Katz E, Nisani S and Chamovitz DA. Indole-3-carbinol: a plant hormone combatting cancer [version 1; peer review: 2 approved]. F1000Research 2018, 7(F1000 Faculty Rev):689 (https://doi.org/10.12688/f1000research.14127.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
Daniel A. Chamovitz
Roles:
Project Administration,
Supervision,
Writing – Original Draft Preparation,
Writing – Review & Editing
OPEN PEER REVIEW
REVIEWER STATUS
Abstract
A diet rich in cruciferous vegetables such as cauliflower, broccoli, and cabbage has long been considered healthy, and various epidemiological studies suggest that the consumption of cruciferous vegetables contributes to a cancer-protecting diet. While these vegetables contain a vast array of phytochemicals, the mechanism by which these vegetables counteract cancer is still largely unresolved. Numerous in situ studies have implicated indole-3-carbinol, a breakdown product of the glucosinolate indole-3-ylmethylglucosinolate, as one of the phytochemicals with anti-cancer properties. Indole-3-carbinol influences a range of cellular processes, but the mechanisms by which it acts on cancer cells are slowly being revealed. Recent studies on the role of indole-3-carbinol in Arabidopsis opens the door for cross-kingdom comparisons that can help in understanding the roles of this important phytohormone in both plant biology and combatting cancer.
Keywords
glucosinolates, indole-3-carbinol, cancer prevention, cruciferous vegetables
Corresponding author:
Daniel A. Chamovitz
Competing interests:
No competing interests were disclosed.
Grant information:
Our research into the role of I3C in Arabidopsis is funded by grants from the Binational Agricultural Research and Development Fund (BARD, IS‐4505‐12R and US‐4846‐15C).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
A diet rich in cruciferous vegetables such as cauliflower, broccoli, and cabbage has long been considered healthy. Even in ancient times, extracts from these vegetables were thought to have medicinal and curative properties, and both Pythagoras and Hippocrates understood the medicinal properties of mustard extracts1. In the 20th century, epidemiological studies pointing to the protective properties of cruciferous vegetables in a cancer-protecting diet started to accumulate2. A meta-analysis of studies carried out over 18 years in Europe revealed an inverse association between weekly consumption of cruciferous vegetables and several common cancers, including colorectal, breast, kidney, and upper digestive tract cancers3. While these vegetables contain a vast array of phytochemicals4, the mechanism by which these vegetables counteract cancer is still largely unresolved.
The anticarcinogenic properties associated with crucifers are primarily attributed to the presence of glucosinolates, a family of secondary metabolites that are synthesized uniquely in this plant family and play a dominant role in plant defenses against insects5. Glucosinolates are derived from glucose and amino acids and contain various modifications to their side chain. The exact glucosinolate profile varies among crucifer species, and more than 140 glucosinolates have been characterized, including approximately 40 in Arabidopsis6,7. Herbivory or other tissue damage initiates the hydrolysis of glucosinolates by an endogenous plant β-thioglucosidase termed “myrosinase”. Glucosinolates and myrosinase are stored in separate plant compartments and meet only following cell rupture. This separation is likely an adaptation to allow the targeted production of glucosinolate breakdown products, which are toxic to not only herbivores and pathogens but also the plants themselves. Further catalysis and spontaneous degradation results in the formation of nitriles, epithionitriles, oxazolidine-2-thiones, thiocyanates, and isothiocyanates8,9. These glucosinolate breakdown products cause the characteristic sharp taste of cruciferous vegetables and typically have a deterrent effect on generalist herbivores10–12.
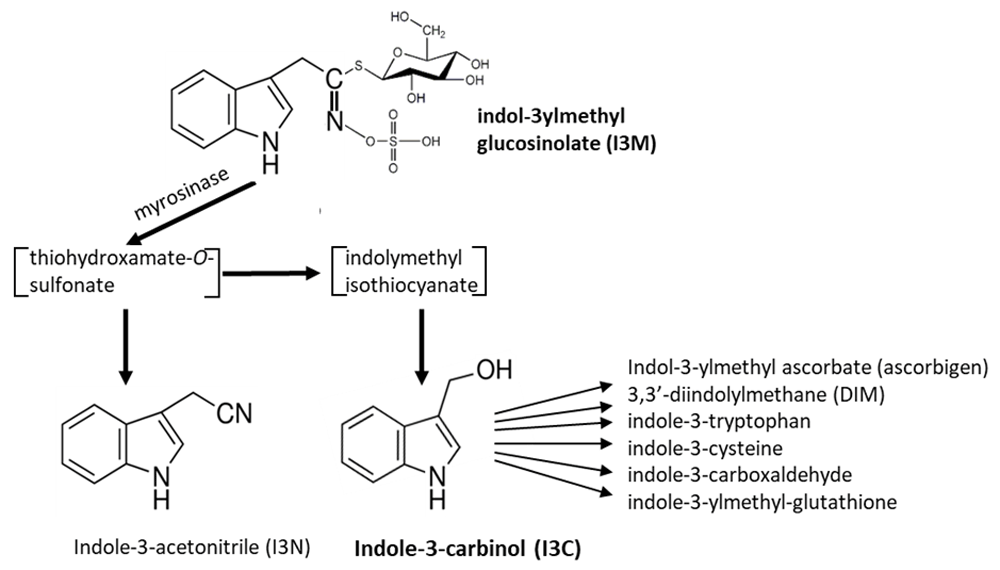
Breakdown of indole-3-ylmethylglucosinolate (I3M-GS), one of the most widely distributed glucosinolates, leads to the formation of indole-3-acetonitrile (I3N) and indole-3-carbinol (I3C) (Figure 1)13. I3C, in turn, can react with itself and a variety of other plant metabolites to form conjugates, some of which are shown in Figure 1. Most of these I3C conjugates have as-yet-unknown functions in plant metabolism, though, interestingly, another function of glucosinolate breakdown products may be to signal further plant defense responses14. Therefore, it is possible that I3M-GS breakdown also triggers other downstream responses in Arabidopsis and other crucifers.
