
11/0/0/0
6649
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6661
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6662
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6670
C
T/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6684
G
A/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6687
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6699
C
T/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6718
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
6726
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6730
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6731
C
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6733
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6753
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6764
C
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6791
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6793
A
G/-/-
24/0/0/0
11/0/0/0
10/3/0/0
11/1/0/0
11/0/0/0
6805
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
10/1/0/0
6808
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6829
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
6841
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6847
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6928
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
6933
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
6982
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
6985
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7027
C
T/-/-
24/0/0/0
9/2/0/0
13/0/0/0
10/2/0/0
10/1/0/0
7045
G
A/-/-
15/9/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
7075
T
C/-/-
24/0/0/0
8/2/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7090
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7099
T
C/G/-
24/0/0/0
11/0/0/0
13/0/0/0
9/2/1/0
11/0/0/0
7105
T
C/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
7108
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
10/2/0/0
10/1/0/0
7116
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
7135
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
7168
T
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7195
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
7207
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
7231
C
T/-/-
24/0/0/0
9/2/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7240
G
A/-/-
9/15/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
7255
G
A/-/-
3/21/0/0
1/10/0/0
0/13/0/0
1/11/0/0
1/10/0/0
7261
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7282
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7290
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7351
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
7360
T
C/-/-
11/13/0/0
2/9/0/0
4/9/0/0
1/11/0/0
1/10/0/0
7381
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
7396
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7402
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
7423
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7432
A
C/-/-
24/0/0/0
11/0/0/0
12/0/0/0
12/0/0/0
10/1/0/0
7450
G
A/-/-
23/1/0/0
10/1/0/0
12/1/0/0
12/0/0/0
9/2/0/0
7478
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7479
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7483
C
G/T/
24/0/0/0
10/1/0/0
11/0/2/0
11/1/0/0
11/0/0/0
7486
C
T/-/-
11/13/0/0
5/6/0/0
4/9/0/0
0/12/0/0
2/9/0/0
7525
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7555
A
G/-/-
24/0/0/0
9/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7578
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7588
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7603
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7606
G
A/-/-
24/0/0/0
10/1/0/0
11/2/0/0
12/0/0/0
11/0/0/0
7627
A
G/-/-
13/11/0/0
7/4/0/0
12/1/0/0
5/7/0/0
5/6/0/0
7672
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7687
G
A/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
7693
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
7735
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/1/0/0
11/0/0/0
7777
C
T/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
7797
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7798
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
7822
A
G/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
7861
T
C/-/-
23/1/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
7923
T
C/-/-
24/0/0/0
11/0/0/0
10/3/0/0
11/1/0/0
11/0/0/0
7952
A
G/-/-
0/24/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
8001
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8034
C
T/-/-
13/11/0/0
6/4/0/0
11/1/0/0
3/9/0/0
5/6/0/0
8091
A
AT/-/-
22/2/0/0
9/1/0/0
12/1/0/0
12/0/0/0
9/2/0/0
8122
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8123
C
T/-/-
22/2/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8149
G
A/-/-
2/22/0/0
0/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
8196
G
A/-/-
23/1/0/0
10/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
8201
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8261
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8297
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8302
G
A/-/-
23/1/0/0
10/0/0/0
10/3/0/0
12/0/0/0
11/0/0/0
8303
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8309
C
T/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8339
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8342
C
T/-/-
24/0/0/0
11/0/0/0
10/3/0/0
12/0/0/0
11/0/0/0
8372
C
T/-/-
24/0/0/0
9/2/0/0
11/2/0/0
11/1/0/0
11/0/0/0
8375
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8382
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
8384
A
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8393
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
8399
G
A/-/-
2/22/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
8404
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8414
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8425
A
G/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8450
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8465
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8471
G
A/T/-
24/0/0/0
6/5/0/0
7/6/0/0
10/2/0/0
10/1/0/0
8513
A
G/-/-
22/2/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8528
A
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8546
A
T/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8564
C
A/T/-
23/0/1/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8582
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
8588
T
C/-/-
24/0/0/0
11/0/0/0
9/4/0/0
11/1/0/0
10/1/0/0
8612
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8645
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8657
T
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
8663
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8678
