Keywords
nicastrin, gamma secretase, polyclonal, human, western blot
This article is included in the Antibody Validations gateway.
nicastrin, gamma secretase, polyclonal, human, western blot
The major change in this revision is the addition of a blot showing the antibody works with murine-derived protein as well. Otherwise we have revised the manuscript to alter some language and include important references suggested by the reviewers. There is also new underlying data for this figure that has been uploaded to FigShare.
See the authors' detailed response to the review by Satoru Funamoto
See the authors' detailed response to the review by Lucia Chavez-Gutierrez
The γ-secretase complex is a multi-subunit, intramembrane protease (reviewed1). It cleaves type-I single-pass transmembrane proteins within their transmembrane domain. This can lead to the release of an intracellular and an extracellular domain that may perform other functions. Examples include the cleavage of amyloid precursor protein (APP) to produce amyloid beta and the cleavage of activated NOTCH receptors to release their intracellular domain for translocation to the nucleus2.
Gamma-secretase is composed of several proteins, including a presenilin protease (PSEN1 or PSEN2), the presenilin enhancer gamma-secretase subunit (PEN2), an anterior pharynx-defective 1 protein (APH1A or APH1B), and nicastrin (NCSTN)3. Nicastrin acquires extensive N-linked glycosylation during its maturation4,5, though the glycosylation may not be required for typical cleavage activity6. The three-dimensional structure of human gamma-secretase shows that the heavily glycosylated ectodomain of nicastrin forms a horseshoe-like clamp on the extracellular portion of the complex7,8. It is thought that NCSTN may help control substrate selectivity9. Understanding the role of nicastrin in gamma-secretase has been challenging. Gamma-secretase can cleave many substrates without nicastrin, though nicastrin does help to exclude some substrates via steric hindrance10–12. There are multiple commercial antibodies for NCSTN available, but they do not agree on the expected product size. We validated one commercial polyclonal antibody (#N1660; Sigma-Aldrich) using HEK293 wildtype and nicastrin knockout cells.
We used a commercially available rabbit anti-human IgG polyclonal antibody that targets human nicastrin (#N1660; Sigma-Aldrich, St. Louis, MO, USA; RRID:AB_477259) which is has performed well in some previous publications13,14. The antibody was raised against Uniprot nicastrin peptide Q92542 (709 amino acid total size). The polyclonal was generated by challenging rabbits with a synthetic peptide corresponding to the C-terminal cytoplasmic domain of nicastrin (peptides 693-709) fused with keyhole limpet hemocyanin as an adjuvant.
The technical documentation claims this subsequence is identical to the matching region of nicastrin in mouse. However, aligning Q92542 to the primary mouse nicastrin peptide sequence (NP_067620.3) with Clustal Omega15,16 actually shows 1 mismatch (94.1% identity; Figure 1). It’s unclear if this discrepancy is due to changes to either the human or mouse peptide sequence for the most common isoform over time as the references have been updated.

Shown is a partial alignment between human and mouse nicastrin. The highlighted area represents the peptides use for generation of the polyclonal antibody. Asterisks represent a matching amino acid between the two sequences, and spaces are mismatches.
We used a mouse anti-human beta actin monoclonal antibody (#AB6276; Abcam, Cambridge, MA, USA; RRID: AB_2223210) as a loading control. The details of all primary and secondary antibodies are summarized in Table 1.
We purchased the Human Embryonic Kidney cell line (HEK293) from the ATCC (CRL-1573). We cultured all cells at 37°C and 5% CO2. For culture media, we used Dulbecco’s Modified Eagle’s Media (DMEM; Gibco, Thermo-Fisher Scientific, #11965-084) supplemented with 5% Fetal Bovine Serum (FBS; Gibco, Thermo-Fisher Scientific, #26140-079), 1% HEPES (Corning, #25-060-CI), 100 U/mL penicillin / streptomycin (Gibco, Thermo-Fisher Scientific, #15140-122), and 2 mM glutamine (Corning, #25-005-CI).