Figure 1. Myrosinase-catalyzed breakdown of indol-3-ylmethylglucosinolate (I3M-GS).
Myrosinase-catalyzed breakdown of I3M-GS leads to the formation of unstable intermediates and then to indole-3-acetonitrile (I3N) and indole-3-carbionol (I3C). I3C reacts with itself and other plant metabolites to form a number of conjugates, some of which are shown.
From a dietary perspective, cooking vegetables affects their profile of glucosinolate breakdown15. Boiling leads to inactivation of the myrosinase enzyme but can also lead to a non-enzymatic breakdown of I3M-GS to I3C and I3N16. In addition, human gut microbes can lead to glucosinolate breakdown17.
Indole-3-carbinol and cancer
The glucosinolate breakdown products, rather than intact glucosinolates, primarily contribute to the anticarcinogenic effects of eating cabbage, broccoli, and related vegetables11,12,18. I3C has long been studied regarding potential roles in cancer management19,20, and many studies showed that I3C suppresses the proliferation of various cancer cell lines, including breast, colon, prostate, and endometrial cancer cells (reviewed in 19,21). One example of its anti-proliferative properties comes from a study conducted on non-tumorigenic and tumorigenic breast epithelial cells (MCF10A and MCF10CA1a, respectively), which showed that I3C induced apoptosis in the breast cancer cells but not in the non-tumorigenic breast epithelial cells22. I3C and one of its reaction products, diindolylmethane (DIM), were implicated in the induction of phase 1 detoxification enzymes, which can result in the breakdown of other dietary carcinogens. Both in situ and in vivo studies point to a role for I3C as a chemoprotective agent in breast and prostate cancer23.
The exact mechanisms by which I3C influences human cells are unclear, though direct interaction with a variety of signaling pathways has been proposed. The treatment of various cancer cells with I3C induces G1 cell cycle arrest24–30. Other studies pointed to a connection between treatment of I3C and stimulation of apoptosis in several tumor cells25,31–34. I3C also induced autophagy in different cell lines. For example, the treatment of human colon cancer HT-29 cells with I3C and genistin induced autophagy and suppressed the cells' viability35. The treatment of human breast cancer cell lines with a cyclic tetrameric derivative of I3C resulted in upregulation of key signaling molecules involved in endoplasmic reticulum stress response and autophagy36. I3C also has the potential to modulate the metabolism of estrogen, and, through this, it may lower the risk of hormone-dependent cancers37–39. In addition, I3C inhibited tumor invasion and metastasis40–44 and modulated the activity of several transcription factors and various protein kinases21. Interestingly, I3C may also be involved in the inhibition of amyloid fibril formation45. I3C was proposed to act as an angiogenesis agent, as it was shown to inhibit the development of new blood vessels46. A number of studies have shown that I3C treatment leads to various changes in gene expression, including changes in key microRNAs (reviewed in 47). The complex mixture of indole metabolites found in cruciferous vegetables likely has synergistic anticarcinogenic effects that are not seen in experiments with individual compounds32.
In vivo studies showed that I3C inhibits the development of different cancers in several animals when given before or in parallel to a carcinogen. However, when I3C was given to the animals after the carcinogen, I3C promoted carcinogenesis48. This concern regarding the long-term effects of I3C treatment on cancer risk in humans resulted in some caution in the use of I3C as a dietary supplement in cancer management protocols49,50.
Mammalian cellular processes attributed to I3C action are as diverse as the different phenotypes presented by the many cancers studied. Indeed, focusing on the effects of I3C on one type of cancer (e.g. breast cancer) may present pleiotropic effects for I3C on multiple molecular targets (reviewed in 51) that may be distinct from those presented in another cancer type. While not often considered, to get a different perspective on the action of I3C in cells in general, it may be instructive to learn from the activity of I3C in plants.