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8684
T
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
10/2/0/0
11/0/0/0
8714
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
8720
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
10/2/0/0
11/0/0/0
8722
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8725
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8726
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
8740
G
A/-/-
24/0/0/0
10/1/0/0
9/4/0/0
9/3/0/0
10/1/0/0
8744
G
A/-/-
23/1/0/0
8/3/0/0
11/2/0/0
10/2/0/0
10/1/0/0
8765
G
A/-/-
22/2/0/0
10/1/0/0
12/1/0/0
11/1/0/0
8/3/0/0
8807
T
C/-/-
22/2/0/0
9/2/0/0
12/1/0/0
11/1/0/0
11/0/0/0
8819
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
11/1/0/0
8/3/0/0
8840
C
T/-/-
11/13/0/0
1/10/0/0
0/13/0/0
0/12/0/0
1/10/0/0
8842
A
G/-/-
24/0/0/0
10/1/0/0
10/3/0/0
12/0/0/0
9/2/0/0
8855
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
8882
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
9/2/0/0
8893
G
A/-/-
24/0/0/0
7/4/0/0
8/6/0/0
9/3/0/0
11/0/0/0
8903
C
T/-/-
23/1/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
8913
A
C/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
8915
G
A/-/-
23/1/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
8927
A
G/-/-
11/13/0/0
2/9/0/0
0/14/0/0
1/11/0/0
2/9/0/0
8942
A
G/-/-
24/0/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
8995
A
G/-/-
21/3/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
8996
A
T/-/-
24/0/0/0
11/0/0/0
13/1/0/0
12/0/0/0
11/0/0/0
9014
A
T/-/-
24/0/0/0
11/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
9035
C
T/-/-
23/1/0/0
11/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
9041
C
T/-/-
3/21/0/0
0/11/0/0
0/14/0/0
0/12/0/0
0/11/0/0
9062
C
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9122
T
C/-/-
24/0/0/0
9/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
9152
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
9158
C
T/-/-
23/1/0/0
7/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
9210
G
A/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9236
C
T/-/-
24/0/0/0
9/2/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9244
G
A/-/-
22/2/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9245
C
A/T/-
23/0/1/0
10/0/1/0
13/0/0/0
12/0/0/0
9/1/1/0
9278
G
A/-/-
24/0/0/0
10/1/0/0
12/1/0/0
11/1/0/0
11/0/0/0
9287
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9311
G
A/-/-
9/15/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
9353
C
T/-/-
24/0/0/0
11/0/0/0
11/1/0/0
12/0/0/0
11/0/0/0
9359
G
A/-/-
9/15/0/0
1/10/0/0
0/13/0/0
0/12/0/0
0/11/0/0
9365
T
C/-/-
24/0/0/0
9/2/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9421
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
9422
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
9459
A
G/-/-
24/0/0/0
10/0/0/0
10/3/0/0
11/1/0/0
11/0/0/0
9493
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
9529
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9532
C
A/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9535
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9550
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9575
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9583
A
G/-/-
20/4/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9611
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9631
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
9660
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9688
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
9690
T
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9711
T
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9724
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
9795
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
9808
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9911
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
9930
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
9976
C
T/-/-
24/0/0/0
10/1/0/0
14/0/0/0
12/0/0/0
11/0/0/0
10011
A
G/-/-
24/0/0/0
11/0/0/0
13/1/0/0
12/0/0/0
10/1/0/0
10047
A
G/-/-
24/0/0/0
11/0/0/0
13/1/0/0
12/0/0/0
11/0/0/0
10063
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10068
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10104
A
G/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
10156
G
A/C/-
24/0/0/0
8/1/2/0
13/1/0/0
10/1/1/0
8/3/0/0
10168
C
A/T/-
24/0/0/0
7/4/0/0
9/5/0/0
9/2/1/0
10/0/1/0
10176
A
G/-/-
11/13/0/0
2/9/0/0
3/11/0/0
0/12/0/0
2/9/0/0
10177
G
A/-/-
24/0/0/0
11/0/0/0
13/1/0/0
12/0/0/0
10/1/0/0
10185
C
A/T/-
16/0/8/0
11/0/0/0
13/1/0/0
12/0/0/0
10/0/1/0
10187
T
C/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
10252
A
G/-/-
24/0/0/0
10/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
10253
C
T/-/-
23/1/0/0
10/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
10269
C
T/-/-
24/0/0/0
7/3/0/0
9/5/0/0
10/2/0/0
11/0/0/0
10284
A
G/-/-
24/0/0/0
10/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
10311
A
G/-/-
24/0/0/0
10/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
10324
T
C/-/-
24/0/0/0
10/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
10332
A
G/-/-
24/0/0/0
10/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
10341
T
C/-/-
24/0/0/0
10/0/0/0
10/3/0/0
12/0/0/0
11/0/0/0
10371
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10383
A
G/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10396
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10401
G
A/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10424
T
A/-/-
16/8/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
10427
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10454
C
T/-/-
24/0/0/0
10/0/0/0
12/1/0/0
10/2/0/0
10/1/0/0
10463
T
C/-/-
24/0/0/0
7/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
10466
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
10481
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10482
G
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10499
C
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10502
C
T/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10535
C
T/-/-
23/1/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10536
T
C/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10542
C
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
9/2/0/0
10559
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10566
C
T/-/-
23/1/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10568
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10583
C
T/-/-
23/1/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10598
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10607
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10628
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10640