We used a HEK293 NCSTN knockout line we had previously generated using CRISPR/Cas9 genome-editing17. Briefly, we synthesized our single-guide RNA as an IDT gBlock and cloned it into the pCR-Blunt TOPO vector. We co-transfected the single-guide RNA vector along with humanized Cas9 (RRID: Addgene_43861) into HEK293 cells, plated to single colonies, and screened for deleted clones by sequencing (Sequence Read Archive project PRJNA268374) and RT-qPCR (Data available from figshare, see source data18). Full methodology for RT-qPCR is provided in the supplementary material of Cao et al.17
Reagent details can be found in Table 2 and Table 3. We harvested cells at ≥90% confluence and pelleted them by centrifugation at 4°C and 400 ×g for 5 minutes. We washed the cell pellet three times in 10 mL of cold phosphate buffered saline (PBS). We then added 300 µL of cold lysis buffer (50 mM Tris-HCl pH 8.0, 2 mM EDTA, 150 mM NaCl, 1.0% NP-40, and 1.5% protease inhibitor cocktail) and lysed the cells with constant agitation for 30 minutes at 4°C. We removed insoluble debris by centrifugation for 15 minutes at 4°C and 10,400 ×g. We determined the concentration of the cleared lysates using a Pierce BCA assay kit (#23227). We stored the lysates in aliquots at -80°C until further use.
We received a snap-frozen mouse liver (2 month-old C57BL/6 mouse) from the Alfred Kim lab, which had been obtained according to their approved IACUC protocol. We minced the liver into pieces and homogenized in ice-cold lysis buffer (5 mM Tris-HCl pH 8.0, 250 mM sucrose, 5 mM EDTA, 1.5% protease inhibitor cocktail) using a Wheaton tissue grinder. We then passed the solution through a QIAshredder spin-column (Qiagen #79656) to facilitate more complete lysis. We spun cellular debris out of solution by a 5 minute spin at 5,000 xg. We then precipitated membrane-enriched fragments by spinning for 5 minutes at 11,000 xg (4°C), then spinning the supernatant for an additional 1 hour at 4°C and 11,000 xg. We then extracted proteins from the membrane pellet by resuspending in buffer containing 2% (v/v) Triton X-100 and incubating on ice for 30 minutes. Any remaining unlysed material was pelleted with a 1 hour, 4°C, 11,000 xg spin. We determined the protein concentration of the lysate using the Pierce BCA assay kit. We stored lysates in aliquots at -80°C until blotting.
We used peptide-N-glycosidase F (PNGase F; #P0704S; New England Biolabs, Ipswich, MA, USA) to remove N-linked sugars. We denatured about 50 µg of protein in glycoprotein denaturing buffer (included with NEB kit; 0.5% SDS, 40 mM DTT) at 100°C for 10 minutes, and then incubated the lysate with PNGase F for 3 hours at 37°C, according to the manufacturer’s instructions. We treated a control in parallel under the same conditions, but omitted the PNGase F enzyme.
We denatured the protein lysate by boiling for 5 minutes in Laemmli sample buffer (5% β-mercaptoethanol). We resolved the proteins on precast 7.5% polyacrylamide gels (Mini-protean TGX; Bio-Rad, Hercules, CA, USA) after loading approximately 20 µg of lysate. We used the Precision Plus Dual-Color Standard as a molecular weight marker (Bio-Rad, Hercules, CA). We prepared PVDF membranes (0.45 µm) by incubating 2 minutes in 100% isopropanol, washing in Milli-Q water for 2 minutes, and equilibrating in transfer buffer for 10 minutes. We transferred separated proteins to the PVDF membrane in transfer buffer without methanol at 200 mA for 2 hours. We blocked the membrane by incubating in blocking buffer (TBST with 5% skim milk powder) for 1 hour at room temperature with gentle rocking. We probed the membrane using primary antibodies to nicastrin (1/1000) and beta actin (1/5000) diluted in blocking buffer overnight at 4°C with gentle rocking. We removed excess unbound antibody by rinsing the membranes 5 times for 10 minutes each in TBST buffer. The anti-mouse and anti-rabbit secondary antibodies were both conjugated to horseradish peroxidase (HRP). We incubated the membranes with secondary antibody (1/7000) in blocking buffer for 1.5 hours at room temperature, followed by washing 5 times for 10 minutes each in TBST. We used the Supersignal West Pico Chemiluminescent Substrate reagent (ThermoFisher, Waltham, MA) to detect secondary antibodies.
We collected protein lysates from wildtype HEK293 cells and HEK293 NCSTN knockouts. The manufacturer provided example blots were derived from HEK293 cells, but used an overexpression construct. In wildtype HEK293 cell lysates, a single, strong band at ~110 kDa can be seen on the blot, and this band is missing in the nicastrin knockout line lysates (Figure 2A, underlying data19,20). The loading controls for the wildtype replicates and knockout replicates all show the expected band for actin (Figure 2B, underlying data19,20), supporting that the loss of the nicastrin band is specific to the knockout and not a loading error. It is worth noting that despite a low background, the nicastrin blots showed an approximately 25 kDa band in both wildtype and knockout lysates. We searched the protein sequence used to develop the antibody (KADVLFIAPREPGAVSY) with protein blast using the Homo sapiens non-redundant peptide database automatically adjusted for short queries, but only matches to nicastrin had a reasonable e-value (2×10-9 to 7×10-11). It is therefore unclear if this band is from a non-specific contaminant in the antibody, a similar peptide that is poorly annotated in the non-redundant protein database, or a nicastrin degradation product.