Indole-3-carbinol and plants
The model plant Arabidopsis thaliana provides an excellent system for elucidating the molecular mechanisms involved in I3C action, as 1) it produces I3C endogenously following herbivory, 2) small amounts of I3C are produced constitutively in the roots, hinting at an endogenous role in maintaining homeostasis, and 3) its short life cycle and small stature coupled with advanced available genetic and genomic resources make Arabidopsis an excellent model system not only for plant biology but also for eukaryotic research in general52.
While the role of I3C in deterring herbivores is well studied53, as is the biochemical pathway leading to the production of I3C13, the secondary responses in plants induced by I3C are only now starting to be revealed. Our recent studies highlight that I3C is not only a defensive chemical targeting herbivores but also a signaling molecule modulating different cellular and developmental pathways.
Using Arabidopsis as a model system, we showed that exogenously applied I3C rapidly and reversibly inhibited root elongation in a dose-dependent manner54. This inhibition was accompanied by three I3C-induced responses that are relevant for our understanding of I3C activity in inhibiting cancer.
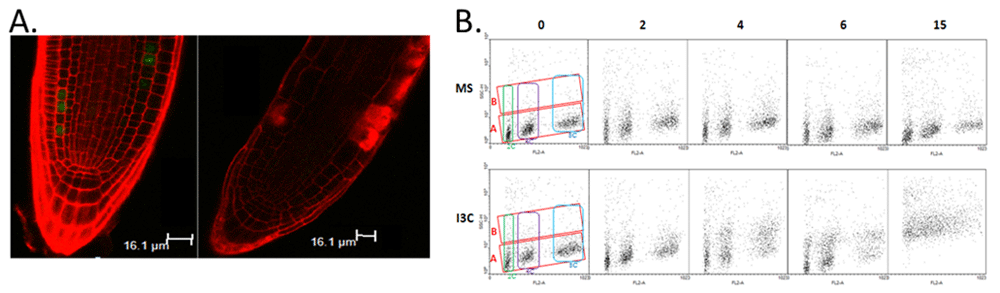
First, the application of I3C led to a cessation of cell division in the root meristem (Figure 2A). While normally a number of CycB1-expressing cells are visible in the root meristem, following I3C treatment, no CycB1-containing cells were detected, indicating a cessation of cell division. This conclusion is supported by transcript-profiling results showing the downregulation of cell cycle genes six hours following exposure to I3C55.
Fluorescence-activated cell sorting (FACS) analysis on nuclei isolated from root tips further indicates a stoppage of the cell cycle. As seen in Figure 2B, three distinct populations of nuclei are detected in untreated roots, corresponding to 2n, 4n, and 8n nuclei. However, following treatment with I3C, there is a progressive loss of the 4n and 8n populations, with a concurrent increase in cells with increased side-scatter (population B).
Figure 2. Indole-3-carbionol (I3C) affects cell cycle and nuclear complexity in the Arabidopsis root meristem.
A. Confocal imaging reveals a lack of Cyclin B–GFP-expressing cells following I3C treatment. Seedlings expressing Cyclin B–GFP were grown on Murashige and Skoog (MS) medium for 4 days, treated with MS (left panel) or 500 μm I3C (right panel) for 6 hours, and imaged using confocal microscopy. Cell walls were stained using propidium iodide. B. Fluorescence-activated cell sorting (FACS) analysis reveals changes in nuclear complexity following I3C treatment. Nuclei were isolated from Arabidopsis roots treated with MS or MS plus I3C for the marked times between 0 and 15 hours and analyzed by FACS for DNA content (FL2-A = propidium iodide fluorescence) and nuclear complexity (SCC-H = light side scatter). The green, purple, and blue boxes represent the populations differing according to nuclear content, 2n, 4n, and the endoreplication population (8n), respectively. The red boxes represent two populations of nuclei ("A" and "B") that differ according to side scatter.
Second, the application of I3C led to a loss of auxin (indole-3-acetic acid [IAA]) activity in the root meristem54. Auxin is the most central plant hormone, controlling nearly all aspects of plant growth and development56. I3C affects plant growth and development by directly modulating auxin signaling. I3C antagonized a number of auxin-induced growth phenotypes, including inhibition of root elongation, formation of root hairs, and secondary root branching. I3C directly interferes with the auxin-dependent binding of the auxin-receptor Transport Inhibitor Response (TIR1) to two of its substrates54. The TIR1 auxin receptor is an F-box-containing subunit of the SCF (Skp, Cullin, F-box) E3 ubiquitin ligase complex. Auxin binding to the SCFTIR1/AFB promotes the degradation of auxin/indole-3-acetic acid (Aux/IAA) transcriptional repressors and through this regulates the transcription of the auxin-induced genes57. I3C inhibits the auxin-dependent dimerization of the receptor with its substrates by competing with auxin for the same binding site in TIR1.