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10664
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10670
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
10/1/0/0
10691
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10700
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10715
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10718
A
G/-/-
24/0/0/0
9/2/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10719
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10724
T
C/-/-
22/2/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10727
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10737
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10742
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10763
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10784
A
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
10796
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
10/1/0/0
10805
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
10808
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10820
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
10841
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10863
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10872
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10877
T
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10880
C
T/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
10881
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10886
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
10889
G
A/-/-
22/2/0/0
9/2/0/0
11/2/0/0
9/3/0/0
11/0/0/0
10892
T
C/-/-
23/1/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10898
T
A/C/-
11/0/13/0
2/0/9/0
0/0/13/0
1/0/11/0
1/0/10/0
10910
A
G/-/-
20/4/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10913
G
A/-/-
22/2/0/0
9/2/0/0
10/3/0/0
10/2/0/0
8/3/0/0
10916
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
10/2/0/0
11/0/0/0
10931
C
T/-/-
24/0/0/0
8/3/0/0
8/5/0/0
10/2/0/0
11/0/0/0
10934
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
10955
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
10988
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
10997
G
A/-/-
23/1/0/0
9/1/0/0
11/2/0/0
10/2/0/0
10/1/0/0
10998
A
G/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11042
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11060
A
G/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
11093
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11117
A
G/-/-
24/0/0/0
10/0/0/0
11/2/0/0
12/0/0/0
11/0/0/0
11150
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11159
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11160
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11171
T
C/-/-
23/1/0/0
9/2/0/0
11/2/0/0
11/1/0/0
8/3/0/0
11174
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11204
T
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11213
A
G/T/-
24/0/0/0
10/1/0/0
12/1/1/0
12/0/0/0
10/0/1/0
11216
T
C/-/-
24/0/0/0
11/0/0/0
13/1/0/0
10/2/0/0
10/1/0/0
11222
A
G/-/-
24/0/0/0
8/3/0/0
9/5/0/0
10/2/0/0
11/0/0/0
11234
C
T/-/-
24/0/0/0
11/0/0/0
12/2/0/0
11/1/0/0
11/0/0/0
11249
C
T/-/-
24/0/0/0
10/1/0/0
13/1/0/0
12/0/0/0
9/2/0/0
11276
G
A/-/-
23/1/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
11282
A
G/-/-
24/0/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
11301
C
T/-/-
9/15/0/0
2/9/0/0
0/14/0/0
0/12/0/0
0/11/0/0
11318
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11339
C
T/-/-
24/0/0/0
8/3/0/0
7/6/0/0
10/2/0/0
11/0/0/0
11348
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11351
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
10/2/0/0
10/1/0/0
11354
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11363
A
G/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11390
C
T/-/-
24/0/0/0
10/1/0/0
14/0/0/0
12/0/0/0
9/2/0/0
11414
G
A/-/-
23/1/0/0
10/1/0/0
10/4/0/0
10/2/0/0
11/0/0/0
11415
T
C/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
11432
A
G/-/-
23/1/0/0
11/0/0/0
14/0/0/0
10/2/0/0
10/1/0/0
11450
C
T/-/-
13/11/0/0
1/10/0/0
1/13/0/0
0/12/0/0
2/9/0/0
11475
T
C/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
11492
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11537
G
A/-/-
24/0/0/0
9/2/0/0
8/5/0/0
10/2/0/0
8/3/0/0
11538
C
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11592
G
A/-/-
0/24/0/0
0/11/0/0
0/13/0/0
0/12/0/0
0/11/0/0
11599
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11610
A
AT/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
11655
G
A/-/-
8/16/0/0
1/9/0/0
0/13/0/0
0/12/0/0
0/11/0/0
11673
G
A/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
11729
G
A/-/-
24/0/0/0
9/2/0/0
11/2/0/0
12/0/0/0
8/3/0/0
11739
A
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11745
T
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11748
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11757
C
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11760
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11763
A
G/-/-
18/6/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
11787
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11793
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11829
A
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11862
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11868
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
11899
G
A/-/-
24/0/0/0
9/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
11907
C
T/-/-
20/4/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
11910
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11916
G
A/-/-
20/4/0/0
5/5/0/0
2/11/0/0
5/7/0/0
8/3/0/0
11925
A
G/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11931
T
C/-/-
23/1/0/0
9/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
11982
A
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
11988
A
C/-/-
12/12/0/0
6/4/0/0
11/2/0/0
3/9/0/0
3/8/0/0
12005
G
A/-/-
23/1/0/0
9/1/0/0
12/1/0/0
9/3/0/0
8/3/0/0
12006
C
T/-/-
23/1/0/0
10/0/0/0
13/0/0/0
11/1/0/0
10/1/0/0
12009
G
A/T/-
24/0/0/0
6/4/0/0
8/2/3/0
8/4/0/0
11/0/0/0
12015
A
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
12020
A
G/-/-
24/0/0/0
9/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
12036
A
G/-/-
13/11/0/0
2/8/0/0
1/12/0/0
2/10/0/0
4/7/0/0
12039
A
G/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12061
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12078
A
G/-/-
24/0/0/0
9/2/0/0
13/0/0/0
11/1/0/0
11/0/0/0
12084
A
G/-/-
24/0/0/0
11/0/0/0
10/3/0/0
11/1/0/0
11/0/0/0
12092
C
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12159
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
12165
C
A/T/-
19/4/1/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12168
C
T/-/-
22/2/0/0
10/1/0/0
12/1/0/0
12/0/0/0
9/2/0/0
12171