A. The NCSTN antibody binds to endogenous levels of protein in wildtype (WT) HEK293 cells with a band at ~110 kDa. The band is absent in NCSTN knockout (KO) cells. Both replicates show an unidentified band at 25 kDa. B. The actin antibody shows the expected ~42 kDa band in both replicates of wildtype and knockout cells. Abbreviations: rep., replicate.
The nicastrin antibody documentation lists the expected fragment size as approximately 110 kDa, and this band size was confirmed on our blots. However, calculating the fragment size of human nicastrin protein sequence Q92542 using Expasy tools21 gives an estimated 78.4 kDa size for the nascent fragment and a reduced 75.2 kDa size after cleavage of the signal peptide. We hypothesized this discrepancy might be due to glycosylation.
We tested this hypothesis by first treating the lysates PNGase F, which will release asparagine-linked oligosaccharides. This reduced the molecular weight of the nicastrin band to less than 75 kDa (Figure 3A, underlying data22,23) without affecting the actin band (Figure 3B underlying data22,23). This phenomenon of a smaller than expected nicastrin band has been observed previously6,24. It is possible that a longer signal sequence than expected is cleaved from the nascent peptide. Given that detailed information is available for the signal cleavage of nicastrin9, a more likely explanation might be that the charge profile of the polypeptide affects its migration.
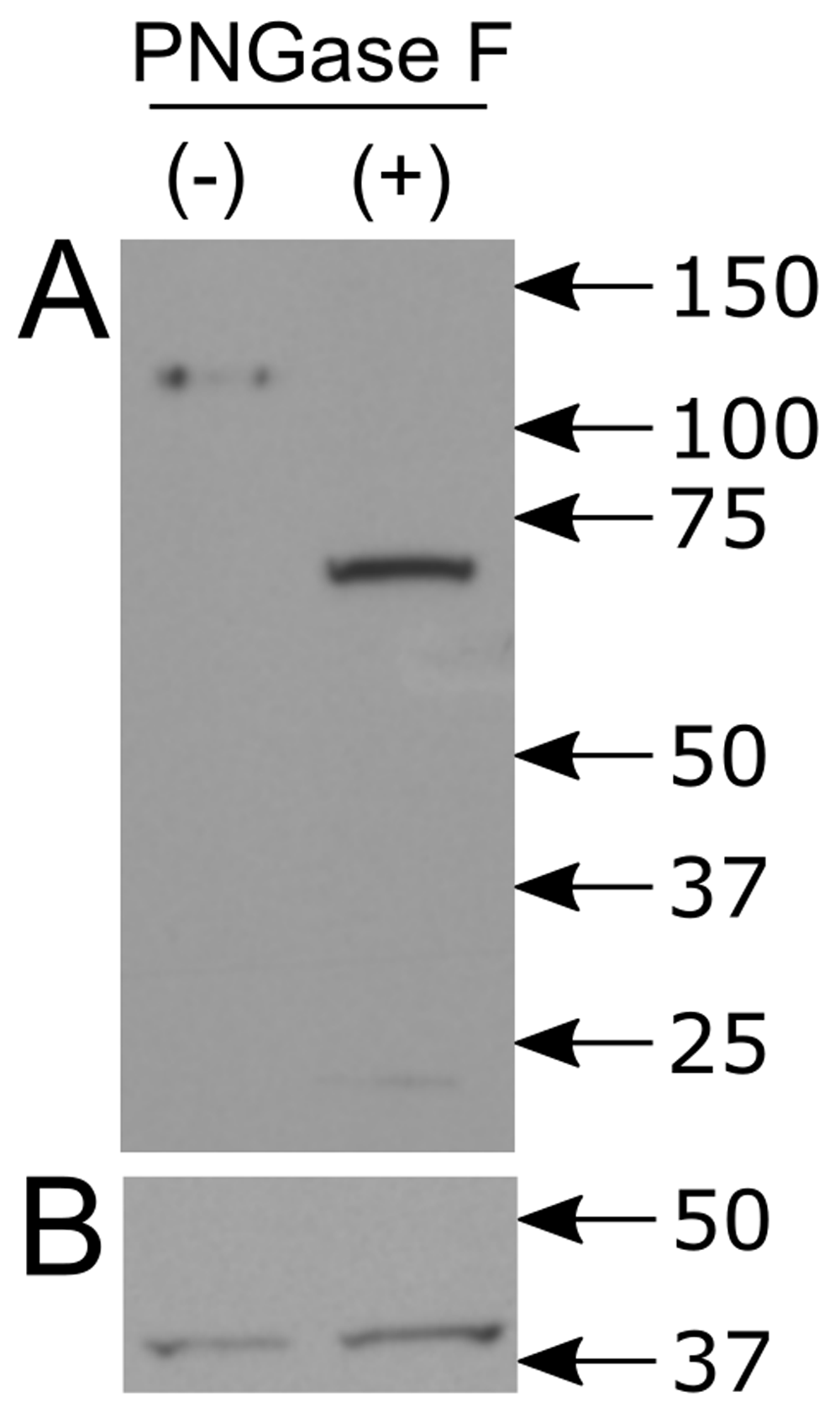
A. In lysates untreated with PNGase F (-), the expected ~110 kDa band is present. With PNGase F treatment (+), the band regresses to less than 75 kDa. B. In both PNGase treated and untreated lysates, the beta actin band is unchanged.