The third I3C-induced response relevant for our understanding of I3C activity in inhibiting cancer is autophagy. Exposure of Arabidopsis roots to I3C leads to the induction of autophagy58. This autophagy is not general, aimed at bulk degradation of general cytoplasmic content for recycling, such as that occuring under starvation conditions, but rather specific. Specific autophagy targets damaged proteins and other cellular components for degradation59 and is seen in the co-localization of the GFP-AtATG8A and mCherry-AtNBR1 marker proteins in autophagosomes following I3C treatment. The I3C-induced autophagy targets the TIR1 auxin receptor, thus connecting I3C-dependent inhibition of auxin signaling55 and the I3C induction of autophagy58.
These two I3C-dependent processes were detected in roots not only after direct exposure to exogenously applied I3C but also following the treatment of leaves with I3C. Most importantly, leaf-wounding also induced autophagy and inhibited the auxin response in the root, and this effect of wounding was lost in glucosinolate-defective mutants. This indicates that an I3C-dependent signal is transported from leaves to the root meristem where auxin signaling is inhibited and autophagy is induced. Thus, I3C is not only a defensive metabolite that repels insects but also involved in long-distance communication regulating growth and development in plants.
Indole-3-carbinol, autophagy, and protein turnover
The connection between I3C and autophagy is quite interesting, as this connection was also found in several human cancer cells, as described earlier35,36. The process of autophagy involves the degradation of unnecessary or dysfunctional cellular components through the actions of lysosomes (in mammals) or vacuoles (in plants). This process is evolutionarily conserved among eukaryotes, and its mechanism is well elucidated60,61. In the context of cancer, autophagy can be viewed as a “double-edged sword”. The activation of autophagy may function as a tumor suppressor (by degrading the defective organelles and cellular component) or can be exploited by the cancer cells to generate nutrients and energy during periods of starvation62.
Although I3C-induced autophagy was detected in both plants and animals, the direct signaling mechanism has yet to be elucidated. However, this might hint at a shared signaling mechanism for both plants and humans. Thus, not only is it instructive for cancer biologists to learn from the activity of I3C in plants but also plant biologists have much to gain from a closer understanding of the mechanistic studies of cancer biologists.
To date, only a few I3C-binding proteins have been identified. In human cells, the enzyme elastase, which mediates the conversion of cyclin E from a higher- to a lower-molecular-weight form associated with cancer cell proliferation, was the first identified specific target protein for I3C63. I3C treatments also inhibited elastase-dependent cleavage of an additional substrate, membrane-associated CD40, a member of the tumor necrosis factor (TNF) receptor superfamily64. Thus, the I3C–elastase nexus may aid in the development of targeted therapies of human breast cancers where high elastase levels are correlated with poor prognosis.
The only I3C-binding protein identified in plants to date is the TIR1 F-box protein. While auxin is a plant-specific hormone, SCF complexes also exist in mammals and play important roles in many mammalian functions65. TIR1 is related to the human protein SKP266, so conceivably I3C could regulate protein turnover in mammals as well. This conjecture is supported by studies which showed that I3C targets and inhibits a different E3 ubiquitin ligase, NEDD4-1 (Neural precursor cell Expressed Developmentally Down regulated gene 4-1) in human melanoma cells67,68. Thus, an I3C-bound, inhibited NEDD4 would need to be cleared from the cell, and conceivably this could occur via specific autophagy, just as I3C-bound, inhibited TIR1 is targeted in plant roots for clearing by specific autophagy. As NEDD4 is frequently overexpressed in different types of human cancers69, I3C could be a potential therapeutic agent inhibiting the activity of the over-accumulated E3 ligase.
While plants do not develop metastatic cancer as mammals do, plants can develop tumors. Plants and animals share numerous pathways and signaling cascades70, and approximately 70% of the genes implicated in cancer have homologs in the Arabidopsis thaliana genome, similar to the percentages of human cancer genes in other established systems such as Drosophila melanogaster, Caenorhabditis elegans, and Saccharomyces cerevisiae52. Furthermore, while plants and animals obviously have independent hormonal regulatory mechanisms, some similarity and cross reactivity exists between the kingdoms. Plants produce phyto-estrogens and other steroid hormones, which also affect human hormone signaling, as well as a number of putative steroid hormone-binding proteins71–75. Thus, the study of I3C in plants can have direct implications for further understanding the role of I3C and perhaps controlling cancer in humans.
Competing interests
The authors declare that they have no competing interests.
Grant information
Our research into the role of I3C in Arabidopsis is funded by grants from the Binational Agricultural Research and Development Fund (BARD, IS-4505-12R and US-4846-15C).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Faculty Opinions recommended
References
1.
Fenwick GR, Heaney RK:
Glucosinolates and their breakdown products in cruciferous crops, foods and feedingstuffs.
Food Chem.
1983; 11(4): 249–71. Publisher Full Text
2.