C
T/-/-
24/0/0/0
9/2/0/0
13/0/0/0
11/1/0/0
10/1/0/0
12204
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
12210
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
12225
T
C/-/-
24/0/0/0
11/0/0/0
11/2/0/0
12/0/0/0
10/1/0/0
12228
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
12258
A
G/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
12273
A
G/-/-
23/1/0/0
11/0/0/0
14/0/0/0
12/0/0/0
11/0/0/0
12278
A
G/-/-
24/0/0/0
11/0/0/0
14/0/0/0
11/1/0/0
11/0/0/0
12333
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
9/2/0/0
12336
A
T/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
12339
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12348
A
G/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
12420
T
C/-/-
24/0/0/0
11/0/0/0
13/1/0/0
12/0/0/0
11/0/0/0
12471
G
A/-/-
23/1/0/0
11/0/0/0
14/0/0/0
10/2/0/0
10/1/0/0
12480
A
G/-/-
24/0/0/0
10/1/0/0
14/0/0/0
12/0/0/0
11/0/0/0
12499
G
A/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12501
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12536
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
12558
T
C/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12601
T
TTA/-/-
24/0/0/0
10/0/0/0
14/0/0/0
12/0/0/0
10/1/0/0
12607
A
T/-/-
24/0/0/0
10/0/0/0
11/3/0/0
12/0/0/0
11/0/0/0
12633
G
A/-/-
23/1/0/0
9/1/0/0
12/2/0/0
11/1/0/0
10/1/0/0
12784
C
T/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
12890
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
12964
T
G/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
12967
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
13109
A
G/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
13123
G
A/-/-
22/2/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
13310
T
A/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
13311
T
C/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
13489
G
A/-/-
23/1/0/0
10/0/0/0
12/0/0/0
12/0/0/0
10/1/0/0
13579
T
C/-/-
24/0/0/0
11/0/0/0
11/0/0/0
12/0/0/0
10/1/0/0
13641
A
G/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
13683
C
T/-/-
24/0/0/0
11/0/0/0
12/0/0/0
11/1/0/0
11/0/0/0
13696
G
A/-/-
22/2/0/0
11/0/0/0
12/0/0/0
11/1/0/0
10/1/0/0
13741
G
A/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
13776
A
G/-/-
23/1/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
13876
A
G/-/-
24/0/0/0
10/1/0/0
12/1/0/0
12/0/0/0
11/0/0/0
13882
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
13941
A
AT/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/0/0/0
13943
T
A/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
10/0/0/0
13955
A
G/-/-
24/0/0/0
10/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
13989
T
C/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
14105
G
A/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
14115
G
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14194
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/1/0/0
14233
T
C/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14291
C
T/-/-
21/3/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14359
G
A/-/-
23/1/0/0
10/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14373
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14384
C
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
14439
A
G/-/-
21/3/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14454
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14536
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/1/0/0
11/0/0/0
14540
C
T/-/-
24/0/0/0
11/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
14543
T
C/-/-
24/0/0/0
11/0/0/0
13/0/0/0
12/0/0/0
10/1/0/0
14549
A
T/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14582
T
TTA/-/-
24/0/0/0
11/0/0/0
12/0/0/0
12/0/0/0
11/0/0/0
14602
T
C/-/-
24/0/0/0
9/2/0/0
12/1/0/0
11/1/0/0
11/0/0/0
14642
C
T/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14686
T
C/-/-
24/0/0/0
10/1/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14691
T
A/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14741
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
14847
T
C/-/-
11/13/0/0
1/9/0/0
2/10/0/0
1/11/0/0
1/9/0/0
14875
A
G/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
10/0/0/0
14881
T
TA/-/-
24/0/0/0
11/0/0/0
11/1/0/0
12/0/0/0
9/0/0/0
14912
T
C/-/-
24/0/0/0
11/0/0/0
10/1/0/0
10/0/0/0
9/0/0/0
14916
C
T/-/-
23/1/0/0
11/0/0/0
11/0/0/0
10/0/0/0
8/1/0/0
14930
A
G/-/-
23/1/0/0
11/0/0/0
9/0/0/0
11/0/0/0
10/0/0/0
14936
A
G/-/-
24/0/0/0
11/0/0/0
9/1/0/0
11/0/0/0
11/0/0/0
14976
T,A
T/-/-
23/1/0/0
10/1/0/0
8/2/0/0
9/1/0/0
7/3/0/0
14978
A
C/T/
20/4/0/0
3/8/0/0
4/6/0/0
3/7/0/0
5/4/1/0
14984
A,T,T
TTT/AT/A
14/2/7/1
2/2/0/3
3/2/0/0
5/2/0/1
4/0/0/2
15002
A
T/-/-
22/2/0/0
5/2/0/0
4/1/0/0
3/5/0/0
4/2/0/0
15012
T
C/-/-
24/0/0/0
9/0/0/0
11/0/0/0
11/1/0/0
10/0/0/0
15035
T
TA/-/-
23/1/0/0
10/0/0/0
12/0/0/0
9/0/0/0
11/0/0/0
15036
A
G/-/-
24/0/0/0
10/0/0/0
12/0/0/0
9/0/0/0
9/2/0/0
15038
A
AATT/-/-
22/2/0/0
7/3/0/0
10/0/0/0
11/0/0/0
7/2/0/0
15039
A
G/-/-
24/0/0/0
10/0/0/0
10/0/0/0
7/4/0/0
8/1/0/0
15049
T
C/-/-
24/0/0/0
10/0/0/0
10/0/0/0
11/0/0/0
7/2/0/0
15050
T
TTA/-/-
23/1/0/0
10/0/0/0
10/0/0/0
11/0/0/0
9/0/0/0
15052
A
G/-/-
23/1/0/0
10/0/0/0
7/1/0/0
11/0/0/0
9/0/0/0
15057
C
T/-/-
23/1/0/0
10/0/0/0
8/0/0/0
8/0/0/0
9/0/0/0
15058
A
G/-/-
22/2/0/0
5/5/0/0
3/5/0/0
4/4/0/0
7/2/0/0
15066
T
C/-/-
23/1/0/0
10/0/0/0
10/0/0/0
11/0/0/0
8/0/0/0
15069
T
C/-/-
23/1/0/0
10/0/0/0
10/0/0/0
11/0/0/0
8/1/0/0
15082
A
G/-/-
22/2/0/0
10/0/0/0
10/0/0/0
11/0/0/0
8/2/0/0
15098
A
G/-/-
23/1/0/0
9/1/0/0
10/0/0/0
8/1/0/0
8/0/0/0
15115
T
C/-/-
21/3/0/0
8/1/0/0
8/2/0/0
8/1/0/0
8/0/0/0
15116
G
A/-/-
14/10/0/0
9/1/0/0
10/0/0/0
8/1/0/0
3/5/0/0
15128
A
G/-/-
23/1/0/0
8/1/0/0
6/1/0/0
8/0/0/0
7/0/0/0
15129
G
A/-/-
1/23/0/0
0/9/0/0
0/7/0/0
1/7/0/0
0/7/0/0
15144
T
A/-/-
24/0/0/0
10/0/0/0
13/0/0/0
11/1/0/0
11/0/0/0
15189
A
G/-/-
18/6/0/0
10/1/0/0
11/0/0/0
10/0/0/0
7/0/0/0
15243
T
C/-/-
15/9/0/0
10/1/0/0
7/1/0/0
7/0/0/0
6/0/0/0
15282
T
C/-/-
24/0/0/0
11/0/0/0
12/1/0/0
12/0/0/0
11/0/0/0
15301
C
A/-/-
24/0/0/0
11/0/0/0
12/1/0/0
11/0/0/0
11/0/0/0
15304
A
G/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/0/0/0
11/0/0/0
15305
T
TA/-/-
24/0/0/0
10/1/0/0
13/0/0/0
11/0/0/0
11/0/0/0
15321
C
T/-/-
23/1/0/0
11/0/0/0
11/2/0/0
11/0/0/0
11/0/0/0
15327
A
G/-/-
23/1/0/0
11/0/0/0
13/0/0/0
12/0/0/0
11/0/0/0
The lack of species-specific markers is also reflected in the phylogenetic tree (Figure 1). An. arabiensis, An. coluzzii and An. gambiae did not cluster separately, which is consistent with previous reports that compared mitochondrial genome sequence data from specimens originating from Kenya, Senegal and South Africa (Besansky et al. (1997)) and Burkina Faso, Cameroon, Kenya, Mali, South Africa, Tanzania and Zimbabwe (Fontaine et al. (2015), supplemental material).