As noted above, there were mismatches between the sequence used to generate the antibody and the mouse sequence for nicastrin. It was possible that this mismatch was enough to reduce the effectiveness of this antibody in mouse extracts. We extracted protein from frozen mouse liver to test this possibility. We were able to confirm the presence of a band of the expected size in the mouse extracts (Figure 4, underlying data25). The same small, non-specific band was present in these blots as well.

Blot showing the results for 35 µg (1) or 25 µg (2) of mouse membrane protein lysate. The expected ~110 kDa band for mature nicastrin is present, as is the non-specific band present in most blots at < 25 kDa. These data suggest the antibody works as well for murine nicastrin as it does for human nicastrin.
We tested by immunoblot an anti-nicastrin antibody using HEK293 cell lysates and mouse liver extracts. Our results show that the antibody is sensitive enough to detect endogenous protein with reasonable specificity. It is able to bind to both glycosylated nicastrin and nicastrin without sugar linkages. The antibody functions for both endogenous human and mouse protein. It is unclear how well the antibody would work for cell staining due to the non-specific 25 kDa band we observed on nicastrin blots. Based on these data obtained with the protocols described above, we can confirm the utility of this nicastrin antibody for immunoblotting.
Home sapiens HEK293 NCSTN knockout by Cas9, Accession number: PRJNA268374
Figshare: HEK293 nicastrin knockout RT-qPCR. https://doi.org/10.6084/m9.figshare.7578539.v118
This project contains the following source data:
knockout_rtqpcr.csv (Raw Ct values of RT-qPCR confirming the knockout (CRISPR-Cas9 mediated) of nicastrin in HEK293 cells.)
Data are available under the terms of the Creative Commons Attribution 4.0 International license (CC-BY 4.0).
Figshare: NCSTN antibody validation - actin antibody in HEK293 knockout line. https://doi.org/10.6084/m9.figshare.8952968.v119
This project contains the following underlying data:
AntibodyValidation_NCSTN_KO_actin_Ab.svg (TIF image of actin antibody blot stored in a scaleable vector graphic file)
Figshare: NCSTN antibody validation - NCSTN antibody in HEK293 knockout line. https://doi.org/10.6084/m9.figshare.8952953.v120
This project contains the following underlying data:
AntibodyValidation_NCSTN_KO_NCSTN_Ab.svg (TIF image of NCSTN antibody blot stored in a scaleable vector graphic file)
Figshare: NCSTN antibody validation - actin antibody in HEK293 knockout line after PNGase treatment. https://doi.org/10.6084/m9.figshare.8952983.v122
This project contains the following underlying data:
AntibodyValidation_NCSTN_PNGase_actin_Ab.svg (TIF image of actin antibody blot stored in a scaleable vector graphic file)
Figshare: NCSTN antibody validation - NCSTN antibody in HEK293 knockout line after PNGase treatment. https://doi.org/10.6084/m9.figshare.8952977.v123
This project contains the following underlying data:
AntibodyValidation_NCSTN_PNGase_NCSTN_Ab.svg (TIF image of NCSTN antibody blot stored in a scaleable vector graphic file)
Figshare: NCSTN antibody validation - NCSTN antibody with murine liver protein. http://www.doi.org/10.6084/m9.figshare.11541864.v125
This project contains the following underlying data:
SuppFig5_MouseNCSTN.svg (TIF image of NCSTN antibody blot stored in a scaleable vector graphic file)
Data are available under the terms of the Creative Commons Zero "No rights reserved" data waiver (CC0 1.0 Public domain dedication).
| Views | Downloads | |
|---|---|---|
| F1000Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Already registered? Sign in
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
Eli
Eli
Eli