Verhoeven DT, Goldbohm RA, van Poppel G, et al.:
Epidemiological studies on brassica vegetables and cancer risk.
Cancer Epidemiol Biomarkers Prev.
1996; 5(9): 733–48. PubMed Abstract
3.
Bosetti C, Filomeno M, Riso P, et al.:
Cruciferous vegetables and cancer risk in a network of case-control studies.
Ann Oncol.
2012; 23(8): 2198–203. PubMed Abstract
| Publisher Full Text
4.
Björkman M, Klingen I, Birch AN, et al.:
Phytochemicals of Brassicaceae in plant protection and human health--influences of climate, environment and agronomic practice.
Phytochemistry.
2011; 72(7): 538–56. PubMed Abstract
| Publisher Full Text
5.
Fahey JW, Zalcmann AT, Talalay P:
The chemical diversity and distribution of glucosinolates and isothiocyanates among plants.
Phytochemistry.
2001; 56(1): 5–51. PubMed Abstract
| Publisher Full Text
7.
Reichelt M, Brown PD, Schneider B, et al.:
Benzoic acid glucosinolate esters and other glucosinolates from Arabidopsis thaliana.
Phytochemistry.
2002; 59(6): 663–71. PubMed Abstract
| Publisher Full Text
8.
Halkier BA, Gershenzon J:
Biology and biochemistry of glucosinolates.
Annu Rev Plant Biol.
2006; 57: 303–33. PubMed Abstract
| Publisher Full Text
9.
Sønderby IE, Geu-Flores F, Halkier BA:
Biosynthesis of glucosinolates--gene discovery and beyond.
Trends Plant Sci.
2010; 15(5): 283–90. PubMed Abstract
| Publisher Full Text
10.
Bones AM, Rossiter JT:
The enzymic and chemically induced decomposition of glucosinolates.
Phytochemistry.
2006; 67(11): 1053–67. PubMed Abstract
| Publisher Full Text
11.
Keck AS, Finley JW:
Cruciferous vegetables: cancer protective mechanisms of glucosinolate hydrolysis products and selenium.
Integr Cancer Ther.
2004; 3(1): 5–12. PubMed Abstract
| Publisher Full Text
12.
Kim YS, Milner JA:
Targets for indole-3-carbinol in cancer prevention.
J Nutr Biochem.
2005; 16(2): 65–73. PubMed Abstract
| Publisher Full Text
13.
Agerbirk N, De Vos M, Kim JH, et al.:
Indole glucosinolate breakdown and its biological effects.
Phytochem Rev.
2009; 8: 101–20. Publisher Full Text
16.
Campbell LD, Slominski BA:
Extent of thermal decomposition of indole glucosinolates during the processing of canola seed.
J Am Oil Chem Soc.
1990; 67(2): 73–5. Publisher Full Text
17.
Shapiro TA, Fahey JW, Wade KL, et al.:
Human metabolism and excretion of cancer chemoprotective glucosinolates and isothiocyanates of cruciferous vegetables.
Cancer Epidemiol Biomarkers Prev.
1998; 7(12): 1091–100. PubMed Abstract
18.
Bones AM, Rossiter JT:
The myrosinase-glucosinolate system, its organisation and biochemistry.
Physiol Plant.
1996; 97(1): 194–208. Publisher Full Text
20.
Verhoeven DT, Verhagen H, Goldbohm RA, et al.:
A review of mechanisms underlying anticarcinogenicity by brassica vegetables.
Chem Biol Interact.
1997; 103(2): 79–129. PubMed Abstract
| Publisher Full Text
21.
Aggarwal BB, Ichikawa H:
Molecular targets and anticancer potential of indole-3-carbinol and its derivatives.
Cell Cycle.
2005; 4(9): 1201–15. PubMed Abstract
| Publisher Full Text
22.
Rahman KM, Aranha O, Sarkar FH:
Indole-3-carbinol (I3C) induces apoptosis in tumorigenic but not in nontumorigenic breast epithelial cells.
Nutr Cancer.
2003; 45(1): 101–12. PubMed Abstract
| Publisher Full Text
23.
Bradlow HL:
Review. Indole-3-carbinol as a chemoprotective agent in breast and prostate cancer.
In Vivo.
2008; 22(4): 441–5. PubMed Abstract
24.
Cover CM, Hsieh SJ, Tran SH, et al.:
Indole-3-carbinol inhibits the expression of cyclin-dependent kinase-6 and induces a G1 cell cycle arrest of human breast cancer cells independent of estrogen receptor signaling.
J Biol Chem.
1998; 273(7): 3838–47. PubMed Abstract
| Publisher Full Text
25.
Chinni SR, Li Y, Upadhyay S, et al.:
Indole-3-carbinol (I3C) induced cell growth inhibition, G1 cell cycle arrest and apoptosis in prostate cancer cells.
Oncogene.
2001; 20(23): 2927–36. PubMed Abstract
| Publisher Full Text
26.