Figure 1. Phylogenetic tree inferred from mtDNA genome sequence data.
With the exception of the outgroup, An. quadriannulatis, this analysis fails to reveal a clear division of the operational taxonomic units included in this analysis. Colors indicate the species or chromosomal form and numbers at the branches indicate the accuracy of the inferred branches on a scale of 0–100, where 100 represents the highest confidence.
Of note, 36 of the samples that we used in our study originated from Kela (Mali). Kela is located near the village of Selinkenyi, where previous studies have shown a history of hybridization and introgression between An. gambiae and An. coluzzii (Lee et al. (2013); Main et al. (2015); Norris et al. (2015)) , which may have resulted in shared polymorphisms in their mitochondrial genomes. Shared polymorphisms in their mitochondrial genomes, where history has not been reported, also appeared to have occurred in Mutengene (Cameroon), where both An. gambiae and An. coluzzii occur sympatrically. Hybridization between either An. coluzzii or An. gambiae with An. arabiensis yields sterile males (Slotman et al. (2004)), but phylogenomic analysis of these species show patterns of introgression between all of them (Fontaine et al. (2015)). Our mitochondrial genome study does not provide conclusive evidence for hybridization and introgression among the taxa under study. However, our data suggest that this is a possibility.
Data availability
Aligned sequences were submitted to the National Center for Biotechnology Information (NCBI) Accession number: MG930826 - MG930896
We thank University of California - Irvine, Malaria Initiatives (UCIMI) for their support.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
We thank Michelle Sanford for her assistance in the field collection in Cameroon in 2011. We thank Clare Marsden for providing the raw data of An. arabiensis samples.
Faculty Opinions recommended
References
Beard CB, Hamm DM, Collins FH:
The mitochondrial genome of the mosquito Anopheles gambiae: DNA sequence, genome organization, and comparisons with mitochondrial sequences of other insects.
Insect Mol Biol.
1993; 2(2): 103–124. PubMed Abstract
| Publisher Full Text
Besansky NJ, Lehmann T, Fahey GT, et al.:
Patterns of mitochondrial variation within and between African malaria vectors, Anopheles gambiae and An. arabiensis, suggest extensive gene flow.
Genetics.
1997; 147(4): 1817–1828. PubMed Abstract
| Free Full Text
Brown WM, George M Jr, Wilson AC:
Rapid evolution of animal mitochondrial DNA.
Proc Natl Acad Sci U S A.
1979; 76(4): 1967–1971. PubMed Abstract
| Free Full Text
Crawford JE, Lazzaro BP:
Assessing the accuracy and power of population genetic inference from low-pass next-generation sequencing data.
Front Genet.
2012; 3: 66. PubMed Abstract
| Publisher Full Text
| Free Full Text
Cronin MA, Palmisciano DA, Vyse ER, et al.:
Mitochondrial DNA in wildlife forensic science: species identification of tissues.
Wildlife Soc B (1973-2006).
1991; 19(1): 94–105. Reference Source
De Barba M, Adams JR, Goldberg CS, et al.:
Molecular species identification for multiple carnivores.
Conserv Genet Resour.
2014; 6(4): 821–824. Publisher Full Text
De Mandal S, Chhakchhuak L, Gurusubramanian G, et al.:
Mitochondrial markers for identification and phylogenetic studies in insects–A Review.
DNA Barcodes.
2014; 2(1). Publisher Full Text
Fontaine MC, Pease JB, Steele A, et al.:
Mosquito genomics. Extensive introgression in a malaria vector species complex revealed by phylogenomics.
Science.
2015; 347(6217): 1258524. PubMed Abstract
| Publisher Full Text
| Free Full Text
Fu Q, Mittnik A, Johnson PLF, et al.:
A revised timescale for human evolution based on ancient mitochondrial genomes.
Curr Biol.
2013; 23(7): 553–559. PubMed Abstract
| Publisher Full Text
| Free Full Text
Garrison E, Marth G:
Haplotype-based variant detection from short-read sequencing.
arXiv: 1207.3907.