Brandi G, Paiardini M, Cervasi B, et al.:
A new indole-3-carbinol tetrameric derivative inhibits cyclin-dependent kinase 6 expression, and induces G1 cell cycle arrest in both estrogen-dependent and estrogen-independent breast cancer cell lines.
Cancer Res.
2003; 63(14): 4028–36. PubMed Abstract
27.
Zhang J, Hsu B A JC, Kinseth B A MA, et al.:
Indole-3-carbinol induces a G1 cell cycle arrest and inhibits prostate-specific antigen production in human LNCaP prostate carcinoma cells.
Cancer.
2003; 98(11): 2511–20. PubMed Abstract
| Publisher Full Text
28.
Cover CM, Hsieh SJ, Cram EJ, et al.:
Indole-3-carbinol and tamoxifen cooperate to arrest the cell cycle of MCF-7 human breast cancer cells.
Cancer Res.
1999; 59(6): 1244–51. PubMed Abstract
29.
Matsuzaki Y, Koyama M, Hitomi T, et al.:
Indole-3-carbinol activates the cyclin-dependent kinase inhibitor p15INK4b gene.
FEBS Lett.
2004; 576(1–2): 137–40. PubMed Abstract
| Publisher Full Text
30.
Chen Z, Tao ZZ, Chen SM, et al.:
Indole-3-carbinol inhibits nasopharyngeal carcinoma growth through cell cycle arrest in vivo and in vitro.
PLoS One.
2013; 8(12): e82288. PubMed Abstract
| Publisher Full Text
| Free Full Text
31.
Ge X, Fares FA, Yannai S:
Induction of apoptosis in MCF-7 cells by indole-3-carbinol is independent of p53 and bax.
Anticancer Res.
1999; 19(4B): 3199–203. PubMed Abstract
32.
Bonnesen C, Eggleston IM, Hayes JD:
Dietary indoles and isothiocyanates that are generated from cruciferous vegetables can both stimulate apoptosis and confer protection against DNA damage in human colon cell lines.
Cancer Res.
2001; 61(16): 6120–30. PubMed Abstract
33.
Safa M, Jafari L, Alikarami F, et al.:
Indole-3-carbinol induces apoptosis of chronic myelogenous leukemia cells through suppression of STAT5 and Akt signaling pathways.
Tumour Biol.
2017; 39(6): 1010428317705768. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
34.
Perez-Chacon G, de Los Rios C, Zapata JM:
Indole-3-carbinol induces cMYC and IAP-family downmodulation and promotes apoptosis of Epstein-Barr virus (EBV)-positive but not of EBV-negative Burkitt's lymphoma cell lines.
Pharmacol Res.
2014; 89: 46–56. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
35.
Nakamura Y, Yogosawa S, Izutani Y, et al.:
A combination of indol-3-carbinol and genistein synergistically induces apoptosis in human colon cancer HT-29 cells by inhibiting Akt phosphorylation and progression of autophagy.
Mol Cancer.
2009; 8: 100. PubMed Abstract
| Publisher Full Text
| Free Full Text
36.
Galluzzi L, De Santi M, Crinelli R, et al.:
Induction of endoplasmic reticulum stress response by the indole-3-carbinol cyclic tetrameric derivative CTet in human breast cancer cell lines.
PLoS One.
2012; 7(8): e43249. PubMed Abstract
| Publisher Full Text
| Free Full Text
37.
Telang NT, Katdare M, Bradlow HL, et al.:
Inhibition of proliferation and modulation of estradiol metabolism: novel mechanisms for breast cancer prevention by the phytochemical indole-3-carbinol.
Proc Soc Exp Biol Med.
1997; 216(2): 246–52. PubMed Abstract
| Publisher Full Text
38.
Meng Q, Yuan F, Goldberg ID, et al.:
Indole-3-carbinol is a negative regulator of estrogen receptor-alpha signaling in human tumor cells.
J Nutr.
2000; 130(12): 2927–31. PubMed Abstract
| Publisher Full Text
39.
Riby JE, Feng C, Chang YC, et al.:
The major cyclic trimeric product of indole-3-carbinol is a strong agonist of the estrogen receptor signaling pathway.
Biochemistry.
2000; 39(5): 910–8. PubMed Abstract
| Publisher Full Text
40.
Meng Q, Goldberg ID, Rosen EM, et al.:
Inhibitory effects of Indole-3-carbinol on invasion and migration in human breast cancer cells.
Breast Cancer Res Treat.
2000; 63(2): 147–52. PubMed Abstract
| Publisher Full Text
41.
Meng Q, Qi M, Chen DZ, et al.:
Suppression of breast cancer invasion and migration by indole-3-carbinol: associated with up-regulation of BRCA1 and E-cadherin/catenin complexes.
J Mol Med (Berl).
2000; 78(3): 155–65. PubMed Abstract
| Publisher Full Text
42.