Cornell University Library; 2013. Reference Source
Green CA:
Cytological maps for the practical identification of females of the three freshwater species of the Anopheles gambiae complex.
Ann Trop Med Parasitol.
1972; 66(1): 143–147. PubMed Abstract
| Publisher Full Text
Haag-Liautard C, Coffey N, Houle D, et al.:
Direct estimation of the mitochondrial DNA mutation rate in Drosophila melanogaster.
PLoS Biol.
2008; 6(8): e204. PubMed Abstract
| Publisher Full Text
| Free Full Text
Hanemaaijer MJ, Houston P, Collier TC, et al.:
Dataset 1: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division.
F1000Research.
2018. Data Source
Harrison RG:
Animal mitochondrial DNA as a genetic marker in population and evolutionary biology.
Trends Ecol Evol.
1989; 4(1): 6–11. PubMed Abstract
| Publisher Full Text
Lanzaro GC, Lee Y:
Speciation in Anopheles gambiae—The distribution of genetic polymorphism and patterns of reproductive isolation among natural populations.
Anopheles mosquitoes-New insights into malaria vectors.
InTech, 2013. Publisher Full Text
Lee Y, Marsden CD, Norris LC, et al.:
Spatiotemporal dynamics of gene flow and hybrid fitness between the M and S forms of the malaria mosquito, Anopheles gambiae.
Proc Natl Acad Sci U S A.
2013; 110(49): 19854–19859. PubMed Abstract
| Publisher Full Text
| Free Full Text
Lee Y, Marsden CD, Nieman C, et al.:
A new multiplex SNP genotyping assay for detecting hybridization and introgression between the M and S molecular forms of Anopheles gambiae.
Mol Ecol Resour.
2014; 14(2): 297–305. PubMed Abstract
| Publisher Full Text
| Free Full Text
Llamas B, Fehren-Schmitz L, Valverde G, et al.:
Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas.
Sci Adv.
2016; 2(4): e1501385. PubMed Abstract
| Publisher Full Text
| Free Full Text
Main BJ, Lee Y, Collier TC, et al.:
Complex genome evolution in Anopheles coluzzii associated with increased insecticide usage in Mali.
Mol Ecol.
2015; 24(20): 5145–5157. PubMed Abstract
| Publisher Full Text
| Free Full Text
Marsden CD, Lee Y, Kreppel K, et al.:
Diversity, differentiation, and linkage disequilibrium: prospects for association mapping in the malaria vector Anopheles arabiensis.
G3 (Bethesda).
2014; 4(1): 121–131. PubMed Abstract
| Publisher Full Text
| Free Full Text
Miles A, Harding N:
cggh/scikit-allel: v1.1.8 (Version v1.1.8).
Zenodo.
2017. Publisher Full Text
Norris LC, Main BJ, Lee Y, et al.:
Adaptive introgression in an African malaria mosquito coincident with the increased usage of insecticide-treated bed nets.
Proc Natl Acad Sci U S A.
2015; 112(3): 815–820. PubMed Abstract
| Publisher Full Text
| Free Full Text
Okonechnikov K, Conesa A, García-Alcalde F:
Qualimap 2: advanced multi-sample quality control for high-throughput sequencing data.
Bioinformatics.
2016; 32(2): 292–294. PubMed Abstract
| Publisher Full Text
| Free Full Text
Pegg GG, Sinclair B, Briskey L, et al.:
MtDNA barcode identification of fish larvae in the southern Great Barrier Reef–Australia.
Sci Mar.
2006; 70(S2): 7–12. Reference Source
Phillips MJ, Haouchar D, Pratt RC, et al.:
Inferring kangaroo phylogeny from incongruent nuclear and mitochondrial genes.
PLoS One.
2013; 8(2): e57745. PubMed Abstract
| Publisher Full Text
| Free Full Text
Pombi M, Caputo B, Simard F, et al.:
Chromosomal plasticity and evolutionary potential in the malaria vector Anopheles gambiae sensu stricto: insights from three decades of rare paracentric inversions.
BMC Evol Biol.
2008; 8(1): 309. PubMed Abstract
| Publisher Full Text
| Free Full Text
Scott JA, Brogdon WG, Collins FH:
Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction.
Am J Trop Med Hyg.
1993; 49(4): 520–529. PubMed Abstract
| Publisher Full Text
Shaw KL:
Conflict between nuclear and mitochondrial DNA phylogenies of a recent species radiation: what mtDNA reveals and conceals about modes of speciation in Hawaiian crickets.
Proc Natl Acad Sci U S A.
2002; 99(25): 16122–16127. PubMed Abstract
| Publisher Full Text
| Free Full Text
Slotman M, Della Torre A, Powell JR:
The genetics of inviability and male sterility in hybrids between Anopheles gambiae and An. arabiensis.
Genetics.
2004; 167(1): 275–287. PubMed Abstract
| Publisher Full Text
| Free Full Text
Sota T, Vogler AP:
Incongruence of mitochondrial and nuclear gene trees in the Carabid beetles Ohomopterus.
Syst Biol.
2001; 50(1): 39–59. PubMed Abstract
| Publisher Full Text
Tahir HM, Mehwish, Kanwal N, et al.:
Genetic diversity in cytochrome c oxidase I gene of Anopheles mosquitoes.
Mitochondrial DNA A DNA Mapp Seq Anal.