Firestone GL, Bjeldanes LF:
Indole-3-carbinol and 3-3'-diindolylmethane antiproliferative signaling pathways control cell-cycle gene transcription in human breast cancer cells by regulating promoter-Sp1 transcription factor interactions.
J Nutr.
2003; 133(7 Suppl): 2448S–55. PubMed Abstract
| Publisher Full Text
44.
Fan S, Meng Q, Auborn K, et al.:
BRCA1 and BRCA2 as molecular targets for phytochemicals indole-3-carbinol and genistein in breast and prostate cancer cells.
Br J Cancer.
2006; 94(3): 407–26. PubMed Abstract
| Publisher Full Text
| Free Full Text
45.
Cohen T, Frydman-Marom A, Rechter M, et al.:
Inhibition of amyloid fibril formation and cytotoxicity by hydroxyindole derivatives.
Biochemistry.
2006; 45(15): 4727–35. PubMed Abstract
| Publisher Full Text
46.
Chang X, Tou JC, Hong C, et al.:
3,3'-Diindolylmethane inhibits angiogenesis and the growth of transplantable human breast carcinoma in athymic mice.
Carcinogenesis.
2005; 26(4): 771–8. PubMed Abstract
| Publisher Full Text
48.
Higdon JV, Delage B, Williams DE, et al.:
Cruciferous vegetables and human cancer risk: epidemiologic evidence and mechanistic basis.
Pharmacol Res.
2007; 55(3): 224–36. PubMed Abstract
| Publisher Full Text
| Free Full Text
49.
Dashwood RH:
Indole-3-carbinol: anticarcinogen or tumor promoter in brassica vegetables?
Chem Biol Interact.
1998; 110(1–2): 1–5. PubMed Abstract
| Publisher Full Text
50.
Lee BM, Park K:
Beneficial and adverse effects of chemopreventive agents.
Mutat Res.
2003; 523–524: 265–78. PubMed Abstract
| Publisher Full Text
53.
Kim JH, Lee BW, Schroeder FC, et al.:
Identification of indole glucosinolate breakdown products with antifeedant effects on Myzus persicae (green peach aphid).
Plant J.
2008; 54(6): 1015–26. PubMed Abstract
| Publisher Full Text
54.
Katz E, Nisani S, Yadav BS, et al.:
The glucosinolate breakdown product indole-3-carbinol acts as an auxin antagonist in roots of Arabidopsis thaliana.
Plant J.
2015; 82(4): 547–55. PubMed Abstract
| Publisher Full Text
55.
Katz E, Nisani S, Sela M, et al.:
The effect of indole-3-carbinol on PIN1 and PIN2 in Arabidopsis roots.
Plant Signal Behav.
2015; 10(9): e1062200. PubMed Abstract
| Publisher Full Text
| Free Full Text
56.
Zažímalová E, Petrášek J, Benková E:
Auxin and Its Role in Plant Development. Vienna: Springer Vienna; 2014. Publisher Full Text
58.
Katz E, Chamovitz DA:
Wounding of Arabidopsis leaves induces indole-3-carbinol-dependent autophagy in roots of Arabidopsis thaliana.
Plant J.
2017; 91(5): 779–87. PubMed Abstract
| Publisher Full Text
59.
Svenning S, Lamark T, Krause K, et al.:
Plant NBR1 is a selective autophagy substrate and a functional hybrid of the mammalian autophagic adapters NBR1 and p62/SQSTM1.
Autophagy.
2011; 7(9): 993–1010. PubMed Abstract
| Publisher Full Text
| Free Full Text
63.
Nguyen HH, Aronchik I, Brar GA, et al.:
The dietary phytochemical indole-3-carbinol is a natural elastase enzymatic inhibitor that disrupts cyclin E protein processing.
Proc Natl Acad Sci U S A.
2008; 105(50): 19750–5. PubMed Abstract
| Publisher Full Text
| Free Full Text
64.
Aronchik I, Bjeldanes LF, Firestone GL:
Direct inhibition of elastase activity by indole-3-carbinol triggers a CD40-TRAF regulatory cascade that disrupts NF-kappaB transcriptional activity in human breast cancer cells.
Cancer Res.
2010; 70(12): 4961–71. PubMed Abstract
| Publisher Full Text
65.
Cardozo T, Pagano M:
The SCF ubiquitin ligase: insights into a molecular machine.
Nat Rev Mol Cell Biol.
2004; 5(9): 739–51. PubMed Abstract
| Publisher Full Text
66.
Ruegger M, Dewey E, Gray WM, et al.:
The TIR1 protein of Arabidopsis functions in auxin response and is related to human SKP2 and yeast grr1p.
Genes Dev.
1998; 12(2): 198–207. PubMed Abstract
| Publisher Full Text
| Free Full Text
67.
Aronchik I, Kundu A, Quirit JG, et al.:
The antiproliferative response of indole-3-carbinol in human melanoma cells is triggered by an interaction with NEDD4-1 and disruption of wild-type PTEN degradation.