2016; 27(6): 4298–4301. PubMed Abstract
| Publisher Full Text
1
Vector Genetics Laboratory, Department of Pathology, Microbiology and Immunology, University of California Davis, Davis, CA, 95616, USA 2
Malaria Research and Training Center, University of Bamako, Bamako, E2528, Mali 3
Mosquito Control Research Laboratory, Kearney Agricultural Center, Department of Entomology and Nematology, University of California Davis, Davis, CA, 93648, USA
Mark J. Hanemaaijer
Roles:
Conceptualization,
Formal Analysis,
Writing – Original Draft Preparation,
Writing – Review & Editing
Parker D. Houston
Roles:
Writing – Original Draft Preparation
Travis C. Collier
Roles:
Data Curation,
Software
Laura C. Norris
Roles:
Investigation
Abdrahamane Fofana
Roles:
Investigation
Gregory C. Lanzaro
Roles:
Writing – Review & Editing
Anthony J. Cornel
Roles:
Investigation,
Writing – Review & Editing
Yoosook Lee
Roles:
Conceptualization,
Data Curation,
Formal Analysis,
Writing – Original Draft Preparation,
Writing – Review & Editing
We thank University of California - Irvine, Malaria Initiatives (UCIMI) for their support.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Hanemaaijer MJ, Houston PD, Collier TC et al. Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.12688/f1000research.13807.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
Sallum MAM. Reviewer Report For: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.5256/f1000research.15009.r33371)
General comment
Phylogenetic analysis need to be improved, and the choice for NJ methods and JC model, justified in the article. There are several programs that have been largely employed for phylogenetic analysis, including for mitogenome data. The paper
... Continue reading
General comment
Phylogenetic analysis need to be improved, and the choice for NJ methods and JC model, justified in the article. There are several programs that have been largely employed for phylogenetic analysis, including for mitogenome data. The paper authored by Foster et al.1 contains useful information about analyses that have been carried out for inferring phylogenetic relationships within Anophelinae mosquitoes. I strongly suggest authors to verify how analyses were done.
Sample collection Authors - “The An. gambiae and An. coluzzii samples were collected as resting adults using mouth aspirators in Kela, Mali (11.88683°N, -8.44744°W) in 2012 and Mutengene, Cameroon (4.0994°N, 9.3081°W) in 2011.”
Comment - Can you please give more details the micro environment where your specimens of An. gambiae and An. coluzzii were resting?
Authors - “Similarly, we did this for the An. gambiae Savannah and Bamako chromosomal forms. We used the same definitions and methods to characterize the chromosomal forms as in Lanzaro & Lee, 2013.”
Comment - It is not clear to me if you examined the polytene chromosome of each specimen you identified as the Savannah, Bamako, Forest and Mopti forms. Please clarify.
Genome sequencing Authors - “For the An. coluzzii and An. gambiae samples we used the same methods as described in Norris et al. (2015) and Main et al. (2015). For the latter species, libraries were created using the Nextera DNA Sample Preparation Kit (FC-121-1031) and TruSeq dual indexing barcodes (FC-121-103) (Illumina) and the samples were sequenced on an Illumina HiSeq2500 with 100-bp paired end reads.”
Comment - Please add a short sentence to clarify if you sequenced the whole genome and from the full sequence data you obtained the positions 1-13,470 of the mitogenome.
Data analysis Authors - “The phylogenetic tree was generated using the Jukes-Cantor genetic distance model and Neighbor-Joining tree methods available in Geneious version 10.1.3.”
Comment - Authors should clarify their choice for sequence analysis. The Geneious software has been developed for editing and aligning DNA / amino acid sequences. There are several softwares, which have been largely used to infer phylogenetic relationships. I suggest authors to refining and improving the phylogenetic analysis using appropriate programs and models that have been chosen for the mitogenome data you have at hand.
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
No
If applicable, is the statistical analysis and its interpretation appropriate?
Partly
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Partly
References
1. Foster PG, de Oliveira TMP, Bergo ES, Conn JE, et al.: Phylogeny of Anophelinae using mitochondrial protein coding genes.R Soc Open Sci. 2017; 4 (11): 170758 PubMed Abstract | Publisher Full Text
Competing Interests: No competing interests were disclosed.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard, however I have significant reservations, as outlined above.
Sallum MAM. Reviewer Report For: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.5256/f1000research.15009.r33371)
Caputo B and Pichler V. Reviewer Report For: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.5256/f1000research.15009.r32266)
General comments
The present research note entitled:” Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division” is well analysed, reported and written. As already reported in previous study the submitted manuscript suggested the
... Continue reading
General comments
The present research note entitled:” Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division” is well analysed, reported and written. As already reported in previous study the submitted manuscript suggested the absence of any species-specific differences in the mitogenome of the three species examined. Although the manuscript is not innovative and the research is not based on any previous evidence, the present note confirms previous suggestions by examining the whole mitogenome of 70 specimens from field specimens and find the lack of species or chromosomal form specific markers.
Title and abstract
Title and abstract are appropriate and summarize well the content of the article.
Introduction
The introduction gives a good description of the aims of the present study, although I would have added some references to previous studies performed on mtDNA of the examined species (for example Besansky 1997) and why you expected to obtain different results compared to previous studies.
Please revise also:
“morphologically identical species that can only be distinguished with molecular markers” (Scott et al., 1993; Coetzee et al., 2013)
The currently used molecular markers are located within genomic islands of divergence located proximal to the centromeres (Lee et al. (2014); Turner et al. (2005)) please rephrase the citation and refer it only to detect genomic differences between An.gambiae e and An.coluzzii.
Please insert a sentence about chromosomal forms of An.gambiae.
Methods
Please specified the method for collecting An. arabiensis as you already described for An.gambiae (e.g. indoor specimens, mouth aspirators, PSC collections).