Mol Cancer Res.
2014; 12(11): 1621–34. PubMed Abstract
| Publisher Full Text
| Free Full Text
| Faculty Opinions Recommendation
68.
Quirit JG, Lavrenov SN, Poindexter K, et al.:
Indole-3-carbinol (I3C) analogues are potent small molecule inhibitors of NEDD4-1 ubiquitin ligase activity that disrupt proliferation of human melanoma cells.
Biochem Pharmacol.
2017; 127: 13–27. PubMed Abstract
| Publisher Full Text
| Faculty Opinions Recommendation
72.
Vert G, Nemhauser JL, Geldner N, et al.:
Molecular mechanisms of steroid hormone signaling in plants.
Annu Rev Cell Dev Biol.
2005; 21: 177–201. PubMed Abstract
| Publisher Full Text
73.
Vriet C, Russinova E, Reuzeau C:
From squalene to brassinolide: the steroid metabolic and signaling pathways across the plant kingdom.
Mol Plant.
2013; 6(6): 1738–57. PubMed Abstract
| Publisher Full Text
74.
Janeczko A:
The presence and activity of progesterone in the plant kingdom.
Steroids.
2012; 77(3): 169–73. PubMed Abstract
| Publisher Full Text
75.
Yang XH, Xu ZH, Xue HW:
Arabidopsis membrane steroid binding protein 1 is involved in inhibition of cell elongation.
Plant Cell.
2005; 17(1): 116–31. PubMed Abstract
| Publisher Full Text
| Free Full Text
1
School of Plant Sciences and Food Security, Tel Aviv University, Tel Aviv, Israel 2
Department of Plant Sciences, University of California, Davis, USA
Ella Katz
Roles:
Conceptualization,
Writing – Original Draft Preparation,
Writing – Review & Editing
Our research into the role of I3C in Arabidopsis is funded by grants from the Binational Agricultural Research and Development Fund (BARD, IS‐4505‐12R and US‐4846‐15C).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Katz E, Nisani S and Chamovitz DA. Indole-3-carbinol: a plant hormone combatting cancer [version 1; peer review: 2 approved]. F1000Research 2018, 7(F1000 Faculty Rev):689 (https://doi.org/10.12688/f1000research.14127.1)
NOTE: If applicable, it is important to ensure the information in square brackets after the title is included in all citations of this article.
track
receive updates on this article
Track an article to receive email alerts on any updates to this article.
Share
Open Peer Review
Current Reviewer Status:
?
Key to Reviewer Statuses
VIEWHIDE
ApprovedThe paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approvedFundamental flaws in the paper seriously undermine the findings and conclusions
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
NOTE: it is important to ensure the information in square brackets after the title is included in this citation.
Reviewer Report01 Jun 2018
Gary L. Firestone,
Department of Molecular and Cell Biology and the Cancer Research Laboratory, University of California at Berkeley, Berkely, California, USA
I confirm that I have read this submission and believe that I have an
... Continue reading
Competing Interests: No competing interests were disclosed.
Faculty Reviews are commissioned and written by members of the prestigious Faculty Opinions Faculty, and are edited as a service to our readers. In order to make these reviews as comprehensive and accessible as possible, we seek the reviewers’ input before publication. The reviewers’ names and any additional comments they may have are published alongside the review, as is usual on F1000Research.
I confirm that I have read this submission and believe that I have an appropriate level of expertise to confirm that it is of an acceptable scientific standard.
Alongside their report, reviewers assign a status to the article:
Approved - the paper is scientifically sound in its current form and only minor, if any, improvements are suggested
Approved with reservations -
A number of small changes, sometimes more significant revisions are required to address specific details and improve the papers academic merit.
Not approved - fundamental flaws in the paper seriously undermine the findings and conclusions
Adjust parameters to alter display
View on desktop for interactive features
Includes Interactive Elements
View on desktop for interactive features
Competing Interests Policy
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Examples of 'Non-Financial Competing Interests'
Within the past 4 years, you have held joint grants, published or collaborated with any of the authors of the selected paper.
You have a close personal relationship (e.g. parent, spouse, sibling, or domestic partner) with any of the authors.
You are a close professional associate of any of the authors (e.g. scientific mentor, recent student).
You work at the same institute as any of the authors.
You hope/expect to benefit (e.g. favour or employment) as a result of your submission.
You are an Editor for the journal in which the article is published.
Examples of 'Financial Competing Interests'
You expect to receive, or in the past 4 years have received, any of the following from any commercial organisation that may gain financially from your submission: a salary, fees, funding, reimbursements.
You expect to receive, or in the past 4 years have received, shared grant support or other funding with any of the authors.
You hold, or are currently applying for, any patents or significant stocks/shares relating to the subject matter of the paper you are commenting on.
Stay Updated
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Comments on this article Comments (0)