Please insert a table with inversion polymorphism of chromosomal forms analyzed.
Please add the source of the An. quadriannulatus specimens you included in the phylogenetic analysis.
Results
Study design is well explained and results are given concisely.
Please add in Table 2 also the number of specimens you included for each species in the analysis.
Please add in Figure two an explanation of what “lineage” means for An. arabiensis specimens.
Please give results (also without table or figure) for each country separately.
Discussion
Discussion is very concise but deals with most major points of interest. We would just suggest to explain better the conclusion on possible introgression (the more plausible hypothesis) between taxa and to evaluate other possible explanations for the absence of fixed differences between species (e.g. absence for divergent selection, or evolutionary characherestic of mitogenomes).
Is the work clearly and accurately presented and does it cite the current literature?
Yes
Is the study design appropriate and is the work technically sound?
Yes
Are sufficient details of methods and analysis provided to allow replication by others?
Yes
If applicable, is the statistical analysis and its interpretation appropriate?
Yes
Are all the source data underlying the results available to ensure full reproducibility?
Yes
Are the conclusions drawn adequately supported by the results?
Yes
Competing Interests: No competing interests were disclosed.
We confirm that we have read this submission and believe that we have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Caputo B and Pichler V. Reviewer Report For: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division [version 1; peer review: 1 approved, 1 approved with reservations]. F1000Research 2018, 7:347 (https://doi.org/10.5256/f1000research.15009.r32266)
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Spreadsheet data files may not format correctly if your computer is using different default delimiters (symbols used to separate values into separate cells) - a spreadsheet created in one region is sometimes misinterpreted by computers in other regions. You can change the regional settings on your computer so that the spreadsheet can be interpreted correctly.
How to fix it
Save downloaded CSV file
Open spreadsheet program (e.g. Excel)
Click the ‘Data’ tab at the top
Click the ‘From text’ icon (top left)
Browse for downloaded CSV file, click ‘Import’
Ensure ‘Delimited’ radio button is selected, click ‘Next’
Check one of the appropriate delimiter checkboxes (you can visualize the formatting by looking at the data preview below these options)
Hanemaaijer MJ, Houston PD, Collier TC et al.. Dataset 1 in: Mitochondrial genomes of Anopheles arabiensis, An. gambiae and An. coluzzii show no clear species division. F1000Research 2018, 7:347 (https://doi.org/10.5256/f1000research.13807.d192892)
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
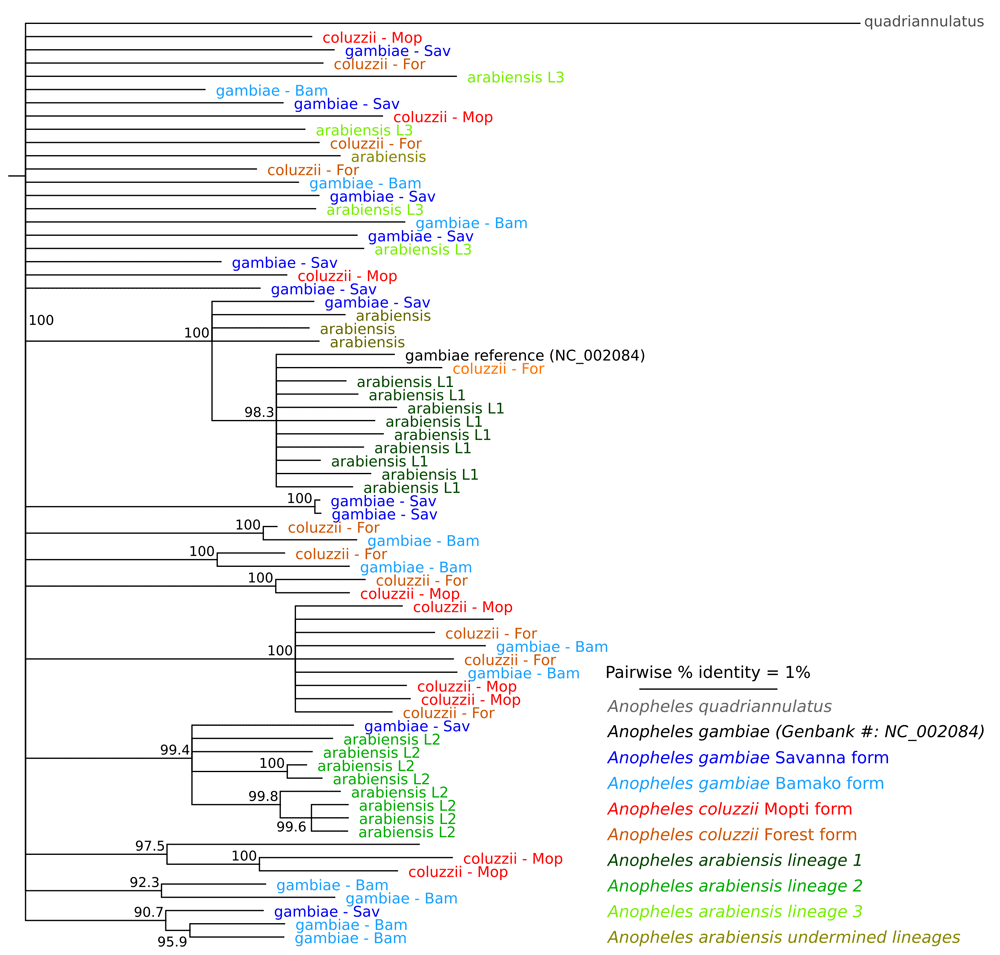
Comments on this article Comments (0)